Serpentovirus (Nidovirus) and Orthoreovirus Coinfection in Captive Veiled Chameleons (Chamaeleo calyptratus) with Respiratory Disease



Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
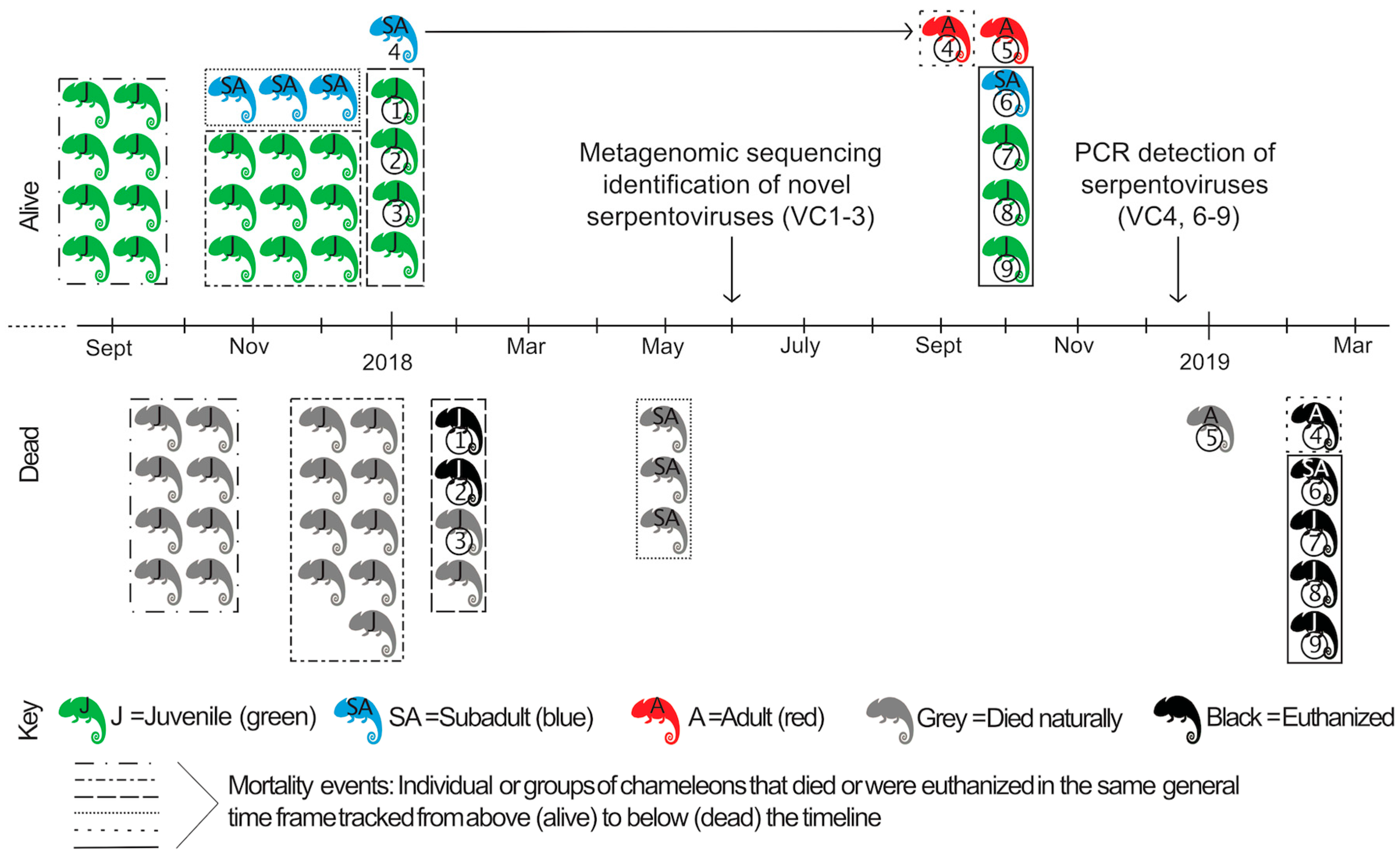
2.2. Case History
2.3. Metagenomic Sequencing and Data Analysis
2.4. PCR Analysis for Viral RNA
2.5. Virus Isolation
3. Results
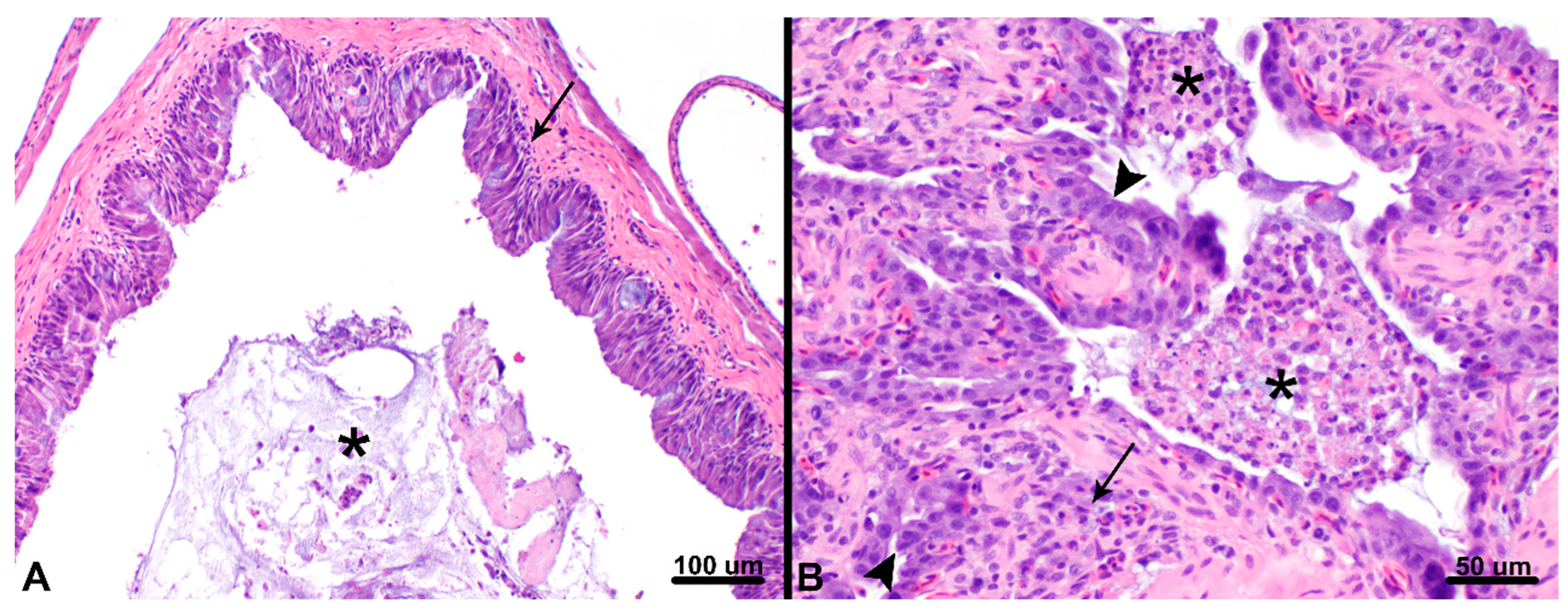
3.1. Postmortem Findings
3.2. Metagenomic Sequencing Findings
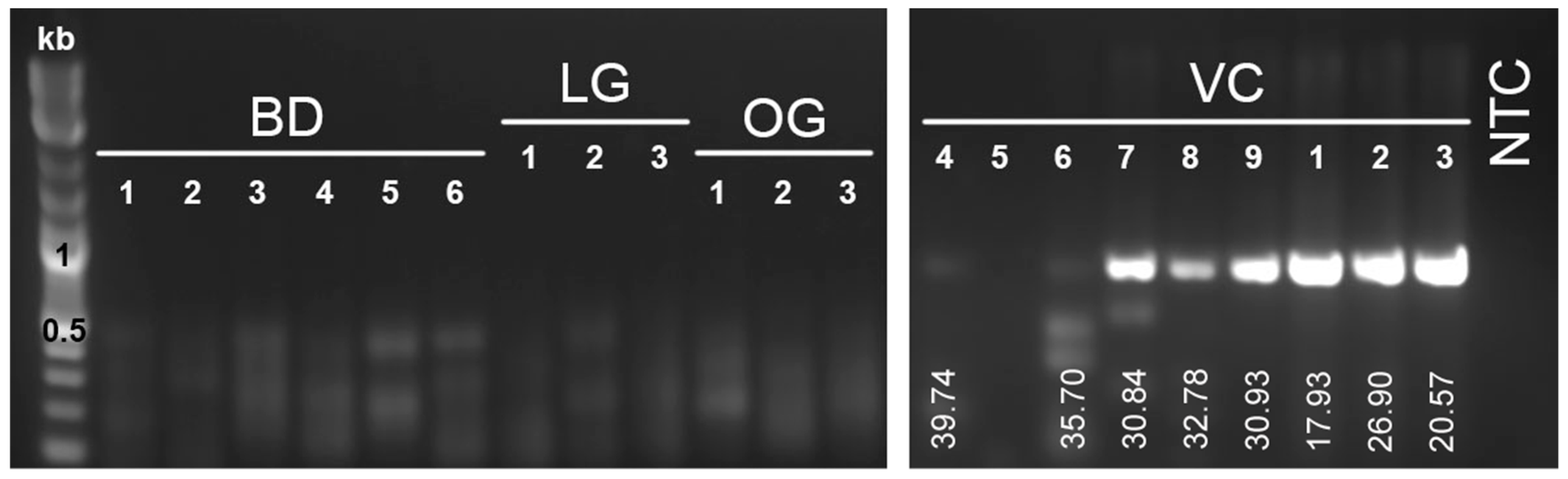
3.3. Targeted Viral RNA Detection
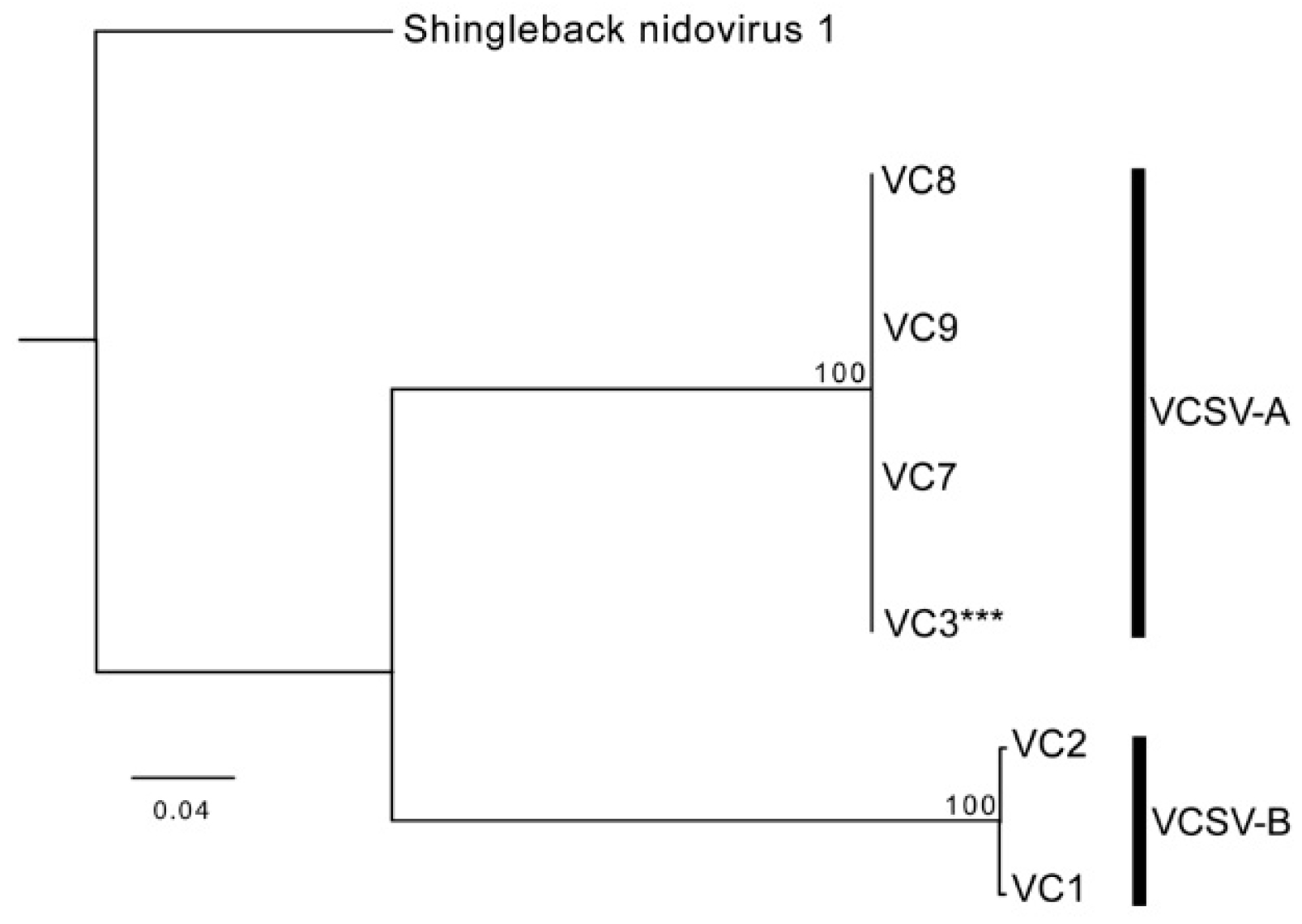
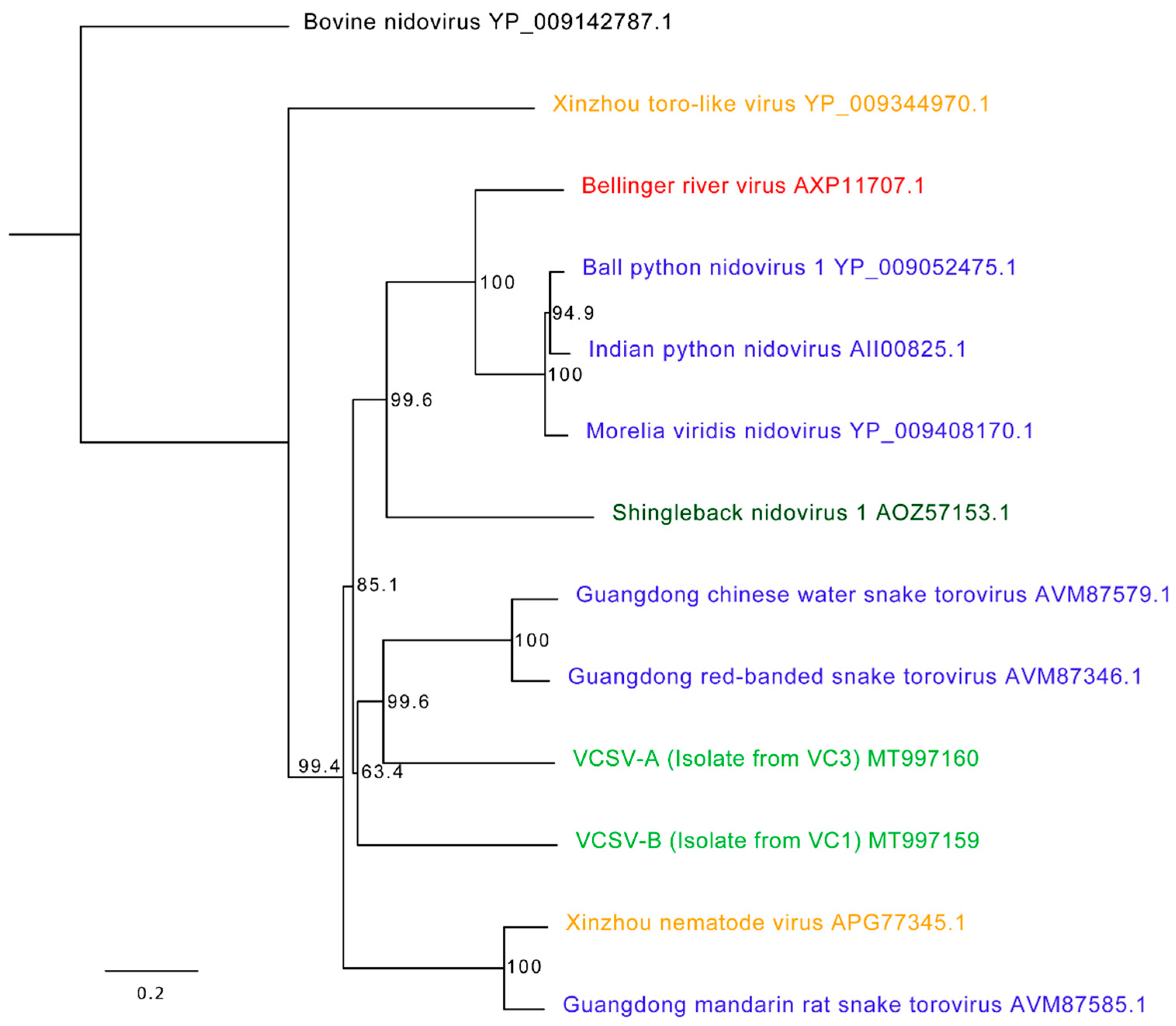
3.4. Chameleon Serpentoviruses: Genomic and Phylogenetic Analysis
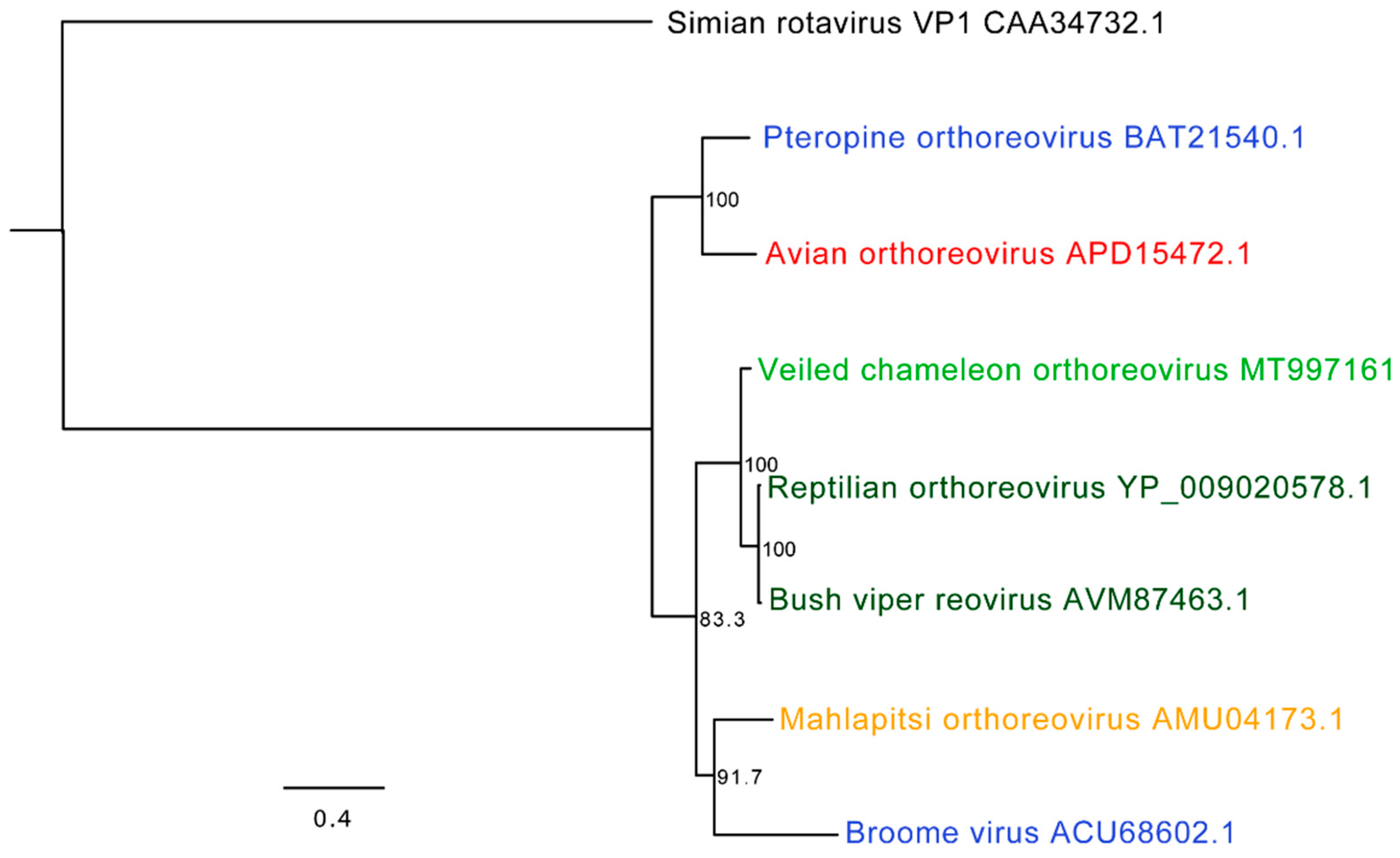
3.5. Chameleon Orthoreoviruses: Genomic and Phylogenetic Analysis
3.6. Virus Isolation
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jacobson, E.R. Infectious Diseases and Pathology of Reptiles; Viruses and Viral Diseases of Reptiles; CRC Press: Boca Raton, FL, USA, 2007; Chapter 9; ISBN 978-1-4200-0403-8. [Google Scholar]
- Marschang, R.E. Viruses infecting reptiles. Viruses 2011, 3, 2087–2126. [Google Scholar]
- Chang, W.-S.; Li, C.-X.; Hall, J.; Eden, J.-S.; Hyndman, T.H.; Holmes, E.C.; Rose, K. Meta-Transcriptomic Discovery of a Divergent Circovirus and a Chaphamaparvovirus in Captive Reptiles with Proliferative Respiratory Syndrome. Viruses 2020, 12, 1073. [Google Scholar] [CrossRef]
- Stenglein, M.D.; Jacobson, E.R.; Wozniak, E.J.; Wellehan, J.F.X.; Kincaid, A.; Gordon, M.; Porter, B.F.; Baumgartner, W.; Stahl, S.; Kelley, K.; et al. Ball python nidovirus: A candidate etiologic agent for severe respiratory disease in Python regius. mBio 2014, 5, e01484-14. [Google Scholar] [CrossRef]
