
Bat-Borne Coronaviruses in Jordan and Saudi Arabia: A Threat to Public Health?

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Mass Influxes and Gatherings
2.1. Jordan
2.2. Saudi Arabia
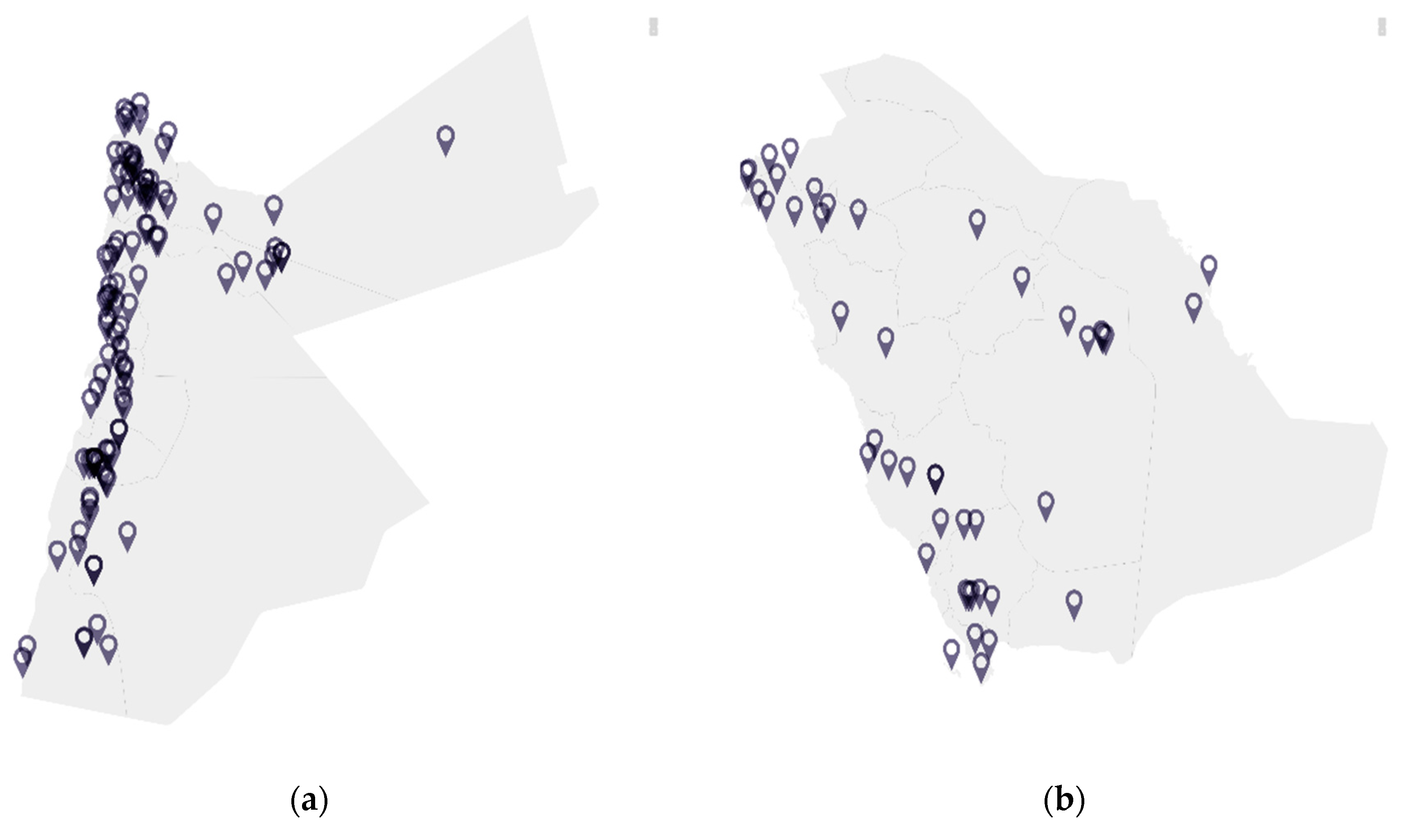
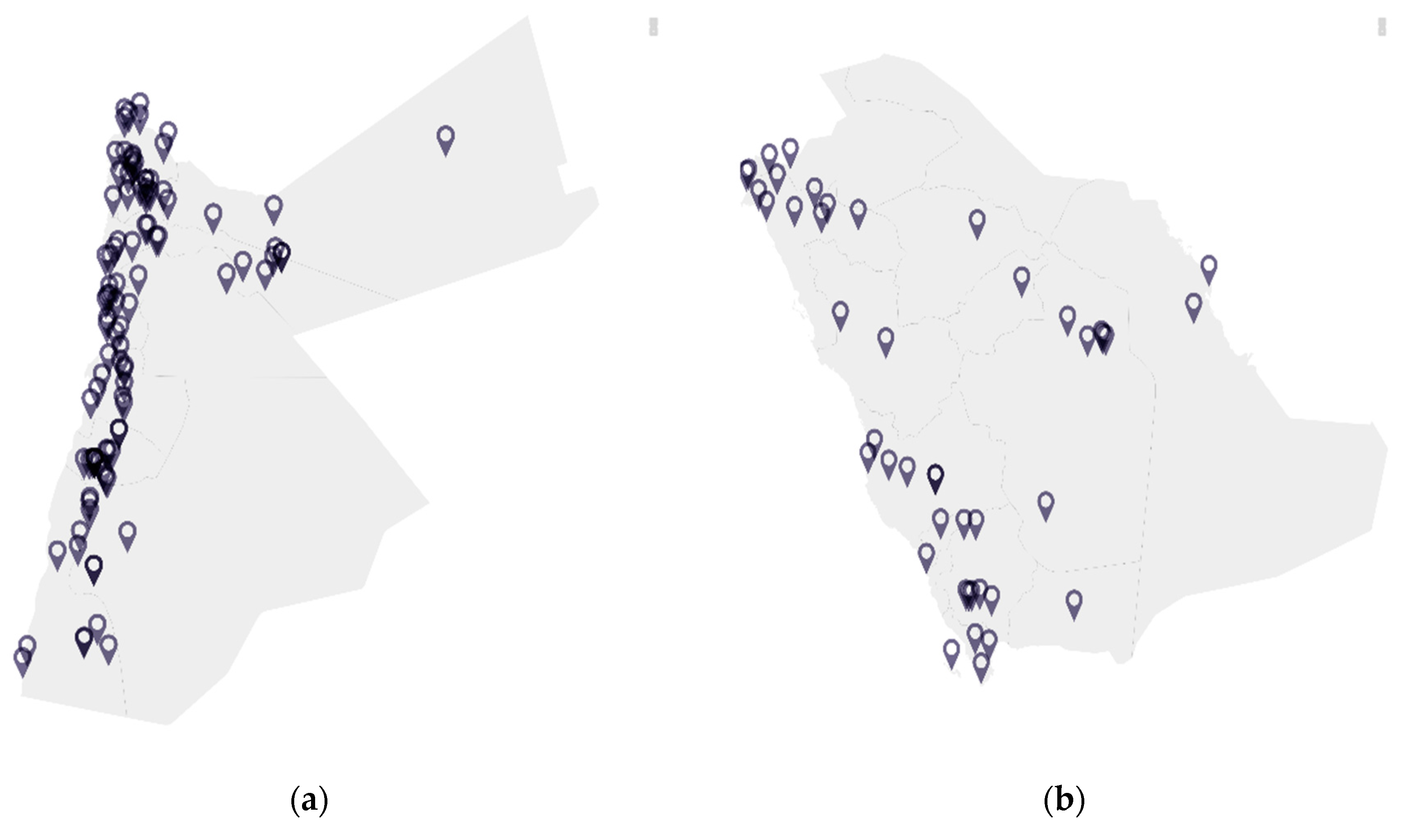
3. Distribution of Bats and Associated Pathogens
3.1. Jordan
3.2. Saudi Arabia
4. Coronaviruses in the Middle East: MERS-CoV and SARS-CoV-2
4.1. Middle East Respiratory Syndrome-Related Coronavirus (MERS-CoV)
4.1.1. Jordan
4.1.2. Saudi Arabia
4.2. SARS-CoV-2
4.2.1. Jordan
4.2.2. Saudi Arabia
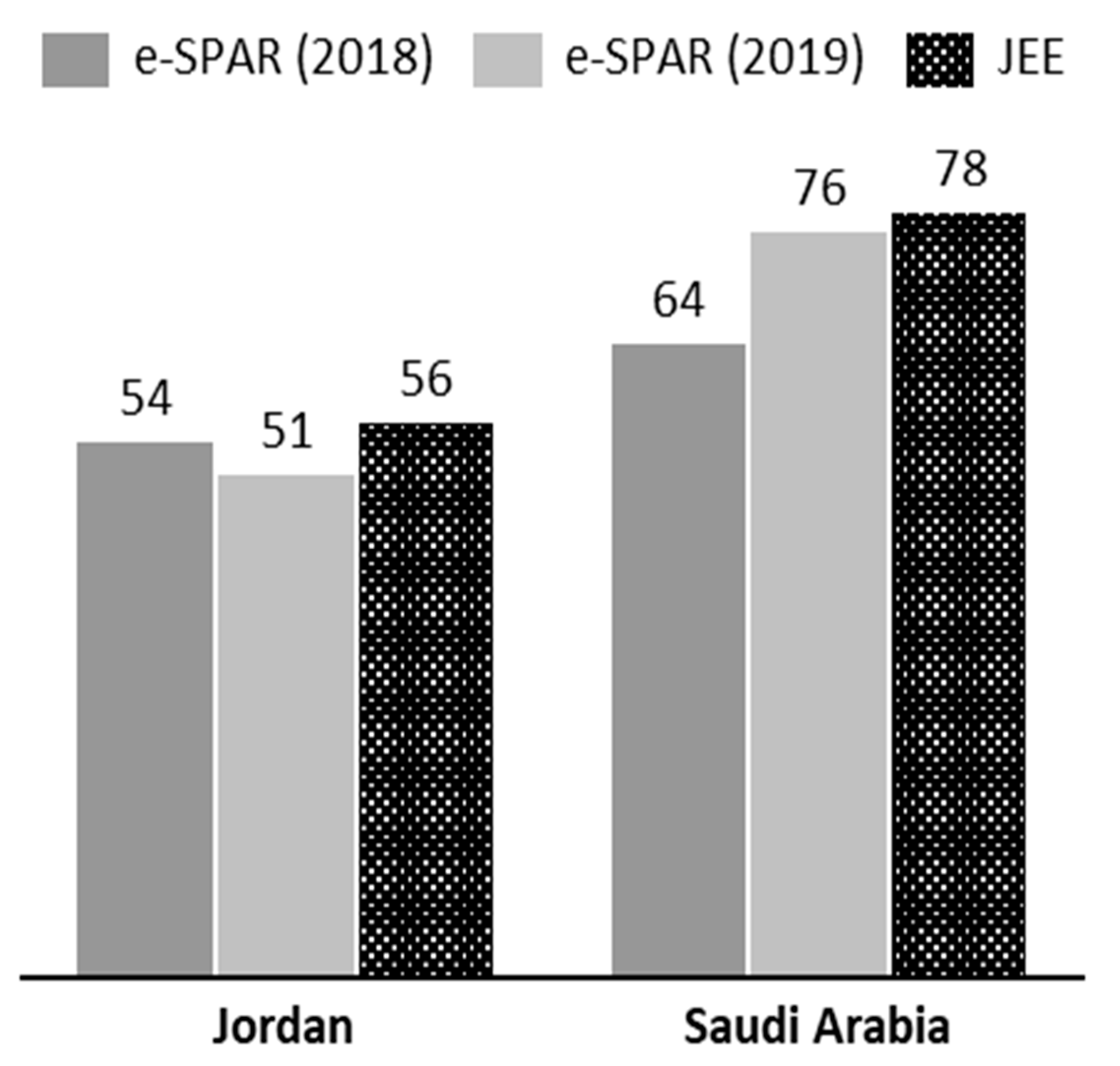
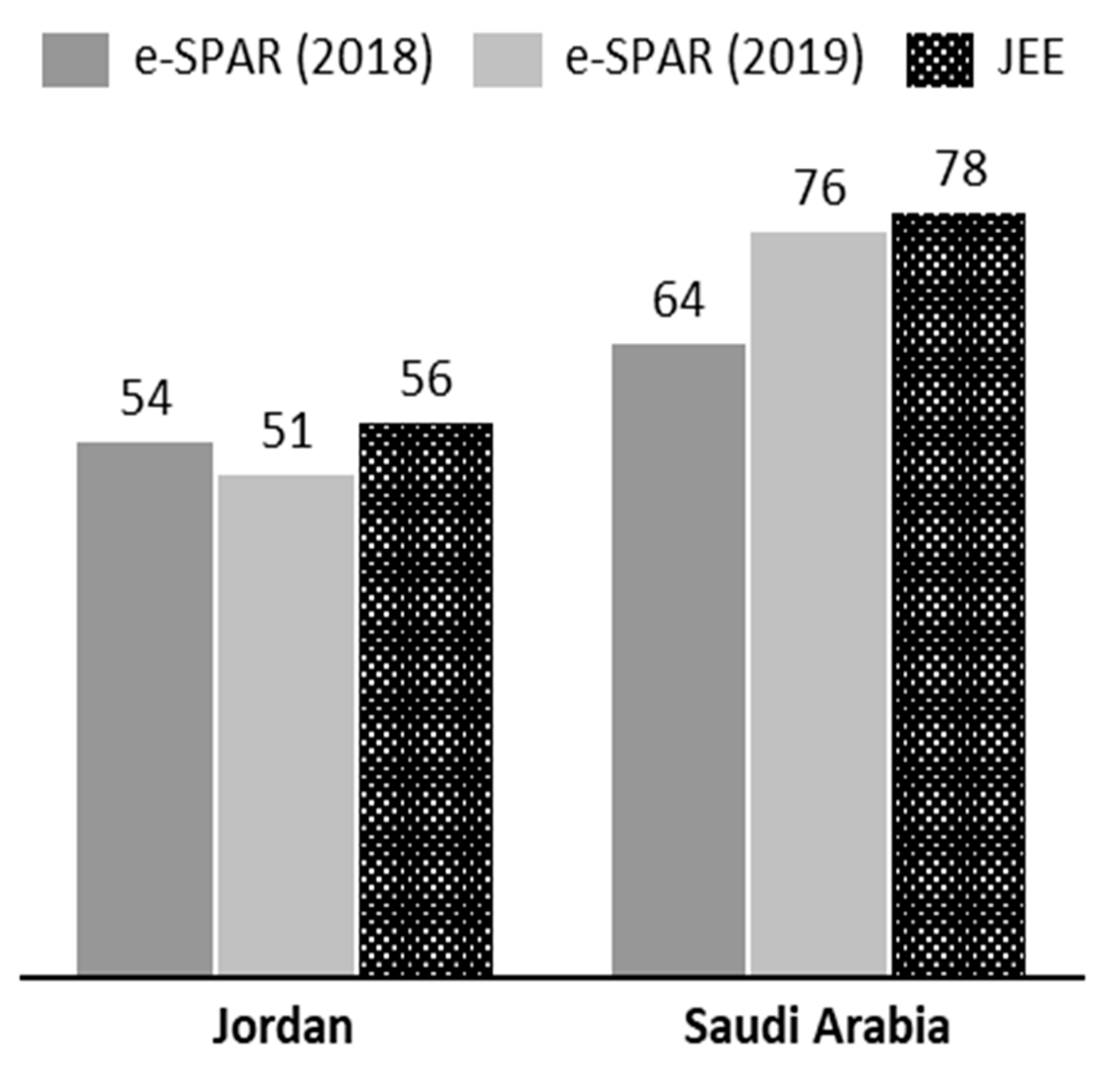
5. Compliance with the International Health Regulations
5.1. Jordan
5.2. Saudi Arabia
6. Conclusions
7. Recommendations and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Henig, R.M. Experts Warned of a Pandemic Decades Ago. Why Weren’t We Ready? Available online: https://www.nationalgeographic.com/science/2020/04/experts-warned-pandemic-decades-ago-why-not-ready-for-coronavirus/ (accessed on 19 July 2020).
- Fauci, A.S.; Lane, H.C.; Redfield, R.R. Covid-19 - Navigating the uncharted. N. Engl. J. Med. 2020, 382, 1268–1269. [Google Scholar] [CrossRef] [PubMed]
- Van Doorn, H.R. Emerging infectious diseases. Medicine (United Kingdom) 2014, 42, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, J.N.S.; Desai, M.A.; Levy, K.; Bates, S.J.; Liang, S.; Naumoff, K.; Scott, J.C. Environmental determinants of infectious disease: A framework for tracking causal links and guiding public health research. Environ. Health Perspect. 2007, 115, 1216–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Childs, J.E.; Richt, J.A.; Mackenzie, J.S. Introduction: Conceptualizing and partitioning the emergence process of zoonotic viruses from wildlife to humans. Curr. Top. Microbiol. Immunol. 2007, 315, 1–31. [Google Scholar]
- Marston, D.A.; Banyard, A.C.; McElhinney, L.M.; Freuling, C.M.; Finke, S.; de Lamballerie, X.; Müller, T.; Fooks, A.R. The lyssavirus host-specificity conundrum—Rabies virus—The exception not the rule. Curr. Opin. Virol. 2018, 28, 68–73. [Google Scholar] [CrossRef]
- Quan, P.L.; Firth, C.; Conte, J.M.; Williams, S.H.; Zambrana-Torrelio, C.M.; Anthony, S.J.; Ellison, J.A.; Gilbert, A.T.; Kuzmin, I.V.; Niezgoda, M.; et al. Bats are a major natural reservoir for hepaciviruses and pegiviruses. Proc. Natl. Acad. Sci. USA 2013, 110, 8194–8199. [Google Scholar] [CrossRef] [Green Version]
- Woo, P.C.Y.; Lau, S.K.P.; Huang, Y.; Yuen, K.Y. Coronavirus diversity, phylogeny and interspecies jumping. Exp. Biol. Med. 2009, 234, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Calisher, C.H.; Childs, J.E.; Field, H.E.; Holmes, K.V.; Schountz, T. Bats: Important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 2006, 19, 531–545. [Google Scholar] [CrossRef] [Green Version]
- Drexler, J.F.; Corman, V.M.; Müller, M.A.; Maganga, G.D.; Vallo, P.; Binger, T.; Gloza-Rausch, F.; Rasche, A.; Yordanov, S.; Seebens, A.; et al. Bats host major mammalian paramyxoviruses. Nat. Commun. 2012, 3, 796. [Google Scholar] [CrossRef] [Green Version]
- Luis, A.D.; Hayman, D.T.S.; O’Shea, T.J.; Cryan, P.M.; Gilbert, A.T.; Pulliam, J.R.C.; Mills, J.N.; Timonin, M.E.; Willis, C.K.R.; Cunningham, A.A.; et al. A comparison of bats and rodents as reservoirs of zoonotic viruses: Are bats special? Proc. R. Soc. B Biol. Sci. 2013, 280, 20122753. [Google Scholar] [CrossRef] [Green Version]
- Mollentze, N.; Streicker, D.G. Viral zoonotic risk is homogenous among taxonomic orders of mammalian and avian reservoir hosts. Proc. Natl. Acad. Sci. USA 2020, 117, 9423–9430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peel, A.J.; Wells, K.; Giles, J.; Boyd, V.; Burroughs, A.; Edson, D.; Crameri, G.; Baker, M.L.; Field, H.; Wang, L.F.; et al. Synchronous shedding of multiple bat paramyxoviruses coincides with peak periods of Hendra virus spillover. Emerg. Microbes Infect. 2019, 8, 1314–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plowright, R.K.; Peel, A.J.; Streicker, D.G.; Gilbert, A.T.; McCallum, H.; Wood, J.; Baker, M.L.; Restif, O. Transmission or Within-Host Dynamics Driving Pulses of Zoonotic Viruses in Reservoir–Host Populations. PLoS Negl. Trop. Dis. 2016, 10, e0004796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plowright, R.K.; Eby, P.; Hudson, P.J.; Smith, I.L.; Westcott, D.; Bryden, W.L.; Middleton, D.; Reid, P.A.; McFarlane, R.A.; Martin, G.; et al. Ecological dynamics of emerging bat virus spillover. Proc. R. Soc. B Biol. Sci. 2014, 282, 20142124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montecino-Latorre, D.; Goldstein, T.; Gilardi, K.; Wolking, D.; Van Wormer, E.; Kazwala, R.; Ssebide, B.; Nziza, J.; Sijali, Z.; Cranfield, M.; et al. Reproduction of East-African bats may guide risk mitigation for coronavirus spillover. One Health Outlook 2020, 2, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Shannan, G. Communicable diseases in the Mediterranean region. Electron. J. Int. Fed. Clin. Chem. Lab. Med. 2018, 29, 164–170. [Google Scholar]
- Abubakar, A.; Malik, M.; Pebody, R.; Elkholy, A.; Khan, W.; Bellos, A.; Mala, P. Burden of acute respiratory disease of epidemic and pandemic potential in the WHO Eastern Mediterranean Region: A literature review. East. Mediterr. Health J. 2016, 22, 509–522. [Google Scholar] [CrossRef]
- Malik, M.R.; Mahjour, J. Preparedness for ebola: Can it transform our current public health system? East. Mediterr. Health J. 2016, 22, 566–567. [Google Scholar] [CrossRef]
