An Engineered Microvirin Variant with Identical Structural Domains Potently Inhibits Human Immunodeficiency Virus and Hepatitis C Virus Cellular Entry

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Protein Expression
2.2. NMR Experiments
2.3. HIV Inhibition Assay
2.4. HCV Infection Assay
2.5. HCV Replication Assay
2.6. HCV Pseudoparticle Assay
2.7. Flow Cytometry Analysis of PBMCs
2.8. MTT Assay
3. Results
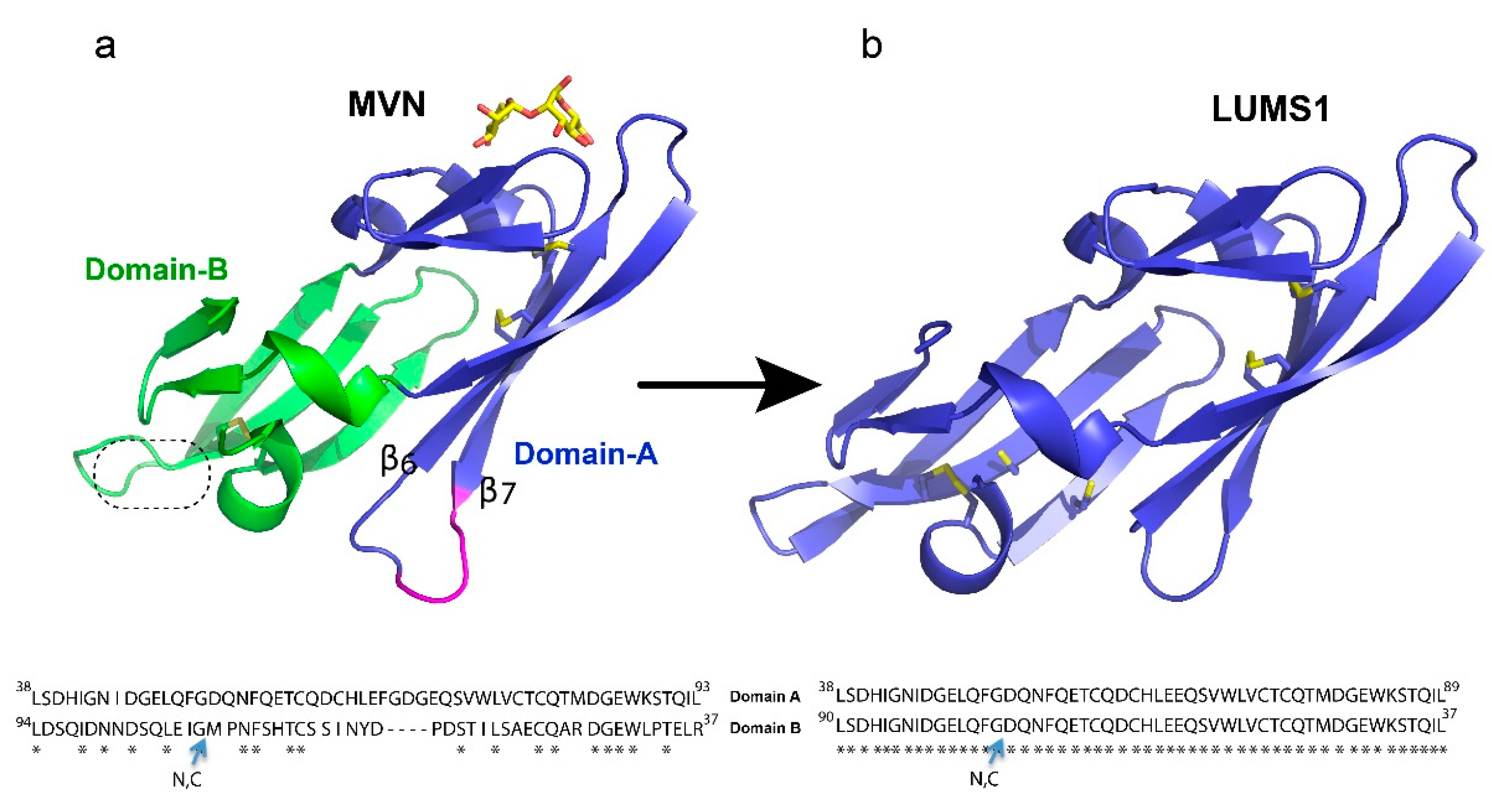
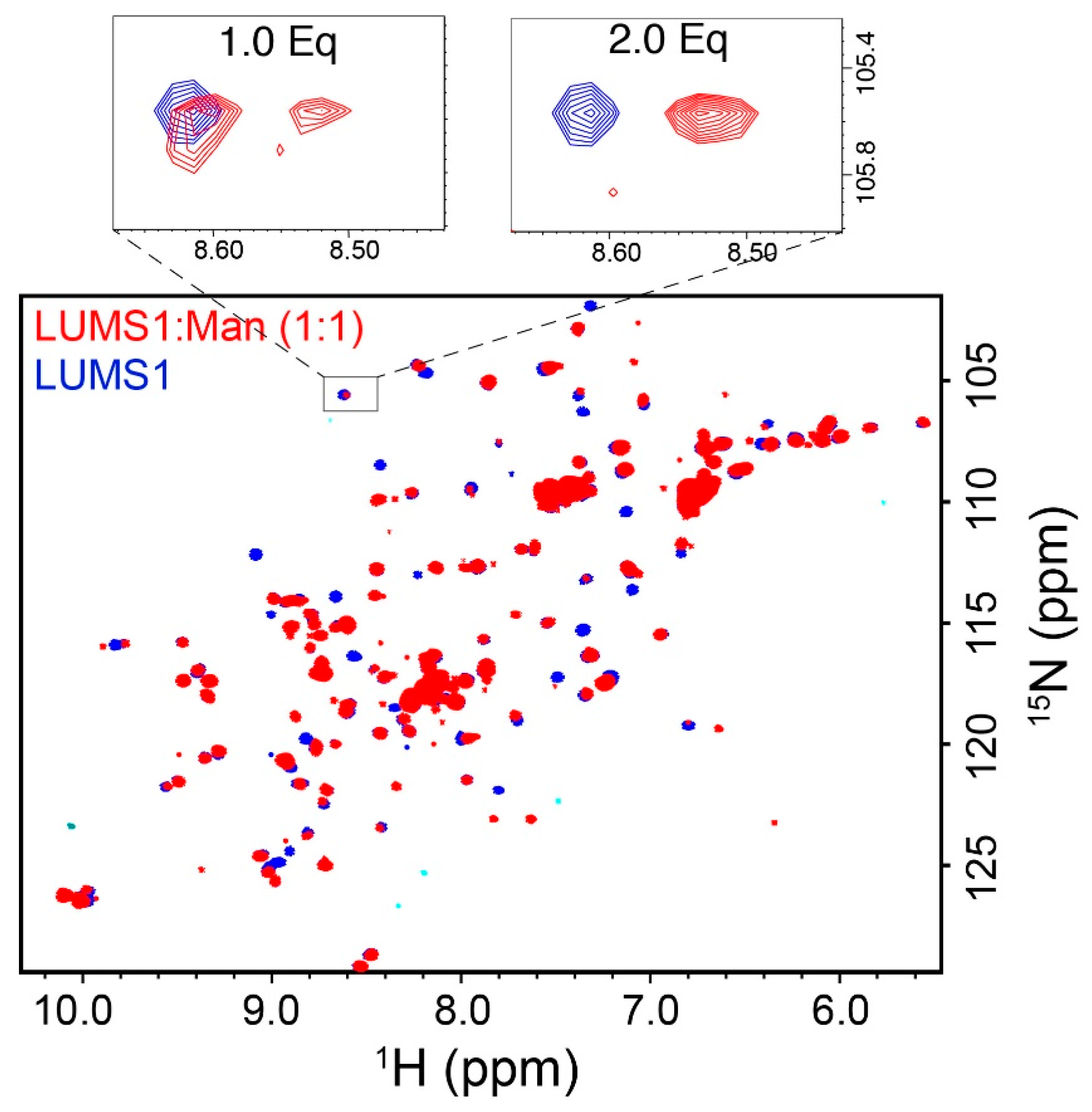
3.1. Designing of the LUMS1 Protein and Characterization of Its Carbohydrate Binding
3.2. LUMS1 Inhibits HIV-1 Cellular Entry
3.3. LUMS1 Inhibits HCV Cellular Entry
3.4. LUMS1 Does Not Stimulate Cellular Activation Markers and Shows Negligible Cytotoxic Effect
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jefferies, M.; Rauff, B.; Rashid, H.; Lam, T.; Rafiq, S. Update on global epidemiology of viral hepatitis and preventive strategies. World J. Clin. Cases 2018, 6, 589–599. [Google Scholar] [CrossRef]
- Mahy, M.; Marsh, K.; Sabin, K.; Wanyeki, I.; Daher, J.; Ghys, P.D. HIV estimates through 2018: Data for decision making. Aids 2019. [CrossRef]
- Ashraf, M.U.; Iman, K.; Khalid, M.F.; Salman, H.M.; Shafi, T.; Rafi, M.; Javaid, N.; Hussain, R.; Ahmad, F.; Shahzad-Ul-Hussan, S.; et al. Evolution of efficacious pangenotypic hepatitis C virus therapies. Med. Res. Rev. 2019, 39, 1091–1136. [Google Scholar] [CrossRef] [PubMed]