- Uccellini, L.; Ossiboff, R.J.; de Matos, R.E.C.; Morrisey, J.K.; Petrosov, A.; Navarrete-Macias, I.; Jain, K.; Hicks, A.L.; Buckles, E.L.; Tokarz, R.; et al. Identification of a novel nidovirus in an outbreak of fatal respiratory disease in ball pythons (Python regius). Virol. J. 2014, 11, 144. [Google Scholar] [CrossRef]
- Bodewes, R.; Lempp, C.; Schurch, A.C.; Habierski, A.; Hahn, K.; Lamers, M.; von Dornberg, K.; Wohlsein, P.; Drexler, J.F.; Haagmans, B.L.; et al. Novel divergent nidovirus in a python with pneumonia. J. Gen. Virol. 2014, 95, 2480–2485. [Google Scholar] [CrossRef]
- Dervas, E.; Hepojoki, J.; Laimbacher, A.; Romero-Palomo, F.; Jelinek, C.; Keller, S.; Smura, T.; Hepojoki, S.; Kipar, A.; Hetzel, U. Nidovirus-associated proliferative pneumonia in the green tree python (Morelia viridis). J. Virol. 2017. [Google Scholar] [CrossRef]
- Ziebuhr, J.; Baric, R.S.; Baker, S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.; Haagmans, B.L.; Neuman, B.W.; Perlman, S.; Poon, L.L.M.; et al. Reorganization of the Family Coronaviridae into Two Families, Coronaviridae (Including the Current Subfamily Coronavirinae and the New Subfamily Letovirinae) and the New Family Tobaniviridae (Accommodating the Current Subfamily Torovirinae and Three Other Subfamilies), Revision of the Genus Rank Structure and Introduction of a New Subgenus Rank 2018; International Committee on Taxonomy of Viruses: London, UK, 2018. [Google Scholar]
- O’Dea, M.A.; Jackson, B.; Jackson, C.; Xavier, P.; Warren, K. Discovery and partial genomic characterisation of a novel nidovirus associated with respiratory disease in wild shingleback lizards (Tiliqua rugosa). PLoS ONE 2016, 11, e0165209. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.-D.; Tian, J.-H.; Chen, L.-J.; Chen, X.; Li, C.-X.; Qin, X.-C.; Li, J.; Cao, J.-P.; Eden, J.-S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef]
