A 19 Year Analysis of Small Mammals Associated with Human Hantavirus Cases in Chile

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
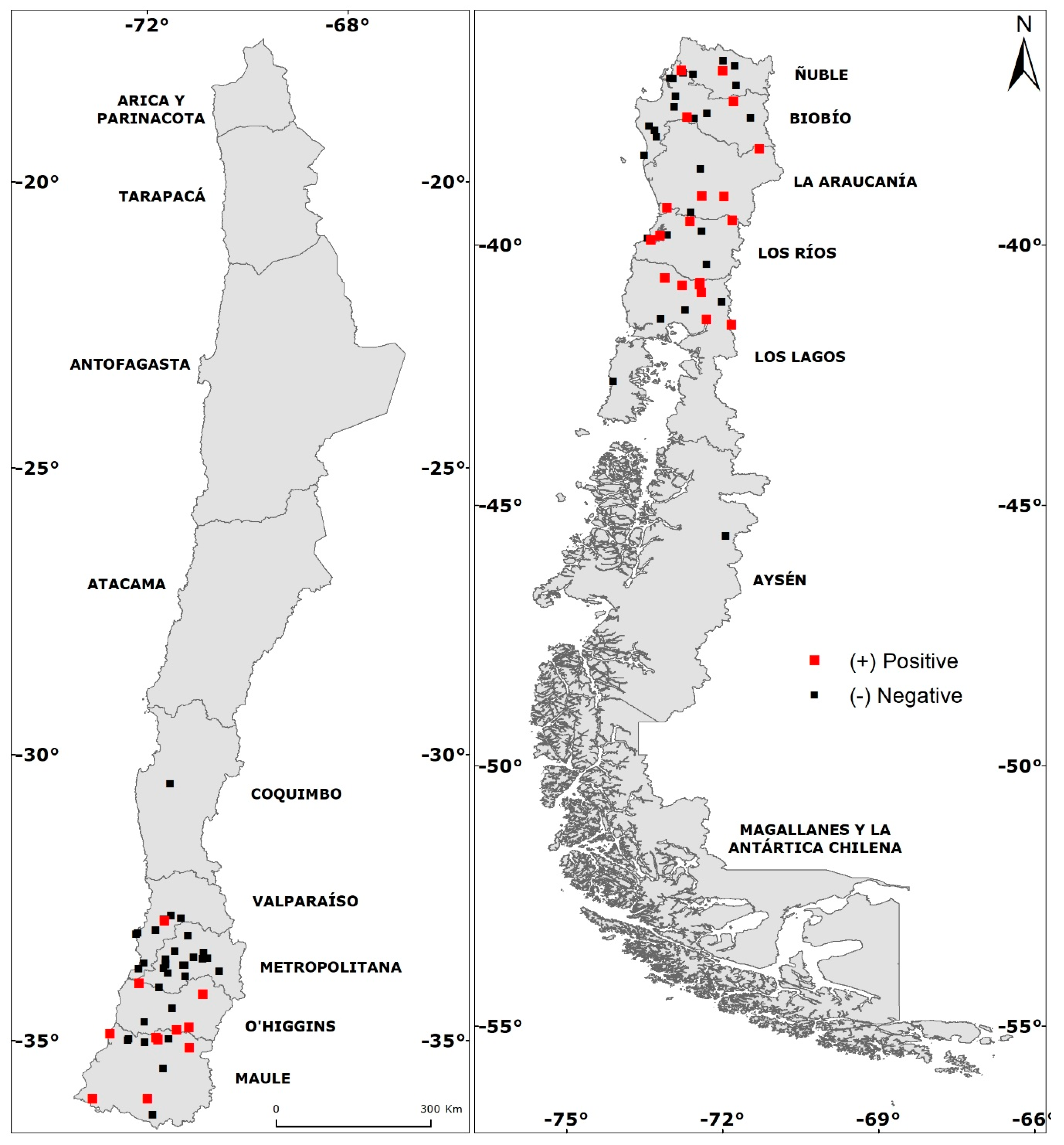
2.2. Sampling
2.3. Relative Abundance
2.4. Relative Seropositivity
2.5. Confirmed Human Cases of Hantavirus
2.6. Ecological Features of the Reservoirs
2.7. Data Analysis and Statistics
2.8. Serology and Human Hantavirus Cases
3. Results
3.1. Rodents Relative Abundance
3.2. Total Seropositivity
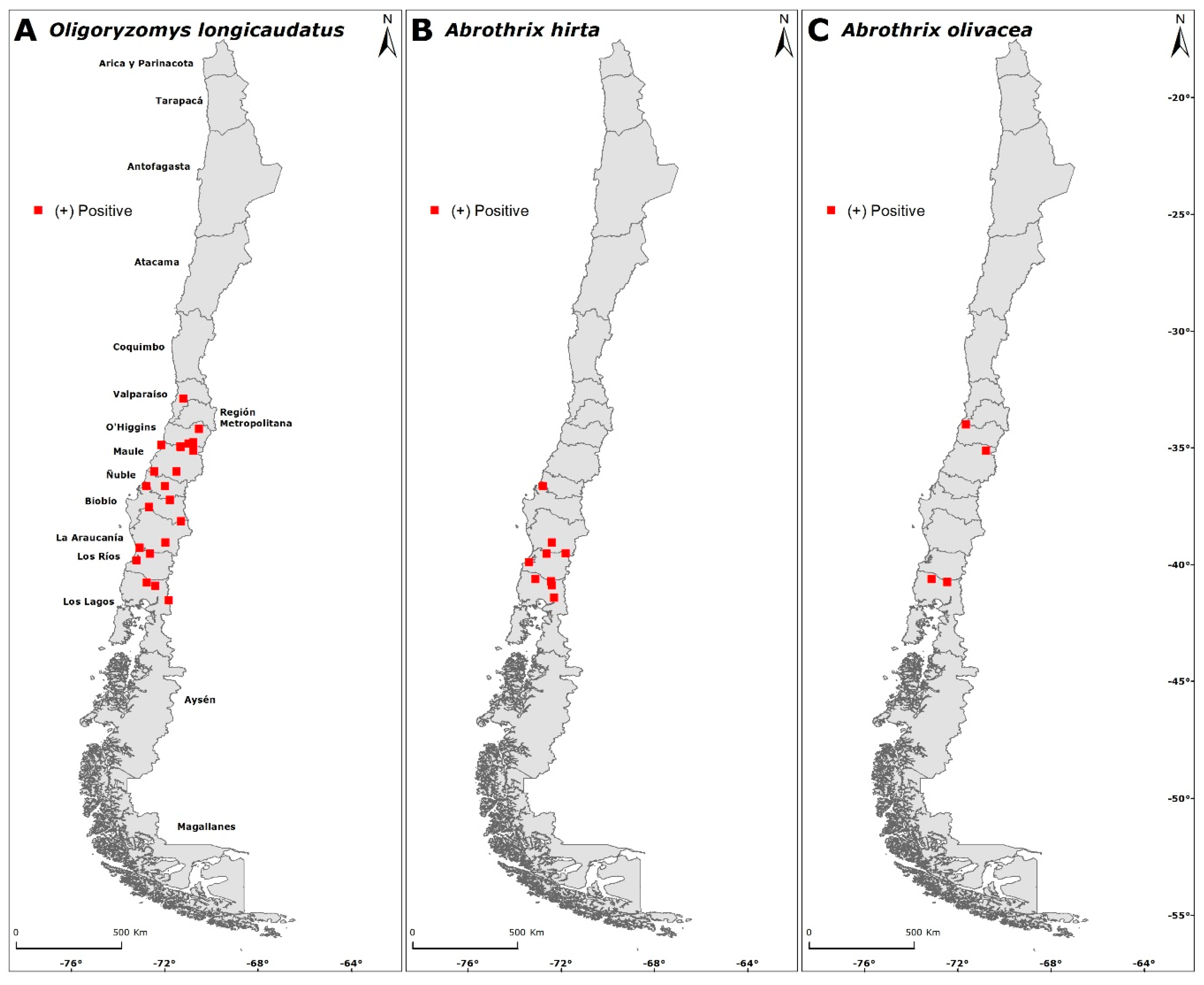
3.3. Intraspecific Seropositivity
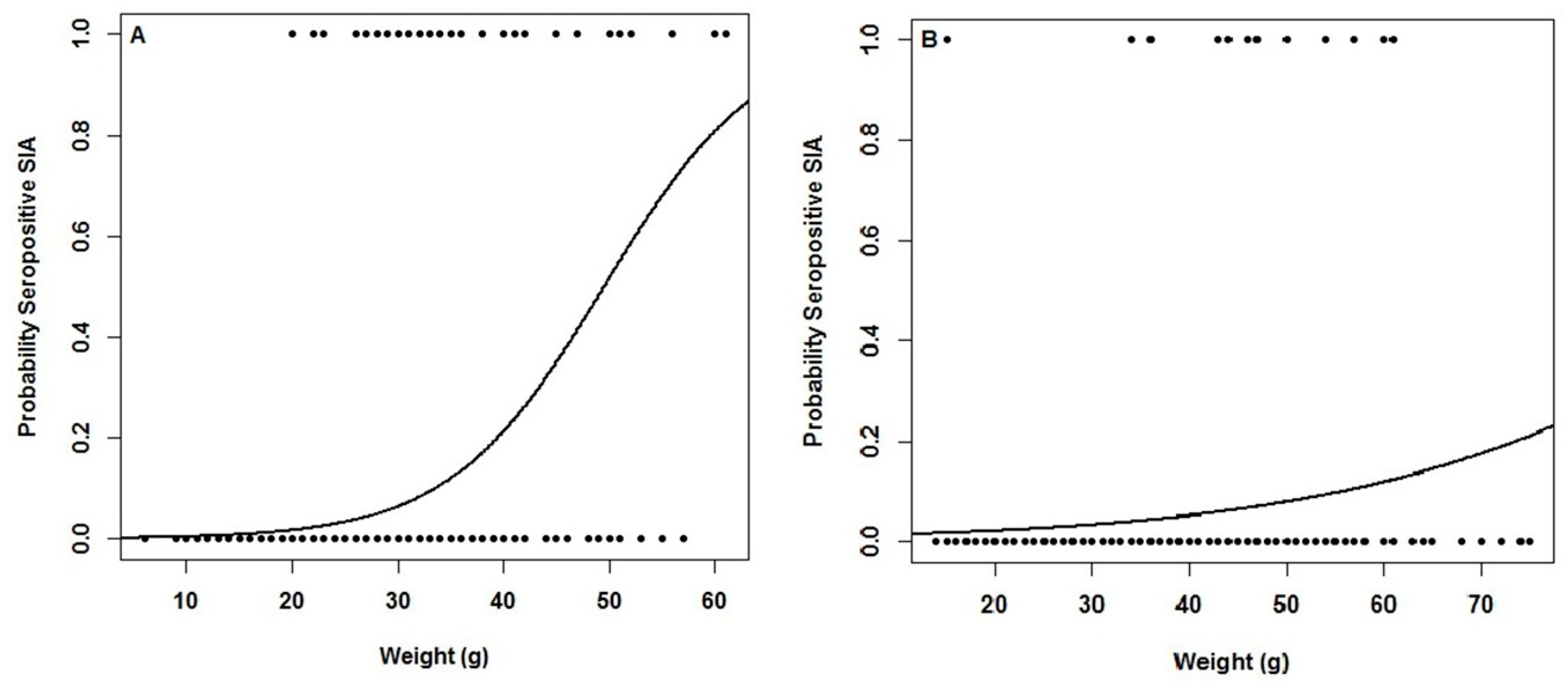
3.4. Association Between Seropositives and Biological Traits
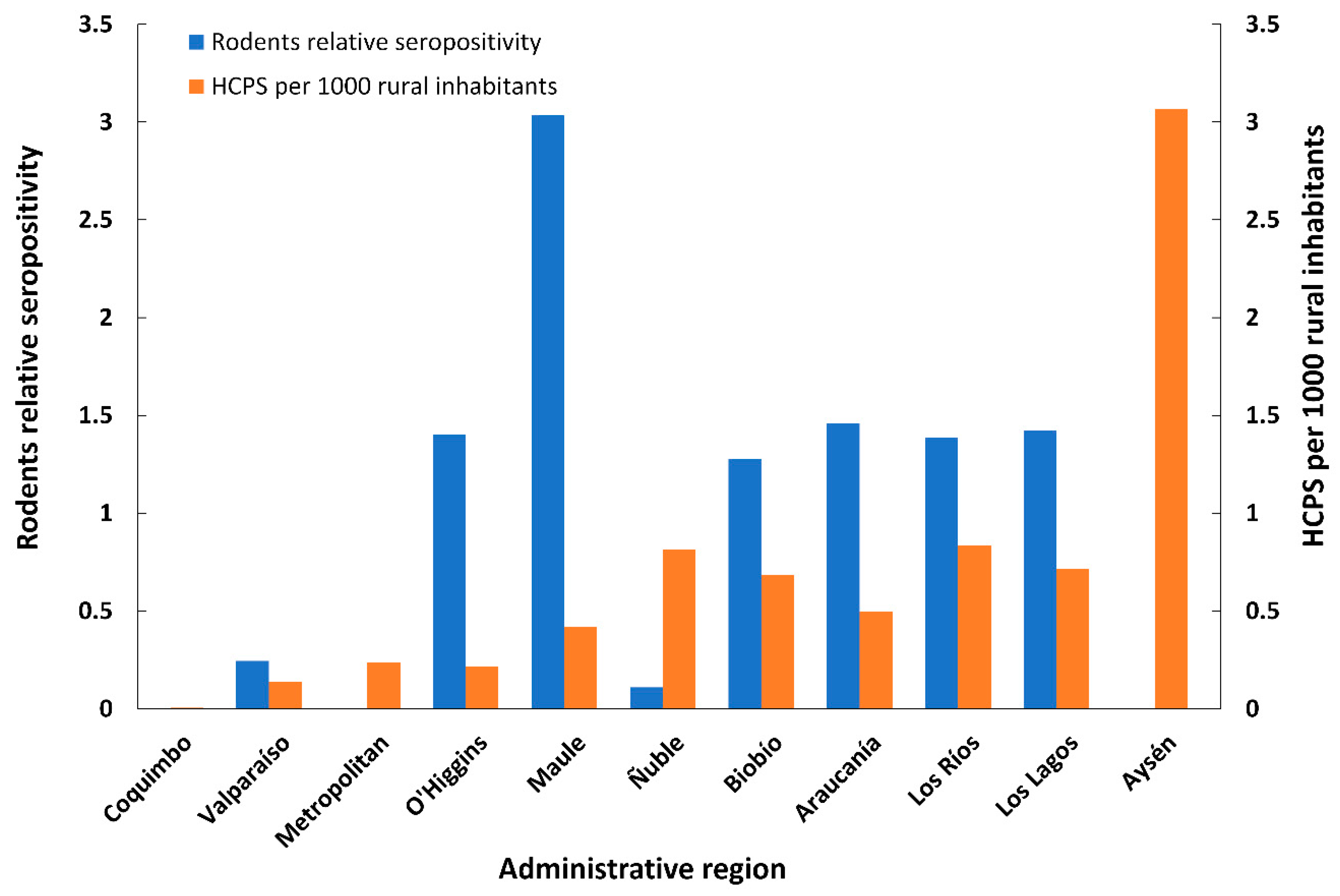
3.5. Human HCPS Cases
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Abundance | Seropositives | Relative Seropositivity | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Locality, County | Latitude | Longitude | Administrative Region | Total Traps | Year | Al | Ah | Ao | As | Am | Lm | Ol | Pd | Mm | Rn | Rr | Ol | Ah | Ao | Ol | Ah | Ao | Total |
| Chiñigue, Ovalle | −30.513 | −71.102 | Coquimbo | 300 | 2006 | 1.00 | 0.00 | 2.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.67 | 0.67 | 0.33 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fundo Chuico Blanco, Hijuelas | −32.812 | −71.089 | Valparaíso | 270 | 2002 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.37 | 0.00 | 0.00 | 1.11 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Llaillay, Llaillay | −32.854 | −70.918 | Valparaíso | 237 | 2001 | 0.00 | 0.00 | 0.42 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.53 | 0.42 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lo