- Dhiman, R.K.; Grover, G.S.; Premkumar, M. Hepatitis C elimination: A Public Health Perspective. Curr. Treat. Options Gastroenterol. 2019. [CrossRef] [PubMed]
- Kanters, S.; Vitoria, M.; Doherty, M.; Socias, M.E.; Ford, N.; Forrest, J.I.; Popoff, E.; Bansback, N.; Nsanzimana, S.; Thorlund, K.; et al. Comparative efficacy and safety of first-line antiretroviral therapy for the treatment of HIV infection: A systematic review and network meta-analysis. Lancet HIV 2016, 3, e510–e520. [Google Scholar] [CrossRef]
- Goffard, A.; Dubuisson, J. Glycosylation of hepatitis C virus envelope proteins. Biochimie 2003, 85, 295–301. [Google Scholar] [CrossRef]
- Doores, K.J.; Bonomelli, C.; Harvey, D.J.; Vasiljevic, S.; Dwek, R.A.; Burton, D.R.; Crispin, M.; Scanlan, C.N. Envelope glycans of immunodeficiency virions are almost entirely oligomannose antigens. Proc. Natl. Acad. Sci. USA 2010, 107, 13800–13805. [Google Scholar] [CrossRef]
- Fenouillet, E.; Gluckman, J.C.; Jones, I.M. Functions of HIV envelope glycans. Trends Biochem. Sci. 1994, 19, 65–70. [Google Scholar] [CrossRef]
- Shahzad-ul-Hussan, S.; Ghirlando, R.; Dogo-Isonagie, C.I.; Igarashi, Y.; Balzarini, J.; Bewley, C.A. Characterization and carbohydrate specificity of pradimicin S. J. Am. Chem. Soc. 2012, 134, 12346–12349. [Google Scholar] [CrossRef]