- Marschang, R.E.; Kolesnik, E. Detection of nidoviruses in live pythons and boas. Tierarztl. Prax. Ausg. K Kleintiere Heimtiere 2017, 45, 22–26. [Google Scholar] [CrossRef]
- Shi, M.; Lin, X.-D.; Chen, X.; Tian, J.-H.; Chen, L.-J.; Li, K.; Wang, W.; Eden, J.-S.; Shen, J.-J.; Liu, L.; et al. The evolutionary history of vertebrate RNA viruses. Nature 2018, 556, 197–202. [Google Scholar] [CrossRef]
- Zhang, J.; Finlaison, D.S.; Frost, M.J.; Gestier, S.; Gu, X.; Hall, J.; Jenkins, C.; Parrish, K.; Read, A.J.; Srivastava, M.; et al. Identification of a novel nidovirus as a potential cause of large scale mortalities in the endangered Bellinger River snapping turtle (Myuchelys georgesi). PLoS ONE 2018, 13, e0205209. [Google Scholar] [CrossRef]
- Rampacci, E.; Masi, M.; Origgi, F.C.; Stefanetti, V.; Bottinelli, M.; Selleri, P.; Coletti, M.; Passamonti, F. First molecular detection of ball python nidovirus in Italy—Short communication. Acta Vet. Hung. 2019, 67, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Hoon-Hanks, L.L.; Layton, M.L.; Ossiboff, R.J.; Parker, J.S.L.; Dubovi, E.J.; Stenglein, M.D. Respiratory disease in ball pythons (Python regius) experimentally infected with ball python nidovirus. Virol. Nidovirus Spec. Issue 2018, 517, 77–87. [Google Scholar] [CrossRef]
- Hoon-Hanks, L.L.; Ossiboff, R.J.; Bartolini, P.; Fogelson, S.B.; Perry, S.M.; Stohr, A.C.; Cross, S.T.; Wellehan, J.F.X.; Jacobson, E.R.; Dubovi, E.J.; et al. Longitudinal and Cross-Sectional Sampling of Serpentovirus (Nidovirus) Infection in Captive Snakes Reveals High Prevalence, Persistent Infection, and Increased Mortality in Pythons and Divergent Serpentovirus Infection in Boas and Colubrids. Front. Vet. Sci. 2019, 6, 338. [Google Scholar] [CrossRef]
