Oxidative Stress in Canine Histiocytic Sarcoma Cells Induced by an Infection with Canine Distemper Virus Led to a Dysregulation of HIF-1α Downstream Pathway Resulting in a Reduced Expression of VEGF-B In Vitro

 ,
,  ,
,  , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Production of Cell Pellets
2.2. Microarray Data Analysis Using a Manually Generated List of Gene Symbols Related to ROS Production and Scavenging, ER Stress and HIF-1α Pathway
2.3. Immunofluorescence and Statistical Analysis
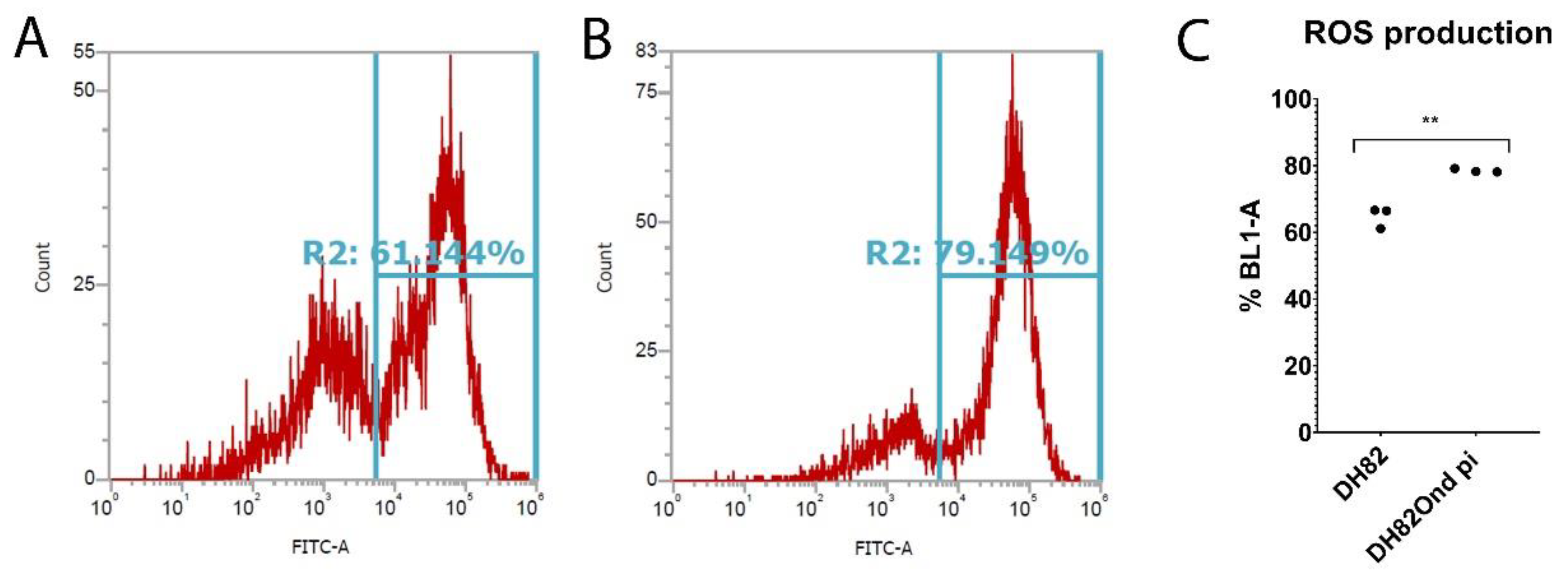
2.4. Determination of Oxidative Burst by Flow Cytometry
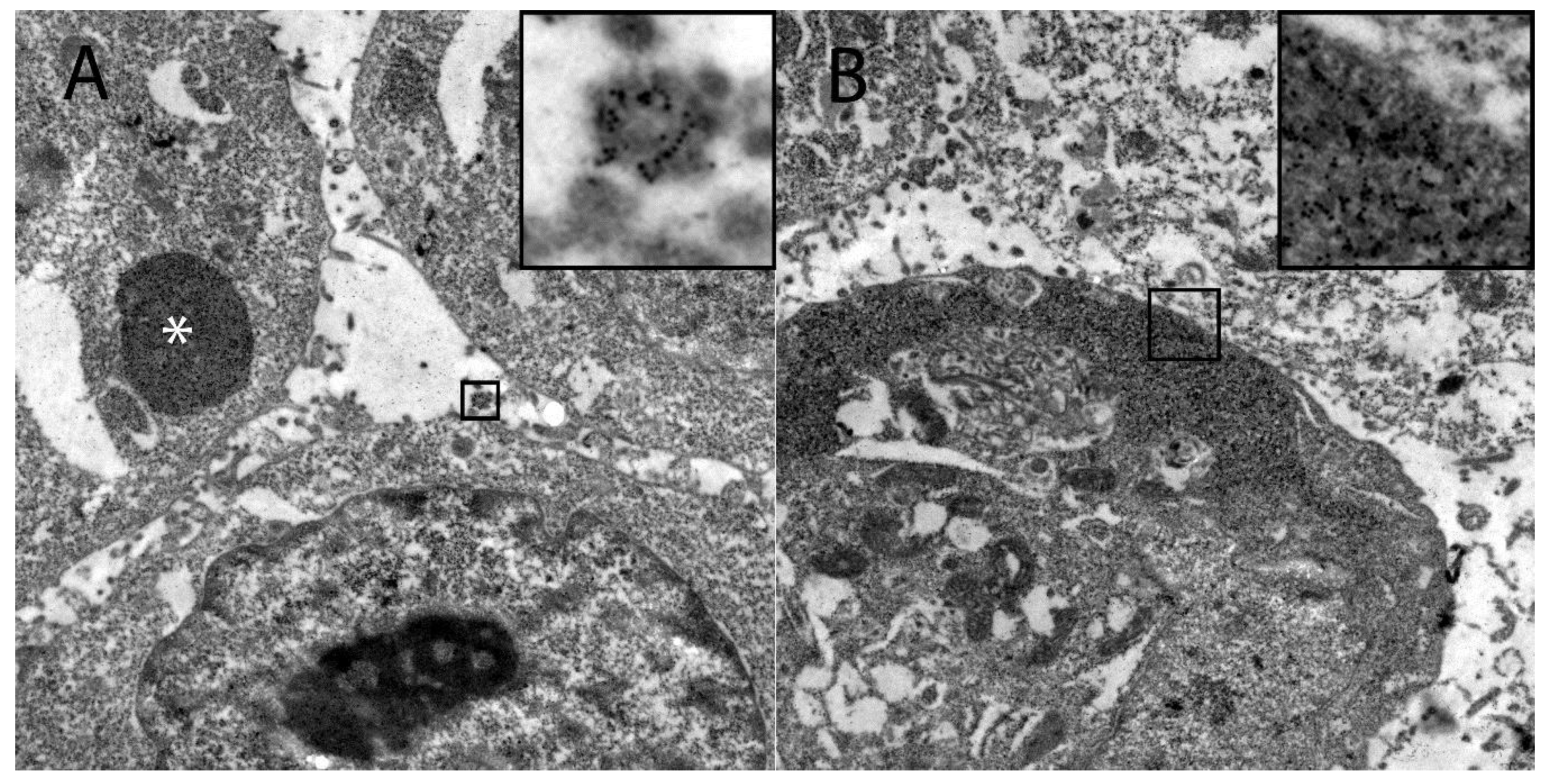
2.5. Immunoelectron Microscopy
2.6. Laser Scanning Confocal Microscopy
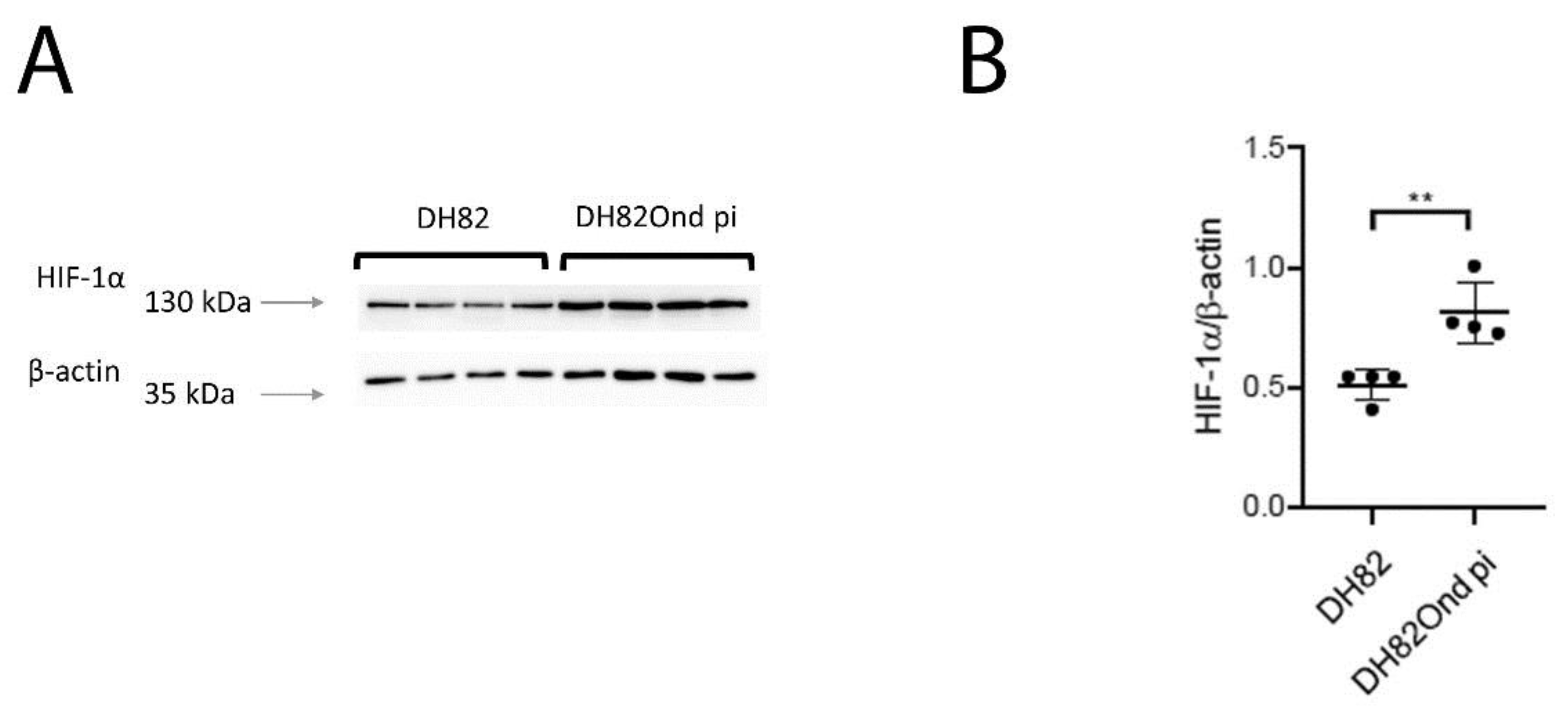
2.7. Immunoblotting
3. Results
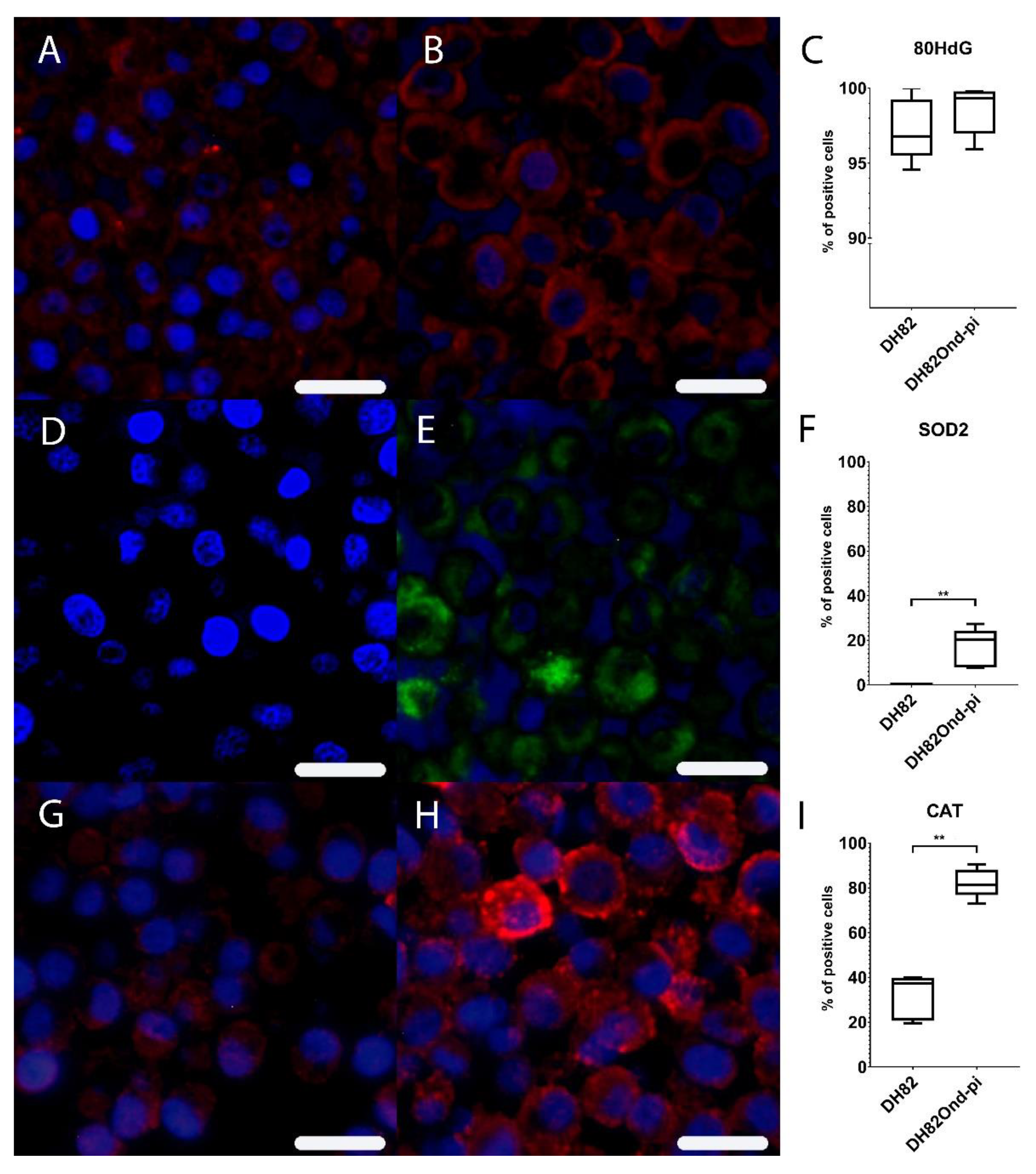
3.1. Persistent CDV Infection of DH82cells Leads to an Increased Level of Intracellular ROS Associated with Increased Catalase and Superoxide Dismutase 2 Protein Expression
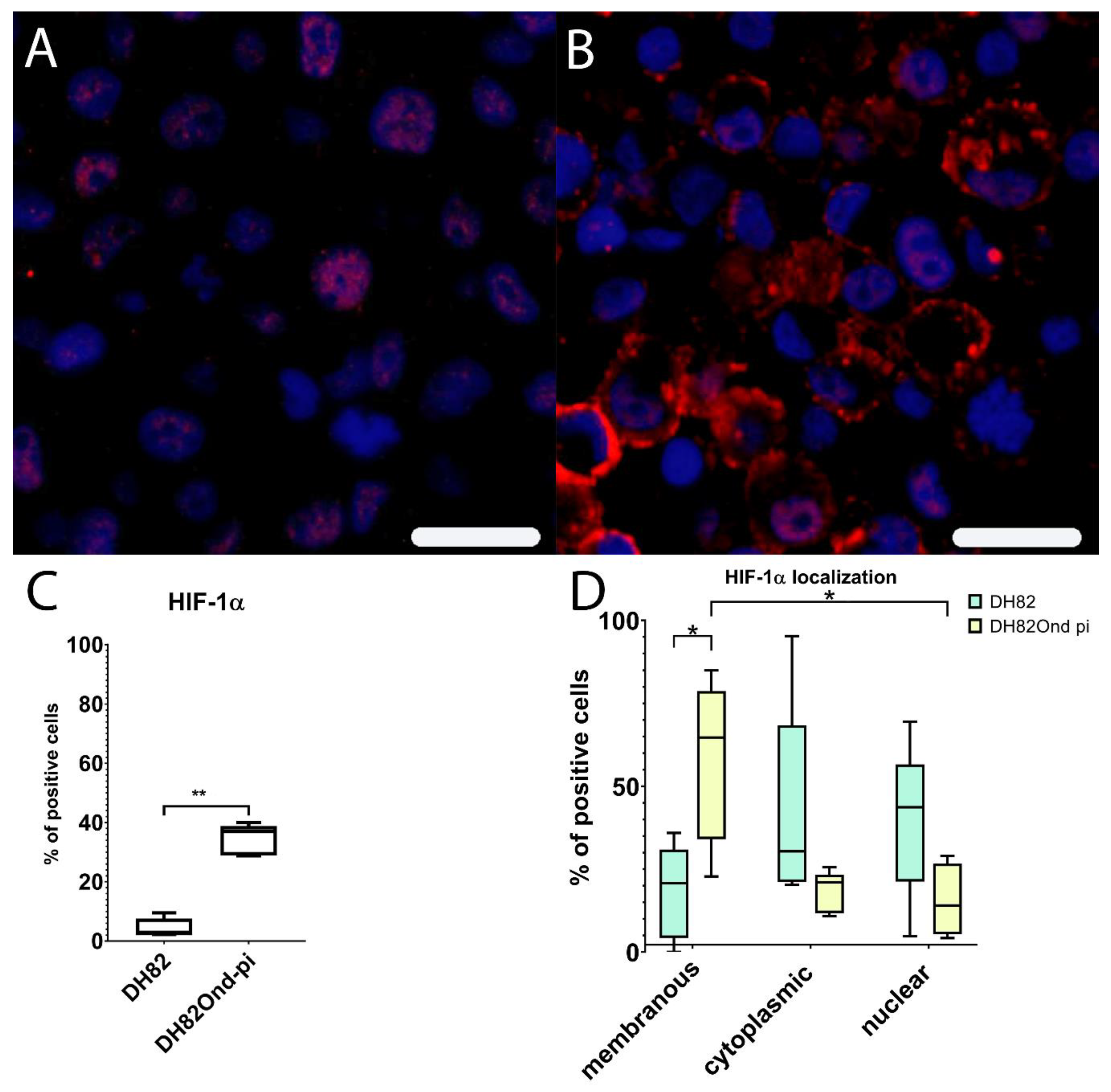
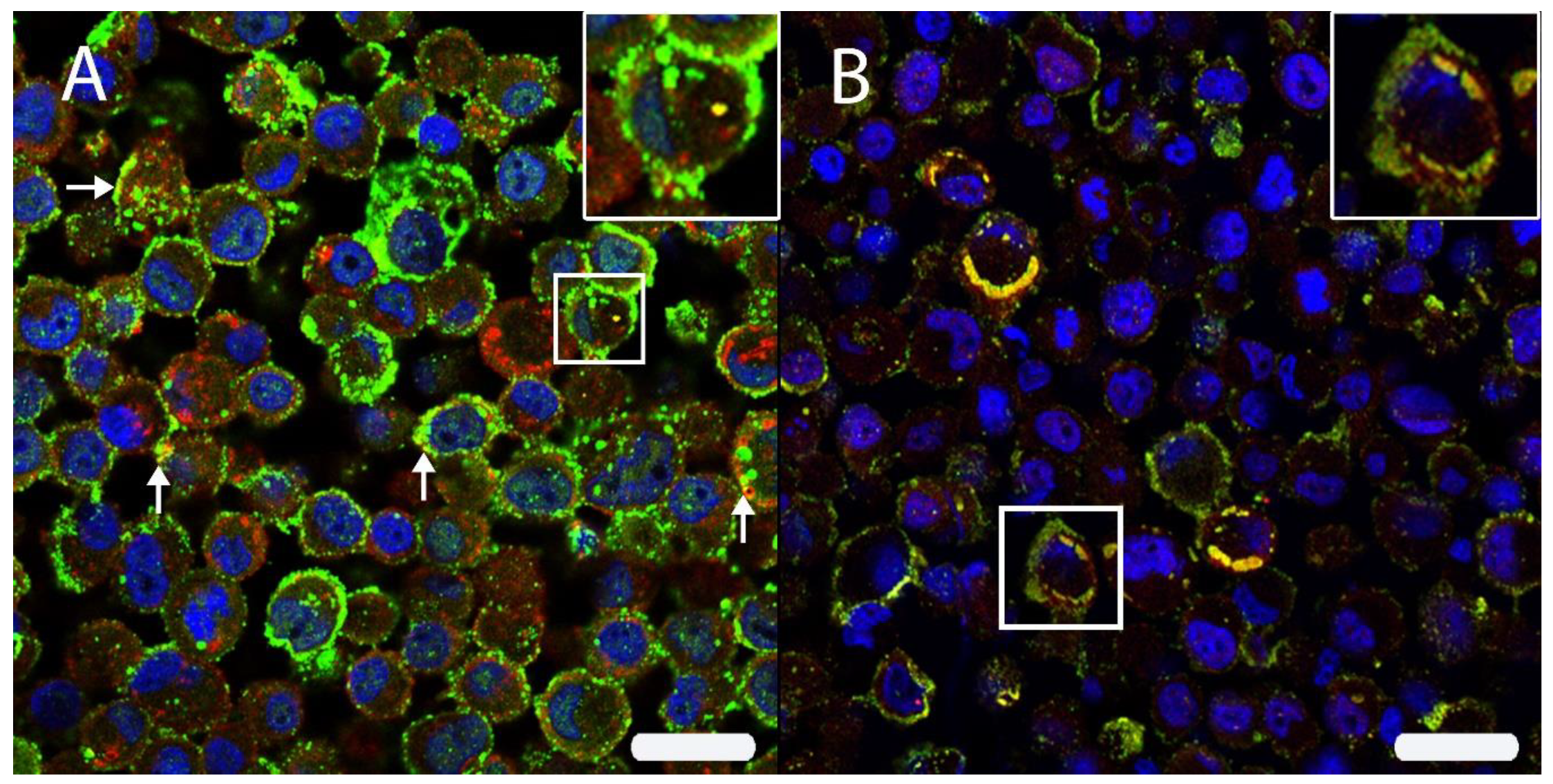
3.2. DH82Ond pi Are Characterized by an Increased HIF-1α Protein Expression Associated with an Altered Intracellular Distribution
3.3. DH82Ond pi Show an Unusual Mainly Sub-Membranous Distribution of HIF-1α
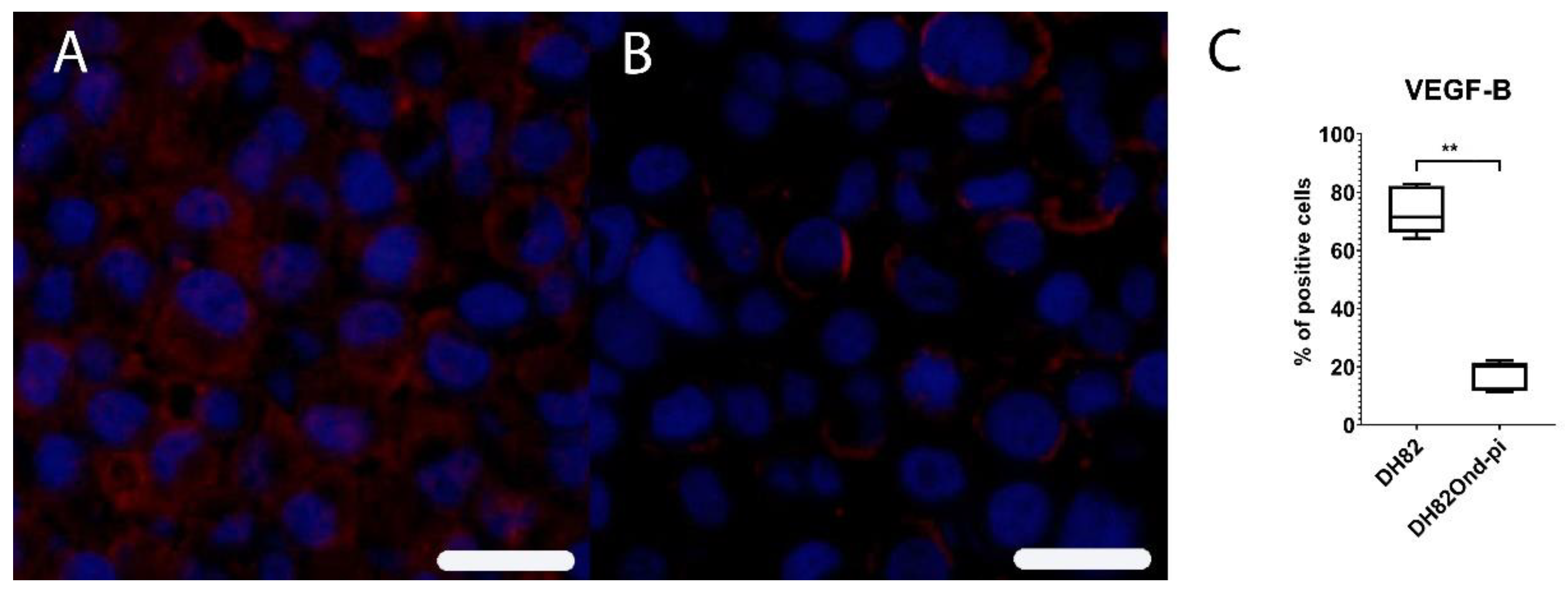
3.4. Unexpected Intracellular HIF-1α Localisation Is Associated with a Dysregulated Expression of Angiogenetic Downstream Targets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hedan, B.; Thomas, R.; Motsinger-Reif, A.; Abadie, J.; Andre, C.; Cullen, J.; Breen, M. Molecular cytogenetic characterization of canine histiocytic sarcoma: A spontaneous model for human histiocytic cancer identifies deletion of tumor suppressor genes and highlights influence of genetic background on tumor behavior. BMC Cancer 2011, 11, 201. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, J.D.; Breen, M. Comparative oncology: What dogs and other species can teach us about humans with cancer. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.C.; Galanis, E.; Kirn, D. Clinical trial results with oncolytic virotherapy: A century of promise, a decade of progress. Nat. Clin. Pract. Oncol. 2007, 4, 101–117. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.; McFadden, G. Viruses for tumor therapy. Cell Host Microbe 2014, 15, 260–265. [Google Scholar] [CrossRef]
- Sinkovics, J.G. Viral oncolysates as human tumor vaccines. Int. Rev. Immunol. 1991, 7, 259–287. [Google Scholar] [CrossRef]
- Heinzerling, L.; Künzi, V.; Oberholzer, P.A.; Kündig, T.; Naim, H.; Dummer, R. Oncolytic measles virus in cutaneous T-cell lymphomas mounts antitumor immune responses in vivo and targets interferon-resistant tumor cells. Blood 2005, 106, 2287–2294. [Google Scholar] [CrossRef]
- Parato, K.A.; Senger, D.; Forsyth, P.A.J.; Bell, J.C. Recent progress in the battle between oncolytic viruses and tumours. Nat. Rev. Cancer 2005, 5, 965–976. [Google Scholar] [CrossRef]
- Lapp, S.; Pfankuche, V.M.; Baumgärtner, W.; Puff, C. Viral oncolysis—Can insights from measles be transferred to canine distemper virus? Viruses 2014, 6, 2340–2375. [Google Scholar] [CrossRef]