- Balzarini, J. Carbohydrate-binding agents: A potential future cornerstone for the chemotherapy of enveloped viruses? Antivir. Chem. Chemother. 2007, 18, 1–11. [Google Scholar] [CrossRef]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C., 2nd; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.W., Jr.; McMahon, J.B.; et al. Isolation and characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J. Biol. Chem. 2005, 280, 9345–9353. [Google Scholar] [CrossRef] [PubMed]
- Akkouh, O.; Ng, T.B.; Singh, S.S.; Yin, C.; Dan, X.; Chan, Y.S.; Pan, W.; Cheung, R.C. Lectins with anti-HIV activity: A review. Molecules 2015, 20, 648–668. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Li, J.; Patel, S.K. Design of Poly(lactic-co-glycolic Acid) (PLGA) Nanoparticles for Vaginal Co-Delivery of Griffithsin and Dapivirine and Their Synergistic Effect for HIV Prophylaxis. Pharmaceutics 2019, 11, 184. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, B.R.; Vojdani, F.; Buffa, V.; Shattock, R.J.; Montefiori, D.C.; Bakke, J.; Mirsalis, J.; d’Andrea, A.L.; Hume, S.D.; Bratcher, B.; et al. Scaleable manufacture of HIV-1 entry inhibitor griffithsin and validation of its safety and efficacy as a topical microbicide component. Proc. Natl. Acad. Sci. USA 2009, 106, 6099–6104. [Google Scholar] [CrossRef] [PubMed]
- Huskens, D.; Schols, D. Algal lectins as potential HIV microbicide candidates. Mar. Drugs 2012, 10, 1476–1497. [Google Scholar] [CrossRef]
- Griffithsin-Based Rectal Microbicide for PREvention of Viral ENTry (PREVENT). Available online: https://clinicaltrials.gov/ct2/show/NCT04032717 (accessed on 9 November 2019).