Mardones, Quillota | −32.870 | −71.221 | Valparaíso | 630 | 2016 | 0.00 | 0.00 | 2.06 | 0.00 | 0.00 | 0.00 | 0.48 | 0.00 | 0.63 | 0.95 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| La Palma, Quillota | −32.908 | −71.206 | Valparaíso | 600 | 2013 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.83 | 0.17 | 1 | 0 | 0 | 3.00 | 0.00 | 0.00 | 0.60 |
| Población Prat, Villa Alemana | −33.070 | −71.357 | Valparaíso | 300 | 2001 | 0.00 | 0.00 | 4.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.33 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Laguna Verde, Valparaíso | −33.115 | −71.666 | Valparaíso | 360 | 2006 | 0.83 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Las Docas, Valparaíso | −33.138 | −71.703 | Valparaíso | 495 | 2018 | 3.23 | 0.00 | 0.40 | 0.00 | 0.00 | 0.00 | 1.41 | 1.01 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Alto el Manzano, TilTil | −33.165 | −70.790 | Metropolitan | 300 | 2010 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.33 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| San Antonio de Naltahua, Isla de Maipo | −33.436 | −71.021 | Metropolitan | 300 | 2004 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.67 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| La Reina, La Reina | −33.456 | −70.517 | Metropolitan | 150 | 2003 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lo Espejo, Lo Espejo | −33.543 | −70.696 | Metropolitan | 600 | 2013 | 0.00 | 0.00 | 0.50 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.17 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| El Canelo, San José de Maipo | −33.556 | −70.454 | Metropolitan | 255 | 2004 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.39 | 0.39 | 0.00 | 0.39 | 0.78 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| La Florida Alto, La Florida | −33.564 | −70.532 | Metropolitan | 270 | 2005 | 0.37 | 0.00 | 3.