- Blahak, S.; Jenckel, M.; Höper, D.; Beer, M.; Hoffmann, B.; Schlottau, K. Investigations into the presence of nidoviruses in pythons. Virol. J. 2020, 17, 6. [Google Scholar] [CrossRef]
- Diaz, R.; Anderson, C.; Baumann, D.; Kupronis, R.; Jewell, D.; Piraquive, C.; Kupronis, J.; Winter, K.; Greek, T.; Trainor, P. Captive Care, Raising, and Breeding of the Veiled Chameleon (Chamaeleo calyptratus). Cold Spring Harb. Protoc. 2015, 2015, 943–949. [Google Scholar] [CrossRef]
- Bovarnick, M.R.; Miller, J.C.; Snyder, J.C. The influence of certain salts, amino acids, sugars, and proteins on the stability of rickettsiae. J. Bacteriol. 1950, 59, 509–522. [Google Scholar]
- Decker, C.J.; Steiner, H.R.; Hoon-Hanks, L.L.; Morrison, J.H.; Haist, K.C.; Stabell, A.C.; Poeschla, E.M.; Morrison, T.E.; Stenglein, M.D.; Sawyer, S.L.; et al. dsRNA-Seq: Identification of Viral Infection by Purifying and Sequencing dsRNA. Viruses 2019, 11, 943. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Wu, T.D.; Nacu, S. Fast and SNP-tolerant detection of complex variants and splicing in short reads. Bioinformatics 2010, 26, 873–881. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Misawa, K.; Katoh, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Wellehan, J.F.X.J.; Childress, A.L.; Marschang, R.E.; Johnson, A.J.; Lamirande, E.W.; Roberts, J.F.; Vickers, M.L.; Gaskin, J.M.; Jacobson, E.R. Consensus nested PCR amplification and sequencing of diverse reptilian, avian, and mammalian orthoreoviruses. Vet. Microbiol. 2009, 133, 34–42. [Google Scholar] [CrossRef]