- Mokdad, A.H.; Forouzanfar, M.H.; Daoud, F.; El Bcheraoui, C.; Moradi-Lakeh, M.; Khalil, I.; Afshin, A.; Tuffaha, M.; Charara, R.; Barber, R.M.; et al. Health in times of uncertainty in the eastern Mediterranean region, 1990–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet Glob. Health 2016, 4, e704–e713. [Google Scholar] [CrossRef] [Green Version]
- Al-Tawfiq, J.A.; Memish, Z.A. COVID-19 in the Eastern Mediterranean Region and Saudi Arabia: Prevention and therapeutic strategies. Int. J. Antimicrob. Agents 2020, 55, 105968. [Google Scholar] [CrossRef]
- Kluge, H.H.P.; Jakab, Z.; Bartovic, J.; D’Anna, V.; Severoni, S. Refugee and migrant health in the COVID-19 response. Lancet 2020, 395, 1237–1239. [Google Scholar] [CrossRef]
- Fehr, A.R.; Perlman, S. Coronaviruses: An overview of their replication and pathogenesis. In Coronaviruses: Methods and Protocols; Maier, H., Bickerton, E., Britton, P., Eds.; Humana Press: New York, NY, USA, 2015; Volume 1282, pp. 1–23. ISBN 9781493924387. [Google Scholar]
- Memish, Z.A.; Steffen, R.; White, P.; Dar, O.; Azhar, E.I.; Sharma, A.; Zumla, A. Mass gatherings medicine: Public health issues arising from mass gathering religious and sporting events. Lancet 2019, 393, 2073–2084. [Google Scholar] [CrossRef]
- Vermeulen, G.; Desmet, E. Essential Texts on European and International Asylum and Migration Law and Policy; Gompel & Svacina: Reebokweg, Belgium, 2018. [Google Scholar]
- Memish, Z.A.; Al-Rabeeah, A.A. Public health management of mass gatherings: The saudi arabian experience with MERS-CoV. Bull. World Health Organ. 2013, 91, 899. [Google Scholar] [CrossRef] [PubMed]
- Alshoubaki, W. The Dynamics of Population Pressure in Jordan: A Focus on Syrian Refugees. In Syrian Crisis, Syrian Refugees; Beaujouan, J., Rasheed, A., Eds.; Palgrave Pivot: Cham, Switzerland, 2020; pp. 27–34. [Google Scholar]
- Achilli, L. Syrian Refugees in Jordan: A Reality Check. Migr. Policy Cent. Eur. Univ. Inst. 2015, 2, 1–12. [Google Scholar]
- Alshoubaki, W. A synopsis of the jordanian governance system in the management of the syrian refugee crisis. J. Intercult. Stud. 2018, 39, 596–603. [Google Scholar] [CrossRef]
- Maani, N. From refugee camp to resilient city: Zaatari refugee camp, Jordan. Footprint 2016, 2016, 145–148. [Google Scholar]
- Al-Tal, R.S.; Ahmad Ghanem, H.H. Impact of the Syrian crisis on the socio-spatial transformation of Eastern Amman, Jordan. Front. Archit. Res. 2019, 8, 591–603. [Google Scholar] [CrossRef]
- Akik, C.; Ghattas, H.; Mesmar, S.; Rabkin, M.; El-Sadr, W.M.; Fouad, F.M. Host country responses to non-communicable diseases amongst Syrian refugees: A review. Confl. Health 2019, 13, 8. [Google Scholar] [CrossRef]
- McNatt, Z.Z.; Freels, P.E.; Chandler, H.; Fawad, M.; Qarmout, S.; Al-Oraibi, A.S.; Boothby, N. “What’s happening in Syria even affects the rocks”: A qualitative study of the Syrian refugee experience accessing noncommunicable disease services in Jordan. Confl. Health 2019, 13, 26. [Google Scholar] [CrossRef] [Green Version]
- International Rescue Committee. Public Health access and Health Seeking Behaviors of Syrian Refugees in Jordan. Available online: https://data2.unhcr.org/en/documents/details/72211 (accessed on 16 October 2020).
- Doocy, S.; Lyles, E.; Roberton, T.; Akhu-Zaheya, L.; Oweis, A.; Burnham, G. Prevalence and care-seeking for chronic diseases among Syrian refugees in Jordan. BMC Public Health 2015, 15, 1097. [Google Scholar] [CrossRef] [Green Version]
- Spiegel, P.; Khalifa, A.; Mateen, F.J. Cancer in refugees in Jordan and Syria between 2009 and 2012: Challenges and the way forward in humanitarian emergencies. Lancet Oncol. 2014, 15, e290–e297. [Google Scholar] [CrossRef]
- Murshidi, M.M.; Hijjawi, M.Q.B.; Jeriesat, S.; Eltom, A. Syrian refugees and Jordan’s health sector. Lancet 2013, 382, 206–207. [Google Scholar] [CrossRef]
- Farag, N.H.; Wannemuehler, K.; Weldon, W.; Arbaji, A.; Belbaisi, A.; Khuri-Bulos, N.; Ehrhardt, D.; Surour, M.R.; ElhajQasem, N.S.; Al-Abdallat, M.M. Estimating population immunity to poliovirus in Jordan’s high-risk areas. Hum. Vaccines Immunother. 2019, 16, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Lam, E.; Diaz, M.; Maina, A.G.K.; Brennan, M. Displaced populations due to humanitarian emergencies and its impact on global eradication and elimination of vaccine-preventable diseases. Confl. Health 2016, 10, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimer, N.A. A Review on Emerging and Reemerging of Infectious Diseases in Jordan: The Aftermath of the Syrian Crises. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 8679174. [Google Scholar] [CrossRef] [Green Version]
- Gatrad, A.R.; Sheikh, A. Erratum: Hajj: Journey of a lifetime (British Medical Journal (2005) 330 (133–137)). Br. Med. J. 2005, 331, 442. [Google Scholar]
- Memish, Z.A.; Stephens, G.M.; Steffen, R.; Ahmed, Q.A. Emergence of medicine for mass gatherings: Lessons from the Hajj. Lancet Infect. Dis. 2012, 12, 56–65. [Google Scholar] [CrossRef]
- Yezli, S.; Yassin, Y.M.; Awam, A.H.; Attar, A.A.; Al-Jahdali, E.A.; Alotaibi, B.M. Umrah. An opportunity for mass gatherings health research. Saudi Med. J. 2017, 38, 868–871. [Google Scholar] [CrossRef]
- Ahmed, Q.A.; Arabi, Y.M.; Memish, Z.A. Health risks at the Hajj. Lancet 2006, 367, 1008–1015. [Google Scholar] [CrossRef]
- Abdelmoety, D.A.; El-Bakri, N.K.; Almowalld, W.O.; Turkistani, Z.A.; Bugis, B.H.; Baseif, E.A.; Melbari, M.H.; AlHarbi, K.; Abu-Shaheen, A. Characteristics of Heat Illness during Hajj: A Cross-Sectional Study. BioMed Res. Int. 2018, 2018, 5629474. [Google Scholar] [CrossRef]