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Mallarauco, Melipilla | −33.569 | −71.180 | Metropolitan | 300 | 2012 | 0.00 | 0.00 | 1.67 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 2.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| San Antonio, San Antonio | −33.643 | −71.563 | Valparaíso | 225 | 2001 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.44 | 0.00 | 1.33 | 0.00 | 0.89 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Pomaire, Melipilla | −33.650 | −71.179 | Metropolitan | 450 | 2006 | 0.00 | 0.00 | 0.89 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.22 | 0.22 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Constructora Inca, Melipilla | −33.673 | −71.180 | Metropolitan | 312 | 2013 | 0.00 | 0.00 | 2.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.32 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Condominio Puerta del Sol, Talagante | −33.677 | −70.845 | Metropolitan | 300 | 2006 | 0.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Talagante, Talagante | −33.681 | −70.877 | Metropolitan | 240 | 2001 | 0.00 | 0.00 | 6.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.42 | 2.08 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Chocalán, Melipilla | −33.735 | −71.217 | Metropolitan | 300 | 2003 | 0.00 | 0.00 | 3.67 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.67 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fundo La Ventolera, Santo Domingo | −33.743 | −71.652 | Valparaíso | 300 | 2002 | 4.00 | 0.00 | 6.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.67 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| El Ingenio, San José de Maipo | −33.786 | −70.248 | Metropolitan | 300 | 2002 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Hijuelas, Hijuelas | −33.813 | −71.147 | Valparaíso | 210 | 2001 | 0.00 | 0.00 | 0.95 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.95 | 0.48 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Abrantes, Paine | −33.868 | −70.837 | Metropolitan | 300 | 2004 | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.33 | 1.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ucúquer, Litueche | −33.998 | −71.650 | O’Higgins | 450 | 2013 | 0.00 | 0.00 | 3.11 | 0.00 | 0.00 | 0.00 | 1.33 | 1.33 | 0.00 | 0.00 | 1.11 | 0 | 0 | 1 | 0.00 | 0.00 | 0.32 | 0.15 |
| Quilamuta-Las Cabras, Las Cabras | −34.070 | −71.300 | O’Higgins | 300 | 2002 | 0.33 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 2.67 | 0.00 | 0.00 | 0.67 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Coya, Machalí | −34.190 | −70.534 | O’Higgins | 145 | 2002 | 0.00 | 0.00 | 0.69 | 0.00 | 0.00 | 0.00 | 0.69 | 1.38 | 0.00 | 3.45 | 0.00 | 1 | 0 | 0 | 1.45 | 0.00 | 0.00 | 0.16 |
| San Vicente de Tagua Tagua, San Vicente de Tagua Tagua | −34.433 | −71.069 | O’Higgins | 714 | 2011 | 0.00 | 0.00 | 0.28 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.42 | 0.00 | 0.14 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Los Maitenes, Lolol | −34.671 | −71.554 | O’Higgins | 652 | 2017 | 0.00 | 0.00 | 0.46 | 0.00 | 0.00 | 0.00 | 0.31 | 0.00 | 0.15 | 