A Sporadic and Lethal Lassa Fever Case in Forest Guinea, 2019

 ,
, {kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
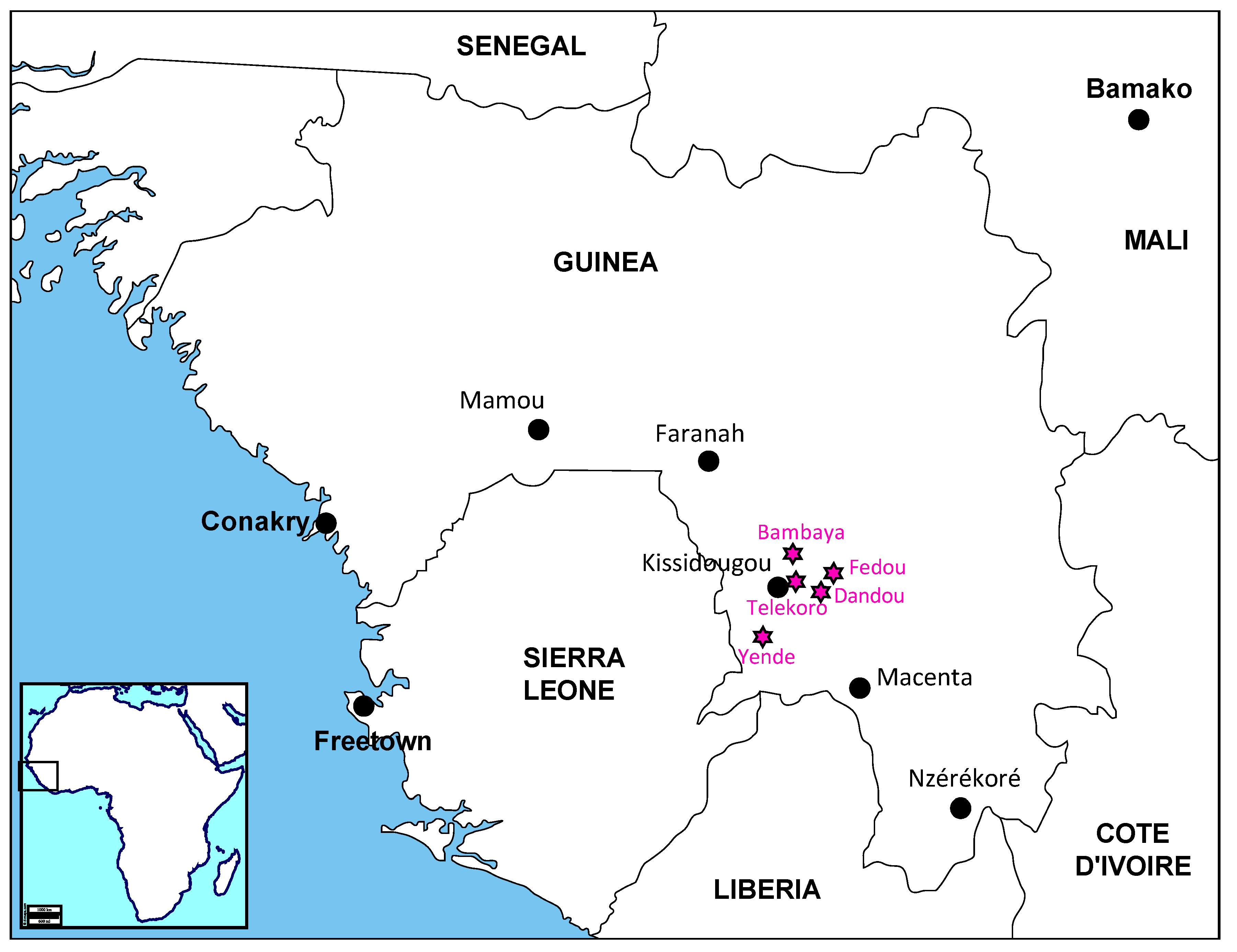
2.1. Field Investigation
2.2. Molecular and Serological Diagnosis
2.3. LASV Sequencing
2.4. Phylogenetic Analysis
3. Results
3.1. The Case Description
3.2. Laboratory Diagnosis, Field Investigation and Contact Tracing
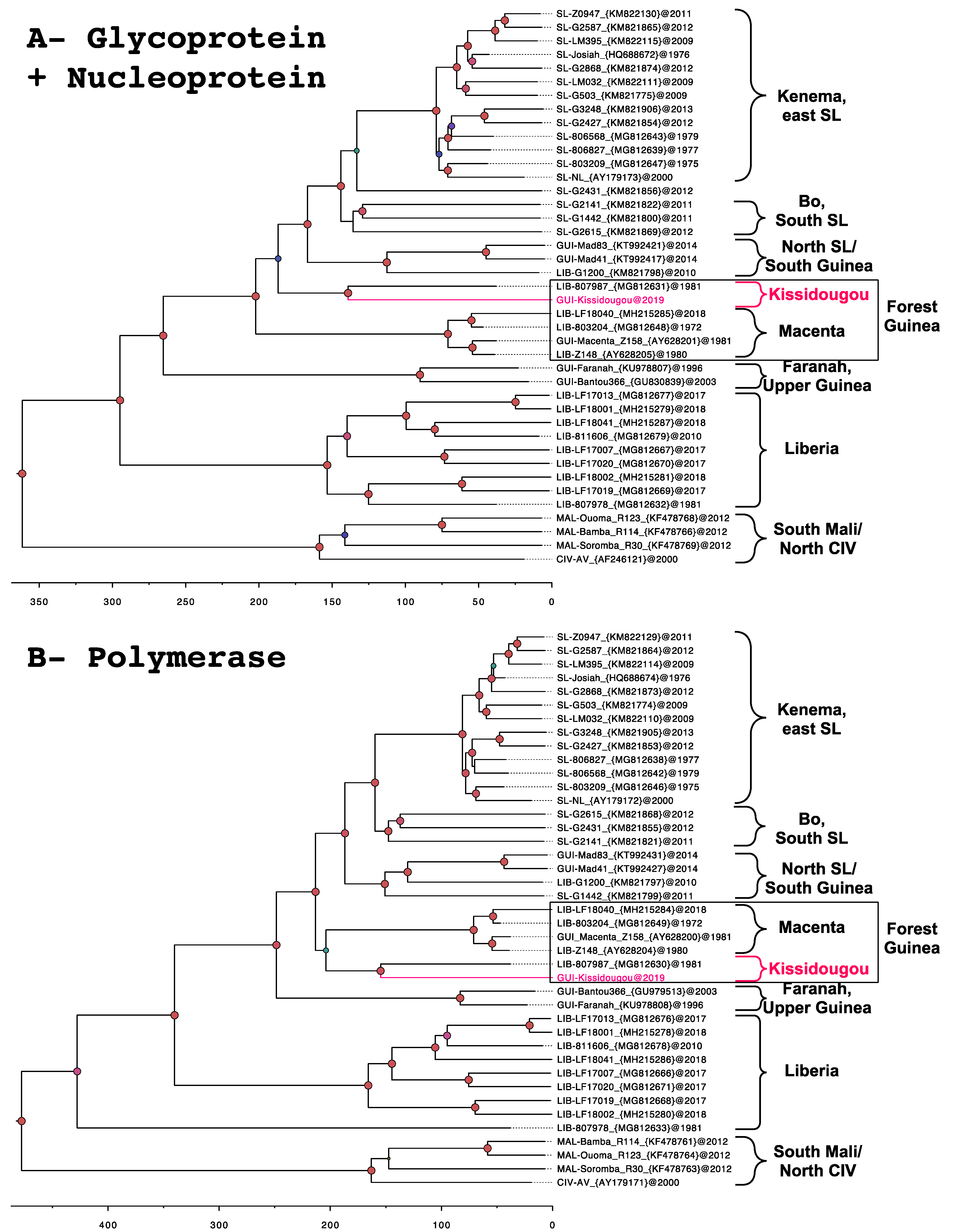
3.3. Phylogeny
4. Discussion
4.1. Identification of the Kissidougou Sub-Lineage
4.2. Strengths and Weaknesses of the Guinean Health System
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buckley, S.M.; Casals, J.; Downs, W.G. Isolation and antigenic characterization of Lassa virus. Nature 1970, 227, 174. [Google Scholar] [CrossRef]
- Frame, J.D.; Baldwin, J.M.J.; Gocke, D.J.; Troup, J.M. Lassa fever, a new virus disease of man from West Africa. Am. J. Trop. Med. Hyg. 1970, 19, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Fichet-Calvet, E.; Rogers, D.J. Risk maps of Lassa fever in West Africa. PLoS Negl. Trop. Dis. 2009, 3, e388. [Google Scholar] [CrossRef] [PubMed]
- Gunther, S.; Lenz, O. Lassa virus. Crit. Rev. Clin. Lab. Sci. 2004, 41, 339–390. [Google Scholar] [CrossRef] [PubMed]