- Memish, Z.A. Meningococcal Disease and Travel. Clin. Infect. Dis. 2002, 34, 84–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautret, P.; Benkouiten, S.; Sridhar, S.; Al-Tawfiq, J.A.; Memish, Z.A. Diarrhea at the Hajj and Umrah. Travel Med. Infect. Dis. 2015, 13, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Abd El Ghany, M.; Alsomali, M.; Almasri, M.; Padron Regalado, E.; Naeem, R.; Tukestani, A.H.; Asiri, A.; Hill-Cawthorne, G.A.; Pain, A.; Memish, Z.A. Enteric infections circulating during Hajj seasons, 2011–2013. Emerg. Infect. Dis. 2017, 23, 1640–1649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olaitan, A.O.; Dia, N.M.; Gautret, P.; Benkouiten, S.; Belhouchat, K.; Drali, T.; Parola, P.; Brouqui, P.; Memish, Z.; Raoult, D.; et al. Acquisition of extended-spectrum cephalosporin- and colistin-resistant Salmonella enterica subsp. enterica serotype Newport by pilgrims during Hajj. Int. J. Antimicrob. Agents 2015, 45, 600–604. [Google Scholar] [CrossRef]
- Leangapichart, T.; Gautret, P.; Griffiths, K.; Belhouchat, K.; Memish, Z.; Raoult, D.; Rolain, J.M. Acquisition of a high diversity of bacteria during the Hajj pilgrimage, including Acinetobacter baumannii with blaOXA-72 and Escherichia coli with blaNDM-5 carbapenemase genes. Antimicrob. Agents Chemother. 2016, 60, 5942–5948. [Google Scholar] [CrossRef] [Green Version]
- Fenton, M.B.; Simmons, N.B. Bats: A World of Science and Mystery, Fenton, Simmons; The University of Chicago Press: Chicago, IL, USA, 2014. [Google Scholar]
- O’Shea, T.J.; Cryan, P.M.; Cunningham, A.A.; Fooks, A.R.; Hayman, D.T.S.; Luis, A.D.; Peel, A.J.; Plowright, R.K.; Wood, J.L.N. Bat flight and zoonotic viruses. Emerg. Infect. Dis. 2014, 20, 741–745. [Google Scholar] [CrossRef] [Green Version]
- Kunz, T.H.; Braun De Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef]
- Olival, K.; Hosseini, P.; Zambrana-Torrelio, C.; Nature, N.R.-; 2017, U. Host and viral traits predict zoonotic spillover from mammals. Nature 2017, 546, 646–650. [Google Scholar] [CrossRef]
- Menachery, V.D.; Yount, B.L.; Debbink, K.; Agnihothram, S.; Gralinski, L.E.; Plante, J.A.; Graham, R.L.; Scobey, T.; Ge, X.Y.; Donaldson, E.F.; et al. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat. Med. 2015, 21, 1508–1513. [Google Scholar] [CrossRef]
- York, A. On the bat’s back I do fly. Nat. Rev. Microbiol. 2019, 17, 265. [Google Scholar] [CrossRef]
- Banerjee, A.; Baker, M.L.; Kulcsar, K.; Misra, V.; Plowright, R.; Mossman, K. Novel Insights Into Immune Systems of Bats. Front. Immunol. 2020, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Phelps, K.L.; Hamel, L.; Alhmoud, N.; Ali, S.; Bilgin, R.; Sidamonidze, K.; Urushadze, L.; Karesh, W.; Olival, K.J. Bat research networks and viral surveillance: Gaps and opportunities in western Asia. Viruses 2019, 11, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelli, D.; Moreno, A.; Lavazza, A.; Bresaola, M.; Canelli, E.; Boniotti, M.B.; Cordioli, P. Identification of Mammalian Orthoreovirus Type 3 in Italian Bats. Zoonoses Public Health 2013, 60, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Wellenberg, G.J.; Audry, L.; Rønsholt, L.; Van der Poel, W.H.M.; Bruschke, C.J.M.; Bourhy, H. Presence of European bat lyssavirus RNAs in apparently healthy Rousettus aegyptiacus bats. Arch. Virol. 2002, 147, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Amr, Z.S.; Abu Baker, M.A.; Botros Qumsiyeh, M. Bat Diversity and Conservation in Jordan. Turkish J. Zool. 2006, 30, 235–244. [Google Scholar]
- Eggerbauer, E.; Pfaff, F.; Finke, S.; Höper, D.; Beer, M.; Mettenleiter, T.C.; Nolden, T.; Teifke, J.-P.; Müller, T.; Freuling, C.M. Comparative analysis of European bat lyssavirus 1 pathogenicity in the mouse model. PLoS Negl. Trop. Dis. 2017, 11, e0005668. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, S.; Ibáñez, C.; Juste, J.; Echevarria, J.E. EBLV1 circulation in natural bat colonies of Eptesicus serotinus: A six year survey. Dev. Biol. (Basel) 2006, 125, 257–261. [Google Scholar]
- Papadatou, E.; Butlin, R.K.; Altringham, J.D. Seasonal Roosting Habits and Population Structure of the Long-fingered Bat Myotis capaccinii in Greece. J. Mammal. 2008, 89, 503–512. [Google Scholar] [CrossRef] [Green Version]
- Pons-Salort, M.; Serra-Cobo, J.; Jay, F.; López-Roig, M.; Lavenir, R.; Guillemot, D.; Letort, V.; Bourhy, H.; Opatowski, L. Insights into Persistence Mechanisms of a Zoonotic Virus in Bat Colonies Using a Multispecies Metapopulation Model. PLoS ONE 2014, 9, e95610. [Google Scholar] [CrossRef] [Green Version]
- NADER, I.A. On the bats (Chiroptera) of the kingdom of Saudi Arabia. J. Zool. 1975, 176, 331–340. [Google Scholar] [CrossRef]
- Memish, Z.A.; Mishra, N.; Olival, K.J.; Fagbo, S.F.; Kapoor, V.; Epstein, J.H.; AlHakeem, R.; Al Asmari, M.; Islam, A.; Kapoor, A.; et al. Middle East respiratory syndrome coronavirus in Bats, Saudi Arabia. Emerg. Infect. Dis. 2013, 19, 1819–1823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcês, A.; Correia, S.; Amorim, F.; Pereira, J.E.; Igrejas, G.; Poeta, P. First report on extended-spectrum beta-lactamase (ESBL) producing Escherichia coli from European free-tailed bats (Tadarida teniotis) in Portugal: A one-health approach of a hidden contamination problem. J. Hazard. Mater. 2019, 370, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Markotter, W.; Monadjem, A.; Nel, L.H. Antibodies against Duvenhage virus in insectivorous bats in Swaziland. J. Wildl. Dis. 2013, 49, 1000–1003. [Google Scholar] [CrossRef] [Green Version]
- Jánoska, M.; Vidovszky, M.; Molnár, V.; Liptovszky, M.; Harrach, B.; Benko, M. Novel adenoviruses and herpesviruses detected in bats. Vet. J. 2011, 189, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Amman, B.R.; Bird, B.H.; Bakarr, I.A.; Bangura, J.; Schuh, A.J.; Johnny, J.; Sealy, T.K.; Conteh, I.; Koroma, A.H.; Foday, I.; et al. Isolation of Angola-like Marburg virus from Egyptian rousette bats from West Africa. Nat. Commun. 2020, 11, 510. [Google Scholar] [CrossRef]
