The Determination of HIV-1 RT Mutation Rate, Its Possible Allosteric Effects, and Its Implications on Drug Resistance


Abstract
1. Introduction
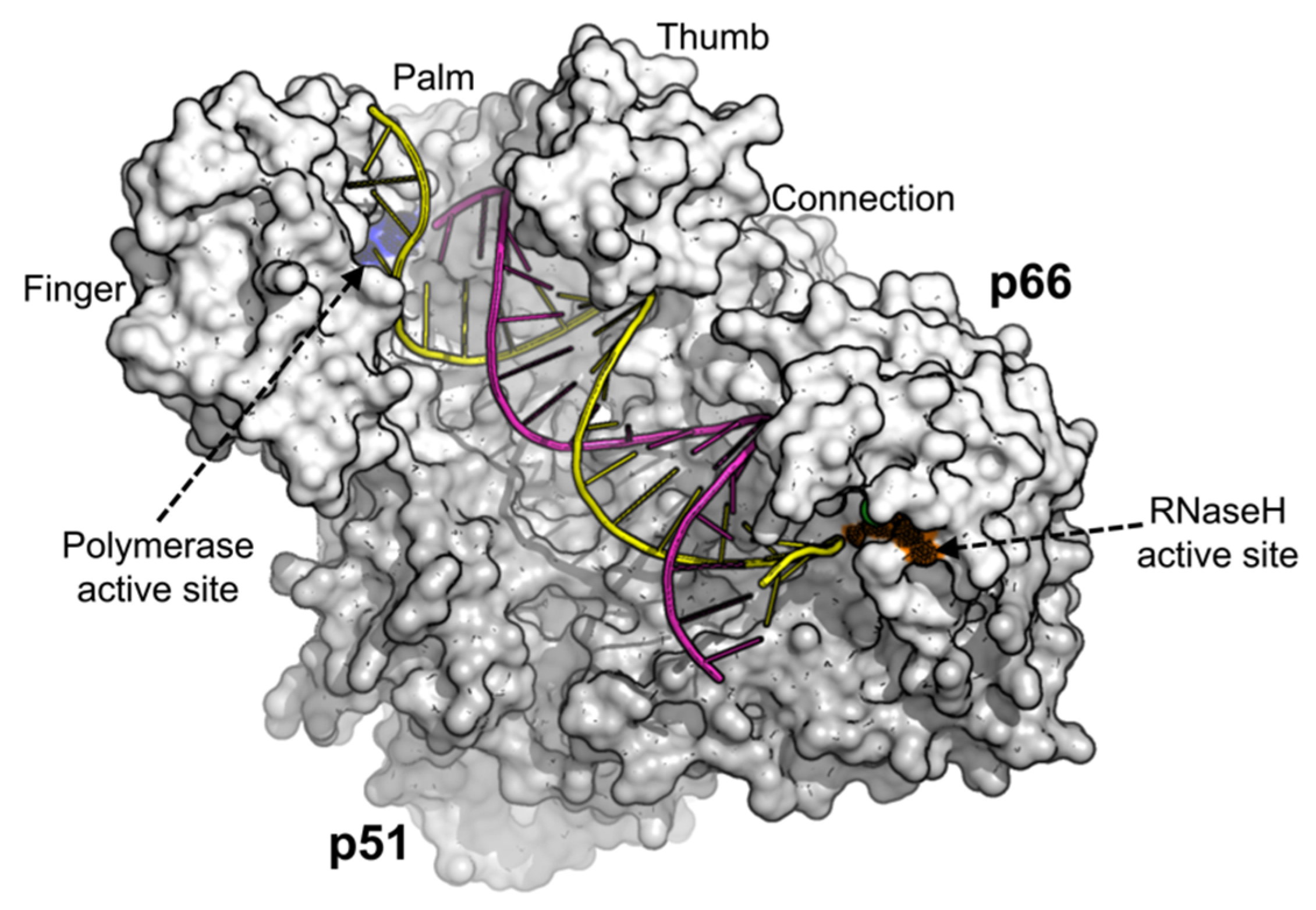
2. HIV-1 Reverse Transcriptase (RT)
3. Comparison of Cell-Free and Cell-Based Fidelity Assays
3.1. Cell-Free Fidelity Assays
3.2. Cell-Based Fidelity Assays
4. Studies on HIV-1 Genes
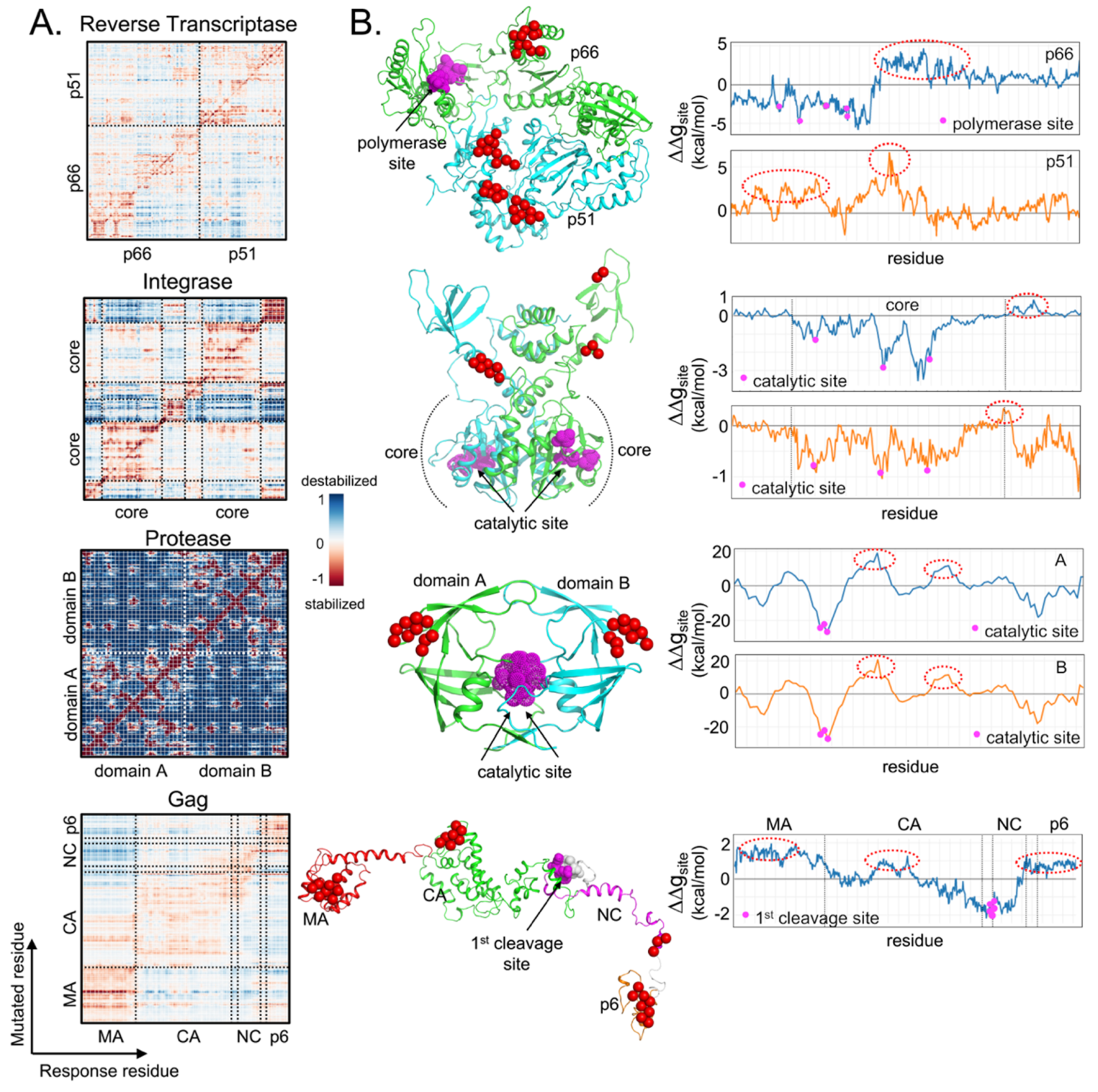
5. Analysis of Allosteric Communication
6. Implications on Sagacious Drug Design
7. Conclusions and Future Perspective
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Alizon, M.; Wain-Hobson, S.; Montagnier, L.; Sonigo, P. Genetic variability of the AIDS virus: Nucleotide sequence analysis of two isolates from African patients. Cell 1986, 46, 63–74. [Google Scholar] [CrossRef]
- Hahn, B.H.; Shaw, G.M.; Taylor, M.E.; Redfield, R.R.; Markham, P.D.; Salahuddin, S.Z.; Wong-Staal, F.; Gallo, R.C.; Parks, E.S.; Parks, W.P. Genetic variation in HTLV-III/LAV over time in patients with AIDS or at risk for AIDS. Science 1986, 232, 1548–1553. [Google Scholar] [CrossRef] [PubMed]
- Saag, M.S.; Hahn, B.H.; Gibbons, J.; Li, Y.; Parks, E.S.; Parks, W.P.; Shaw, G.M. Extensive variation of human immunodeficiency virus type-1 in vivo. Nature 1988, 334, 440–444. [Google Scholar] [CrossRef] [PubMed]
- Balfe, P.; Simmonds, P.; Ludlam, C.A.; Bishop, J.O.; Brown, A.J. Concurrent evolution of human immunodeficiency virus type 1 in patients infected from the same source: Rate of sequence change and low frequency of inactivating mutations. J. Virol. 1990, 64, 6221–6233. [Google Scholar] [CrossRef] [PubMed]
- Mansky, L.M. Retrovirus mutation rates and their role in genetic variation. J. Gen. Virol. 1998, 79, 1337–1345. [Google Scholar] [CrossRef] [PubMed]
- Menéndez-Arias, L. Mutation Rates and Intrinsic Fidelity of Retroviral Reverse Transcriptases. Viruses 2009, 1, 1137–1165. [Google Scholar] [CrossRef]
- Wei, X.; Ghosh, S.K.; Taylor, M.E.; Johnson, V.A.; Emini, E.A.; Deutsch, P.; Lifson, J.D.; Bonhoeffer, S.; Nowak, M.A.; Hahn, B.H. Viral dynamics in human immunodeficiency virus type 1 infection. Nature 1995, 373, 117–122. [Google Scholar] [CrossRef]
- Hu, W.S.; Temin, H.M. Retroviral recombination and reverse transcription. Science 1990, 250, 1227–1233. [Google Scholar] [CrossRef]
- Coffin, J.M. HIV population dynamics in vivo: Implications for genetic variation, pathogenesis, and therapy. Science 1995, 267, 483–489. [Google Scholar] [CrossRef]
- Fisher, A.G.; Ensoli, B.; Looney, D.; Rose, A.; Gallo, R.C.; Saag, M.S.; Shaw, G.M.; Hahn, B.H.; Wong-Staal, F. Biologically diverse molecular variants within a single HIV-1 isolate. Nature 1988, 334, 444–447. [Google Scholar] [CrossRef]
- Meyerhans, A.; Cheynier, R.; Albert, J.; Seth, M.; Kwok, S.; Sninsky, J.; Morfeldt-Månson, L.; Asjö, B.; Wain-Hobson, S. Temporal fluctuations in HIV quasispecies in vivo are not reflected by sequential HIV isolations. Cell 1989, 58, 901–910. [Google Scholar] [CrossRef]
- Hedskog, C.; Mild, M.; Jernberg, J.; Sherwood, E.; Bratt, G.; Leitner, T.; Lundeberg, J.; Andersson, B.; Albert, J. Dynamics of HIV-1 Quasispecies during Antiviral Treatment Dissected Using Ultra-Deep Pyrosequencing. PLoS ONE 2010, 5, e11345. [Google Scholar] [CrossRef] [PubMed]
- Ode, H.; Matsuda, M.; Matsuoka, K.; Hachiya, A.; Hattori, J.; Kito, Y.; Yokomaku, Y.; Iwatani, Y.; Sugiura, W. Quasispecies Analyses of the HIV-1 Near-full-length Genome With Illumina MiSeq. Front. Microbiol. 2015, 6, 1258. [Google Scholar] [CrossRef] [PubMed]