- Kafetzopoulou, L.E.; Pullan, S.T.; Lemey, P.; Suchard, M.A.; Ehichioya, D.U.; Pahlmann, M.; Thielebein, A.; Hinzmann, J.; Oestereich, L.; Wozniak, D.M.; et al. Metagenomic sequencing at the epicenter of the Nigeria 2018 Lassa fever outbreak. Science 2019, 363, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Lukashevich, I.S.; Clegg, J.C.; Sidibe, K. Lassa virus activity in Guinea: Distribution of human antiviral antibody defined using enzyme-linked immunosorbent assay with recombinant antigen. J. Med. Virol. 1993, 40, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Fair, J.; Jentes, E.; Inapogui, A.; Kourouma, K.; Goba, A.; Bah, A.; Tounkara, M.; Coulibaly, M.; Garry, R.F.; Bausch, D.G. Lassa virus-infected rodents in refugee camps in Guinea: A looming threat to public health in a politically unstable region. Vector Borne Zoonotic Dis. 2007, 7, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Fichet-Calvet, E.; Lecompte, E.; Koivogui, L.; Soropogui, B.; Dore, A.; Kourouma, F.; Sylla, O.; Daffis, S.; Koulemou, K.; Ter Meulen, J. Fluctuation of abundance and Lassa virus prevalence in Mastomys natalensis in Guinea, West Africa. Vector Borne Zoonotic Dis. 2007, 7, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Fichet-Calvet, E.; Olschlager, S.; Strecker, T.; Koivogui, L.; Becker-Ziaja, B.; Camara, A.B.; Soropogui, B.; Magassouba, N.; Gunther, S. Spatial and temporal evolution of Lassa virus in the natural host population in Upper Guinea. Sci. Rep. 2016, 6, 21977. [Google Scholar] [CrossRef] [PubMed]
