Stability of African Swine Fever Virus in Carcasses of Domestic Pigs and Wild Boar Experimentally Infected with the ASFV “Estonia 2014” Isolate



Abstract
1. Introduction
2. Materials and Methods
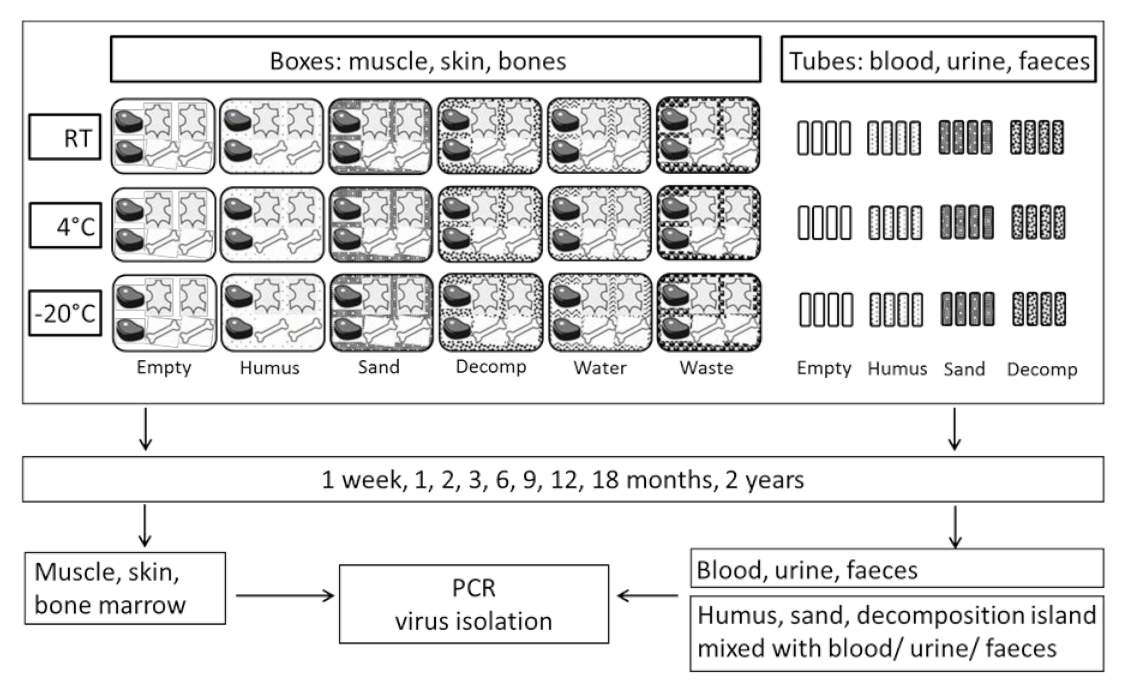
2.1. Study Design and Sample Collection
2.2. Laboratory Analyses
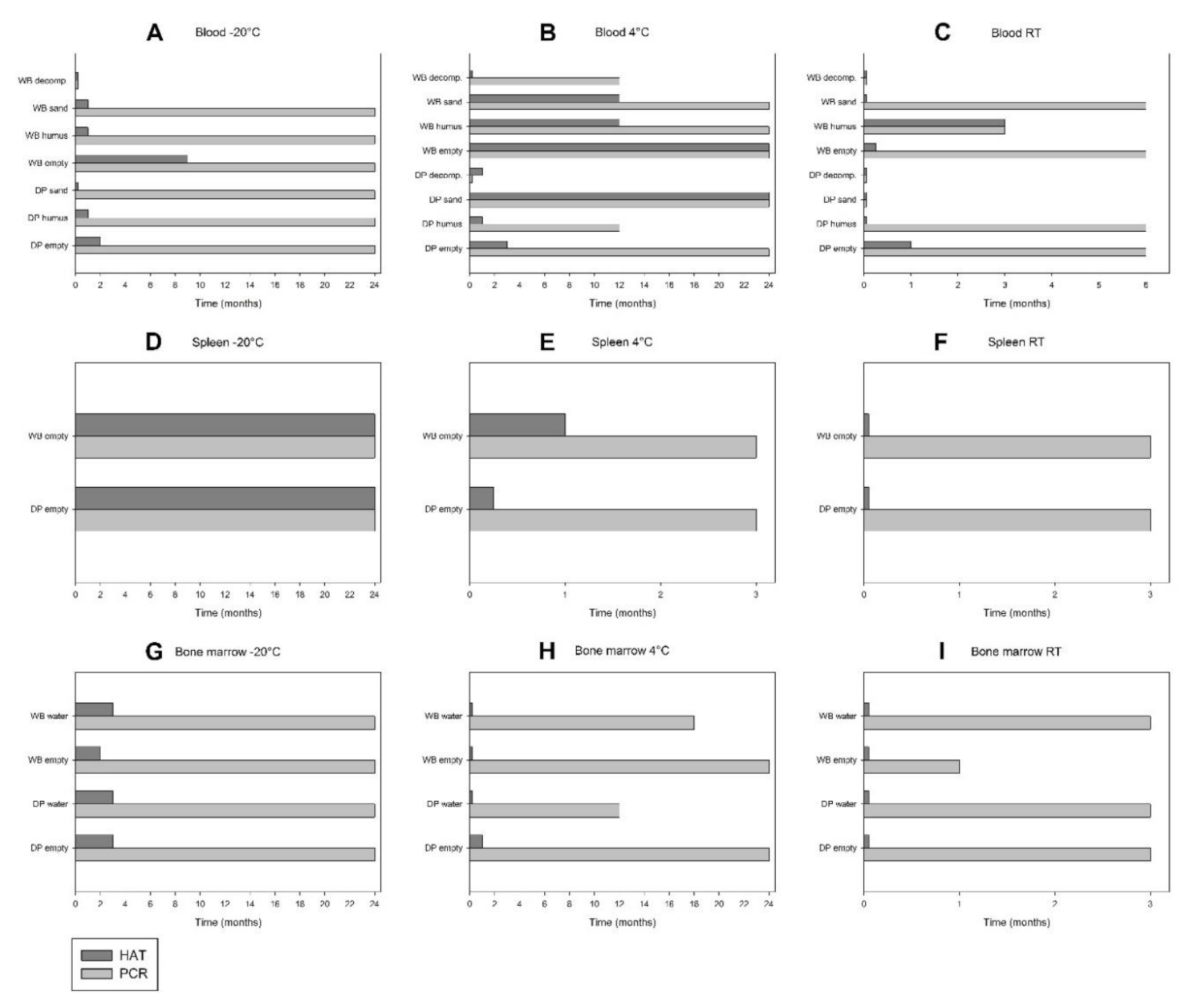
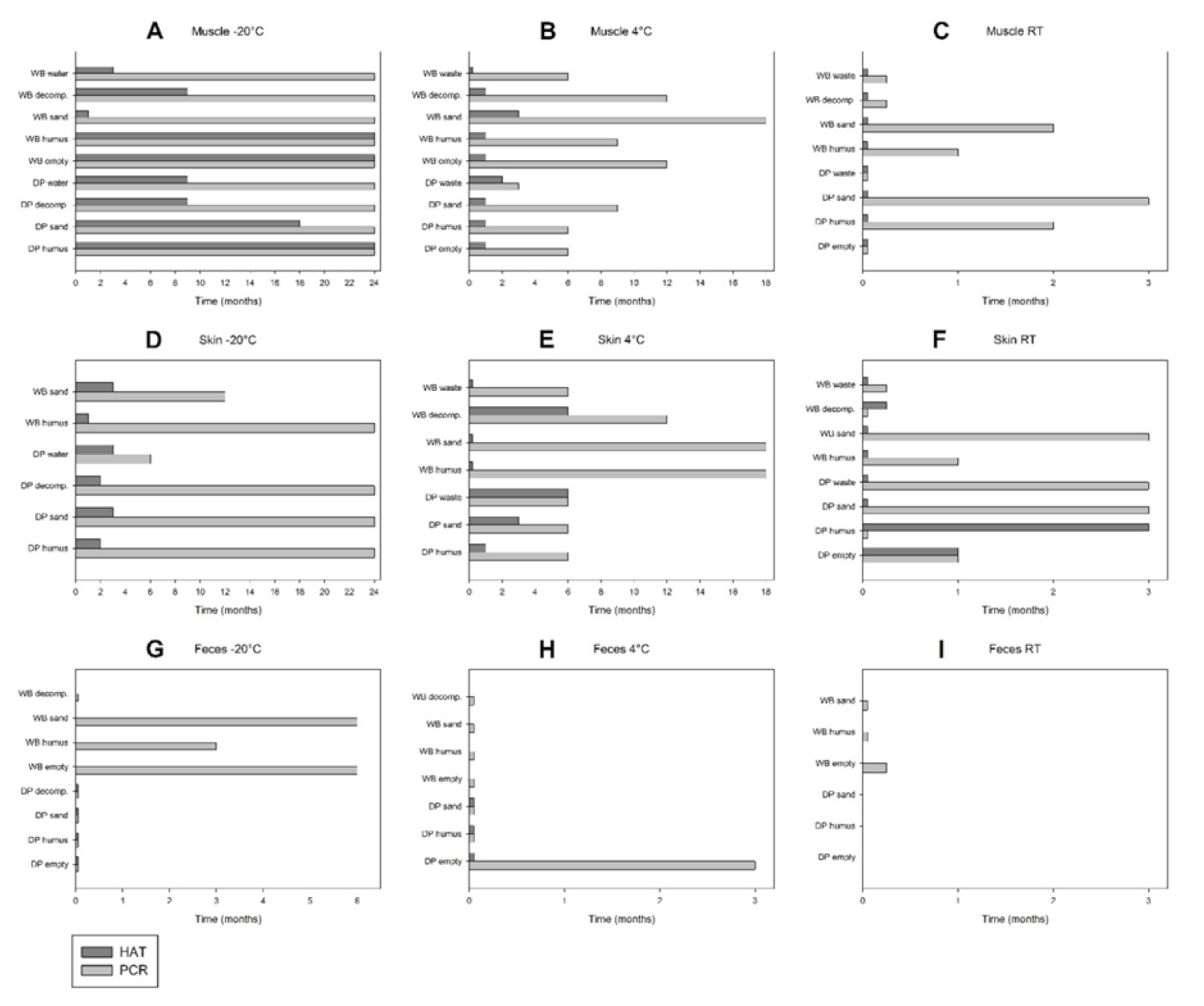
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Penrith, M.L.; Vosloo, W. Review of African swine fever: Transmission, spread and control. J. S. Afr. Vet. Assoc. 2009, 80, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Galindo, I.; Alonso, C. African Swine Fever Virus: A Review. Viruses 2017, 9, 103. [Google Scholar] [CrossRef] [PubMed]
- Animal Disease Notification System. Available online: https://ec.europa.eu/food/animals/animal-diseases/not-system_en (accessed on 31 August 2020).
- Sánchez-Vizcaíno, J.M.; Mur, L.; Martínez-López, B. African Swine Fever: An Epidemiological Update. Transbound. Emerg. Dis. 2012, 59 (Suppl. 1), 27–35. [Google Scholar] [CrossRef]
- Kolbasov, D.; Tsybanov, S.; Malogolovkin, A.; Gazaev, I.; Mikolaychuk, S.V. Detection of ASF virus in pork products. Veterinaria 2011, 10, 54–56. [Google Scholar]
- McKercher, P.D.; Yedloutschnig, R.J.; Callis, J.J.; Murphy, R.; Panina, G.F.; Civardi, A.; Bugnetti, M.; Foni, E.; Laddomada, A.; Scarano, C.; et al. Survival of Viruses in “Prosciutto di Parma” (Parma Ham). Can. Inst. Food Sci. Technol. J. 1987, 20, 267–272. [Google Scholar] [CrossRef]
- Petrini, S.; Feliziani, F.; Casciari, C.; Giammarioli, M.; Torresi, C.; De Mia, G.M. Survival of African swine fever virus (ASFV) in various traditional Italian dry-cured meat products. Prev. Vet. Med. 2019, 162, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Mebus, C.A.; House, C.; Gonzalvo, F.R.; Pineda, J.M.; Tapiador, J.; Pire, J.J.; Bergada, J.; Yedloutschnig, R.J.; Sahu, S.; Becerra, V.; et al. Survival of foot-and-mouth disease, African swine fever, and hog cholera viruses in Spanish serrano cured hams and Iberian cured hams, shoulders and loins. Food Microbiol. 1993, 10, 133–143. [Google Scholar] [CrossRef]