0.31 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Las Peñas, San Fernando 2002 | −34.766 | −70.776 | O’Higgins | 360 | 2002 | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.28 | 0.00 | 0.00 | 0.00 | 0.00 | 1 | 0 | 0 | 3.60 | 0.00 | 0.00 | 1.20 |
| Las Peñas, San Fernando 2018 | −34.767 | −70.776 | O’Higgins | 600 | 2018 | 0.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.50 | 1.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| El Sauce, Chimbarongo | −34.815 | −70.994 | O’Higgins | 390 | 2012 | 0.00 | 0.00 | 2.31 | 0.00 | 0.00 | 0.00 | 0.77 | 0.00 | 0.00 | 0.00 | 0.26 | 1 | 0 | 0 | 1.30 | 0.00 | 0.00 | 0.30 |
| Lipimávida, Vichuquén | −34.870 | −72.147 | Maule | 450 | 2010 | 0.44 | 0.00 | 0.22 | 0.00 | 0.00 | 0.00 | 1.78 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Duao, Licantén | −34.882 | −72.154 | Maule | 300 | 2003 | 1.00 | 0.00 | 8.00 | 0.00 | 0.00 | 0.00 | 12.33 | 0.00 | 0.00 | 0.67 | 0.00 | 4 | 0 | 0 | 0.32 | 0.00 | 0.00 | 0.18 |
| Escuela Quilpoco, Rauco | −34.955 | −71.346 | Maule | 270 | 2018 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.37 | 0.00 | 0.37 | 1.11 | 0.00 | 1 | 0 | 0 | 2.70 | 0.00 | 0.00 | 0.54 |
| El Pumaitén, Romeral | −34.967 | −71.125 | Maule | 300 | 2002 | 0.67 | 0.00 | 3.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.67 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Hualañé, Hualañé 2010 | −34.971 | −71.833 | Maule | 450 | 2010 | 0.00 | 0.00 | 0.22 | 0.00 | 0.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| El Trapiche, Curicó | −34.983 | −71.317 | Maule | 600 | 2004 | 0.00 | 0.00 | 3.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.33 | 0.17 | 0.50 | 1 | 0 | 0 | 1.00 | 0.00 | 0.00 | 0.20 |