- Dampier, W.; Nonnemacher, M.R.; Mell, J.; Earl, J.; Ehrlich, G.D.; Pirrone, V.; Aiamkitsumrit, B.; Zhong, W.; Kercher, K.; Passic, S.; et al. HIV-1 Genetic Variation Resulting in the Development of New Quasispecies Continues to Be Encountered in the Peripheral Blood of Well-Suppressed Patients. PLoS ONE 2016, 11, e0155382. [Google Scholar] [CrossRef] [PubMed]
- Domingo, E.; Holland, J.J. Rna Virus Mutations and Fitness for Survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef] [PubMed]
- Gaschen, B.; Taylor, J.; Yusim, K.; Foley, B.; Gao, F.; Lang, D.; Novitsky, V.; Haynes, B.; Hahn, B.H.; Bhattacharya, T.; et al. Diversity considerations in HIV-1 vaccine selection. Science 2002, 296, 2354–2360. [Google Scholar] [CrossRef]
- Kellam, P.; Larder, B.A. Retroviral recombination can lead to linkage of reverse transcriptase mutations that confer increased zidovudine resistance. J. Virol. 1995, 69, 669–674. [Google Scholar] [CrossRef]
- Gu, Z.; Gao, Q.; Faust, E.A.; Wainberg, M.A. Possible involvement of cell fusion and viral recombination in generation of human immunodeficiency virus variants that display dual resistance to AZT and 3TC. J. Gen. Virol. 1995, 76, 2601–2605. [Google Scholar] [CrossRef]
- Moutouh, L.; Corbeil, J.; Richman, D.D. Recombination leads to the rapid emergence of HIV-1 dually resistant mutants under selective drug pressure. Proc. Natl. Acad. Sci. USA 1996, 93, 6106–6111. [Google Scholar] [CrossRef]
- Burke, D.S. Recombination in HIV: An important viral evolutionary strategy. Emerg. Infect. Dis. 1997, 3, 253–259. [Google Scholar] [CrossRef]
- Santoro, M.M.; Perno, C.F. HIV-1 Genetic Variability and Clinical Implications. ISRN Microbiol. 2013, 2013, 481314. [Google Scholar] [CrossRef] [PubMed]
- Lightfoote, M.M.; Coligan, J.E.; Folks, T.M.; Fauci, A.S.; Martin, M.A.; Venkatesan, S. Structural characterization of reverse transcriptase and endonuclease polypeptides of the acquired immunodeficiency syndrome retrovirus. J. Virol. 1986, 60, 771–775. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.D.; Bebenek, K.; Kunkel, T.A. The accuracy of reverse transcriptase from HIV-1. Science 1988, 242, 1171–1173. [Google Scholar] [CrossRef]
- Kim, T.; Mudry, R.A.; Rexrode, C.A.; Pathak, V.K. Retroviral mutation rates and A-to-G hypermutations during different stages of retroviral replication. J. Virol. 1996, 70, 7594–7602. [Google Scholar] [CrossRef] [PubMed]
- O’Neil, P.K.; Sun, G.; Yu, H.; Ron, Y.; Dougherty, J.P.; Preston, B.D. Mutational Analysis of HIV-1 Long Terminal Repeats to Explore the Relative Contribution of Reverse Transcriptase and RNA Polymerase II to Viral Mutagenesis. J. Biol. Chem. 2002, 277, 38053–38061. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, J.M.; Geller, R.; Garijo, R.; López-Aldeguer, J.; Sanjuán, R. Extremely High Mutation Rate of HIV-1 In Vivo. PLoS Biol. 2015, 13, e1002251. [Google Scholar] [CrossRef]
- Tuske, S.; Sarafianos, S.G.; Clark, A.D.; Ding, J.; Naeger, L.K.; White, K.L.; Miller, M.D.; Gibbs, C.S.; Boyer, P.L.; Clark, P.; et al. Structures of HIV-1 RT–DNA complexes before and after incorporation of the anti-AIDS drug tenofovir. Nature Structural & Molecular Biology 2004, 11, 469–474. [Google Scholar]
- Schrodinger, L. The PyMOL Molecular Graphics System. 2019. Available online: http://www.pymol.org (accessed on 5 February 2020).