- Mebus, C.; Arias, M.; Pineda, J.M.; Tapiador, J.; House, C.; Sánchez-Vizcaíno, J.M. Survival of several porcine viruses in different Spanish dry-cured meat products. Food Chem. 1997, 59, 555–559. [Google Scholar] [CrossRef]
- Jurado, C.; Mur, L.; Pérez Aguirreburualde, M.S.; Cadenas-Fernández, E.; Martínez-López, B.; Sánchez-Vizcaíno, J.M.; Perez, A. Risk of African swine fever virus introduction into the United States through smuggling of pork in air passenger luggage. Sci. Rep. 2019, 9, 14423. [Google Scholar] [CrossRef]
- Taylor, R.A.; Condoleo, R.; Simons, R.R.L.; Gale, P.; Kelly, L.A.; Snary, E.L. The Risk of Infection by African Swine Fever Virus in European Swine Through Boar Movement and Legal Trade of Pigs and Pig Meat. Front. Vet. Sci. 2020, 6. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on African swine fever. EFSA J. 2014, 12, 3628. [Google Scholar]
- FLI. Qualitative Risikobewertung zur Einschleppung der Afrikanischen Schweinepest aus Verbreitungsgebieten in Europa nach Deutschland. OpenAgrar 2019. Available online: https://www.openagrar.de/servlets/MCRFileNodeServlet/openagrar_derivate_00030315/ASP_Risikobewertung_2020-05-25.pdf (accessed on 31 August 2020).
- Berg, C.; Botner, A.; Browman, H.; De Koeijer, A.; Domingo, M.; Ducrot, C.; Edwards, S.; Fourichon, C.; Koenen, F.; More, S.; et al. African Swine Fever. EFSA J. 2015, 13, 4163. [Google Scholar] [CrossRef]
- Chenais, E.; Depner, K.; Guberti, V.; Dietze, K.; Viltrop, A.; Ståhl, K. Epidemiological considerations on African swine fever in Europe 2014–2018. Porc. Health Manag. 2019, 5. [Google Scholar] [CrossRef]
- Guinat, C.; Gogin, A.; Blome, S.; Keil, G.; Pollin, R.; Pfeiffer, D.U.; Dixon, L. Transmission routes of African swine fever virus to domestic pigs: Current knowledge and future research directions. Vet. Rec. 2016, 178, 262–267. [Google Scholar] [CrossRef]