- Suter, S.E.; Chein, M.B.; Von Messling, V.; Yip, B.; Cattaneo, R.; Vernau, W.; Madewell, B.R.; London, C.A. In vitro canine distemper virus infection of canine lymphoid cells: A prelude to oncolytic therapy for lymphoma. Clin. Cancer Res. 2005, 11, 1579–1587. [Google Scholar] [CrossRef]
- Dispenzieri, A.; Tong, C.; Laplant, B.; Lacy, M.Q.; Laumann, K.; Dingli, D.; Zhou, Y.; Federspiel, M.J.; Gertz, M.A.; Hayman, S.; et al. Phase I trial of systemic administration of edmonston strain of measles virus genetically engineered to express the sodium iodide symporter in patients with recurrent or refractory multiple myeloma. Leukemia 2017, 31, 2791–2798. [Google Scholar] [CrossRef]
- Pfankuche, V.M.; Spitzbarth, I.; Lapp, S.; Ulrich, R.; Deschl, U.; Kalkuhl, A.; Baumgärtner, W.; Puff, C. Reduced angiogenic gene expression in morbillivirus-triggered oncolysis in a translational model for histiocytic sarcoma. J. Cell. Mol. Med. 2017, 21, 816–830. [Google Scholar] [CrossRef] [PubMed]
- Gröne, A.; Fonfara, S.; Baumgärtner, W. Cell type-dependent cytokine expression after canine distemper virus infection. Viral Immunol. 2002, 15, 493–505. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA. Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Adams, V.J.; Evans, K.M.; Sampson, J.; Wood, J.L.N. Methods and mortality results of a health survey of purebred dogs in the UK. J. Small Anim. Pract. 2010, 51, 512–524. [Google Scholar] [CrossRef]
- Moore, P.F. A Review of Histiocytic Diseases of Dogs and Cats. Vet. Pathol. 2014, 51, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Schlick, K.; Aigelsreiter, A.; Pichler, M.; Reitter, S.; Neumeister, P.; Hoefler, G.; Beham-Schmid, C.; Linkesch, W. Histiocytic sarcoma–Targeted therapy: Novel therapeutic options? A series of 4 cases. Onkologie 2012, 35, 447–450. [Google Scholar] [CrossRef]
- Fidel, J.; Schiller, I.; Hauser, B.; Jausi, Y.; Rohrer-Bley, C.; Roos, M.; Kaser-Hotz, B. Histiocytic sarcomas in flat-coated retrievers: a summary of 37 cases (November 1998-March 2005). Vet. Comp. Oncol. 2006, 4, 63–74. [Google Scholar] [CrossRef]
- Hornick, J.L.; Jaffe, E.S.; Fletcher, C.D.M. Extranodal histiocytic sarcoma: Clinicopathologic analysis of 14 cases of a rare epithelioid malignancy. Am. J. Surg. Pathol. 2004, 28, 1133–1144. [Google Scholar] [CrossRef]
- Wellman, M.L.; Krakowka, S.; Jacobs, R.M.; Kociba, G.J. A macrophage-monocyte cell line from a dog with malignant histiocytosis. Vitr. Cell. Dev. Biol. 1988, 24, 223–229. [Google Scholar] [CrossRef]
- Puff, C.; Krudewig, C.; Imbschweiler, I.; Baumgärtner, W.; Alldinger, S. Influence of persistent canine distemper virus infection on expression of RECK, matrix-metalloproteinases and their inhibitors in a canine macrophage/monocytic tumour cell line (DH82). Vet. J. 2009, 182, 100–107. [Google Scholar] [CrossRef]