- Loeb, L.A.; Kunkel, T.A. Fidelity of DNA synthesis. Annu. Rev. Biochem. 1982, 51, 429–457. [Google Scholar] [CrossRef]
- Weymouth, L.A.; Loeb, L.A. Mutagenesis during in vitro DNA synthesis. Proc. Natl. Acad. Sci. USA 1978, 75, 1924–1928. [Google Scholar] [CrossRef]
- Bebenek, K.; Kunkel, T.A. Analyzing fidelity of DNA polymerases. In Methods in Enzymology; DNA Replication; Elsevier: Amsterdam, The Netherlands, 1995; Volume 262, pp. 217–232. [Google Scholar]
- Preston, B.D.; Poiesz, B.J.; Loeb, L.A. Fidelity of HIV-1 reverse transcriptase. Science 1988, 242, 1168–1171. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Nagumo, T.; Hoshino, H. Low fidelity of cell-free DNA synthesis by reverse transcriptase of human immunodeficiency virus. J. Virol. 1988, 62, 3900–3902. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.; Grosse, F. Fidelity of human immunodeficiency virus type I reverse transcriptase in copying natural DNA. Nucleic Acids Res. 1989, 17, 1379–1393. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Loeb, L.A. Fidelity of HIV-1 reverse transcriptase copying RNA in vitro. Biochemistry 1992, 31, 954–958. [Google Scholar] [CrossRef]
- Ji, J.; Loeb, L.A. Fidelity of HIV-1 Reverse Transcriptase Copying a Hypervariable Region of the HIV-1 env Gene. Virology 1994, 199, 323–330. [Google Scholar] [CrossRef]
- Stuke, A.W.; Ahmad-Omar, O.; Hoefer, K.; Hunsmann, G.; Jentsch, K.D. Mutations in the SIV env and the M13 lacZa gene generated in vitro by reverse transcriptases and DNA polymerases. Arch. Virol. 1997, 142, 1139–1154. [Google Scholar] [CrossRef]
- Rezende, L.F.; Curr, K.; Ueno, T.; Mitsuya, H.; Prasad, V.R. The Impact of Multidideoxynucleoside Resistance-Conferring Mutations in Human Immunodeficiency Virus Type 1 Reverse Transcriptase on Polymerase Fidelity and Error Specificity. J. Virol. 1998, 72, 2890–2895. [Google Scholar] [CrossRef]
- Drosopoulos, W.C.; Prasad, V.R. Increased Misincorporation Fidelity Observed for Nucleoside Analog Resistance Mutations M184V and E89G in Human Immunodeficiency Virus Type 1 Reverse Transcriptase Does Not Correlate with the Overall Error Rate Measured In Vitro. J. Virol. 1998, 72, 4224–4230. [Google Scholar] [CrossRef]
- Rezende, L.F.; Drosopoulos, W.C.; Prasad, V.R. The influence of 3TC resistance mutation M184I on the fidelity and error specificity of human immunodeficiency virus type 1 reverse transcriptase. Nucleic Acids Res. 1998, 26, 3066–3072. [Google Scholar] [CrossRef]
- Boyer, P.L.; Hughes, S.H. Effects of Amino Acid Substitutions at Position 115 on the Fidelity of Human Immunodeficiency Virus Type 1 Reverse Transcriptase. J. Virol. 2000, 74, 6494–6500. [Google Scholar] [CrossRef][Green Version]
- Shah, F.S.; Curr, K.A.; Hamburgh, M.E.; Parniak, M.A.; Mitsuya, H.; Arnez, J.G.; Prasad, V.R. Differential influence of nucleoside analog-resistance mutations K65R and L74V on the overall mutation rate and error specificity of human immunodeficiency virus type 1 reverse transcriptase. J. Biol. Chem. 2000, 35, 27037–27044. [Google Scholar] [CrossRef]
- Rezende, L.F.; Kew, Y.; Prasad, V.R. The effect of increased processivity on overall fidelity of human immunodeficiency virus type 1 reverse transcriptase. J. Biomed. Sci. 2001, 8, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Boyer, P.L.; Stenbak, C.R.; Hoberman, D.; Linial, M.L.; Hughes, S.H. In Vitro Fidelity of the prototype primate foamy virus (PFV) RT compared to HIV-1 RT. Virology 2007, 367, 253–264. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.; Matamoros, T.; Menéndez-Arias, L. Increased thermostability and fidelity of DNA synthesis of wild-type and mutant HIV-1 group O reverse transcriptases. J. Mol. Biol. 2009, 392, 872–884. [Google Scholar] [CrossRef][Green Version]
- Álvarez, M.; Barrioluengo, V.; Afonso-Lehmann, R.N.; Menéndez-Arias, L. Altered error specificity of RNase H-deficient HIV-1 reverse transcriptases during DNA-dependent DNA synthesis. Nucleic Acids Res. 2013, 41, 4601–4612. [Google Scholar] [CrossRef][Green Version]
- Álvarez, M.; Sebastián-Martín, A.; García-Marquina, G.; Menéndez-Arias, L. Fidelity of classwide-resistant HIV-2 reverse transcriptase and differential contribution of K65R to the accuracy of HIV-1 and HIV-2 reverse transcriptases. Sci. Rep. 2017, 7, 44834. [Google Scholar] [CrossRef]
- Sebastián-Martín, A.; Barrioluengo, V.; Menéndez-Arias, L. Transcriptional inaccuracy threshold attenuates differences in RNA-dependent DNA synthesis fidelity between retroviral reverse transcriptases. Sci. Rep. 2018, 8, 627. [Google Scholar] [CrossRef]
- Bebenek, K.; Abbotts, J.; Roberts, J.D.; Wilson, S.H.; Kunkel, T.A. Specificity and mechanism of error-prone replication by human immunodeficiency virus-1 reverse transcriptase. J. Biol. Chem. 1989, 264, 16948–16956. [Google Scholar]
- Hübner, A.; Kruhoffer, M.; Grosse, F.; Krauss, G. Fidelity of human immunodeficiency virus type I reverse transcriptase in copying natural RNA. J. Mol. Biol. 1992, 223, 595–600. [Google Scholar] [CrossRef]