- Lecompte, E.; Fichet-Calvet, E.; Daffis, S.; Koulemou, K.; Sylla, O.; Kourouma, F.; Dore, A.; Soropogui, B.; Aniskin, V.; Allali, B.; et al. Mastomys natalensis and Lassa fever, West Africa. Emerg. Infect. Dis. 2006, 12, 1971–1974. [Google Scholar] [CrossRef] [PubMed]
- Olayemi, A.; Cadar, D.; Magassouba, N.; Obadare, A.; Kourouma, F.; Oyeyiola, A.; Fasogbon, S.; Igbokwe, J.; Rieger, T.; Bockholt, S.; et al. New Hosts of The Lassa Virus. Sci. Rep. 2016, 6, 25280. [Google Scholar] [CrossRef] [PubMed]
- Bausch, D.G.; Demby, A.H.; Coulibaly, M.; Kanu, J.; Goba, A.; Bah, A.; Condé, N.; Wurtzel, H.L.; Cavallaro, K.F.; Lloyd, E.; et al. Lassa fever in Guinea: I. Epidemiology of human disease and clinical observations. Vector Borne Zoonotic Dis. 2001, 1, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Boiro, I.; Lomonossov, N.N.; Sotsinski, V.A.; Constantinov, O.K.; Tkachenko, E.A.; Inapogui, A.P.; Balde, C. Eléments de recherches clinico-épidémiologiques et de laboratoire sur les fièvres hémorragiques en Guinée. B. Soc. Pathol. Exot. 1987, 80, 607–612. [Google Scholar]
- Bowen, M.D.; Rollin, P.E.; Ksiazek, T.G.; Hustad, H.L.; Bausch, D.G.; Demby, A.H.; Bajani, M.D.; Peters, C.J.; Nichol, S.T. Genetic diversity among Lassa virus strains. J. Virol. 2000, 74, 6992–7004. [Google Scholar] [CrossRef] [PubMed]
- ProMED-mail. Lassa fever-West Africa (09): Liberia ex Guinea. Available online: https://promedmail.org/promed-post/?id=20180210.5620420 (accessed on 10 February 2018).