- Amman, B.R.; Carroll, S.A.; Reed, Z.D.; Sealy, T.K.; Balinandi, S.; Swanepoel, R.; Kemp, A.; Erickson, B.R.; Comer, J.A.; Campbell, S.; et al. Seasonal Pulses of Marburg Virus Circulation in Juvenile Rousettus aegyptiacus Bats Coincide with Periods of Increased Risk of Human Infection. PLoS Pathog. 2012, 8, e1002877. [Google Scholar] [CrossRef]
- Drexler, J.F.; Gloza-Rausch, F.; Glende, J.; Corman, V.M.; Muth, D.; Goettsche, M.; Seebens, A.; Niedrig, M.; Pfefferle, S.; Yordanov, S.; et al. Genomic Characterization of Severe Acute Respiratory Syndrome-Related Coronavirus in European Bats and Classification of Coronaviruses Based on Partial RNA-Dependent RNA Polymerase Gene Sequences. J. Virol. 2010, 84, 11336–11349. [Google Scholar] [CrossRef] [Green Version]
- Markotter, W.; Geldenhuys, M.; Jansen van Vuren, P.; Kemp, A.; Mortlock, M.; Mudakikwa, A.; Nel, L.; Nziza, J.; Paweska, J.; Weyer, J. Paramyxo- and Coronaviruses in Rwandan Bats. Trop. Med. Infect. Dis. 2019, 4, 99. [Google Scholar] [CrossRef] [Green Version]
- Ar Gouilh, M.; Puechmaille, S.J.; Diancourt, L.; Vandenbogaert, M.; Serra-Cobo, J.; Lopez Roïg, M.; Brown, P.; Moutou, F.; Caro, V.; Vabret, A.; et al. SARS-CoV related Betacoronavirus and diverse Alphacoronavirus members found in western old-world. Virology 2018, 517, 88–97. [Google Scholar] [CrossRef]
- Rizzo, F.; Edenborough, K.M.; Toffoli, R.; Culasso, P.; Zoppi, S.; Dondo, A.; Robetto, S.; Rosati, S.; Lander, A.; Kurth, A.; et al. Coronavirus and paramyxovirus in bats from Northwest Italy. BMC Vet. Res. 2017, 13, 396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra-Cobo, J.; Amengual, B.; Carlos Abellán, B.; Bourhy, H. European bat Lyssavirus infection in Spanish bat populations. Emerg. Infect. Dis. 2002, 8, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, K.; Kuwata, R.; Shimoda, H.; Mizutani, T.; Hondo, E.; Maeda, K. The complete genomic sequence of Rhinolophus gammaherpesvirus 1 isolated from a greater horseshoe bat. Arch. Virol. 2019, 164, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Pauly, M.; Pir, J.B.; Loesch, C.; Sausy, A.; Snoeck, C.J.; Hübschen, J.M.; Muller, C.P. Novel alphacoronaviruses and paramyxoviruses cocirculate with type 1 and severe acute respiratory system (SARS)-related betacoronaviruses in synanthropic bats of Luxembourg. Appl. Environ. Microbiol. 2017, 83, e01326-17. [Google Scholar] [CrossRef] [Green Version]
- Rihtarič, D.; Hostnik, P.; Steyer, A.; Grom, J.; Toplak, I. Identification of SARS-like coronaviruses in horseshoe bats (Rhinolophus hipposideros) in Slovenia. Arch. Virol. 2010, 155, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Fagbo, S.F.; Alagaili, A.N.; Nitido, A.; Williams, S.H.; Ng, J.; Lee, B.; Durosinlorun, A.; Garcia, J.A.; Jain, K.; et al. A viral metagenomic survey identifies known and novel mammalian viruses in bats from Saudi Arabia. PLoS ONE 2019, 14, e0214227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dufkova, L.; Straková, P.; Širmarová, J.; Salát, J.; Moutelíková, R.; Chrudimský, T.; Bartonička, T.; Nowotny, N.; Růžek, D. Detection of Diverse Novel Bat Astrovirus Sequences in the Czech Republic. Vector-Borne Zoonotic Dis. 2015, 15, 518–521. [Google Scholar] [CrossRef] [PubMed]
- De Benedictis, P.; Marciano, S.; Scaravelli, D.; Priori, P.; Zecchin, B.; Capua, I.; Monne, I.; Cattoli, G. Alpha and lineage C betaCoV infections in Italian bats. Virus Genes 2014, 48, 366–371. [Google Scholar] [CrossRef]
- Smith, G.C.; Aegerter, J.N.; Allnutt, T.R.; MacNicoll, A.D.; Learmount, J.; Hutson, A.M.; Atterby, H. Bat population genetics and Lyssavirus presence in Great Britain. Epidemiol. Infect. 2011, 139, 1463–1469. [Google Scholar] [CrossRef] [Green Version]
- Robardet, E.; Borel, C.; Moinet, M.; Jouan, D.; Wasniewski, M.; Barrat, J.; Boué, F.; Montchâtre-Leroy, E.; Servat, A.; Gimenez, O.; et al. Longitudinal survey of two serotine bat (Eptesicus serotinus) maternity colonies exposed to EBLV-1 (European Bat Lyssavirus type 1): Assessment of survival and serological status variations using capture-recapture models. PLoS Negl. Trop. Dis. 2017, 11, e0006048. [Google Scholar] [CrossRef] [Green Version]
- Picard-Meyer, E.; Dubourg-Savage, M.J.; Arthur, L.; Barataud, M.; Bécu, D.; Bracco, S.; Borel, C.; Larcher, G.; Meme-Lafond, B.; Moinet, M.; et al. Active surveillance of bat rabies in France: A 5-year study (2004–2009). Vet. Microbiol. 2011, 151, 390–395. [Google Scholar] [CrossRef] [Green Version]
- Molnár, V.; Jánoska, M.; Harrach, B.; Glávits, R.; Pálmai, N.; Rigó, D.; Sós, E.; Liptovszky, M. Detection of a novel bat gammaherpesvirus in Hungary. Acta Vet. Hung. 2008, 56, 529–538. [Google Scholar] [CrossRef]
- Ceballos, N.A.; Morón, S.V.; Berciano, J.M.; Nicolás, O.; López, C.A.; Juste, J.; Nevado, C.R.; Setién, Á.A.; Echevarría, J.E. Novel lyssavirus in bat, Spain. Emerg. Infect. Dis. 2013, 19, 793–795. [Google Scholar] [CrossRef] [Green Version]
- Picard-Meyer, E.; Beven, V.; Hirchaud, E.; Guillaume, C.; Larcher, G.; Robardet, E.; Servat, A.; Blanchard, Y.; Cliquet, F. Lleida Bat Lyssavirus isolation in Miniopterus schreibersii in France. Zoonoses Public Health 2019, 66, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Kemenesi, G.; Kurucz, K.; Dallos, B.; Zana, B.; Földes, F.; Boldogh, S.; Görföl, T.; Carroll, M.W.; Jakab, F. Re-emergence of Lloviu virus in Miniopterus schreibersii bats, Hungary, 2016. Emerg. Microbes Infect. 2018, 7, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzmin, I.V.; Niezgoda, M.; Franka, R.; Agwanda, B.; Markotter, W.; Beagley, J.C.; Urazova, O.Y.; Breiman, R.F.; Rupprecht, C.E. Possible emergence of West Caucasian bat virus in Africa. Emerg. Infect. Dis. 2008, 14, 1887–1889. [Google Scholar] [CrossRef] [PubMed]