- Boyer, J.C.; Bebenek, K.; Kunkel, T.A. Unequal human immunodeficiency virus type 1 reverse transcriptase error rates with RNA and DNA templates. Proc. Natl. Acad. Sci. USA 1992, 89, 6919–6923. [Google Scholar] [CrossRef]
- Bebenek, K.; Abbotts, J.; Wilson, S.H.; Kunkel, T.A. Error-prone polymerization by HIV-1 reverse transcriptase. Contribution of template-primer misalignment, miscoding, and termination probability to mutational hot spots. J. Biol. Chem. 1993, 268, 10324–10334. [Google Scholar] [PubMed]
- Beard, W.A.; Stahl, S.J.; Kim, H.R.; Bebenek, K.; Kumar, A.; Strub, M.P.; Becerra, S.P.; Kunkel, T.A.; Wilson, S.H. Structure/function studies of human immunodeficiency virus type 1 reverse transcriptase. Alanine scanning mutagenesis of an alpha-helix in the thumb subdomain. J. Biol. Chem. 1994, 269, 28091–28097. [Google Scholar] [PubMed]
- Bebenek, K.; Beard, W.A.; Casas-Finet, J.R.; Kim, H.-R.; Darden, T.A.; Wilson, S.H.; Kunkel, T.A. Reduced Frameshift Fidelity and Processivity of HIV-1 Reverse Transcriptase Mutants Containing Alanine Substitutions in Helix H of the Thumb Subdomain. J. Biol. Chem. 1995, 270, 19516–19523. [Google Scholar] [CrossRef] [PubMed]
- Beard, W.A.; Minnick, D.T.; Wade, C.L.; Prasad, R.; Won, R.L.; Kumar, A.; Kunkel, T.A.; Wilson, S.H. Role of the “Helix Clamp” in HIV-1 Reverse Transcriptase Catalytic Cycling as Revealed by Alanine-scanning Mutagenesis. J. Biol. Chem. 1996, 271, 12213–12220. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Hathaway, T.R.; Loeb, L.A. Fidelity of Mutant HIV-1 Reverse Transcriptases: Interaction with the Single-Stranded Template Influences the Accuracy of DNA Synthesis. Biochemistry 1998, 37, 5831–5839. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Ayran, J.C.; Sagar, S.G.; Adman, E.T.; Fuller, S.M.; Tran, N.H.; Horrigan, J. New Human Immunodeficiency Virus, Type 1 Reverse Transcriptase (HIV-1 RT) Mutants with Increased Fidelity of DNA Synthesis. Accuracy, template binding, and processivity. J. Biol. Chem. 1999, 274, 27666–27673. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lewis, D.A.; Bebenek, K.; Beard, W.A.; Wilson, S.H.; Kunkel, T.A. Uniquely Altered DNA Replication Fidelity Conferred by an Amino Acid Change in the Nucleotide Binding Pocket of Human Immunodeficiency Virus Type 1 Reverse Transcriptase. J. Biol. Chem. 1999, 274, 32924–32930. [Google Scholar] [CrossRef] [PubMed]
- Jonckheere, H.; Clercq, E.D.; Anné, J. Fidelity analysis of HIV-1 reverse transcriptase mutants with an altered amino-acid sequence at residues Leu74, Glu89, Tyr115, Tyr183 and Met184. Eur. J. Biochem. 2000, 267, 2658–2665. [Google Scholar] [CrossRef]
- Weiss, K.K.; Isaacs, S.J.; Tran, N.H.; Adman, E.T.; Kim, B. Molecular Architecture of the Mutagenic Active Site of Human Immunodeficiency Virus Type 1 Reverse Transcriptase: Roles of the β8-αE Loop in Fidelity, Processivity, and Substrate Interactions. Biochemistry 2000, 39, 10684–10694. [Google Scholar] [CrossRef]
- Fisher, T.S.; Prasad, V.R. Substitutions of Phe61 Located in the Vicinity of Template 5′-Overhang Influence Polymerase Fidelity and Nucleoside Analog Sensitivity of HIV-1 Reverse Transcriptase. J. Biol. Chem. 2002, 277, 22345–22352. [Google Scholar] [CrossRef]
- Weiss, K.K.; Chen, R.; Skasko, M.; Reynolds, H.M.; Lee, K.; Bambara, R.A.; Mansky, L.M.; Kim, B. A Role for dNTP Binding of Human Immunodeficiency Virus Type 1 Reverse Transcriptase in Viral Mutagenesis. Biochemistry 2004, 43, 4490–4500. [Google Scholar] [CrossRef]
- Hamburgh, M.E.; Curr, K.A.; Monaghan, M.; Rao, V.R.; Tripathi, S.; Preston, B.D.; Sarafianos, S.; Arnold, E.; Darden, T.; Prasad, V.R. Structural Determinants of Slippage-mediated Mutations by Human Immunodeficiency Virus Type 1 Reverse Transcriptase. J. Biol. Chem. 2006, 281, 7421–7428. [Google Scholar] [CrossRef] [PubMed]
- Curr, K.; Tripathi, S.; Lennerstrand, J.; Larder, B.A.; Prasad, V.R. Influence of naturally occurring insertions in the fingers subdomain of human immunodeficiency virus type 1 reverse transcriptase on polymerase fidelity and mutation frequencies in vitro. J. Gen. Virol. 2006, 87, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Matamoros, T.; Kim, B.; Menéndez-Arias, L. Mechanistic Insights into the Role of Val75 of HIV-1 Reverse Transcriptase in Misinsertion and Mispair Extension Fidelity of DNA Synthesis. J. Mol. Biol. 2008, 375, 1234–1248. [Google Scholar] [CrossRef] [PubMed]
- Barrioluengo, V.; Álvarez, M.; Barbieri, D.; Menéndez-Arias, L. Thermostable HIV-1 group O reverse transcriptase variants with the same fidelity as murine leukaemia virus reverse transcriptase. Biochem. J. 2011, 436, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Mansky, L.M.; Temin, H.M. Lower in vivo mutation rate of human immunodeficiency virus type 1 than that predicted from the fidelity of purified reverse transcriptase. J. Virol. 1995, 69, 5087–5094. [Google Scholar] [CrossRef] [PubMed]
