
Sindbis Virus Infection in Non-Blood-Fed Hibernating Culex pipiens Mosquitoes in Sweden

Abstract
:1. Introduction
2. Materials and Methods
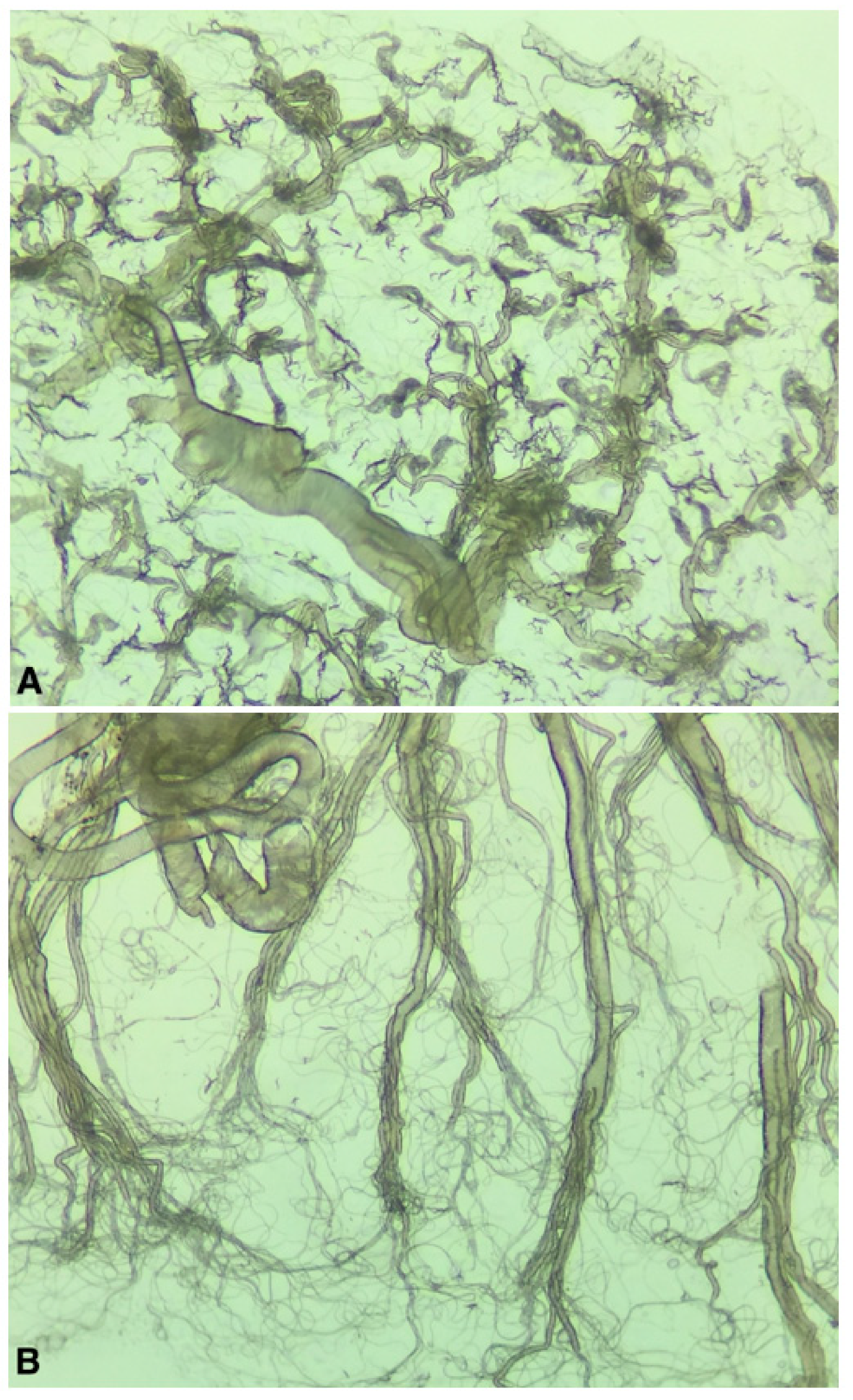
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rosen, L. The Natural history of Japanese encephalitis virus. Ann. Rev. Mirobiol. 1986, 40, 395–414. [Google Scholar] [CrossRef] [PubMed]
- Nasci, R.S.; Savage, H.M.; White, D.J.; Miller, J.R.; Cropp, B.C.; Godsey, M.S.; Kerst, A.J.; Bennett, P.; Gottfried, K.; Lanciotti, R.S. West Nile virus in overwintering Culex mosquitoes, New York City, 2000. Emerg. Infect. Dis. 2001, 7, 742–744. [Google Scholar] [CrossRef] [PubMed]
- Farajollahi, A.; Crans, W.J.; Bryant, P.; Wolf, B.; Burkhalter, K.L.; Godsey, M.S.; Aspen, S.E.; Nasci, R.S. Detection of West Nile viral RNA from an overwintering pool of Culex pipens pipiens (Diptera: Culicidae) in New Jersey, 2003. J. Med. Entomol. 2005, 42, 490–494. [Google Scholar] [CrossRef] [Green Version]
- Monaco, F.; Savini, G.; Calistri, P.; Polci, A.; Pinoni, C.; Bruno, R.; Lelli, R. 2009 West Nile disease epidemic in Italy: First evidence of overwintering in Western Europe? Res. Vet. Sci. 2011, 91, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Rudolf, I.; Betášová1, L.; Blažejová, H.; Venclíková, K.; Straková, P.; Šebesta, O.; Mendel, J.; Bakonyi, T.; Schaffner, F.; Nowotny, N.; et al. West Nile virus in overwintering mosquitoes, central Europe. Parasites Vectors 2017, 10, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Service, M.W. The taxonomy and biology of two sympatric sibling species of Culex, C. pipiens and C. torrentium (Diptera, Culicidae). J. Zool. 1968, 156, 313–323. [Google Scholar] [CrossRef]
- Jaenson, T.G.T. Overwintering of Culex mosquitoes in Sweden and their potential as reservoirs of human pathogens. Med. Vet. Entomol. 1987, 1, 151–156. [Google Scholar] [CrossRef]
- Kurkela, S.; Helve, T.; Vaheri, A.; Vapalahti, O. Arthritis and arthralgia three years after Sindbis virus infection: Clinical follow-up of a cohort of 49 patients. Scand. J. Infect. Dis. 2008, 40, 167–173. [Google Scholar] [CrossRef]
- Hesson, J.C.; Lundström, J.O.; Tok, A.; Östman, Ö.; Lundkvist, Å. Temporal variation in Sindbis virus antibody prevalence in bird hosts in an endemic area in Sweden. PLoS ONE 2016, 11, e0162005. [Google Scholar] [CrossRef] [Green Version]
- Adouchief, S.; Smura, T.; Sane, J.; Vapalahti, O.; Kurkela, S. Sindbis virus as a human pathogen—Epidemiology, clinical picture and pathogenesis. Rev. Med. Virol. 2016, 26, 221–241. [Google Scholar] [CrossRef]