- Cukor, J.; Linda, R.; Václavek, P.; Mahlerová, K.; Šatrán, P.; Havránek, F. Confirmed cannibalism in wild boar and its possible role in African swine fever transmission. Transbound. Emerg. Dis. 2019. [Google Scholar] [CrossRef]
- Probst, C.; Globig, A.; Knoll, B.; Conraths, F.J.; Depner, K. Behaviour of free ranging wild boar towards their dead fellows: Potential implications for the transmission of African swine fever. R. Soc. Open Sci. 2017, 4. [Google Scholar] [CrossRef]
- Pepin, K.M.; Golnar, A.J.; Abdo, Z.; Podgórski, T. Ecological drivers of African swine fever virus persistence in wild boar populations: Insight for control. Ecol. Evol. 2020, 10, 2846–2859. [Google Scholar] [CrossRef]
- Boklund, A.; Cay, B.; Depner, K.; Földi, Z.; Guberti, V.; Masiulis, M.; Miteva, A.; More, S.; Olsevskis, E.; Šatrán, P.; et al. Epidemiological analyses of African swine fever in the European Union (November 2017 until November 2018). EFSA J. 2018, 16, e05494. [Google Scholar] [CrossRef]
- Depner, K.; Blome, S.; Staubach, C.; Probst, C.; Globig, A.; Dietze, K.; Sauter-Louis, C.; Conraths, F.J. African swine fever—A habitat-borne disease often of low contagiosity. Prakt. Tierarzt 2016, 6, 536–544. [Google Scholar]
- Álvarez, J.; Bicout, D.; Boklund, A.; Bøtner, A.; Depner, K.; More, S.; Roberts, H.; Ståhl, K.; Thulke, H.-H.; Viltrop, A.; et al. Research gap analysis on African swine fever. EFSA J. 2019, 17. [Google Scholar] [CrossRef]
- Brown, V.R.; Bevins, S.N. A Review of African Swine Fever and the Potential for Introduction into the United States and the Possibility of Subsequent Establishment in Feral Swine and Native Ticks. Front. Vet. Sci. 2018, 5, 11. [Google Scholar] [CrossRef] [PubMed]