- Pfankuche, V.M.; Sayed-Ahmed, M.; Contioso, V.B.; Spitzbarth, I.; Rohn, K.; Ulrich, R.; Deschl, U.; Kalkuhl, A.; Baumgärtner, W.; Puff, C. Persistent morbillivirus infection leads to altered cortactin distribution in histiocytic sarcoma cells with decreased cellular migration capacity. PLoS ONE 2016, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Wojton, J.; Kaur, B. Impact of tumor microenvironment on oncolytic viral therapy. Cytokine Growth Factor Rev. 2010, 21, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Chung, Y.H.; Johnston, R.N. Reovirus and tumor oncolysis. J. Microbiol. 2007, 45, 187–192. [Google Scholar]
- Gopas, J.; Itzhaky, D.; Segev, Y.; Salzberg, S.; Trink, B.; Isakov, N.; Rager-Zisman, B. Persistent measles virus infection enhances major histocompatibility complex class I expression and immunogenicity of murine neuroblastoma cells. Cancer Immunol. Immunother. 1992, 34, 313–320. [Google Scholar] [CrossRef]
- Schattner, A.; Rager-Zisman, B.; Bloom, B.R. Persistent viral infection affects tumorigenicity of a neuroblastoma cell line. Cell. Immunol. 1985, 90, 103–114. [Google Scholar] [CrossRef]
- Challapalli, A.; Carroll, L.; Aboagye, E.O. Molecular mechanisms of hypoxia in cancer. Clin. Transl. Imaging 2017, 5, 225–253. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxidants Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr Biol 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Attig, F.; Spitzbarth, I.; Kalkuhl, A.; Deschl, U.; Puff, C.; Baumgärtner, W.; Ulrich, R. Reactive oxygen species are key mediators of demyelination in canine distemper leukoencephalitis but not in Theiler’s murine encephalomyelitis. Int. J. Mol. Sci. 2019, 20, 3217. [Google Scholar] [CrossRef] [PubMed]
- Bürge, T.; Griot, C.; Vandevelde, M.; Peterhans, E. Antiviral antibodies stimulate production of reactive oxygen species in cultured canine brain cells infected with canine distemper virus. J. Virol. 1989, 63, 2790–2797. [Google Scholar] [CrossRef] [PubMed]
- Griot, C.; Bürge, T.; Vandevelde, M.; Peterhans, E. Antibody-induced generation of reactive oxygen radicals by brain macrophages in canine distemper encephalitis: a mechanism for bystander demyelination. Acta Neuropathol. 1989, 78, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Karadeniz, A.; Hanedan, B.; Cemek, M.; Börkü, M.K. Relationship between canine distemper and oxidative stress in dogs. Rev. Med. Vet. (Toulouse). 2008, 159, 462–467. [Google Scholar]
- Stein, V.M.; Czub, M.; Schreiner, N.; Moore, P.F.; Vandevelde, M.; Zurbriggen, A.; Tipold, A. Microglial cell activation in demyelinating canine distemper lesions. J. Neuroimmunol. 2004, 153, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Brunner, J.M.; Plattet, P.; Doucey, M.A.; Rosso, L.; Curie, T.; Montagner, A.; Wittek, R.; Vandelvelde, M.; Zurbriggen, A.; Hirling, H.; et al. Morbillivirus glycoprotein expression induces ER stress, alters Ca2+ homeostasis and results in the release of vasostatin. PLoS ONE 2012, 7, 1–12. [Google Scholar] [CrossRef]
- Bhandary, B.; Marahatta, A.; Kim, H.R.; Chae, H.J. An involvement of oxidative stress in endoplasmic reticulum stress and its associated diseases. Int. J. Mol. Sci. 2013, 14, 434–456. [Google Scholar] [CrossRef]
- Fiaschi, T.; Chiarugi, P. Oxidative stress, tumor microenvironment, and metabolic reprogramming: A diabolic liaison. Int. J. Cell Biol. 2012, 2012. [Google Scholar] [CrossRef]