- Kehr, J.C.; Zilliges, Y.; Springer, A.; Disney, M.D.; Ratner, D.D.; Bouchier, C.; Seeberger, P.H.; de Marsac, N.T.; Dittmann, E. A mannan binding lectin is involved in cell-cell attachment in a toxic strain of Microcystis aeruginosa. Mol. Microbiol. 2006, 59, 893–906. [Google Scholar] [CrossRef]
- Huskens, D.; Ferir, G.; Vermeire, K.; Kehr, J.C.; Balzarini, J.; Dittmann, E.; Schols, D. Microvirin, a novel alpha(1,2)-mannose-specific lectin isolated from Microcystis aeruginosa, has anti-HIV-1 activity comparable with that of cyanovirin-N but a much higher safety profile. J. Biol. Chem. 2010, 285, 24845–24854. [Google Scholar] [CrossRef]
- Shahzad-ul-Hussan, S.; Gustchina, E.; Ghirlando, R.; Clore, G.M.; Bewley, C.A. Solution structure of the monovalent lectin microvirin in complex with Man(alpha)(1-2)Man provides a basis for anti-HIV activity with low toxicity. J. Biol. Chem. 2011, 286, 20788–20796. [Google Scholar] [CrossRef]
- Gustchina, E.; Louis, J.M.; Lam, S.N.; Bewley, C.A.; Clore, G.M. A monoclonal Fab derived from a human nonimmune phage library reveals a new epitope on gp41 and neutralizes diverse human immunodeficiency virus type 1 strains. J. Virol. 2007, 81, 12946–12953. [Google Scholar] [CrossRef]
- Lee, M.; Yang, J.; Park, S.; Jo, E.; Kim, H.Y.; Bae, Y.S.; Windisch, M.P. Micrococcin P1, a naturally occurring macrocyclic peptide inhibiting hepatitis C virus entry in a pan-genotypic manner. Antivir. Res. 2016, 132, 287–295. [Google Scholar] [CrossRef]
- Hsu, M.; Zhang, J.; Flint, M.; Logvinoff, C.; Cheng-Mayer, C.; Rice, C.M.; McKeating, J.A. Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped retroviral particles. Proc. Natl. Acad. Sci. USA 2003, 100, 7271–7276. [Google Scholar] [CrossRef] [PubMed]
- Baldick, C.J.; Wichroski, M.J.; Pendri, A.; Walsh, A.W.; Fang, J.; Mazzucco, C.E.; Pokornowski, K.A.; Rose, R.E.; Eggers, B.J.; Hsu, M.; et al. A novel small molecule inhibitor of hepatitis C virus entry. PLoS Pathog. 2010, 6, e1001086. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of cell viability by the MTT assay. Cold Spring Harb. Protoc. 2018, 2018, pdbprot095505. [Google Scholar] [CrossRef] [PubMed]
- Bewley, C.A.; Shahzad-Ul-Hussan, S. Characterizing carbohydrate-protein interactions by nuclear magnetic resonance spectroscopy. Biopolymers 2013, 99, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.P.; Reynolds, H.M.; Lumicisi, B.; Bryson, C.J. Immunogenicity of protein therapeutics: The key causes, consequences and challenges. Self Nonself 2010, 1, 314–322. [Google Scholar] [CrossRef] [PubMed]
- Ito, S.; Ikuno, T.; Mishima, M.; Yano, M.; Hara, T.; Kuramochi, T.; Sampei, Z.; Wakabayashi, T. In vitro human helper T-cell assay to screen antibody drug candidates for immunogenicity. J. Immunotoxicol. 2019, 16, 125–132. [Google Scholar] [CrossRef]
- Huskens, D.; Vermeire, K.; Vandemeulebroucke, E.; Balzarini, J.; Schols, D. Safety concerns for the potential use of cyanovirin-N as a microbicidal anti-HIV agent. Int. J. Biochem. Cell. Biol. 2008, 40, 2802–2814. [Google Scholar] [CrossRef]
- Lee, J.H.; Ozorowski, G.; Ward, A.B. Cryo-EM structure of a native, fully glycosylated, cleaved HIV-1 envelope trimer. Science 2016, 351, 1043–1048. [Google Scholar] [CrossRef]
- Dam, T.K.; Brewer, C.F. Effects of clustered epitopes in multivalent ligand-receptor interactions. Biochemistry 2008, 47, 8470–8476. [Google Scholar] [CrossRef]