- ProMED-mail. Lassa fever-West Africa (08): Guinea (MM). Available online: https://promedmail.org/promed-post/?id=6292809 (accessed on 3 February 2019).
- Henderson, B.E.; Gary, G.W.; Kissling, R.E.; Frame, J.D.; Carey, D.E. Lassa fever virological and serological studies. Trans. Roy. Soc. Trop. Med. Hyg. 1972, 66, 409–416. [Google Scholar] [CrossRef]
- Olschlager, S.; Lelke, M.; Emmerich, P.; Panning, M.; Drosten, C.; Hass, M.; Asogun, D.; Ehichioya, D.; Omilabu, S.; Gunther, S. Improved detection of Lassa virus by reverse transcription-PCR targeting the 5’ region of S RNA. J. Clin. Microbiol. 2010, 48, 2009–2013. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed]
- Marien, J.; Lo Iacono, G.; Rieger, T.; Magassouba, N.; Gunther, S.; Fichet-Calvet, E. Households as hotspots of Lassa fever? Assessing the spatial distribution of Lassa virus-infected rodents in rural villages of Guinea. Emerg. Microbes Infect. 2020, 9, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Olayemi, A.; Adesina, A.S.; Strecker, T.; Magassouba, N.; Fichet-Calvet, E. Determining Ancestry between Rodent- and Human-Derived Virus Sequences in Endemic Foci: Towards a More Integral Molecular Epidemiology of Lassa Fever within West Africa. Biology 2020, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Jahrling, P.B.; Geisbert, J.; Ibrahim, M.S. Direct Submission Lassa mammarenavirus. Available online: https://www.ncbi.nlm.nih.gov/nuccore/AY628201 (accessed on 18 May 2004).
- Rossi, S.L.; Guerbois, M.; Forrester, N.L.; Ksiazek, T.; Lin, D.; Hari, K.; Weaver, S.C. Direct Submission Lassa mammarenavirus. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KU978807 (accessed on 24 March 2016).
- Wiley, M.R.; Fakoli, L.; Letizia, A.G.; Welch, S.R.; Ladner, J.T.; Prieto, K.; Reyes, D.; Espy, N.; Chitty, J.A.; Pratt, C.B.; et al. Lassa virus circulating in Liberia: A retrospective genomic characterisation. Lancet Infect. Dis. 2019, 19, 1371–1378. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magassouba, N.; Koivogui, E.; Conde, S.; Kone, M.; Koropogui, M.; Soropogui, B.; Kekoura, I.; Hinzmann, J.; Günther, S.; Keita, S.; et al. A Sporadic and Lethal Lassa Fever Case in Forest Guinea, 2019. Viruses 2020, 12, 1062. https://doi.org/10.3390/v12101062
Magassouba N, Koivogui E, Conde S, Kone M, Koropogui M, Soropogui B, Kekoura I, Hinzmann J, Günther S, Keita S, et al. A Sporadic and Lethal Lassa Fever Case in Forest Guinea, 2019. Viruses. 2020; 12(10):1062. https://doi.org/10.3390/v12101062
Chicago/Turabian StyleMagassouba, N’Faly, Enogo Koivogui, Sory Conde, Moussa Kone, Michel Koropogui, Barrè Soropogui, Ifono Kekoura, Julia Hinzmann, Stephan Günther, Sakoba Keita, and et al. 2020. "A Sporadic and Lethal Lassa Fever Case in Forest Guinea, 2019" Viruses 12, no. 10: 1062. https://doi.org/10.3390/v12101062
APA StyleMagassouba, N., Koivogui, E., Conde, S., Kone, M., Koropogui, M., Soropogui, B., Kekoura, I., Hinzmann, J., Günther, S., Keita, S., Duraffour, S., & Fichet-Calvet, E. (2020). A Sporadic and Lethal Lassa Fever Case in Forest Guinea, 2019. Viruses, 12(10), 1062. https://doi.org/10.3390/v12101062