- Adkin, A.; Coburn, H.; England, T.; Hall, S.; Hartnett, E.; Marooney, C.; Wooldridge, M.; Watson, E.; Cooper, J.; Cox., T.; et al. Risk Assessment for the Illegal Import of Contaminated Meat and Meat Products into Great Britain and the Subsequent Exposure of GB Livestock (IIRA): Foot and Mouth Disease (FMD), Classical Swine Fever (CSF), African Swine Fever (ASF), Swine Vesicular Disease (SVD); Veterinary Laboratories Agency: New Hawk, UK, 2004. [Google Scholar]
- Haas, B.; Ahl, R.; Bohm, R.; Strauch, D. Inactivation of viruses in liquid manure. Rev. Sci. Tech. 1995, 14, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Plowright, W.; Thomson, G.R.; Neser, J.A. African Swine Fever. In Infectious Diseases of Livestock with special reference to Southern Africa; Coetzer, J.A.W., Thomson, G.R., Tustin, R.C., Eds.; Oxford Press: Oxford, UK, 1994; Volume 1, pp. 568–599. [Google Scholar]
- Montgomery, E.R. On A Form of Swine Fever Occurring in British East Africa (Kenya Colony). J. Comp. Pathol. Ther. 1921, 34, 159–191. [Google Scholar] [CrossRef]
- Walker, J. East African Swine Fever. Thesis (accepted), University of Zürich, Zürich, Switzerland, 1933. [Google Scholar]
- Kovalenko, J.R.; Sidorov, M.A.; Burba, L.G. Afrikanskaia chuma svinei. African swine fever. Unpublished manuscript translated from Russian to German. 1965. [Google Scholar]
- Steyn, D.G. East African Virus Disease in Pigs. 18th Report of the Director of Veterinary Services and Animal Industry, Onderstepoort, Pretoria/Union of South Africa, Department of Agriculture; Department of Agriculture: Pretoria, South Africa, 1932; pp. 99–109. [Google Scholar]
- Plowright, W.; Parker, J. The stability of African swine fever virus with particular reference to heat and pH inactivation. Arch. Gesamte Virusforsch. 1967, 21, 383–402. [Google Scholar] [CrossRef]
- De Kock, G.; Robinson, E.M.; Keppel, J.J.G.; DuToit, P.J. Swine fever in South Africa. Onderstepoort J. Vet. Sci. Anim. Ind. 1940, 14, 31–93. [Google Scholar]