| Hualañé, Hualañé 2012 | −34.990 | −71.847 | Maule | 360 | 2012 | 0.00 | 0.00 | 3.06 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Palquibudi, Rauco | −35.030 | −71.549 | Maule | 270 | 2002 | 0.00 | 0.00 | 0.74 | 0.00 | 0.00 | 0.00 | 0.37 | 0.00 | 0.00 | 0.37 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Los Queñes, Romeral | −35.125 | −70.773 | Maule | 360 | 2012 | 0.00 | 0.00 | 0.28 | 0.00 | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.28 | 1.39 | 4 | 0 | 1 | 3.60 | 0.00 | 3.60 | 1.64 |
| Las Lomas, San Clemente | −35.484 | −71.227 | Maule | 300 | 2002 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.67 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Canal Melado, Longaví | −36.017 | −71.501 | Maule | 375 | 2002 | 0.00 | 0.27 | 1.87 | 0.00 | 0.00 | 0.00 | 0.80 | 0.00 | 0.00 | 0.00 | 0.00 | 1 | 0 | 0 | 1.25 | 0.00 | 0.00 | 0.34 |
| Retupel, Cauquenes | −36.018 | −72.462 | Maule | 300 | 2003 | 0.00 | 0.00 | 2.33 | 0.00 | 0.00 | 0.00 | 1.67 | 0.00 | 0.00 | 0.00 | 0.00 | 1 | 0 | 0 | 0.60 | 0.00 | 0.00 | 0.25 |
| Bullileo, Parral | −36.289 | −71.413 | Maule | 240 | 2001 | 0.00 | 0.42 | 0.00 | 0.00 | 0.00 | 0.00 | 1.25 | 0.00 | 0.00 | 0.42 | 1.25 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| San Miguel de Ablemo, San Carlos | −36.452 | −71.990 | Ñuble | 324 | 2003 | 0.00 | 0.00 | 7.10 | 0.00 | 0.00 | 0.00 | 3.09 | 0.00 | 0.00 | 0.93 | 1.54 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Tres Esquinas, Coihueco | −36.556 | −71.766 | Biobío | 675 | 2013 | 0.00 | 0.00 | 0.15 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Agua Tendida, Tomé | −36.641 | −72.796 | Biobío | 300 | 2004 | 0.00 | 3.67 | 5.67 | 0.00 | 0.00 | 0.00 | 3.33 | 0.67 | 0.00 | 0.33 | 0.33 | 2 | 1 | 0 | 0.60 | 0.27 | 0.00 | 0.21 |