- Stenglein, M.D.; Sanders, C.; Kistler, A.L.; Ruby, J.G.; Franco, J.Y.; Reavill, D.R.; Dunker, F.; Derisi, J.L. Identification, characterization, and in vitro culture of highly divergent arenaviruses from boa constrictors and annulated tree boas: Candidate etiological agents for snake inclusion body disease. mBio 2012, 3, e00180-12. [Google Scholar] [CrossRef]
- Abbas, M.D.; Marschang, R.E.; Schmidt, V.; Kasper, A.; Papp, T. A unique novel reptilian paramyxovirus, four atadenovirus types and a reovirus identified in a concurrent infection of a corn snake (Pantherophis guttatus) collection in Germany. Vet. Microbiol. 2011, 150, 70–79. [Google Scholar]
- Saberi, A.; Gulyaeva, A.A.; Brubacher, J.L.; Newmark, P.A.; Gorbalenya, A.E. A planarian nidovirus expands the limits of RNA genome size. PLoS Pathog. 2018, 14, e1007314. [Google Scholar] [CrossRef]
- Dervas, E.; Hepojoki, J.; Smura, T.; Prähauser, B.; Windbichler, K.; Blümich, S.; Ramis, A.; Hetzel, U.; Kipar, A. Serpentoviruses: More than Respiratory Pathogens. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Fields, B.N.; Knipe, D.M.; David, M.; Howley, P.M. Fields Virology, 6th ed.; Virology; Wolters Kluwer/Lippincott Williams & Wilkins Health: Philadelphia, PA, USA, 2013. [Google Scholar]
- Marschang, R.E.; Donahoe, S.; Manvell, R.; Lemos-Espinal, J. Paramyxovirus and reovirus infections in wild-caught Mexican lizards (Xenosaurus and Abronia spp.). J. Zoo Wildl. Med. 2002, 33, 317–321. [Google Scholar] [PubMed]
- Blahak, S.; Ott, I.; Vieler, E. Comparison of 6 different reoviruses of various reptiles. Vet. Res. 1995, 26, 470–476. [Google Scholar]
- Ahne, W.; Thomsen, I.; Winton, J. Isolation of a reovirus from the snake, Python regius. Brief report. Arch. Virol. 1987, 94, 135–139. [Google Scholar]