- Semenza, G.L. Oxygen sensing, hypoxia-inducible factors, and disease pathophysiology. Annu. Rev. Pathol. Mech. Dis. 2014, 9, 47–71. [Google Scholar] [CrossRef]
- Zepeda, A.B.; Pessoa, A.; Castillo, R.L.; Figueroa, C.A.; Pulgar, V.M.; Farías, J.G. Cellular and molecular mechanisms in the hypoxic tissue: Role of HIF-1 and ROS. Cell Biochem. Funct. 2013, 31, 451–459. [Google Scholar] [CrossRef]
- Ushio-Fukai, M.; Nakamura, Y. Reactive oxygen species and angiogenesis NADPH oxidase as target for cancer therapy. Cancer Lett. 2008, 266, 37–52. [Google Scholar] [CrossRef]
- Krock, B.L.; Skuli, N.; Simon, M.C. Hypoxia-induced angiogenesis: good and evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef] [PubMed]
- Alldinger, S.; Baumgärtner, W.; Kremmer, E.; Fonfara, S. Characterization of a canine CD44 specific monoclonal antibody. J. Vet. Med. Ser. A Physiol. Pathol. Clin. Med. 1999, 46, 19–32. [Google Scholar] [CrossRef]
- Kolesnikov, N.; Hastings, E.; Keays, M.; Melnichuk, O.; Tang, Y.A.; Williams, E.; Dylag, M.; Kurbatova, N.; Brandizi, M.; Burdett, T.; et al. ArrayExpress update-simplifying data submissions. Nucleic Acids Res. 2015, 43, D1113–D1116. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Reactive oxygen species and cancer paradox: To promote or to suppress? Free Radic. Biol. Med. 2017, 104, 144–164. [Google Scholar] [CrossRef]
- Klaunig, J.E.; Kamendulis, L.M.; Hocevar, B.A. Oxidative stress and oxidative damage in carcinogenesis. Toxicol. Pathol. 2010, 38, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Yates, B.; Braschi, B.; Gray, K.A.; Seal, R.L.; Tweedie, S.; Bruford, E.A. Genenames.org: The HGNC and VGNC resources in 2017. Nucleic Acids Res. 2017, 45, D619–D625. [Google Scholar] [CrossRef] [PubMed]
- Dalman, M.R.; Deeter, A.; Nimishakavi, G.; Duan, Z.H. Fold change and p-value cutoffs significantly alter microarray interpretations. BMC Bioinform. 2012, 13, S11. [Google Scholar] [CrossRef] [PubMed]
- Haverkamp, A.K.; Lehmbecker, A.; Spitzbarth, I.; Widagdo, W.; Haagmans, B.L.; Segalés, J.; Vergara-Alert, J.; Bensaid, A.; Van Den Brand, J.M.A.; Osterhaus, A.D.M.E.; et al. Experimental infection of dromedaries with Middle East respiratory syndrome-Coronavirus is accompanied by massive ciliary loss and depletion of the cell surface receptor dipeptidyl peptidase. Sci. Rep. 2018, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Allnoch, L.; Baumgärtner, W.; Hansmann, F. Impact of astrocyte depletion upon inflammation and demyelination in a murine animal model of multiple sclerosis. Int. J. Mol. Sci. 2019, 20, 3922. [Google Scholar] [CrossRef] [PubMed]
- Brogden, G.; Shammas, H.; Maalouf, K.; Naim, S.L.; Wetzel, G.; Amiri, M.; Von Köckritz-Blickwede, M.; Das, A.M.; Naim, H.Y. Case study on the pathophysiology of Fabry disease: Abnormalities of cellular membranes can be reversed by substrate reduction in vitro. Biosci. Rep. 2017, 37, 1–10. [Google Scholar] [CrossRef]