- Blome, S.; Dietze, K. Report on the stability of African swine fever virus strain “Armenia 2008” in different diagnostic materials after storage at different ambient temperatures. Unpublished FAO Project Report. 2011. [Google Scholar]
- Mazur-Panasiuk, N.; Wozniakowski, G. Natural inactivation of African swine fever virus in tissues: Influence of temperature and environmental conditions on virus survival. Vet. Microbiol. 2020, 242, 108609. [Google Scholar] [CrossRef]
- Zani, L.; Masiulis, M.; Bušauskas, P.; Dietze, K.; Pridotkas, G.; Globig, A.; Blome, S.; Mettenleiter, T.; Depner, K.; Karvelienė, B. African swine fever virus survival in buried wild boar carcasses. Transbound. Emerg. Dis. 2020, 67, 2086–2092. [Google Scholar] [CrossRef]
- Davies, K.; Goatley, L.C.; Guinat, C.; Netherton, C.L.; Gubbins, S.; Dixon, L.K.; Reis, A.L. Survival of African Swine Fever Virus in Excretions from Pigs Experimentally Infected with the Georgia 2007/1 Isolate. Transbound. Emerg. Dis. 2017, 64, 425–431. [Google Scholar] [CrossRef]
- Fischer, M.; Mohnke, M.; Probst, C.; Pikalo, J.; Conraths, F.J.; Beer, M.; Blome, S. Stability of African swine fever virus on heat-treated field crops. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Turner, C.; Williams, S.M. Laboratory-scale inactivation of African swine fever virus and swine vesicular disease virus in pig slurry. J. Appl. Microbiol. 1999, 87, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Krug, P.W.; Larson, C.R.; Eslami, A.C.; Rodriguez, L.L. Disinfection of foot-and-mouth disease and African swine fever viruses with citric acid and sodium hypochlorite on birch wood carriers. Vet. Microbiol. 2012, 156, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Michels, I.; Menge, C. Desinfektionsverfahren bei den einzelnen Tierseuchen: Afrikanische Schweinepest. In Empfehlungen zur Desinfektion bei Tierseuchen; Friedrich-Loeffler-Inst.: Greifswald, Germany, 2020; Available online: https://www.openagrar.de/receive/openagrar_mods_00054329 (accessed on 31 August 2020).