- Picard-Meyer, E.; Servat, A.; Robardet, E.; Moinet, M.; Borel, C.; Cliquet, F. Isolation of Bokeloh bat lyssavirus in Myotis nattereri in France. Arch. Virol. 2013, 158, 2333–2340. [Google Scholar] [CrossRef]
- Freuling, C.M.; Beer, M.; Conraths, F.J.; Finke, S.; Hoffmann, B.; Keller, B.; Kliemt, J.; Mettenleiter, T.C.; Mühlbach, E.; Teifke, J.P.; et al. Novel lyssavirus in Natterer’s bat, Germany. Emerg. Infect. Dis. 2011, 17, 1519–1522. [Google Scholar] [CrossRef]
- Wibbelt, G.; Kurth, A.; Yasmum, N.; Bannert, M.; Nagel, S.; Nitsche, A.; Ehlers, B. Discovery of herpesviruses in bats. J. Gen. Virol. 2007, 88, 2651–2655. [Google Scholar] [CrossRef]
- Lelli, D.; Papetti, A.; Sabelli, C.; Rosti, E.; Moreno, A.; Boniotti, M.B. Detection of coronaviruses in bats of various species in Italy. Viruses 2013, 5, 2679–2689. [Google Scholar] [CrossRef] [Green Version]
- Straková, P.; Dufkova, L.; Širmarová, J.; Salát, J.; Bartonička, T.; Klempa, B.; Pfaff, F.; Höper, D.; Hoffmann, B.; Ulrich, R.G.; et al. Novel hantavirus identified in European bat species Nyctalus noctula. Infect. Genet. Evol. 2017, 48, 127–130. [Google Scholar] [CrossRef]
- Moreno, A.; Lelli, D.; De Sabato, L.; Zaccaria, G.; Boni, A.; Sozzi, E.; Prosperi, A.; Lavazza, A.; Cella, E.; Castrucci, M.R.; et al. Detection and full genome characterization of two beta CoV viruses related to Middle East respiratory syndrome from bats in Italy. Virol. J. 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Lelli, D.; Prosperi, A.; Moreno, A.; Chiapponi, C.; Gibellini, A.M.; De Benedictis, P.; Leopardi, S.; Sozzi, E.; Lavazza, A. Isolation of a novel Rhabdovirus from an insectivorous bat (Pipistrellus kuhlii) in Italy. Virol. J. 2018, 15, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cusi, M. Toscana Virus Epidemiology: From Italy to Beyond. Open Virol. J. 2010, 4, 22–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schatz, J.; Fooks, A.R.; Mcelhinney, L.; Horton, D.; Echevarria, J.; Vázquez-Moron, S.; Kooi, E.A.; Rasmussen, T.B.; Müller, T.; Freuling, C.M. Bat Rabies Surveillance in Europe. Zoonoses Public Health 2013, 60, 22–34. [Google Scholar] [CrossRef]
- Al-Malabeh, A.; Kempe, S.; Henschel, H.V.; Hofmann, H.; Tobschall, H.J. The possibly hypogene karstic iron ore deposit of Warda near Ajloun (Northern Jordan), its mineralogy, geochemistry and historic mine. Acta Carsol. 2008, 37, 241–253. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, K.; Peiris, J.S.M. Coronaviruses. In Clinical Virology, 3rd ed.; Richman, D., Whitley, R., Hayden, F., Eds.; ASM Press: Washington, DC, USA, 2009; pp. 1155–1171. [Google Scholar]
- AL-Eitan, L.N.; Alahmad, S.Z. Pharmacogenomics of genetic polymorphism within the genes responsible for SARS-CoV-2 susceptibility and the drug-metabolising genes used in treatment. Rev. Med. Virol. 2020, e2194. [Google Scholar] [CrossRef]
- Graham, R.L.; Donaldson, E.F.; Baric, R.S. A decade after SARS: Strategies for controlling emerging coronaviruses. Nat. Rev. Microbiol. 2013, 11, 836–848. [Google Scholar] [CrossRef] [Green Version]
- Van Der Hoek, L. Human coronaviruses: What do they cause? Antivir. Ther. 2007, 12, 651–658. [Google Scholar]
- Paden, C.R.; Yusof, M.F.B.M.; Al Hammadi, Z.M.; Queen, K.; Tao, Y.; Eltahir, Y.M.; Elsayed, E.A.; Marzoug, B.A.; Bensalah, O.K.A.; Khalafalla, A.I.; et al. Zoonotic origin and transmission of Middle East respiratory syndrome coronavirus in the UAE. Zoonoses Public Health 2018, 65, 322–333. [Google Scholar] [CrossRef]
- Bolles, M.; Donaldson, E.; Baric, R. SARS-CoV and emergent coronaviruses: Viral determinants of interspecies transmission. Curr. Opin. Virol. 2011, 1, 624–634. [Google Scholar] [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony, S.J.; Ojeda-Flores, R.; Rico-Chávez, O.; Navarrete-Macias, I.; Zambrana-Torrelio, C.M.; Rostal, M.K.; Epstein, J.H.; Tipps, T.; Liang, E.; Sanchez-Leon, M.; et al. Coronaviruses in bats from Mexico. J. Gen. Virol. 2013, 94, 1028–1038. [Google Scholar] [CrossRef]
- Ithete, N.L.; Stoffberg, S.; Corman, V.M.; Cottontail, V.M.; Richards, L.R.; Schoeman, M.C.; Drosten, C.; Drexler, J.F.; Preiser, W. Close relative of human middle east respiratory syndrome coronavirus in bat, South Africa. Emerg. Infect. Dis. 2013, 19, 1697–1699. [Google Scholar] [CrossRef] [PubMed]
- Al-Osail, A.M.; Al-Wazzah, M.J. The history and epidemiology of Middle East respiratory syndrome corona virus. Multidiscip. Respir. Med. 2017, 12, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H.; et al. Bats are natural reservoirs of SARS-like coronaviruses. Science (80-. ) 2005, 310, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Menachery, V.D.; Yount, B.L.; Sims, A.C.; Debbink, K.; Agnihothram, S.S.; Gralinski, L.E.; Graham, R.L.; Scobey, T.; Plante, J.A.; Royal, S.R.; et al. SARS-like WIV1-CoV poised for human emergence. Proc. Natl. Acad. Sci. USA 2016, 113, 3048–3053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, N.; Li, S.Y.; Yang, X.L.; Huang, H.M.; Zhang, Y.J.; Guo, H.; Luo, C.M.; Miller, M.; Zhu, G.; Chmura, A.A.; et al. Serological Evidence of Bat SARS-Related Coronavirus Infection in Humans, China. Virol. Sin. 2018, 33, 104–107. [Google Scholar] [CrossRef] [Green Version]
- CDC MERS Symptoms & Complications|CDC. Available online: https://www.cdc.gov/coronavirus/mers/about/symptoms.html (accessed on 27 February 2020).