- Gravendyck, M.; Ammermann, P.; Marschang, R.E.; Kaleta, E.F. Paramyxoviral and reoviral infections of iguanas on Honduran Islands. J. Wildl. Dis. 1998, 34, 33–38. [Google Scholar]
- Raynaud, A.; Adrian, M. Cutaneous lesions with papillomatous structure associated with viruses in the green lizard (Lacerta viridis laurenti 1768). C. R. Acad. Hebd. Seances. Acad. Sci. D 1976, 283, 845–847. [Google Scholar]
- Drury, S.E.N.; Gough, R.E.; de Welchman, D.B. Isolation and identification of a reovirus from a lizard, Uromastyx hardwickii, in the United Kingdom. Vet. Rec. 2002, 151, 637–638. [Google Scholar]
- Marschang, R.E.; Papp, T. Isolation and Partial Characterization of Three Reoviruses from Lizards. J. Herpetol. Med. Surg. 2009, 19, 13–15. [Google Scholar] [CrossRef]
- Lamirande, E.W.; Nichols, D.K.; Owens, J.W.; Gaskin, J.M.; Jacobson, E.R. Isolation and experimental transmission of a reovirus pathogenic in ratsnakes (Elaphe species). Virus Res. 1999, 63, 135–141. [Google Scholar]
- Schragen, S. Experimetelle Infektion von Juvenilen Boa Constrictor Mit Einem Orthoreovirusisolat; Justus-Liebig-Universität Giessen: Giessen, Germany, 2006. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample and Seq Run | Total Reads (A) | Remove Low Quality Reads (B) | Collapse to Unique Reads (C) | Host Filter (D) | VCSV Reads | VCOrV Reads | |
|---|---|---|---|---|---|---|---|
| Shotgun RNA Sequencing | VC1 | 6.4 × 106 | 5.7 × 106 (89%) | 1 × 106 (15%) | 0.5 × 106 (8%) | 914 | 0 |
| VC2 | 1.5 × 106 | 1.4 × 106 (91%) | 0.3 × 106 (19%) | 0.1 × 106 (9%) | 0 | 0 | |
| VC3 | 1.1 × 106 | 1 × 106 (92%) | 0.2 × 106 (20%) | 0.1 × 106 (10%) | 212 | 0 | |
| dsRNA Sequencing | VC1 | 16.9 × 106 | 15.7 × 106 (93%) | 2.7 × 106 (16%) | 2.6 × 106 (15%) | 901 | 39 |
| VC2 | 42.7 × 106 | 38.5 × 106 (90%) | 6.6 × 106 (16%) | 6 × 106 (14%) | 11 | 2921 | |