- Dahlgren, C.; Karlsson, A.; Bylund, J. Intracellular neutrophil oxidants: from laboratory curiosity to clinical reality. J. Immunol. 2019, 202, 3127–3134. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yu, Q.; Chng, W.J. TXNIP (VDUP-1, TBP-2): A major redox regulator commonly suppressed in cancer by epigenetic mechanisms. Int. J. Biochem. Cell Biol. 2011, 43, 1668–1673. [Google Scholar] [CrossRef] [PubMed]
- Cha, Y.J.; Baik, J.E.; Rhee, W.J. Inhibition of endoplasmic reticulum stress-induced apoptosis by silkworm storage protein 1. Biotechnol. Bioprocess Eng. 2018, 23, 194–200. [Google Scholar] [CrossRef]
- Kaufman, R.J. Stress signaling from the lumen of the endoplasmic reticulum: Coordination of gene transcriptional and translational controls. Genes Dev. 1999, 13, 1211–1233. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. The endoplasmic reticulum and the unfolded protein response. Semin. Cell Dev. Biol. 2007, 18, 716–731. [Google Scholar] [CrossRef]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef]
- Aga, M.; Bentz, G.L.; Raffa, S.; Torrisi, M.R.; Kondo, S.; Wakisaka, N.; Yoshizaki, T.; Pagano, J.S.; Shackelford, J. Exosomal HIF1α supports invasive potential of nasopharyngeal carcinoma-associated LMP1-positive exosomes. Oncogene 2014, 33, 4613–4622. [Google Scholar] [CrossRef]
- Marie, J.C.; Saltel, F.; Escola, J.-M.; Jurdic, P.; Wild, T.F.; Horvat, B. Cell surface delivery of the measles virus nucleoprotein: a viral strategy to induce immunosuppression. J. Virol. 2004, 78, 11952–11961. [Google Scholar] [CrossRef]
- Weis, S.M.; Cheresh, D.A. Tumor angiogenesis: Molecular pathways and therapeutic targets. Nat. Med. 2011, 17, 1359–1370. [Google Scholar] [CrossRef]
- Pani, G.; Galeotti, T.; Chiarugi, P. Metastasis: Cancer cell’s escape from oxidative stress. Cancer Metastasis Rev. 2010, 29, 351–378. [Google Scholar] [CrossRef] [PubMed]
- Van Horssen, J.; Schreibelt, G.; Drexhage, J.; Hazes, T.; Dijkstra, C.D.; van der Valk, P.; de Vries, H.E. Severe oxidative damage in multiple sclerosis lesions coincides with enhanced antioxidant enzyme expression. Free Radic. Biol. Med. 2008, 45, 1729–1737. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Stephenson, J.R.; Ter Meulen, V. Antigenic relationships between measles and canine distemper viruses: Comparison of immune response in animals and humans to individual virus-specific polypeptides. Proc. Natl. Acad. Sci. USA 1979, 76, 6601–6605. [Google Scholar] [CrossRef]
- Lal, N.; Puri, K.; Rodrigues, B. Vascular Endothelial Growth Factor B and Its Signaling. Front. Cardiovasc. Med. 2018, 5, 1–9. [Google Scholar] [CrossRef]
- Bry, M.; Kivelä, R.; Leppänen, V.M.; Alitalo, K. Vascular endothelial growth factor-B in physiology and disease. Physiol. Rev. 2014, 94, 779–794. [Google Scholar] [CrossRef]
- Li, X.; Lee, C.; Tang, Z.; Zhang, F.; Arjunan, P.; Li, Y.; Hou, X.; Kumar, A.; Dong, L. VEGF-B. A survival, or an angiogenic factor? Cell Adhes. Migr. 2009, 3, 322–327. [Google Scholar] [CrossRef]
- Abd-Aziz, N.; Stanbridge, E.J.; Shafee, N. Newcastle disease virus degrades HIF-1α through proteasomal pathways independent of VHL and p53. J. Gen. Virol. 2016, 97, 3174–3182. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primary Antibody | Host Species, Clonality | Epitope Retrieval | Serum Blocking | Dilution | Secondary Antibody (1:200) | Positive Control |