- de Carvalho Ferreira, H.C.; Weesendorp, E.; Quak, S.; Stegeman, J.A.; Loeffen, W.L. Suitability of faeces and tissue samples as a basis for non-invasive sampling for African swine fever in wild boar. Vet. Microbiol. 2014, 172, 449–454. [Google Scholar] [CrossRef] [PubMed]
- McKercher, P.D.; Hess, W.R.; Hamdy, F. Residual viruses in pork products. Appl. Environ. Microbiol. 1978, 35, 142–145. [Google Scholar] [CrossRef]
- Zani, L.; Forth, J.H.; Forth, L.; Nurmoja, I.; Leidenberger, S.; Henke, J.; Carlson, J.; Breidenstein, C.; Viltrop, A.; Höper, D.; et al. Deletion at the 5’-end of Estonian ASFV strains associated with an attenuated phenotype. Sci. Rep. 2018, 8, 6510. [Google Scholar] [CrossRef]
- Carter, D.O.; Yellowlees, D.; Tibbett, M. Cadaver decomposition in terrestrial ecosystems. Naturwissenschaften 2007, 94, 12–24. [Google Scholar] [CrossRef]
- Carlson, J.; Fischer, M.; Zani, L.; Eschbaumer, M.; Fuchs, W.; Mettenleiter, T.; Beer, M.; Blome, S. Stability of African swine fever virus in soil and options to mitigate the potential transmission risk. Authorea 2020. [Google Scholar] [CrossRef]
- King, D.P.; Reid, S.M.; Hutchings, G.H.; Grierson, S.S.; Wilkinson, P.J.; Dixon, L.K.; Bastos, A.D.; Drew, T.W. Development of a TaqMan PCR assay with internal amplification control for the detection of African swine fever virus. J. Virol. Methods 2003, 107, 53–61. [Google Scholar] [CrossRef]
- Hoffmann, B.; Depner, K.; Schirrmeier, H.; Beer, M. A universal heterologous internal control system for duplex real-time RT-PCR assays used in a detection system for pestiviruses. J. Virol. Methods 2006, 136, 200–209. [Google Scholar] [CrossRef]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Benninger, L.A.; Carter, D.O.; Forbes, S.L. The biochemical alteration of soil beneath a decomposing carcass. Forensic Sci. Int. 2008, 180, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Larizza, M. Physical and chemical analysis of pig carcass decomposition in a fine sand. Master of Science Thesis, University of Ontario, Oshawa, ON, Canada, 2010. Available online: http://hdl.handle.net/10155/115 (accessed on 31 August 2020).