- Ling, J.; Smura, T.; Lundström, J.O.; Pettersson, J.H.-O.; Sironen, T.; Vapalahti, O.; Lundkvist, Å.; Hesson, J.C. The introduction and dispersal of Sindbis virus from central Africa to Europe. J. Virol. 2019, 93, e00620-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, N.; Petric, D.; Boase, C.; Lane, J.; Zgomba, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control; Springer: Boston, MA, USA, 2003; pp. 113–286. ISBN 978-1-4757-5897-9. [Google Scholar]
- Kardos, E.H.; Bellamy, R.E. Examination of the ovarian tracheation. Ann. Entomol. Soc. Am. 1961, 54, 448–451. [Google Scholar] [CrossRef]
- Jöst, H.; Bialonski, A.; Storch, V.; Günther, S.; Becker, N.; Schmidt-Chanasit, J. Isolation and Phylogenetic Analysis of Sindbis Viruses from Mosquitoes in Germany. J. Clin. Microbiol. 2010, 48, 1900–1903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesson, J.C.; Lundström, J.O.; Halvarsson, P.; Erixon, P.; Collado, A. A sensitive and reliable restriction enzyme assay to distinguish between the mosquitoes Culex torrentium and Culex pipiens. Med. Vet. Entomol. 2010, 24, 142–149. [Google Scholar] [CrossRef]
- Hesson, J.C.; Verner-Carlsson, J.; Larsson, A.; Ahmed, R.; Lundkvist, Å.; Lundström, J.O. Culex torrentium Mosquito Role as Major Enzootic Vector Defined by Rate of Sindbis Virus Infection, Sweden, 2009. Emerg. Infect. Dis. 2015, 21, 875–878. [Google Scholar] [CrossRef]
- Lundström, J.O.; Hesson, J.C.; Schäfer, M.L.; Östman, Ö.; Semmler, T.; Bekaert, M.; Weidmann, M.; Lundkvist, Å.; Pfeffer, M. Sindbis virus polyarthritis outbreak signalled by virus prevalence in the mosquito vectors. PLoS Negl. Trop. Dis. 2019, 13, e0007702. [Google Scholar] [CrossRef] [Green Version]
- Bailey, C.L.; Faran, M.E.; Gargan, T.P., 2nd; Hayes, D.E. Winter survival of blood-fed and nonblood-fed Culex pipiens L. Am. J. Trop. Med. Hyg. 1982, 31, 1054–1061. [Google Scholar] [CrossRef]
- Hayes, E.B.; Komar, N.; Nasci, R.S.; Montgomery, S.P.; O’Leary, D.R.; Campbell, G.L. Epidemiology and transmission dynamics of West Nile virus disease. Emerg. Infect. Dis. 2005, 11, 1167–1173. [Google Scholar] [CrossRef]
- Oda, T.; Wada, Y.; Hayashi, K. Considerations on the possibility of Japanese encephalitis virus in Culex tritaeniorhynchus females. Trop. Med. 1978, 20, 153–155. [Google Scholar]
- Oda, T.; Mori, A.; Fujita, K.; Wada, Y. Follicular development in Culex tritaeniorhynchus females showing gonotrophic dissociation. Trop. Med. 1981, 23, 105–110. [Google Scholar]