|---|---|---|---|---|---|---|
| CDV-NP (University of Bern) | Mouse, monoclonal, clone D110 | Citrate buffer, microwave (800 W, 20´) | PBST + 3% BSA + 5% goat serum | 1:100 | GaM-Cy3 or GaM-Cy2 | n/a |
| 8OHdG (Abcam, Cambridge, USA) | Goat, polyclonal | Proteinase K | PBST + 3% BSA + 5% horse serum | 1:200 | DaG-Cy3 | Canine pyo-granu-lomatous endo- metritis |
| SOD2 (Abcam, Cambridge, USA) | Rabbit, polyclonal | Citrate buffer, microwave (800 W, 20´) | PBST + 3% BSA + 5% goat serum | 1:200 | GaR-Cy2 | Canine brain and spinal cord |
| CAT (Abcam, Cambridge, USA) | Goat, polyclonal | Citrate buffer, microwave (800 W, 20´) | PBST + 3% BSA + 5% horse serum | 1:50 | DaG-Cy3 | Canine spinal cord with fibro-carti- lagineous embolus |
| HIF-1α (Novus Biologicals, Colorado, USA) | Rabbit, polyclonal | Citrate buffer, microwave (800 W, 20´) | PBST + 3% BSA + 5% goat serum | 1:500 | GaR-Cy3 or DaR-Cy2 | Canine mammary adeno- carcinoma with central necrosis |
| Wheat germ agglutinin (WGA) AF633 conjugated (Invitrogen, California, USA) | none | Citrate buffer, microwave (800 W, 20´) | n/a | 1:20 | n/a | n/a |
| CD63 (Sicgen, Coimbra, Portugal) | Goat, polyclonal | Citrate buffer, microwave (800 W, 20´) | PBST + 3% BSA + 5% horse serum | 1:200 | DaG-Cy3 | MDCK cell pellet |
| GM-130 (BD Transduction Laboratories, North Carolina, USA) | Mouse, monoclonal, clone 35/GM130 (RUO) | Citrate buffer, microwave (800 W, 20´) | PBST + 3% BSA + 5% goat serum | 1:200 | GaM-Cy2 | n/a |
| VEGF-B (My Biosource, California, USA) | Rabbit, polyclonal | Citrate buffer, microwave (800 W, 20´) | PBST + 3% BSA + 5% goat serum | 1:40 | GaR-Cy3 | Canine fetal brain, liver and kidney |
| Canine Gene Symbol | Gene Name | Functional Group | Fold Change | p-Value | References |
|---|---|---|---|---|---|
| VEGF-B | vascular endothelial growth factor B | HIF-1a downstream | −593.197 | <0.001 | [29,39,40,41,45] |
| THBS2 | thrombospondin 2 | HIF-1a downstream | −451.295 | <0.001 | [42] |
| EDN1 | endothelin 1 | HIF-1a downstream | −47.795 | <0.001 | [42] |
| CXCR4 | chemokine (C-X-C motif) receptor 4 | HIF-1a downstream | −13.485 | <0.001 | [39] |
| SERPINE1 | serine (or cysteine) peptidase inhibitor, clade E, member 1 | HIF-1a downstream | −13.116 | <0.001 | [41,42] |
| COX7B2 | cytochrome c oxidase subunit VIIb2 | ROS production; ER stress | −6.015 | <0.001 | [29,31,45] |
| ITPR3 | inositol 1,4,5-triphosphate receptor, type 3 | ER stress | −4.646 | <0.001 | [37] |
| THBS1 | thrombospondin 1 | HIF-1a downstream | −4.461 | <0.001 | [42] |
| ERO1L | ERO1-like (S. cerevisiae) | ROS production; ER stress | −3.995 | <0.001 | [37] |
| Cxcl12 | chemokine (C-X-C motif) ligand 12 | HIF-1a downstream | −3.683 | <0.001 | [39] |
| NT5E | 5’-nucleotidase, ecto (CD73) | HIF-1a downstream | −3.041 | <0.001 | [39] |
| CANX | calnexin | ER stress | −2.780 | <0.001 | [36] |
| TXNRD3 | thioredoxin reductase 3 | ROS scavenging | −2.464 | <0.001 | [29,45] |