- Heaton, V.; Moffatt, C.; Simmons, T. Quantifying the Temperature of Maggot Masses and its Relationship to Decomposition. J. Forensic Sci. 2014, 59. [Google Scholar] [CrossRef]
- Probst, C.; Gethmann, J.; Amendt, J.; Lutz, L.; Teifke, J.; Conraths, F. Estimating the Postmortem Interval of Wild Boar Carcasses. Vet. Sci. 2020, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Probst, C.; Gethmann, J.; Hohmann, U.; Knoll, B.; Amendt, J.; Teifke, J.P.; Conraths, F.J. Zersetzungsstadien bei Wildschweinkadavern — und wie die Liegezeit geschätzt werden kann. Amtstierärztlicher Dienst. Lebensm. 2020, 2, 85–94. [Google Scholar]
- Gallardo, C.; Nieto, R.; Soler, A.; Pelayo, V.; Fernández-Pinero, J.; Markowska-Daniel, I.; Pridotkas, G.; Nurmoja, I.; Granta, R.; Simón, A.; et al. Assessment of African Swine Fever Diagnostic Techniques as a Response to the Epidemic Outbreaks in Eastern European Union Countries: How To Improve Surveillance and Control Programs. J. Clin. Microbiol. 2015, 53, 2555–2565. [Google Scholar] [CrossRef]
- Rai, A.; Pruitt, S.; Ramirez-Medina, E.; Vuono, E.A.; Silva, E.; Velazquez-Salinas, L.; Carrillo, C.; Borca, M.V.; Gladue, D.P. Identification of a Continuously Stable and Commercially Available Cell Line for the Identification of Infectious African Swine Fever Virus in Clinical Samples. Viruses 2020, 12, 820. [Google Scholar] [CrossRef] [PubMed]
- OIE. Chapter 3.8.1—African swine fever (infection with African swine fever virus). OIE Terr. Man. 2019. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.08.01_ASF.pdf (accessed on 31 August 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Material | ASFV Stability | Method |
|---|---|---|---|
| Blood | |||
| Montgomery 1921 [27] | Defibrinated blood (RT) | 140 days | In vivo assay |
| Preserved blood (RT) | 404 days | ||
| Preserved blood (37 °C) | 104 h | ||
| Preserved blood (40 °C) | 15 days | ||
| Preserved blood (50 °C) | 4 h | ||
| Preserved blood (55 °C) | 1 h | ||
| Filtered serum (RT) | 428 days | ||
| Steyn 1932 [30] | Preserved blood in ice chests | 12 months | In vivo assay |
| Clotted blood from decomposed carcass | 5 days | ||
| Walker 1933 [28] | Preserved blood (1–4 °C) | 1076 days | In vivo assay |
| De Kock et al., 1940 [32] | Blood stored “in a cold room in the dark” | 6 years | In vivo assay |
| Kovalenko et al., 1965 [29] | Dried blood | 2165 days | |
| Native blood in sealed ampules | 1526 days | ||
| Frozen meat (3–4 °C) | 104 days | ||
| Kovalenko et al., 1965 [29] | Fresh blood on wood, buried | 81 days | In vivo assay |
| Fresh blood on wood, surface | 6 months | ||
| Fresh blood on bricks, buried | 112 days | ||
| Clotted blood buried in garden soil | 112 days | ||
| Clotted blood buried in sandy forest | 81 days | ||
| Clotted blood in pond water | 70 days | ||
| Plowright and Parker 1967 [31] | Preserved blood (4 °C) | 18 months | In vivo assay |
| Blome and Dietze 2011 [33] | Blood (4 °C, 18–22 °C, 37 °C) | 3 months | Virus isolation |
| Organs | |||
| Kovalenko et al., 1965 [29] | Spleen buried in soil | 280 days | In vivo assay |
| Plowright and Parker 1967 [31] | Spleen suspension (−70 °C) | 82–105 weeks | In vivo assay |
| Spleen suspension (−20 °C) | marked instability | ||
| Blome and Dietze 2011 [33] | Spleen (4 °C, 18–22 °C, 37 °C) | 3 months | Virus isolation |
| de Carvalho Ferreira et al., 2014 [41] | Spleen, liver, lymph nodes, tonsil | Half-life: 4–80/2.7–7.4/1.8–4.6/1.7–3.8 days | PCR |
| Mazur-Panasiuk and Woźniakowski 2020 [34] | Spleen (−20 °C, 4 °C, 23 °C) | Predicted survival time: 483/126/15 days | PCR |
| Putrescent spleen in water/soil/leaf litter/straw/hay/grain (4 °C) | Predicted survival time: 36/33/19/97/113/69 days | ||