- Dong, E.; Du, H.; Gardner, L. An interactive web-based dashboard to track COVID-19 in real time. Lancet Infect. Dis. 2020, 20, 533–534. [Google Scholar] [CrossRef]
- Hu, T.; Liu, Y.; Zhao, M.; Zhuang, Q.; Xu, L.; He, Q. A comparison of COVID-19, SARS and MERS. PeerJ 2020, 8, e9725. [Google Scholar] [CrossRef]
- Killerby, M.E.; Biggs, H.M.; Midgley, C.M.; Gerber, S.I.; Watson, J.T. Middle east respiratory syndrome coronavirus transmission. Emerg. Infect. Dis. 2020, 26, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Katul, G.G.; Mrad, A.; Bonetti, S.; Manoli, G.; Parolari, A.J. Global convergence of COVID-19 basic reproduction number and estimation from early-time SIR dynamics. PLoS ONE 2020, 15, e0239800. [Google Scholar] [CrossRef] [PubMed]
- Arabi, Y.M.; Arifi, A.A.; Balkhy, H.H.; Najm, H.; Aldawood, A.S.; Ghabashi, A.; Hawa, H.; Alothman, A.; Khaldi, A.; Al Raiy, B. Clinical course and outcomes of critically ill patients with middle east respiratory syndrome coronavirus infection. Ann. Intern. Med. 2014, 160, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Mohd, H.A.; Al-Tawfiq, J.A.; Memish, Z.A. Middle East Respiratory Syndrome Coronavirus (MERS-CoV) origin and animal reservoir. Virol. J. 2016, 13, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anthony, S.J.; Gilardi, K.; Menachery, V.D.; Goldstein, T.; Ssebide, B.; Mbabazi, R.; Navarrete-Macias, I.; Liang, E.; Wells, H.; Hicks, A.; et al. Further evidence for bats as the evolutionary source of middle east respiratory syndrome coronavirus. MBio 2017, 8, e00373-17. [Google Scholar] [CrossRef] [Green Version]
- Zumla, A.; Hui, D.S.; Perlman, S. Middle East respiratory syndrome. Lancet 2015, 386, 995–1007. [Google Scholar] [CrossRef] [Green Version]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef]
- Munster, V.J.; Adney, D.R.; Van Doremalen, N.; Brown, V.R.; Miazgowicz, K.L.; Milne-Price, S.; Bushmaker, T.; Rosenke, R.; Scott, D.; Hawkinson, A.; et al. Replication and shedding of MERS-CoV in Jamaican fruit bats (Artibeus jamaicensis). Sci. Rep. 2016, 6, 21878. [Google Scholar] [CrossRef]
- Eifan, S.A.; Nour, I.; Hanif, A.; Zamzam, A.M.M.; AlJohani, S.M. A pandemic risk assessment of middle east respiratory syndrome coronavirus (MERS-CoV) in Saudi Arabia. Saudi J. Biol. Sci. 2017, 24, 1631–1638. [Google Scholar] [CrossRef]
- Chu, D.K.W.; Poon, L.L.M.; Gomaa, M.M.; Shehata, M.M.; Perera, R.A.P.M.; Zeid, D.A.; El Rifay, A.S.; Siu, L.Y.; Guan, Y.; Webby, R.J.; et al. MERS coronaviruses in dromedary camels, Egypt. Emerg. Infect. Dis. 2014, 20, 1049–1053. [Google Scholar] [CrossRef] [Green Version]
- Reusken, C.B.; Ababneh, M.; Raj, V.S.; Meyer, B.; Eljarah, A.; Abutarbush, S.; Godeke, G.J.; Bestebroer, T.M.; Zutt, I.; Müller, M.A.; et al. Middle east respiratory syndrome coronavirus (MERS-CoV) serology in major livestock species in an affected region in Jordan, June to September 2013. Eurosurveillance 2013, 18, 20662. [Google Scholar] [CrossRef]
- Reusken, C.B.E.M.; Haagmans, B.L.; Müller, M.A.; Gutierrez, C.; Godeke, G.J.; Meyer, B.; Muth, D.; Raj, V.S.; De Vries, L.S.; Corman, V.M.; et al. Middle East respiratory syndrome coronavirus neutralising serum antibodies in dromedary camels: A comparative serological study. Lancet Infect. Dis. 2013, 13, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Perera, R.A.; Wang, P.; Gomaa, M.R.; El-Shesheny, R.; Kandeil, A.; Bagato, O.; Siu, L.Y.; Shehata, M.M.; Kayed, A.S.; Moatasim, Y.; et al. Seroepidemiology for MERS coronavirus using microneutralisation and pseudoparticle virus neutralisation assays reveal a high prevalence of antibody in dromedary camels in Egypt, June 2013. Eurosurveillance 2013, 18, 20574. [Google Scholar] [CrossRef] [PubMed]
- Hemida, M.; Perera, R.; Wang, P.; Alhammadi, M.; Siu, L.; Li, M.; Poon, L.; Saif, L.; Alnaeem, A.; Peiris, M. Middle East Respiratory Syndrome (MERS) coronavirus seroprevalence in domestic livestock in Saudi Arabia, 2010 to 2013. Eurosurveillance 2013, 18, 20659. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Müller, M.A.; Corman, V.M.; Reusken, C.B.E.M.; Ritz, D.; Godeke, G.J.; Lattwein, E.; Kallies, S.; Siemens, A.; van Beek, J.; et al. Antibodies against MERS coronavirus in dromedaries, United Arab Emirates, 2003 and 2013. Emerg. Infect. Dis. 2014, 20, 552–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, M.A.; Meyer, B.; Corman, V.M.; Al-Masri, M.; Turkestani, A.; Ritz, D.; Sieberg, A.; Aldabbagh, S.; Bosch, B.J.; Lattwein, E.; et al. Presence of Middle East respiratory syndrome coronavirus antibodies in Saudi Arabia: A nationwide, cross-sectional, serological study. Lancet Infect. Dis. 2015, 15, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Hunter, J.C.; Nguyen, D.; Aden, B.; Al Bandar, Z.; Al Dhaheri, W.; Abu Elkheir, K.; Khudair, A.; Al Mulla, M.; El Saleh, F.; Imambaccus, H.; et al. Transmission of middle east respiratory syndrome coronavirus infections in healthcare settings, abu dhabi. Emerg. Infect. Dis. 2016, 22, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Sharif-Yakan, A.; Kanj, S.S. Emergence of MERS-CoV in the Middle East: Origins, Transmission, Treatment, and Perspectives. PLoS Pathog. 2014, 10, e1004457. [Google Scholar] [CrossRef] [Green Version]
- Al-Jasser, F.S.; Nouh, R.M.; Youssef, R.M. Epidemiology and predictors of survival of MERS-CoV infections in Riyadh region, 2014–2015. J. Infect. Public Health 2019, 12, 171–177. [Google Scholar] [CrossRef]
- Al-Abdallat, M.M.; Payne, D.C.; Alqasrawi, S.; Rha, B.; Tohme, R.A.; Abedi, G.R.; Nsour, M.A.; Iblan, I.; Jarour, N.; Farag, N.H.; et al. Hospital-associated outbreak of middle east respiratory syndrome coronavirus: A serologic, epidemiologic, and clinical description. Clin. Infect. Dis. 2014, 59, 1225–1233. [Google Scholar] [CrossRef] [Green Version]
- Khuri-Bulos, N.; Payne, D.C.; Lu, X.; Erdman, D.; Wang, L.; Faouri, S.; Shehabi, A.; Johnson, M.; Becker, M.M.; Denison, M.R.; et al. Middle East respiratory syndrome coronavirus not detected in children hospitalized with acute respiratory illness in Amman, Jordan, March 2010 to September 2012. Clin. Microbiol. Infect. 2014, 20, 678–682. [Google Scholar] [CrossRef] [Green Version]
- van Doremalen, N.; Hijazeen, Z.S.K.; Holloway, P.; Al Omari, B.; McDowell, C.; Adney, D.; Talafha, H.A.; Guitian, J.; Steel, J.; Amarin, N.; et al. High Prevalence of Middle East Respiratory Coronavirus in Young Dromedary Camels in Jordan. Vector-Borne Zoonotic Dis. 2017, 17, 155–159. [Google Scholar] [CrossRef] [Green Version]
- Payne, D.C.; Biggs, H.M.; Al-Abdallat, M.M.; Alqasrawi, S.; Lu, X.; Abedi, G.R.; Haddadin, A.; Iblan, I.; Alsanouri, T.; Nsour, M.A.; et al. Multihospital outbreak of a Middle East respiratory syndrome coronavirus deletion variant, Jordan: A molecular, serologic, and epidemiologic investigation. Open Forum Infect. Dis. 2018, 5, ofy095. [Google Scholar] [CrossRef] [Green Version]
- Lamers, M.M.; Raj, S.V.; Shafei, M.; Ali, S.S.; Abdallh, S.M.; Gazo, M.; Nofal, S.; Lu, X.; Erdman, D.D.; Koopmans, M.P.; et al. Deletion variants of middle east respiratory syndrome coronavirus from humans, Jordan, 2015. Emerg. Infect. Dis. 2016, 22, 716–719. [Google Scholar] [CrossRef]
- World Health Organization. Middle East Respiratory Syndrome Coronavirus (MERS-CoV). Available online: https://www.who.int/health-topics/middle-east-respiratory-syndrome-coronavirus-mers#tab=tab_1 (accessed on 17 February 2020).
- Al-Tawfiq, J.A.; Zumla, A.; Memish, Z.A. Respiratory tract infections during the annual Hajj. Curr. Opin. Pulm. Med. 2013, 19, 192–197. [Google Scholar] [CrossRef]
- Gautret, P.; Benkouiten, S.; Al-Tawfiq, J.A.; Memish, Z.A. Hajj-associated viral respiratory infections: A systematic review. Travel Med. Infect. Dis. 2016, 14, 92–109. [Google Scholar] [CrossRef]
- Memish, Z.A.; Almasri, M.; Turkestani, A.; Al-Shangiti, A.M.; Yezli, S. Etiology of severe community-acquired pneumonia during the 2013 Hajj-part of the MERS-CoV surveillance program. Int. J. Infect. Dis. 2014, 25, 186–190. [Google Scholar] [CrossRef] [Green Version]
- Alfaraj, S.H.; Al-Tawfiq, J.A.; Alzahrani, N.A.; Altwaijri, T.A.; Memish, Z.A. The impact of co-infection of influenza A virus on the severity of Middle East Respiratory Syndrome Coronavirus. J. Infect. 2017, 74, 521–523. [Google Scholar] [CrossRef] [Green Version]
- Memish, Z.A.; Zumla, A.; Alhakeem, R.F.; Assiri, A.; Turkestani, A.; Al Harby, K.D.; Alyemni, M.; Dhafar, K.; Gautret, P.; Barbeschi, M.; et al. Hajj: Infectious disease surveillance and control. Lancet 2014, 383, 2073–2082. [Google Scholar] [CrossRef]
- Premila Devi, J.; Noraini, W.; Norhayati, R.; Chee Kheong, C.; Badrul, A.S.; Zainah, S.; Fadzilah, K.; Hirman, I.; Lokman Hakim, S.; Noor Hisham, A. Laboratory-confirmed case of Middle East respiratory syndrome coronavirus (MERS-CoV) infection in Malaysia: Preparedness and response, April 2014. Eurosurveillance 2014, 19, 20797. [Google Scholar] [CrossRef]
- Kumar, A.; Beckett, G.; Wiselka, M. Middle East respiratory syndrome coronavirus (MERS-CoV) in pilgrims returning from the Hajj. BMJ 2015, 351, h5185. [Google Scholar] [CrossRef]
- Gautret, P.; Charrel, R.; Benkouiten, S.; Belhouchat, K.; Nougairede, A.; Drali, T.; Salez, N.; Memish, Z.A.; al Masri, M.; Lagier, J.C.; et al. Lack of MERS coronavirus but prevalence of influenza virus in French pilgrims after 2013 Hajj. Emerg. Infect. Dis. 2014, 20, 728–730. [Google Scholar] [CrossRef]
- Benkouiten, S.; Charrel, R.; Belhouchat, K.; Drali, T.; Salez, N.; Nougairede, A.; Zandotti, C.; Memish, Z.A.; al Masri, M.; Gaillard, C.; et al. Circulation of Respiratory Viruses Among Pilgrims During the 2012 Hajj Pilgrimage. Clin. Infect. Dis. 2013, 57, 992–1000. [Google Scholar] [CrossRef] [Green Version]
- Gautret, P.; Charrel, R.; Belhouchat, K.; Drali, T.; Benkouiten, S.; Nougairede, A.; Zandotti, C.; Memish, Z.A.; al Masri, M.; Gaillard, C.; et al. Lack of nasal carriage of novel corona virus (HCoV-EMC) in French Hajj pilgrims returning from the Hajj 2012, despite a high rate of respiratory symptoms. Clin. Microbiol. Infect. 2013, 19, E315–E317. [Google Scholar] [CrossRef] [Green Version]