- Mitchell, C.J. Differentiation of Host-Seeking Behavior from Blood-Feeding Behavior in Overwintering Culex Pipiens (Diptera: Culicidae) and Observations on Gonotrophic Dissociation. J. Med. Entomol. 1983, 20, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.J.; Briegel, H. Inability of diapausing Culex pipiens (Diptera, Culicidae) to use blood for producing lipid reserves for overwinter survival. J. Med. Entomol. 1989, 26, 318–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelms, B.M.; Macedo, P.A.; Kothera, L.; Savage, H.M.; Reisen, W.K. Overwintering Biology of Culex (Diptera: Culicidae) Mosquitoes in the Sacramento Valley of California. J. Med. Entomol. 2013, 50, 773–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lequime, S.; Paul, R.E.; Lambrechts, L. Determinants of Arbovirus Vertical Transmission in Mosquitoes. PLoS Pathog. 2016, 12, e1005548. [Google Scholar] [CrossRef] [PubMed]
- Dohm, D.J.; Turell, M.J. Effect of Incubation at Overwintering Temperatures on the Replication of West Nile Virus in New York Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2001, 38, 462–464. [Google Scholar] [CrossRef] [PubMed]
- Lundström, J.O.; Niklasson, B.; Francy, D.B. Swedish Culex torrentium and Cx. pipiens (Diptera: Culicidae) as experimental vectors of Ockelbo virus. J. Med. Entomol. 1990, 27, 561–563. [Google Scholar] [CrossRef]
- Hesson, J.C.; Östman, Ö.; Schäfer, M.; Lundström, J.O. Geographic distribution and relative abundance of the sibling vector species Culex torrentium and Culex pipiens in Sweden. Vector Borne Zoonotic Dis. 2011, 11, 1383–1389. [Google Scholar] [CrossRef]
- Leggewie, M.; Badusche, M.; Rudolf, M.; Jansen, S.; Börstler, J.; Krumkamp, R.; Huber, K.; Krüger, A.; Schmidt-Chanasit, J.; Tannich, E.; et al. Culex pipiens and Culex torrentium populations from Central Europe are susceptible to West Nile virus infection. One Health 2016, 2, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Jansen, S.; Heitmann, A.; Lühken, R.; Leggewie, M.; Helms, M.; Badusche, M.; Rossini, G.; Schmidt-Chanasit, J.; Tannich, E. Culex torrentium: A Potent Vector for the Transmission of West Nile Virus in Central Europe. Viruses 2019, 11, 492. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Parity Status | Collection Period | Incubation Time (Days) | PCR Positive | Sequencing Positive |
|---|---|---|---|---|---|
| Cx. pipiens | nulliparous | 6 December | 5 | yes | yes |
| Cx. pipiens | unknown | 6 December | 5 | yes | no |
| Cx. pipiens | unknown | 6 December | 5 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 10 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 10 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 10 | yes | no |