- Mansky, L.M. Forward mutation rate of human immunodeficiency virus type 1 in a T lymphoid cell line. AIDS Res. Hum. Retroviruses 1996, 12, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Mansky, L.M. The mutation rate of human immunodeficiency virus type 1 is influenced by the vpr gene. Virology 1996, 222, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.J.; Wooley, D.P. A new cell-based assay for measuring the forward mutation rate of HIV-1. J. Virol. Methods 2005, 124, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Rawson, J.M.O.; Landman, S.R.; Reilly, C.S.; Mansky, L.M. HIV-1 and HIV-2 exhibit similar mutation frequencies and spectra in the absence of G-to-A hypermutation. Retrovirology 2015, 12, 60. [Google Scholar] [CrossRef]
- Abram, M.E.; Ferris, A.L.; Shao, W.; Alvord, W.G.; Hughes, S.H. Nature, Position, and Frequency of Mutations Made in a Single Cycle of HIV-1 Replication. J. Virol. 2010, 84, 9864–9878. [Google Scholar] [CrossRef]
- Abram, M.E.; Ferris, A.L.; Das, K.; Quinoñes, O.; Shao, W.; Tuske, S.; Alvord, W.G.; Arnold, E.; Hughes, S.H. Mutations in HIV-1 Reverse Transcriptase Affect the Errors Made in a Single Cycle of Viral Replication. J. Virol. 2014, 88, 7589–7601. [Google Scholar] [CrossRef] [PubMed]
- Geller, R.; Domingo-Calap, P.; Cuevas, J.M.; Rossolillo, P.; Negroni, M.; Sanjuán, R. The external domains of the HIV-1 envelope are a mutational cold spot. Nat. Commun. 2015, 6, 8571. [Google Scholar] [CrossRef] [PubMed]
- Mansky, L.M.; Rouzic, E.L.; Benichou, S.; Gajary, L.C. Influence of Reverse Transcriptase Variants, Drugs, and Vpr on Human Immunodeficiency Virus Type 1 Mutant Frequencies. J. Virol. 2003, 77, 2071–2080. [Google Scholar] [CrossRef] [PubMed]
- Su, C.T.-T.; Koh, D.W.-S.; Gan, S.K.-E. Reviewing HIV-1 Gag Mutations in Protease Inhibitors Resistance: Insights for Possible Novel Gag Inhibitor Designs. Molecules 2019, 24, 3243. [Google Scholar] [CrossRef] [PubMed]
- Eckert, K.A.; Kunkel, T.A. Fidelity of DNA synthesis catalyzed by human DNA polymerase alpha and HIV-1 reverse transcriptase: Effect of reaction pH. Nucleic Acids Res. 1993, 21, 5212–5220. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, A.B.; Usman, M.; Kandi, V. Current Scenario of HIV/AIDS, Treatment Options, and Major Challenges with Compliance to Antiretroviral Therapy. Cureus 2016, 8, e515. [Google Scholar] [CrossRef]
- Wensing, A.M.; Calvez, V.; Ceccherini-Silberstein, F.; Charpentier, C.; Günthard, H.F.; Paredes, R.; Shafer, R.W.; Richman, D.D. 2019 Update of the Drug Resistance Mutations in HIV-1. Top. Antivir. Med. 2019, 27, 111–121. [Google Scholar]
- Seckler, J.M.; Barkley, M.D.; Wintrode, P.L. Allosteric suppression of HIV-1 reverse transcriptase structural dynamics upon inhibitor binding. Biophys. J. 2011, 100, 144–153. [Google Scholar] [CrossRef]
- Patel, D.; Antwi, J.; Koneru, P.C.; Serrao, E.; Forli, S.; Kessl, J.J.; Feng, L.; Deng, N.; Levy, R.M.; Fuchs, J.R.; et al. A New Class of Allosteric HIV-1 Integrase Inhibitors Identified by Crystallographic Fragment Screening of the Catalytic Core Domain. J. Biol. Chem. 2016, 291, 23569–23577. [Google Scholar] [CrossRef]
- Ung, P.M.-U.; Dunbar, J.B.; Gestwicki, J.E.; Carlson, H.A. An allosteric modulator of HIV-1 protease shows equipotent inhibition of wild-type and drug-resistant proteases. J. Med. Chem. 2014, 57, 6468–6478. [Google Scholar] [CrossRef]
- Chan, K.-F.; Phua, S.-X.; Su, C.T.-T.; Gan, S.K.-E. Inhibiting HIV-1 and MMLV Reverse Transcriptase: The potential of an Allosteric Broad-Spectrum Inhibitor. Manuscript in preparation.
- Su, C.T.-T.; Lua, W.-H.; Ling, W.-L.; Gan, S.K.-E. Allosteric Effects between the Antibody Constant and Variable Regions: A Study of IgA Fc Mutations on Antigen Binding. Antibodies 2018, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Chiang, R.Z.-H.; Gan, S.K.-E.; Su, C.T.-T. A computational study for rational HIV-1 non-nucleoside reverse transcriptase inhibitor selection and the discovery of novel allosteric pockets for inhibitor design. Biosci. Rep. 2018, 38, BSR20171113. [Google Scholar] [CrossRef]
- Freed, E.O. HIV-1 assembly, release and maturation. Nat. Rev. Microbiol. 2015, 13, 484–496. [Google Scholar] [CrossRef] [PubMed]
- Su, C.T.-T.; Kwoh, C.-K.; Verma, C.S.; Gan, S.K.-E. Modeling the full length HIV-1 Gag polyprotein reveals the role of its p6 subunit in viral maturation and the effect of non-cleavage site mutations in protease drug resistance. J. Biomol. Struct. Dyn. 2018, 36, 4366–4377. [Google Scholar] [CrossRef] [PubMed]
- Gatanaga, H.; Suzuki, Y.; Tsang, H.; Yoshimura, K.; Kavlick, M.F.; Nagashima, K.; Gorelick, R.J.; Mardy, S.; Tang, C.; Summers, M.F. Amino acid substitutions in Gag protein at non-cleavage sites are indispensable for the development of a high multitude of HIV-1 resistance against protease inhibitors. J. Biol. Chem. 2002, 277, 5952–5961. [Google Scholar] [CrossRef]
- Guarnera, E.; Berezovsky, I.N. Structure-Based Statistical Mechanical Model Accounts for the Causality and Energetics of Allosteric Communication. PLoS Comput. Biol. 2016, 12, e1004678. [Google Scholar] [CrossRef]
- Guarnera, E.; Tan, Z.W.; Zheng, Z.; Berezovsky, I.N. AlloSigMA: Allosteric signaling and mutation analysis server. Bioinformatics 2017, 33, 3996–3998. [Google Scholar] [CrossRef]