- Li, Y.; Kang, C. Solution NMR Spectroscopy in Target-Based Drug Discovery. Molecules 2017, 22, 1399. [Google Scholar] [CrossRef]
- Min, Y.Q.; Duan, X.C.; Zhou, Y.D.; Kulinich, A.; Meng, W.; Cai, Z.P.; Ma, H.Y.; Liu, L.; Zhang, X.L.; Voglmeir, J. Effects of microvirin monomers and oligomers on hepatitis C virus. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Takebe, Y.; Saucedo, C.J.; Lund, G.; Uenishi, R.; Hase, S.; Tsuchiura, T.; Kneteman, N.; Ramessar, K.; Tyrrell, D.L.; Shirakura, M.; et al. Antiviral lectins from red and blue-green algae show potent in vitro and in vivo activity against hepatitis C virus. PLoS ONE 2013, 8, e64449. [Google Scholar] [CrossRef] [PubMed]
- Kachko, A.; Loesgen, S.; Shahzad-Ul-Hussan, S.; Tan, W.; Zubkova, I.; Takeda, K.; Wells, F.; Rubin, S.; Bewley, C.A.; Major, M.E. Inhibition of hepatitis C virus by the cyanobacterial protein Microcystis viridis lectin: Mechanistic differences between the high-mannose specific lectins MVL, CV-N, and GNA. Mol. Pharm. 2013, 10, 4590–4602. [Google Scholar] [CrossRef] [PubMed]
- Helle, F.; Wychowski, C.; Vu-Dac, N.; Gustafson, K.R.; Voisset, C.; Dubuisson, J. Cyanovirin-N inhibits hepatitis C virus entry by binding to envelope protein glycans. J. Biol. Chem. 2006, 281, 25177–25183. [Google Scholar] [CrossRef] [PubMed]
- Goffard, A.; Callens, N.; Bartosch, B.; Wychowski, C.; Cosset, F.L.; Montpellier, C.; Dubuisson, J. Role of N-linked glycans in the functions of hepatitis C virus envelope glycoproteins. J. Virol. 2005, 79, 8400–8409. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay System | Inhibitor | EC50 (nM) |
|---|---|---|
| HCVcc | LUMS1 | 45.3 ± 18.6 |
| MVN # | 35.6 ± 3.98 | |
| HCV replicon | LUMS1 | n.d. |
| Sofosbuvir | 54.0 ± 32.8 | |
| HCVpp | LUMS1 | 142.1 ± 23.0 |
| HIV-1 single
round infectivity | LUMS1 | 37.2 ± 4.4 |
| MVN | 8.0 ± 1.4 |
| Cell Types | CC50 of LUMS1 (nM) |
|---|---|
| Huh7.5 | >10,000 |
| PBMCs | >10,000 |
| TZMbl | 4900 ± 166 |
| HepG2 | >10,000 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahid, M.; Qadir, A.; Yang, J.; Ahmad, I.; Zahid, H.; Mirza, S.; Windisch, M.P.; Shahzad-ul-Hussan, S. An Engineered Microvirin Variant with Identical Structural Domains Potently Inhibits Human Immunodeficiency Virus and Hepatitis C Virus Cellular Entry. Viruses 2020, 12, 199. https://doi.org/10.3390/v12020199
Shahid M, Qadir A, Yang J, Ahmad I, Zahid H, Mirza S, Windisch MP, Shahzad-ul-Hussan S. An Engineered Microvirin Variant with Identical Structural Domains Potently Inhibits Human Immunodeficiency Virus and Hepatitis C Virus Cellular Entry. Viruses. 2020; 12(2):199. https://doi.org/10.3390/v12020199
Chicago/Turabian StyleShahid, Munazza, Amina Qadir, Jaewon Yang, Izaz Ahmad, Hina Zahid, Shaper Mirza, Marc P. Windisch, and Syed Shahzad-ul-Hussan. 2020. "An Engineered Microvirin Variant with Identical Structural Domains Potently Inhibits Human Immunodeficiency Virus and Hepatitis C Virus Cellular Entry" Viruses 12, no. 2: 199. https://doi.org/10.3390/v12020199
APA StyleShahid, M., Qadir, A., Yang, J., Ahmad, I., Zahid, H., Mirza, S., Windisch, M. P., & Shahzad-ul-Hussan, S. (2020). An Engineered Microvirin Variant with Identical Structural Domains Potently Inhibits Human Immunodeficiency Virus and Hepatitis C Virus Cellular Entry. Viruses, 12(2), 199. https://doi.org/10.3390/v12020199