| VC3 | 13.1 × 106 | 11.4 × 106 (87%) | 3.1 × 106 (24%) | 3 × 106 (23%) | 11,881 | 11 | |
| Species | ID | Sex | Age | Sample Type | Sample Tissue | Analysis | VCSV | VCOrV |
|---|---|---|---|---|---|---|---|---|
| Veiled chameleon (Chameleo calyptratus) | VC1 | M | Juvenile | PM | LLK | MGS, PCR | + | + |
| VC2 | F | Juvenile | PM | LLK | MGS, PCR | + | + | |
| VC3 | M | Juvenile | PM | LTOM | MGS, PCR | + | + | |
| VC4 | M | Adult | AM, PM | OS, LT | PCR | + | - * | |
| VC5 | F | Adult | AM | OS | PCR | - | - * | |
| VC6 | F | Subadult | AM, PM | OS, LT | PCR | + | - * | |
| VC7 | F | Juvenile | AM, PM | OS, LT | PCR | + | - * | |
| VC8 | F | Juvenile | AM, PM | OS, LT | PCR | + | - * | |
| VC9 | F | Juvenile | AM, PM | OS, LT | PCR | + | - * | |
| Central bearded dragon (Pogona vitticeps) | BD1 | F | Adult | AM | OS | PCR | - | NT |
| BD2 | F | Adult | AM | OS | PCR | - | NT | |
| BD3 | M | Adult | AM | OS | PCR | - | NT | |
| BD4 | F | Adult | AM | OS | PCR | - | NT | |
| BD5 | M | Adult | AM | OS | PCR | - | NT | |
| BD6 | F | Adult | AM | OS | PCR | - | NT | |
| Common leopard gecko (Eublepharis macularius) | LG1 | U | U | AM | OS | PCR | - | NT |
| LG2 | U | U | AM | OS | PCR | - | NT | |
| LG3 | U | U | AM | OS | PCR | - | NT | |
| Ocelot gecko (Paroedura pictus) | OG1 | U | U | AM | OS | PCR | - | NT |
| OG2 | U | U | AM | OS | PCR | - | NT | |
| OG3 | U | U | AM | OS | PCR | - | NT |
| Segment | Gene | Size (bp) | Protein Size (aa) | GenBank Accession |
|---|---|---|---|---|
| L1 | Lambda (λ) A | 4000 | 1131 | MT997161 |
| L2 | Lambda (λ) C | 3685 | 872 | MT997162 |
| L3 | Lambda (λ) B (RdRp) | 3795 | 1165 | MT997163 |
| M1 | Mu (μ) NS | 2467 | 793 | MT997164 |
| M2 | Mu (μ) A | 2289 | 754 | MT997165 |