- Guarnera, E.; Berezovsky, I.N. On the perturbation nature of allostery: Sites, mutations, and signal modulation. Curr. Opin. Struct. Biol. 2019, 56, 18–27. [Google Scholar] [CrossRef]
- Guarnera, E.; Berezovsky, I.N. Toward Comprehensive Allosteric Control over Protein Activity. Structure 2019, 27, 866–878. [Google Scholar] [CrossRef]
- Tan, Z.W.; Tee, W.-V.; Guarnera, E.; Booth, L.; Berezovsky, I.N. AlloMAPS: Allosteric mutation analysis and polymorphism of signaling database. Nucleic Acids Res. 2018, 47, D265–D270. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Miller, J.T.; Lapkouski, M.; Tian, L.; Yang, W.; Le Grice, S.F.J. Examining the Role of the HIV-1 Reverse Transcriptase p51 Subunit in Positioning and Hydrolysis of RNA/DNA Hybrids. J. Biol. Chem. 2013, 288, 16177–16184. [Google Scholar] [CrossRef] [PubMed]
- Appadurai, R.; Senapati, S. Dynamical Network of HIV-1 Protease Mutants Reveals the Mechanism of Drug Resistance and Unhindered Activity. Biochemistry 2016, 55, 1529–1540. [Google Scholar] [CrossRef] [PubMed]
- Su, C.T.-T.; Ling, W.-L.; Lua, W.-H.; Haw, Y.-X.; Gan, S.K.-E. Structural analyses of 2015-updated drug-resistant mutations in HIV-1 protease: An implication of protease inhibitor cross-resistance. BMC Bioinformatics 2016, 17, 500. [Google Scholar] [CrossRef] [PubMed]
- Maguire, M.F.; Guinea, R.; Griffin, P.; Macmanus, S.; Elston, R.C.; Wolfram, J.; Richards, N.; Hanlon, M.H.; Porter, D.J.T.; Wrin, T.; et al. Changes in Human Immunodeficiency Virus Type 1 Gag at Positions L449 and P453 Are Linked to I50V Protease Mutants In Vivo and Cause Reduction of Sensitivity to Amprenavir and Improved Viral Fitness In Vitro. J. Virol. 2002, 76, 7398. [Google Scholar] [CrossRef] [PubMed]
- Doyon, L.; Croteau, G.; Thibeault, D.; Poulin, F.; Pilote, L.; Lamarre, D. Second locus involved in human immunodeficiency virus type 1 resistance to protease inhibitors. J. Virol. 1996, 70, 3763. [Google Scholar] [CrossRef]
- Turner, D.; Brenner, B.; Wainberg, M.A. Multiple Effects of the M184V Resistance Mutation in the Reverse Transcriptase of Human Immunodeficiency Virus Type 1. Clin. Diagn. Lab. Immunol. 2003, 10, 979–981. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Assay | RT Mutant | Vector | Reporter Gene/Template | Error Rate (× 10−4 errors/bp/cycle) | Reference |
|---|---|---|---|---|---|
| Base reversion | WT 1 | ΦX174 am3 | Position 587 of ΦX174 am3, DNA | 2.50 | [32] |
| Base reversion | WT 1 | M13mp2 | Position 89 of lacZα coding sequence, DNA | 0.55 | [23] |
| Forward mutation | lacZα, DNA | 5.00–6.67 | |||
| Misincorporation | WT (HTLV-IIIB) | - | Polyadenylic acid | 0.31–0.57 | [33] |
| WT (HIV[GUN-1]) | - | 0.26–0.38 | |||
| Base reversion | WT (HTLV-III) | ΦX174 am16 | ΦX174 am16, DNA | 1.43–2.00 | [34] |
| Forward mutation | WT 1 | - | lacZα, RNA | 1.45 | [35] |
| - | M13mp2 (+), DNA | 1.69 | |||
| Forward mutation | WT 1 | - | env V-1, DNA | 1.90 | [36] |
| - | env V-1, RNA | 2.00 | |||
| - | env V-1, RNA/DNA | 3.80 | |||
| Forward mutation | WT (BH10) | M13mp19 (CSIVM13B) | SIVagmTYO-7 env (minus-strand), DNA | 0.18 | [37] |
| M13mp19 (CSIVM13B) | lacZα, DNA | 0.53 | |||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 0.45 | [38] |
| Q151M (BH10) | 0.40 | ||||
| A62V/V75I/F77L/F116Y/Q151M (BH10) | 0.23 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 0.57 | [39] |
| E89G (HXB2) | 0.41 | ||||
| M184V (HXB2) | 0.36 | ||||
| E89G/M184V (HXB2) | 0.81 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 0.71 | [39,40] |
| M184V (HXB2) | 0.43 | ||||
| Forward mutation | M184I (HXB2) | M13mp2 | lacZα, DNA | 0.17 | [40] |
| Forward mutation | WT 1 | U-DNA Litmus 29 (Not) | lacZα, DNA | 1.60 | [41] |
| Y115F 1 | 1.00 | ||||
| Y115V 1 | 4.70 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 0.57 | [42] |
| K65R (HXB2) | 0.07 | ||||
| L74V (HXB2) | 0.30 | ||||
| Forward mutation | WT (NL4-3) | M13mp2 | lacZα, DNA | 0.63 | [43] |
| FE20 (NL4-3) | 0.56 | ||||
| FE103 (NL4-3) | 0.53 | ||||
| Forward mutation | WT 1 | U-DNA Litmus 29 (Not) | lacZα, DNA | 0.75 | [44] |
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 1.36 | [45] |
| WT (ESP49) | 0.55 | ||||
| V75I (ESP49) | 0.29 | ||||
| Forward mutation | D433N (ESP49) | M13mp2 | lacZα, DNA | 0.14 | [46] |
| E478Q (ESP49) | 0.1 | ||||
| V75I/D443N (ESP49) | 0.14 | ||||
| V75I/E478Q (ESP49) | 0.2 | ||||
| E478Q (BH10) | 0.42 | ||||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 1.52 | [47] |
| WT (ROD) | 1.00 | ||||
| K65R (ROD) | 0.84 | ||||
| K65R/Q151M/M184V (ROD) | 0.74 | ||||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, RNA | 0.35 | [48] |
| WT (ESP49) | 0.27 | ||||
| K65R (ESP49) | 0.26 | ||||
| K65R/V75I (ESP49) | 0.25 |
| Assay | RT Mutant | Vector | Reporter Gene/Template | Mutant Frequency (× 10−4 Mutants/Clones) | Reference |
|---|---|---|---|---|---|
| Base reversion | WT 1 | M13mp2 | Position 89 of lacZα coding sequence, DNA | 1.00 | [23] |
| Forward mutation | WT 1 | M13mp2 | lacZα, DNA | 340–460 | |
| Forward mutation | WT 1 | M13mp2 | lacZα, DNA | 390.00 | [49] |
| Base reversion | WT 1 | pTZ18R | ΦX174 am16, RNA | 26.00 | [50] |