| Cx. pipiens | nulliparous | 7 December | 10 | yes | no |
| Cx. pipiens | nulliparous | 7 December | 11 | yes | yes |
| Cx. pipiens | parous | 7 December | 11 | yes | yes |
| unknown | unknown | 7 December | 12 | yes | no |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | unknown | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | unknown | 7 December | 12 | yes | no |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | no |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | no |
| unknown | nulliparous | 7 December | 12 | yes | no |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | unknown | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | no |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | parous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | nulliparous | 7 December | 12 | yes | yes |
| Cx. pipiens | unknown | 7 December | 12 | yes | yes |
| unknown | unknown | 15 February | 11 | yes | no |
| Cx. pipiens | nulliparous | 15 February | 11 | yes | no |
| Incubation Time (Days) | Collection Period | Number Analyzed | SINV Positive: Number (Percent) |
|---|---|---|---|
| 4 | December | 10 | 0 (0) |
| February | 40 | 0 (0) | |
| 5 | December | 50 | 3 (6) |
| February | NA | NA | |
| 6 | December | NA | NA |
| February | 50 | 0 (0) | |
| 7 | December | 10 | 0 (0) |
| February | 140 | 0 (0) | |
| 8 | December | 32 | 0 (0) |
| February | NA | NA | |
| 10 | December | 88 | 4 (4.5) |
| February | 40 | 0 (0) | |
| 11 | December | 80 | 2 (2.5) |
| February | 17 | 2 (12) | |
| 12 | December | 110 | 24 (23) |
| February | NA | NA | |
| 13 | December | 100 | 0 (0) |
| February | NA | NA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergman, A.; Dahl, E.; Lundkvist, Å.; Hesson, J.C. Sindbis Virus Infection in Non-Blood-Fed Hibernating Culex pipiens Mosquitoes in Sweden. Viruses 2020, 12, 1441. https://doi.org/10.3390/v12121441
Bergman A, Dahl E, Lundkvist Å, Hesson JC. Sindbis Virus Infection in Non-Blood-Fed Hibernating Culex pipiens Mosquitoes in Sweden. Viruses. 2020; 12(12):1441. https://doi.org/10.3390/v12121441
Chicago/Turabian StyleBergman, Alexander, Emma Dahl, Åke Lundkvist, and Jenny C. Hesson. 2020. "Sindbis Virus Infection in Non-Blood-Fed Hibernating Culex pipiens Mosquitoes in Sweden" Viruses 12, no. 12: 1441. https://doi.org/10.3390/v12121441
APA StyleBergman, A., Dahl, E., Lundkvist, Å., & Hesson, J. C. (2020). Sindbis Virus Infection in Non-Blood-Fed Hibernating Culex pipiens Mosquitoes in Sweden. Viruses, 12(12), 1441. https://doi.org/10.3390/v12121441