| NDUFAF2 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, assembly factor 2 | ROS production; ER stress | −2.292 | <0.001 | [29,45] |
| NDUFAB1 | NADH dehydrogenase (ubiquinone) 1, alpha/beta subcomplex, 1, 8kDa | ROS production; ER stress | −2.261 | <0.001 | [29,45] |
| DDIT3 | DNA-damage-inducible transcript 3 | ER stress | −2.087 | <0.001 | [36] |
| EGLN1 | Egl nine homolog 1 (C. elegans) | HIF-1a transcription & regulation | −1.976 | 0.001 | [39,40,42,46] |
| PRDX6 | peroxiredoxin 6 | ROS scavenging | −1.895 | <0.001 | [29,45] |
| EGLN3 | egl nine homolog 3 (C. elegans) | HIF-1a transcription & regulation | −1.875 | 0.004 | [39,40,42,46] |
| SDHD | succinate dehydrogenase complex, subunit D, integral membrane protein | ROS production; ER stress | −1.857 | <0.001 | [29,45] |
| FGF2 | fibroblast growth factor 2 (basic) | HIF-1a downstream | −1.842 | 0.003 | [42] |
| PDIA6 | protein disulfide isomerase family A, member 6 | ROS production; ER stress | −1.801 | <0.001 | [37] |
| VHL | von Hippel-Lindau tumor suppressor, E3 ubiquitin protein ligase | HIF-1a transcription & regulation | −1.771 | 0.005 | [39,40,42,45,46] |
| SOD1 | superoxide dismutase 1, soluble | ROS scavenging | −1.712 | <0.001 | [29,45] |
| PDIA4 | protein disulfide isomerase family A, member 4 | ROS production; ER stress | −1.678 | 0.010 | [37] |
| ADM | adrenomedullin | HIF-1a downstream | −1.665 | <0.001 | [42] |
| GSS | glutathione synthetase | ROS scavenging; ER stress | −1.648 | 0.001 | [37] |
| NDUFC2 | NADH dehydrogenase (ubiquinone) 1, subcomplex unknown, 2, 14.5kDa | ROS production; ER stress | −1.630 | 0.001 | [29,45] |
| GCLM | glutamate-cysteine ligase, modifier subunit | ROS scavenging; ER stress | −1.565 | <0.001 | [37] |
| PDIA3 | protein disulfide isomerase family A, member 3 | ROS production; ER stress | −1.533 | 0.001 | [37] |
| CD274 | CD274 molecule | HIF-1a downstream | −1.515 | 0.025 | [39] |
| PDGFRL | platelet-derived growth factor receptor-like | HIF-1a downstream | 1.554 | 0.004 | [39] |
| UQCR11 | ubiquinol-cytochrome c reductase (6.4kD) subunit | ROS production; ER stress | 1.563 | 0.002 | [29,45] |
| UQCRC2 | ubiquinol cytochrome c reductase core protein 2 | ROS production; ER stress | 1.590 | 0.021 | [29,45] |
| NDUFS1 | NADH dehydrogenase (ubiquinone) Fe-S protein 1, 75kDa (NADH-coenzyme Q reductase) | ROS production; ER stress | 1.622 | <0.001 | [29,45] |
| NCF2 | neutrophil cytosolic factor 2 | ROS production | 1.639 | 0.004 | [29] |
| UQCRC1 | ubiquinol-cytochrome c reductase core protein 1 | ROS production; ER stress | 1.678 | <0.001 | [29,45] |
| ITPR1 | inositol 1,4,5-triphosphate receptor, type 1 | ER stress | 1.844 | 0.001 | [37] |
| NDUFS7 | NADH dehydrogenase (ubiquinone) Fe-S protein 7, 20kDa (NADH-coenzyme Q reductase) | ROS production; ER stress | 1.846 | <0.001 | [29,45] |
| LONP1 | lon peptidase 1, mitochondrial | ER stress | 1.850 | 0.001 | [37] |
| CCL2 | chemokine (C-C motif) ligand 2 | HIF-1a downstream | 1.866 | <0.001 | [29,42] |
| HMOX1 | heme oxygenase (decycling) 1 | ROS scavenging | 1.940 | <0.001 | [29] |
| NDUFA10 | NADH dehydrogenase (ubiquinone) 1 alpha subcomplex, 10, 42kDa | ROS production; ER stress | 2.009 | <0.001 | [29,45] |