| ΦX174 am16, DNA | 26.00 | ||||
| Forward mutation | WT 1 | M13mp2 | lacZα, DNA | 340–540 | [51] |
| lacZα, RNA | 91–210 | ||||
| Forward mutation | WT 1 | M13mp2 | lacZα, RNA | 40.70 | [35] |
| pBluescript SK+ | M13mp2, (+) DNA | 47.30 | |||
| Forward mutation | WT 1 | M13mp2 | pseudowild type 1 (pwt1) lacZα, DNA | 490 | [52] |
| pseudowild type 2 (pwt2) lacZα, DNA | 450 | ||||
| lacZα, DNA | 500 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 200 | [53] |
| D256A (HXB2) | 240 | ||||
| Q258A (HXB2) | 390 | ||||
| K259A (HXB2) | 300 | ||||
| L260A (HXB2) | 230 | ||||
| G262A (HXB2) | 880 | ||||
| K263A (HXB2) | 290 | ||||
| W266A (HXB2) | 640 | ||||
| Q269A (HXB2) | 510 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 210 | [54] |
| G262A (HXB2) | 860 | ||||
| W266A (HXB2) | 630 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 160 | [55] |
| R277A (HXB2) | 140 | ||||
| Q278A (HXB2) | 190 | ||||
| L279A (HXB2) | 150 | ||||
| C280A (HXB2) | 300 | ||||
| K281A (HXB2) | 140 | ||||
| L282A (HXB2) | 120 | ||||
| R284A (HXB2) | 170 | ||||
| G285A (HXB2) | 160 | ||||
| K287A (HXB2) | 120 | ||||
| Forward mutation | WT (BH10) | M13mp19 (CSIVM13B) | SIVagmTYO-7 env (minus-strand), DNA | 31.40 | [37] |
| lacZα, DNA | 60.90 | ||||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 232 | [56] |
| D76V (BH10) | 26 | ||||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 64.00 | [38] |
| Q151M (BH10) | 55.00 | ||||
| A62V/V75I/F77L/F116Y/Q151M (BH10) | 31.00 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 86.00 | [39] |
| E89G (HXB2) | 62.60 | ||||
| M184V (HXB2) | 55.30 | ||||
| E89G/M184V (HXB2) | 123.00 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 97 | [39,40] |
| M184V (HXB2) | 59 | ||||
| Forward mutation | M184I (HXB2) | M13mp2 | lacZα, DNA | 24 | [40] |
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 250 | [57] |
| R78A (BH10) | 28 | ||||
| Base reversion | WT (HXB2) | TGA codon (position 87–89) in lacZα, DNA | 2.2 | [58] | |
| R72A (HXB2) | 82 | ||||
| One-nucleotide deletion reversion | WT (HXB2) | TTTT run in M13mp2 DNA | 32 | ||
| R72A (HXB2) | 1.6 | ||||
| Forward mutation | WT (HXB2) | lacZα, DNA | 210 | ||
| R72A (HXB2) | 340 | ||||
| Forward mutation | WT 1 | U-DNA Litmus 29 (Not) | lacZα, DNA | 278.00 | [41] |
| Y115F 1 | 175.00 | ||||
| Y115V 1 | 82.00 | ||||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 192 | [59] |
| L74V (BH10) | 55 | ||||
| E89G (BH10) | 96 | ||||
| M184V (BH10) | 228 | ||||
| Y183F (BH10) | 303 | ||||
| Y115A (BH10) | 763 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 86 | [42] |
| K65R (HXB2) | 10.6 | ||||
| L74V (HXB2) | 50.5 | ||||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 261 | [60] |
| Q151N (BH10) | 20 | ||||
| K154A (BH10) | 125 | ||||
| Forward mutation | WT (NL4-3) | M13mp2 | lacZα, DNA | 86.00 | [43] |
| FE20 (NL4-3) | 77.00 | ||||
| FE103 (NL4-3) | 74.00 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 97 | [61] |
| F61A (HXB2) | 8.3 | ||||
| Forward mutation | V184I (HXB2) | M13mp2 | lacZα, DNA | 30 | [62] |
| Forward mutation | E89K (HXB2) | M13mp2 | lacZα, DNA | 77 | [63] |
| E89V (HXB2) | 64 | ||||
| E89S (HXB2) | 53 | ||||
| Forward mutation | WT (HXB2) | M13mp2 | lacZα, DNA | 97 | [64] |
| T69S-AG (HXB2) | 20 | ||||
| T69S-SG (HXB2) | 12 | ||||
| T69S-SS (HXB2) | 24 | ||||
| A62V/T69S-AG/L210W/R211K/L214F/T215Y | 8.5 | ||||
| A62V/T69S-SG/L210W/R211K/L214F/T215Y | 19 | ||||
| A62V/T69S-SS/L210W/R211K/L214F/T215Y | 11 | ||||
| M41L/T69S-AG/L210W/R211K/L214F/T215Y | 6.3 | ||||
| M41L/T69S-SG/L210W/R211K/L214F/T215Y | 5.9 | ||||
| Forward mutation | WT 1 | U-DNA Litmus 29 (Not) | lacZα, DNA | 130.00 | [44] |
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 206 | [65] |
| V75A (BH10) | 281 | ||||
| V75F (BH10) | 112 | ||||
| V75I (BH10) | 69.6 | ||||
| Base reversion | WT (BH10) | 27 | |||
| V75I (BH10) | 7.8 | ||||
| Forward mutation | WT (ESP49) | M13mp2 | lacZα, DNA | 83.1 | [45] |
| V75I (ESP49) | 43.4 | ||||
| Forward mutation | K65R (ESP49) | M13mp2 | lacZα, DNA | 7.7 | [66] |
| K65R/V75I (ESP49) | 8.9 | ||||
| R78A (ESP49) | 5.9 | ||||
| Forward mutation | WT (ESP49) | M13mp2 | lacZα, DNA | 96 | [46] |
| D433N (ESP49) | 19.8 | ||||
| E478Q (ESP49) | 13.5 | ||||
| V75I/D443N (ESP49) | 18.2 | ||||
| V75I/E478Q (ESP49) | 29.1 | ||||
| WT (BH10) | 113.4–132.3 | ||||
| E478Q (BH10) | 57.6 | ||||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 199.00 | [47] |
| WT (ROD) | 124.20 | ||||
| K65R (ROD) | 117.90 | ||||
| K65R/Q151M/M184V (ROD) | 103.10 | ||||
| Forward mutation | WT (BH10) | M13mp2 | lacZα, DNA | 40.50 | [48] |
| WT (ESP49) | 34.5 | ||||
| K65R (ESP49) | 29.5 | ||||
| K65R/V75I (ESP49) | 29.3 |
| Assay | RT Mutant | Vector | Reporter Gene/Template | Error Rate (× 10−4 errors/bp/cycle) | Reference |
|---|---|---|---|---|---|
| Forward mutation | WT (NL4-3) | HIV-1 vector (HIV shuttle 3.12 & 5.1) | lacZα, DNA | 0.34 | [67] |
| Forward mutation | WT (NL4-3) | HIV-1 vector (HIV shuttle 3.12) | lacZα, DNA | 0.40 | [68] |