- Rashid, H.; Barasheed, O.; Booy, R. Acute febrile respiratory infection symptoms in Australian Hajjis at risk of exposure to middle east respiratory syndrome coronavirus. Med. J. Aust. 2013, 199, 453. [Google Scholar] [CrossRef]
- Koul, P.A.; Mir, H.; Saha, S.; Chadha, M.S.; Potdar, V.; Widdowson, M.A.; Lal, R.B.; Krishnan, A. Influenza not MERS CoV among returning Hajj and Umrah pilgrims with respiratory illness, Kashmir, north India, 2014–2015. Travel Med. Infect. Dis. 2017, 15, 45–47. [Google Scholar] [CrossRef]
- Refaey, S.; Amin, M.M.; Roguski, K.; Azziz-Baumgartner, E.; Uyeki, T.M.; Labib, M.; Kandeel, A. Cross-sectional survey and surveillance for influenza viruses and MERS-CoV among Egyptian pilgrims returning from Hajj during 2012–2015. Influenza Other Respi. Viruses 2017, 11, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Liu, F.; Liu, L.; Zhang, L.; Lu, M.; Abudukadeer, A.; Wang, L.; Tian, F.; Zhen, W.; Yang, P.; et al. No MERS-CoV but positive influenza viruses in returning Hajj pilgrims, China, 2013–2015. BMC Infect. Dis. 2017, 17, 715. [Google Scholar] [CrossRef] [Green Version]
- Annan, A.; Owusu, M.; Marfo, K.S.; Larbi, R.; Sarpong, F.N.; Adu-Sarkodie, Y.; Amankwa, J.; Fiafemetsi, S.; Drosten, C.; Owusu-Dabo, E.; et al. High prevalence of common respiratory viruses and no evidence of Middle East Respiratory Syndrome Coronavirus in Hajj pilgrims returning to Ghana, 2013. Trop. Med. Int. Heal. 2015, 20, 807–812. [Google Scholar] [CrossRef] [Green Version]
- Amin, M.; Bakhtiar, A.; Subarjo, M.; Aksono, E.B.; Widiyanti, P.; Shimizu, K.; Mori, Y. Screening for Middle East respiratory syndrome coronavirus among febrile Indonesian Hajj pilgrims: A study on 28,197 returning pilgrims. J. Infect. Prev. 2018, 19, 236–239. [Google Scholar] [CrossRef] [Green Version]
- Yavarian, J.; Shafiei Jandaghi, N.Z.; Naseri, M.; Hemmati, P.; Dadras, M.; Gouya, M.M.; Mokhtari Azad, T. Influenza virus but not MERS coronavirus circulation in Iran, 2013–2016: Comparison between pilgrims and general population. Travel Med. Infect. Dis. 2018, 21, 51–55. [Google Scholar] [CrossRef]
- Al-Abdallat, M.M.; Rha, B.; Alqasrawi, S.; Payne, D.C.; Iblan, I.; Binder, A.M.; Haddadin, A.; Nsour, M.A.; Alsanouri, T.; Mofleh, J.; et al. Acute respiratory infections among returning Hajj pilgrims—Jordan, 2014. J. Clin. Virol. 2017, 89, 34–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Memish, Z.A.; Assiri, A.; Almasri, M.; Alhakeem, R.F.; Turkestani, A.; Al Rabeeah, A.A.; Al-Tawfiq, J.A.; Alzahrani, A.; Azhar, E.; Makhdoom, H.Q.; et al. Prevalence of MERS-CoV Nasal Carriage and Compliance With the Saudi Health Recommendations Among Pilgrims Attending the 2013 Hajj. J. Infect. Dis. 2014, 210, 1067–1072. [Google Scholar] [CrossRef] [Green Version]
- Kraaij – Dirkzwager, M.; Timen, A.; Dirksen, K.; Gelinck, L.; Leyten, E.; Groeneveld, P.; Jansen, C.; Jonges, M.; Raj, S.; Thurkow, I.; et al. Middle East respiratory syndrome coronavirus (MERS-CoV) infections in two returning travellers in the Netherlands, May 2014. Eurosurveillance 2014, 19, 20817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almasri, M.; Ahmed, Q.A.; Turkestani, A.; Memish, Z.A. Hajj abattoirs in Makkah: Risk of zoonotic infections among occupational workers. Vet. Med. Sci. 2019, 5, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Hakawi, A.; Rose, E.B.; Biggs, H.M.; Lu, X.; Mohammed, M.; Abdalla, O.; Abedi, G.R.; Alsharef, A.A.; Alamri, A.A.; Bereagesh, S.A.; et al. Middle East respiratory syndrome coronavirus, Saudi Arabia, 2017–2018. Emerg. Infect. Dis. 2019, 25, 2149–2151. [Google Scholar] [CrossRef] [PubMed]
- El-Kafrawy, S.A.; Corman, V.M.; Tolah, A.M.; Al Masaudi, S.B.; Hassan, A.M.; Müller, M.A.; Bleicker, T.; Harakeh, S.M.; Alzahrani, A.A.; Alsaaidi, G.A.; et al. Enzootic patterns of Middle East respiratory syndrome coronavirus in imported African and local Arabian dromedary camels: A prospective genomic study. Lancet Planet. Health 2019, 3, e521–e528. [Google Scholar] [CrossRef] [Green Version]
- Kuwait News Agency (KUNA). Camel Slaughtering in Hajj Banned for Fear of Coronavirus. Available online: https://www.kuna.net.kw/ArticleDetails.aspx?id=2460041&language=en (accessed on 19 November 2020).
- Zheng, J. SARS-coV-2: An emerging coronavirus that causes a global threat. Int. J. Biol. Sci. 2020, 16, 1678–1685. [Google Scholar] [CrossRef] [Green Version]
- Symptoms of Coronavirus. Available online: https://www.cdc.gov/coronavirus/2019-ncov/symptoms-testing/symptoms.html (accessed on 19 July 2020).
- Li, G.; He, X.; Zhang, L.; Ran, Q.; Wang, J.; Xiong, A.; Wu, D.; Chen, F.; Sun, J.; Chang, C. Assessing ACE2 expression patterns in lung tissues in the pathogenesis of COVID-19. J. Autoimmun. 2020, 112, 102463. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, X.; Hu, T.; Li, J.; Song, H.; Liu, Y.; Wang, P.; Liu, D.; Yang, J.; Holmes, E.C.; et al. A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Curr. Biol. 2020, 30, 2196–2203.e3. [Google Scholar] [CrossRef]
- Chan, J.F.W.; Kok, K.H.; Zhu, Z.; Chu, H.; To, K.K.W.; Yuan, S.; Yuen, K.Y. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg. Microbes Infect. 2020, 9, 221–236. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Xiao, X.; Wei, X.; Li, J.; Yang, J.; Tan, H.; Zhu, J.; Zhang, Q.; Wu, J.; Liu, L. Composition and divergence of coronavirus spike proteins and host ACE2 receptors predict potential intermediate hosts of SARS-CoV-2. J. Med. Virol. 2020, 92, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Boni, M.F.; Lemey, P.; Jiang, X.; Lam, T.T.; Perry, B.W.; Castoe, T.A.; Rambaut, A.; Robertson, D.L. Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. Nat. Microbiol. 2020, 5. [Google Scholar] [CrossRef]
- Lee, J.; Hughes, T.; Lee, M.-H.; Field, H.; Rovie-Ryan, J.J.; Sitam, F.T.; Sipangkui, S.; Nathan, S.; Ramirez, D.; Kumar, S.V.; et al. No Evidence of Coronaviruses or Other Potentially Zoonotic Viruses in Sunda pangolins (Manis javanica) Entering the Wildlife Trade via Malaysia. Ecohealth 2020. [Google Scholar] [CrossRef]
- Alqutob, R.; Al Nsour, M.; Tarawneh, M.R.; Ajlouni, M.; Khader, Y.; Aqel, I.; Kharabsheh, S.; Obeidat, N. COVID-19 crisis in Jordan: Response, scenarios, strategies, and recommendations (Preprint). JMIR Public Health Surveill. 2020, 6, e19332. [Google Scholar] [CrossRef] [PubMed]
- Santucci, E. What Lies ahead as Jordan Faces the Fallout of COVID-19 - Atlantic Council. Available online: https://www.atlanticcouncil.org/blogs/menasource/what-lies-ahead-as-jordan-faces-the-fallout-of-covid-19/ (accessed on 19 July 2020).
- El-Khatib, Z.; Al Nsour, M.; Khader, Y.S.; Abu Khudair, M. Mental health support in Jordan for the general population and for the refugees in the Zaatari camp during the period of COVID-19 lockdown. Psychol. Trauma Theory Res. Pract. Policy 2020, 12, 511–514. [Google Scholar] [CrossRef]
- Fawad, M.; Rawashdeh, F.; Parmar, P.K.; Ratnayake, R. Simple ideas to mitigate the impacts of the COVID-19 epidemic on refugees with chronic diseases. Confl. Health 2020, 14, 23. [Google Scholar] [CrossRef]
- Yezli, S.; Khan, A. COVID-19 social distancing in the Kingdom of Saudi Arabia: Bold measures in the face of political, economic, social and religious challenges. Travel Med. Infect. Dis. 2020, 37, 101692. [Google Scholar] [CrossRef]
- Ebrahim, S.H.; Memish, Z.A. COVID-19: Preparing for superspreader potential among Umrah pilgrims to Saudi Arabia. Lancet 2020, 395, e48. [Google Scholar] [CrossRef] [Green Version]
- Ebrahim, S.H.; Memish, Z.A. Saudi Arabia’s drastic measures to curb the COVID-19 outbreak: Temporary suspension of the Umrah pilgrimage. J. Travel Med. 2020, 27, taaa029. [Google Scholar] [CrossRef] [Green Version]
- Gostin, L.O. International infectious disease law: Revision of the World Health Organization’s International Health Regulations. J. Am. Med. Assoc. 2004, 291, 2623–2627. [Google Scholar] [CrossRef]
- Fidler, D.P.; Gostin, L.O. The New International Health Regulations: An Historic Development for International Law and Public Health. J. Law Med. Ethics 2006, 34, 85–94. [Google Scholar] [CrossRef] [Green Version]
- Kandel, N.; Chungong, S.; Omaar, A.; Xing, J. Health security capacities in the context of COVID-19 outbreak: An analysis of International Health Regulations annual report data from 182 countries. Lancet 2020, 395, 1047–1053. [Google Scholar] [CrossRef]
- Nsubuga, P.; White, M.E.; Thacker, S.B.; Anderson, M.A.; Blount, S.B.; Broome, C.V.; Chiller, T.M.; Espitia, V.; Imtiaz, R.; Sosin, D.; et al. Chapter 53. Public Health Surveillance: A Tool for Targeting and Monitoring Interventions. In Disease Control Priorities in Developing Countries, 2nd ed.; The World Bank: Washington, DC, USA, 2006; pp. 997–1016. ISBN 0821361791. [Google Scholar]
- Subramanian, R.E.; Herrera, D.G.; Kelly, P.M. An evaluation of the global network of field epidemiology and laboratory training programmes: A resource for improving public health capacity and increasing the number of public health professionals worldwide. Hum. Resour. Health 2013, 11, 45. [Google Scholar] [CrossRef] [Green Version]
- Al Nsour, M.; Bashier, H.; Al Serouri, A.; Malik, E.; Khader, Y.; Saeed, K.; Ikram, A.; Abdalla, A.M.; Belalia, A.; Assarag, B.; et al. The role of the global health development/eastern mediterranean public health network and the eastern mediterranean field epidemiology training programs in preparedness for COVID-19. J. Med. Internet Res. 2020, 22, e18503. [Google Scholar] [CrossRef]
- Sheikhali, S.A.; Abdallat, M.; Mabdalla, S.; Qaseer, B.A.; Khorma, R.; Malik, M.; Profili, M.C.; Rø, G.; Haskew, J. Design and implementation of a national public health surveillance system in Jordan. Int. J. Med. Inform. 2016, 88, 58–61. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Comprehensive Assessment of Jordan’s Health Information System 2016. Available online: https://applications.emro.who.int/docs/9789290222583-eng.pdf?ua=1 (accessed on 27 February 2020).