| Lung (−20 °C, 4 °C, 23 °C) | Predicted survival time: 714/136/9 days | ||
| Kidney (−20 °C, 4 °C, 23 °C) | Predicted survival time: 353/35/17 days | ||
| Muscle | |||
| Blome and Dietze 2011 [33] | Muscle (37 °C) | 3 months | PCR, virus isolation |
| Muscle (4 °C) | 2 months | ||
| Urine | |||
| Kovalenko et al., 1965 [29] | Urine (4 °C) | 60 days | In vivo assay |
| Davies et al., 2017 [36] | Urine (4° C, 12 °C, 21 °C, 37 °C) | Estimated survival: 15.3/7.5/4.8/2.9 days | PCR, virus isolation |
| Faeces | |||
| Kovalenko et al., 1965 [29] | Faeces (4 °C) | 160 days | In vivo assay |
| Blome and Dietze 2011 [33] | Faeces (18–22 °C) | 3 months | Virus isolation |
| de Carvalho Ferreira et al., 2014 [41] | Faeces (5 °C, 12 °C, 20 °C, 30 °C) | Half-life: 1568/660/21.7/15.3 days | PCR |
| Davies et al., 2017 [36] | Faeces (4 °C, 12 °C, 21 °C, 37 °C) | Estimated survival: 8.5/6.5/5.1/3.7 days | PCR, virus isolation |
| Pork Products | |||
| McKercher et al., 1978 [42] | Pepperoni sausage | 8 days | In vivo assay |
| McKercher et al., 1987 [6] | Muscle of Parma ham | 183 days | Virus isolation (HAT) |
| Bone marrow of Parma ham | 94 days | ||
| Fat of ham | 183 days | ||
| Pooled muscle, bone marrow, fat of ham | 291 days | In vivo assay | |
| Mebus et al., 1993 [8] | Muscle of dry cured Iberian ham/shoulder/loin/dry cured Serrano ham | 112/112/98/112 days | In vivo assay |
| Fat of dry cured Iberian ham/shoulder/dry cured Serrano ham | 112/84/112 days | ||
| Bone marrow of dry cured Iberian ham/shoulder/dry cured Serrano ham | 112/84/112 days | ||
| Lymph node of dry cured Iberian ham/dry cured Serrano ham | 112/56 days | ||
| Kolbasov et al., 2011 [5] | Meat and fat (22–27 °C, salted) | 16 days | PCR |
| Pork products (4–6 °C) | 84 days | ||
| Pork products (−18–20 °C) | 118 days | ||
| Petrini et al., 2019 [7] | Italian salami | 18 days | In vivo assay |
| Pork belly | 60 days | ||
| Loin | 83 days | ||
| Sample | ASFV Genome Copies (log10) | |||||
|---|---|---|---|---|---|---|
| DP1 | DP2 | DP3 | WB1 | WB2 | WB3 | |
| Blood | 4.73 | 5.98 | 4.40 | 5.50 | 6.98 | 6.50 |
| Bone marrow | 4.17 | 3.27 | 3.34 | 3.17 | 3.95 | 3.04 |
| Muscle | 3.10 | 3.31 | 2.90 | 3.92 | 3.79 | 3.36 |
| Skin | 3.93 | 3.47 | 3.37 | 4.51 | 4.49 | 3.95 |
| Spleen | 4.91 | 4.79 | 6.10 | 5.87 | 5.79 | 5.26 |
| Faeces | 1.97 | 1.88 | neg. | 3.62 | 3.47 | 2.18 |
| Urine | 3.07 | 2.47 | neg. | 3.38 | 3.91 | 2.30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fischer, M.; Hühr, J.; Blome, S.; Conraths, F.J.; Probst, C. Stability of African Swine Fever Virus in Carcasses of Domestic Pigs and Wild Boar Experimentally Infected with the ASFV “Estonia 2014” Isolate. Viruses 2020, 12, 1118. https://doi.org/10.3390/v12101118
Fischer M, Hühr J, Blome S, Conraths FJ, Probst C. Stability of African Swine Fever Virus in Carcasses of Domestic Pigs and Wild Boar Experimentally Infected with the ASFV “Estonia 2014” Isolate. Viruses. 2020; 12(10):1118. https://doi.org/10.3390/v12101118
Chicago/Turabian StyleFischer, Melina, Jane Hühr, Sandra Blome, Franz J. Conraths, and Carolina Probst. 2020. "Stability of African Swine Fever Virus in Carcasses of Domestic Pigs and Wild Boar Experimentally Infected with the ASFV “Estonia 2014” Isolate" Viruses 12, no. 10: 1118. https://doi.org/10.3390/v12101118
APA StyleFischer, M., Hühr, J., Blome, S., Conraths, F. J., & Probst, C. (2020). Stability of African Swine Fever Virus in Carcasses of Domestic Pigs and Wild Boar Experimentally Infected with the ASFV “Estonia 2014” Isolate. Viruses, 12(10), 1118. https://doi.org/10.3390/v12101118