| Forward mutation | WT (NL4-3) | HIV-1 vector (HIV shuttle 3.12 vpr+) | lacZα, DNA | 0.30 | [69] |
| HIV-1 vector (HIV shuttle 3.12 vpr ATG−) | 1.20 | ||||
| Forward mutation (SSCP) | WT (HXB2) | HIV-1 vector (pHIV-gpt) | HIV-1 LTR, DNA | 0.92 | [25] |
| WT (NL4-3) | HIV-1 vector (NL4-3gpt) | 0.79 | |||
| Forward mutation | WT (NL4-3) | HIV-1 vector (pNL4-3deltaΔ +cass) | tk, DNA | 0.22 | [70] |
| Forward mutation | WT (NL4-3) | HIV-1 vector (pNL4-3 HIG) | U373-MAGI-X4 cells, DNA | 6.90 | [71] |
| Forward mutation | WT (NL4-3) | HIV-1 vector (pSICO-LZF) | lacZα, DNA | 0.22 | [72] |
| HIV-1 vector (pSICO-LZR) | 0.17 | ||||
| Forward mutation | WT (NL4-3) | HIV-1 vector (pSICO-LZF/R) | lacZα, DNA | 0.14 | [73] |
| Y115F (NL4-3) | 0.37 | ||||
| Q151M (NL4-3) | 0.17 | ||||
| M184I (NL4-3) | 0.21 | ||||
| M184V (NL4-3) | 0.18 | ||||
| Forward mutation | WT (HX2B2) | HIV-1 vector (pSDY-dCK) | HIV env, RNA | 0.36 | [74] |
| HIV Int-vif-vpr, RNA | 0.75 |
| Assay | RT Mutant | Vector | Reporter Gene/Template | Mutant Frequency (× 10−4 Mutants/Clones) | Reference |
|---|---|---|---|---|---|
| Forward mutation | WT (NL4-3) | HIV-1 vector (HIV shuttle 3.12) | lacZα, DNA | 44 | [67] |
| HIV-1 vector (HIV shuttle 5.2) | 42 | ||||
| Forward mutation | WT (NL4-3) | HIV-1 vector (HIV shuttle 3.12) | lacZα, DNA | 50 | [68] |
| Forward mutation | WT (NL4-3) | HIV-1 vector (HIV shuttle 3.12 vpr ATG−) | lacZα, DNA | 150 | [69] |
| HIV-1 vector (HIV shuttle 3.12 vpr A30F) | 140 | ||||
| HIV-1 vector (HIV shuttle 3.12 vpr+) | 40 | ||||
| Forward mutation | WT (NL4-3) | HIV-1 vector | lacZα, DNA | 1500–1510 2 | [75] |
| K65R (NL4-3) | 450 2 | ||||
| D67N (NL4-3) | 1490 2 | ||||
| K70R (NL4-3) | 14702 | ||||
| L74V (NL4-3) | 11202 | ||||
| D76V (NL4-3) | 590–600 2 | ||||
| R78A (NL4-3) | 420–430 2 | ||||
| E89G (NL4-3) | 1202 | ||||
| Y115A (NL4-3) | 3400–3480 2 | ||||
| Q151N (NL4-3) | 250–280 2 | ||||
| K154A (NL4-3) | 1520 2 | ||||
| F227A (NL4-3) | 930 2 | ||||
| W229A (NL4-3) | 720 2 | ||||
| Y501W (NL4-3) | 4300 2 | ||||
| I505A (NL4-3) | 1410 2 | ||||
| D76V/R78A (NL4-3) | 150 2 | ||||
| R78A/Q151N (NL4-3) | 110 2 | ||||
| Y115A/Q151N (NL4-3) | 1050 2 | ||||
| Forward mutation | WT (NL4-3) | HIV-1 vector (HIV shuttle 3.12) | lacZα, DNA | 1490 | [62] |
| V148I (NL4-3) | 390 | ||||
| Q151N (NL4-3) | 260 | ||||
| Forward mutation | WT (NL4-3) | HIV-1 vector (pSICO-LZF/R) | lacZα (F), DNA | 38 | [72] |
| HIV-1 vector (pSICO-LZF/R) | lacZα (R), DNA | 21.8 | |||
| HIV-1 vector (pNLZeoIN-R-E-.LZF/R) | lacZα (F), DNA | 21.7 | |||
| HIV-1 vector (pNLZeoIN-R-E-.LZF/R) | lacZα (R), DNA | 18.2 | |||
| Forward mutation | WT (NL4-3) | HIV-1 vector (pSICO-LZF/R) | lacZα, DNA | 21.98 | [73] |
| Y115F (NL4-3) | 55.91 | ||||
| Q151M (NL4-3) | 25.69 | ||||
| M184I (NL4-3) | 31.9 | ||||
| M184V (NL4-3) | 27 |
| Template | Base Substitutions | Frameshifts | Others | Reference | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Transversions | Transitions | Insertions | Deletions | |||||||||||||
| A -> C | C -> A | A -> T | T -> A | C -> G | G -> C | G -> T | T -> G | G -> A | A -> G | C -> T | T -> C | |||||
| HIV-1 env V-1, DNA | 0 | 0 | 1.59 (1) | 1.59 (1) | 1.59 (1) | 0 | 4.76 (3) | 6.35 (4) | 9.52 (6) | 26.98 (17) | 15.87 (10) | 4.76 (3) | 23.81 (15) | 3.17 (2) | 0 | [36] |
| HIV-1 env V-1, DNA/RNA | 7.55 (4) | 0 | 3.77 (2) | 3.77 (2) | 0 | 0 | 16.98 (9) | 0 | 3.77 (2) | 20.75 (11) | 16.98 (9) | 13.21 (7) | 7.55 (4) | 5.66 (3) | 0 | |
| HIV-1 env | 27.88 (29) | 46.15 (48) | 15.38 (16) | 8.65 (9) | 1.92 (2) | 0 | [74] | |||||||||
| HIV-1 int-vif-vpr | 19.61 (20) | 50.98 (52) | 22.55 (23) | 4.90 (2) | 1.96 (2) | 0 | ||||||||||
| LacZα, RNA | - | - | - | - | - | 10.64 (5) | 23.40 (11) | - | 0 | - | 31.91 (15) | 2.13 (1) | 14.89 (7) | 17.02 (8) | [35] | |
| LacZα, DNA | 0 | 31.53 (70) | 0 | 30.63 (68) | 0.45 (1) | 0.45 (1) | 0 | 0.90 (2) | 12.61 (28) | 3.60 (8) | 4.95 (11) | 0 | 14.86 (33) | 0 | [41] | |
| LacZα, DNA | 0.57 (1) | 11.93 (21) | 0 | 9.66 (17) | 0 | 2.27 (4) | 0 | 0.57 (1) | 45.45 (80) | 6.25 (11) | 7.39 (13) | 1.14 (2) | 10.23 (18) | 4.55 (8) | [44] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeo, J.Y.; Goh, G.-R.; Su, C.T.-T.; Gan, S.K.-E. The Determination of HIV-1 RT Mutation Rate, Its Possible Allosteric Effects, and Its Implications on Drug Resistance. Viruses 2020, 12, 297. https://doi.org/10.3390/v12030297
Yeo JY, Goh G-R, Su CT-T, Gan SK-E. The Determination of HIV-1 RT Mutation Rate, Its Possible Allosteric Effects, and Its Implications on Drug Resistance. Viruses. 2020; 12(3):297. https://doi.org/10.3390/v12030297
Chicago/Turabian StyleYeo, Joshua Yi, Ghin-Ray Goh, Chinh Tran-To Su, and Samuel Ken-En Gan. 2020. "The Determination of HIV-1 RT Mutation Rate, Its Possible Allosteric Effects, and Its Implications on Drug Resistance" Viruses 12, no. 3: 297. https://doi.org/10.3390/v12030297
APA StyleYeo, J. Y., Goh, G.-R., Su, C. T.-T., & Gan, S. K.-E. (2020). The Determination of HIV-1 RT Mutation Rate, Its Possible Allosteric Effects, and Its Implications on Drug Resistance. Viruses, 12(3), 297. https://doi.org/10.3390/v12030297