- Abdulrahim, N.; Alasasfeh, I.; Khader, Y.S.; Iblan, I. Knowledge, Awareness, and Compliance of Disease Surveillance and Notification Among Jordanian Physicians in Residency Programs. Inquiry 2019, 56, 0046958019856508. [Google Scholar] [CrossRef]
- Al Nsour, M.; Iblan, I.; Tarawneh, M.R. Jordan field epidemiology training program: Critical role in national and regional capacity building. J. Med. Internet Res. 2018, 20, 1. [Google Scholar] [CrossRef]
- Al-Nsour, M.; Zindah, M.; Belbeisi, A.; Hadaddin, R.; Brown, D.W.; Walke, H. Prevalence of selected chronic, noncommunicable disease risk factors in Jordan: Results of the 2007 Jordan Behavioral Risk Factor Surveillance Survey. Prev. Chronic Dis. 2012, 9, E25. [Google Scholar] [CrossRef] [Green Version]
- Boyd, A.T.; Cookson, S.T.; Almashayek, I.; Yaacoub, H.; Qayyum, M.S.; Galev, A. An evaluation of a tuberculosis case-finding and treatment program among Syrian refugees - Jordan and Lebanon, 2013–2015. Confl. Health 2019, 13, 32. [Google Scholar] [CrossRef]
- Saikal, S.L.; Ge, L.; Mir, A.; Pace, J.; Abdulla, H.; Leong, K.F.; Benelkahla, M.; Olabi, B.; Medialdea-Carrera, R.; Padovese, V. Skin disease profile of Syrian refugees in Jordan: A field-mission assessment. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 419–425. [Google Scholar] [CrossRef]
- Lessler, J.; Rodriguez-Barraquer, I.; Cummings, D.A.T.; Garske, T.; Van Kerkhove, M.; Mills, H.; Truelove, S.; Hakeem, R.; Albarrak, A.; Ferguson, N.M. Estimating Potential Incidence of MERS-CoV Associated with Hajj Pilgrims to Saudi Arabia, 2014. PLoS Curr. 2014, 6. [Google Scholar] [CrossRef]
- Aljoudi, A.S. A University of the Hajj? Lancet 2013, 382, 1689. [Google Scholar] [CrossRef]
- Al Turki, Y.A. Mass Gathering Medicine New discipline to Deal with Epidemic and Infectious Diseases in the Hajj Among Muslim Pilgrimage: A Mini Review Article. J. Relig. Health 2016, 55, 1270–1274. [Google Scholar] [CrossRef]
- Field Epidemiology Training Program—Supervisor’s-General Message. Available online: https://www.moh.gov.sa/depten/Epidemiology/Pages/default.aspx (accessed on 27 February 2020).
- Field Epidemiology Training Program|College of Medicine. Available online: https://medicine.ksu.edu.sa/en/node/5783 (accessed on 27 February 2020).
- Razavi, S.; Saeednejad, M.; Salamati, P. Vaccination in Hajj: An overview of the recent findings. Int. J. Prev. Med. 2016, 7, 129. [Google Scholar]
- Yusef, D.; Hayajneh, W.; Awad, S.; Momany, S.; Khassawneh, B.; Samrah, S.; Obeidat, B.; Raffee, L.; Al-Faouri, I.; Bani Issa, A.; et al. Large outbreak of coronavirus disease among wedding attendees, Jordan. Emerg. Infect. Dis. 2020, 26, 2165–2167. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Bat Species (Common Name) | Jordan | Saudi Arabia | Associated Pathogen |
|---|---|---|---|
| Emballonuridae | |||
| Taphozous nudiventris | ● | ● | n/a |
| Taphozous perforatus | ● | ● | MERS-CoV [67] |
| Hipposideridae | |||
| Asellia tridens | ● | ● | n/a |
| Molossidae | |||
| Chaerephon pumilus | ● | Chikungunya virus; Dakar bat virus; Entebbe bat virus [10] | |
| Mops midas | ● | n/a | |
| Tadarida teniotis | ● | MRV [59]; multi-drug resistant E. coli [68] | |
| Tadarida aegyptiaca | ● | n/a | |
| Nycteridae | |||
| Nycteris thebaica | ● | Duvenhage lyssavirus [69] | |
| Pteropodidae | |||
| Rousettus aegyptiacus | ● | ● | EBLV-1 [60]; herpesviruses [70]; Marburg viruses [71,72] |
| Rhinolophidae | |||
| Rhinolophus blasii | ● | SARS-related CoV [73] | |
| Rhinolophus clivosus | ● | ● | Novel β-CoV [74] |
| Rhinolophus euryale | ● | SARS-related CoV [73] | |
| Rhinolophus ferrumequinum | ● | Adenoviruses [70]; α-CoV [75]; β-CoV [76]; EBLV-1 [77]; γ-herpesvirus [78]; SARS-related CoV [73,75,79] | |
| Rhinolophus hipposideros | ● | ● | β-CoV and SARS-like CoV [80]; MRV [59] |
| Rhinolophus mehelyi | ● | SARS-related CoV [73] | |
| Rhinopomatidae | |||
| Rhinopoma hardwickii | ● | ● | Hepe-Astroviruses, hepatoviruses, paramyxoviruses & rotaviruses [81] |
| Rhinopoma microphyllum | ● | ● | n/a |
| Vespertilionidae | |||
| Eptesicusbottae | ● | n/a | |
| Eptesicus nasutus | ● | n/a | |
| Eptesicus serotinus | ● | Astroviruses [82]; β-CoV [83]; EBLV-1 [84,85,86]; γ-herpesvirus [87] | |
| Miniopterus schreibersi | ● | EBLV-1 [77]; Lleida bat lyssavirus [88,89]; Lloviu virus [90]; WCBL [91] | |
| Myotis capaccinii | ● | EBLV-1 [65] | |
| Myotis emarginatus | ● | α-CoV & paramyxoviruses [79]; astroviruses [82] | |
| Myotis nattereri | ● | α-CoV [76]; Bokeloh bat lyssavirus [92,93]; EBLV-1 [77]; herpesviruses [94] | |
| Nycticeinops schlieffeni | ● | n/a | |
| Nyctalus noctula | ● | Adenoviruses [70]; astroviruses [82]; β-CoV [95]; hantavirus [96] | |
| Otonycteris hemprichii | ● | ● | n/a |
| Pipistrellus ariel | ● | n/a | |
| Pipistrellus bodenheimeri | ● | n/a | |
| Pipistrellus kuhlii | ● | ● | α-CoV [95]; β-CoV [97]; MRV [59]; novel rhabdovirus [98]; paramyxoviruses [76]; Toscana virus [99] |
| Pipistrellus nathusii | ● | EBLV-1 [100] | |
| Pipistrellus pipistrellus | ● | EBLV-1 [100] | |
| Pipistrellus rueppellii | ● | n/a | |
| Plecotus austriacus | ● | n/a |
| Index | Jordan (2016) | Saudi Arabia (2017) |
|---|---|---|
| Capacity to prevent | 3 | 4 |
| Capacity to detect | 4 | 4 |
| Capacity to respond | 3 | 5 |
| Enabling function | 3 | 4 |
| Operational index | 3 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
AL-Eitan, L.N.; Tarkhan, A.H.; Alghamdi, M.A.; Marston, D.A.; Wu, G.; McElhinney, L.M.; Brown, I.H.; Fooks, A.R. Bat-Borne Coronaviruses in Jordan and Saudi Arabia: A Threat to Public Health? Viruses 2020, 12, 1413. https://doi.org/10.3390/v12121413
AL-Eitan LN, Tarkhan AH, Alghamdi MA, Marston DA, Wu G, McElhinney LM, Brown IH, Fooks AR. Bat-Borne Coronaviruses in Jordan and Saudi Arabia: A Threat to Public Health? Viruses. 2020; 12(12):1413. https://doi.org/10.3390/v12121413
Chicago/Turabian StyleAL-Eitan, Laith N., Amneh H. Tarkhan, Mansour A. Alghamdi, Denise A. Marston, Guanghui Wu, Lorraine M. McElhinney, Ian H. Brown, and Anthony R. Fooks. 2020. "Bat-Borne Coronaviruses in Jordan and Saudi Arabia: A Threat to Public Health?" Viruses 12, no. 12: 1413. https://doi.org/10.3390/v12121413
APA StyleAL-Eitan, L. N., Tarkhan, A. H., Alghamdi, M. A., Marston, D. A., Wu, G., McElhinney, L. M., Brown, I. H., & Fooks, A. R. (2020). Bat-Borne Coronaviruses in Jordan and Saudi Arabia: A Threat to Public Health? Viruses, 12(12), 1413. https://doi.org/10.3390/v12121413