| M3 | Mu (μ) B | 2091 | 613 | MT997166 |
| S1 | P14 (fusion), Sigma (σ) C | 1463 | 126,350 | MT997167 |
| S2 | Sigma (σ) A | 1260 | 390 | MT997168 |
| S3 | Sigma (σ) B | 1252 | 388 | MT997169 |
| S4 | Sigma (σ) NS | 1129 | 282 | MT997170 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoon-Hanks, L.L.; Stöhr, A.C.; Anderson, A.J.; Evans, D.E.; Nevarez, J.G.; Díaz, R.E.; Rodgers, C.P.; Cross, S.T.; Steiner, H.R.; Parker, R.R.; et al. Serpentovirus (Nidovirus) and Orthoreovirus Coinfection in Captive Veiled Chameleons (Chamaeleo calyptratus) with Respiratory Disease. Viruses 2020, 12, 1329. https://doi.org/10.3390/v12111329
Hoon-Hanks LL, Stöhr AC, Anderson AJ, Evans DE, Nevarez JG, Díaz RE, Rodgers CP, Cross ST, Steiner HR, Parker RR, et al. Serpentovirus (Nidovirus) and Orthoreovirus Coinfection in Captive Veiled Chameleons (Chamaeleo calyptratus) with Respiratory Disease. Viruses. 2020; 12(11):1329. https://doi.org/10.3390/v12111329
Chicago/Turabian StyleHoon-Hanks, Laura L., Anke C. Stöhr, Amanda J. Anderson, Dawn E. Evans, Javier G. Nevarez, Raúl E. Díaz, Case P. Rodgers, Shaun T. Cross, Halley R. Steiner, Roy R. Parker, and et al. 2020. "Serpentovirus (Nidovirus) and Orthoreovirus Coinfection in Captive Veiled Chameleons (Chamaeleo calyptratus) with Respiratory Disease" Viruses 12, no. 11: 1329. https://doi.org/10.3390/v12111329
APA StyleHoon-Hanks, L. L., Stöhr, A. C., Anderson, A. J., Evans, D. E., Nevarez, J. G., Díaz, R. E., Rodgers, C. P., Cross, S. T., Steiner, H. R., Parker, R. R., & Stenglein, M. D. (2020). Serpentovirus (Nidovirus) and Orthoreovirus Coinfection in Captive Veiled Chameleons (Chamaeleo calyptratus) with Respiratory Disease. Viruses, 12(11), 1329. https://doi.org/10.3390/v12111329