| PDGFA | platelet-derived growth factor alpha polypeptide | HIF-1a downstream | 2.089 | <0.001 | [39] |
| PPID | peptidylprolyl isomerase D (cyclophilin D) | ER stress | 2.286 | <0.001 | [29,45] |
| NDUFV3 | NADH dehydrogenase (ubiquinone) flavoprotein 3 | ROS production; ER stress | 2.362 | <0.001 | [29,45] |
| ALOX5AP | arachidonate 5-lipoxygenase-activating protein | ROS production | 2.509 | <0.001 | [45] |
| COX17 | COX17 homolog, cytochrome c oxidase assembly protein | ROS production; ER stress | 2.557 | 0.001 | [29,31,45] |
| CAT | Catalase | ROS scavenging | 3.584 | <0.001 | [29,45] |
| NQO1 | NAD(P)H dehydrogenase, quinone 1 | ROS scavenging | 3.868 | <0.001 | [29] |
| XDH | xanthine dehydrogenase | ROS production; ER stress | 3.913 | 0.002 | [29,37,45] |
| KITLG | KIT ligand | HIF-1a downstream | 4.174 | <0.001 | [39] |
| LOC100856470 | peroxiredoxin-2-like | ROS scavenging | 5.351 | <0.001 | [29,45] |
| TEK | endothelial-specific receptor tyrosine kinase | HIF-1a downstream | 5.639 | <0.001 | [39,41,42] |
| PDGFC | platelet derived growth factor C | HIF-1a downstream | 6.578 | <0.001 | [39] |
| TXNIP | thioredoxin interacting protein | ROS scavenging | 11.227 | 0.001 | [29] |
| NCF4 | neutrophil cytosolic factor 4, 40kDa | ROS production | 67.304 | <0.001 | [29,41] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armando, F.; Gambini, M.; Corradi, A.; Giudice, C.; Pfankuche, V.M.; Brogden, G.; Attig, F.; von Köckritz-Blickwede, M.; Baumgärtner, W.; Puff, C. Oxidative Stress in Canine Histiocytic Sarcoma Cells Induced by an Infection with Canine Distemper Virus Led to a Dysregulation of HIF-1α Downstream Pathway Resulting in a Reduced Expression of VEGF-B In Vitro. Viruses 2020, 12, 200. https://doi.org/10.3390/v12020200
Armando F, Gambini M, Corradi A, Giudice C, Pfankuche VM, Brogden G, Attig F, von Köckritz-Blickwede M, Baumgärtner W, Puff C. Oxidative Stress in Canine Histiocytic Sarcoma Cells Induced by an Infection with Canine Distemper Virus Led to a Dysregulation of HIF-1α Downstream Pathway Resulting in a Reduced Expression of VEGF-B In Vitro. Viruses. 2020; 12(2):200. https://doi.org/10.3390/v12020200
Chicago/Turabian StyleArmando, Federico, Matteo Gambini, Attilio Corradi, Chiara Giudice, Vanessa Maria Pfankuche, Graham Brogden, Friederike Attig, Maren von Köckritz-Blickwede, Wolfgang Baumgärtner, and Christina Puff. 2020. "Oxidative Stress in Canine Histiocytic Sarcoma Cells Induced by an Infection with Canine Distemper Virus Led to a Dysregulation of HIF-1α Downstream Pathway Resulting in a Reduced Expression of VEGF-B In Vitro" Viruses 12, no. 2: 200. https://doi.org/10.3390/v12020200
APA StyleArmando, F., Gambini, M., Corradi, A., Giudice, C., Pfankuche, V. M., Brogden, G., Attig, F., von Köckritz-Blickwede, M., Baumgärtner, W., & Puff, C. (2020). Oxidative Stress in Canine Histiocytic Sarcoma Cells Induced by an Infection with Canine Distemper Virus Led to a Dysregulation of HIF-1α Downstream Pathway Resulting in a Reduced Expression of VEGF-B In Vitro. Viruses, 12(2), 200. https://doi.org/10.3390/v12020200