| Chillán-Pinto, Chillán | −36.651 | −71.999 | Ñuble | 300 | 2003 | 0.00 | 1.33 | 7.67 | 0.00 | 0.00 | 0.00 | 3.00 | 0.00 | 0.33 | 0.33 | 0.67 | 1 | 0 | 0 | 0.33 | 0.00 | 0.00 | 0.08 |
| Lloicura, Tomé | −36.689 | −72.762 | Biobío | 306 | 2003 | 0.00 | 1.63 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Peñablanca, Quillón | −36.713 | −72.572 | Ñuble | 288 | 2001 | 0.00 | 0.35 | 1.39 | 0.00 | 0.00 | 0.00 | 2.08 | 0.00 | 0.00 | 1.04 | 0.69 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Vilumanque, Concepción | −36.784 | −73.012 | Biobío | 218 | 2003 | 0.00 | 8.72 | 8.26 | 0.00 | 0.00 | 0.00 | 11.93 | 0.00 | 0.00 | 0.00 | 0.46 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Chaimávida, Concepción | −36.799 | −72.952 | Biobío | 450 | 2009 | 0.00 | 0.44 | 1.11 | 0.00 | 0.00 | 0.00 | 2.89 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Forestal Millalemu, El Carmen | −36.931 | −71.744 | Ñuble | 450 | 2005 | 0.00 | 0.67 | 0.89 | 0.00 | 0.00 | 0.00 | 4.22 | 0.00 | 0.00 | 0.00 | 0.22 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Talcamávida, Hualqui | −37.142 | −72.908 | Biobío | 450 | 2005 | 0.00 | 1.56 | 0.67 | 0.00 | 0.00 | 0.00 | 1.56 | 0.00 | 0.44 | 0.00 | 0.22 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Forestal Millalemu, Tucapel | −37.241 | −71.794 | Biobío | 300 | 2002 | 0.00 | 1.33 | 7.00 | 0.00 | 0.00 | 0.00 | 3.00 | 0.00 | 0.00 | 0.00 | 0.33 | 2 | 0 | 0 | 0.67 | 0.00 | 0.00 | 0.17 |
| Santa Juana, Santa Juana | −37.342 | −72.932 | Biobío | 450 | 2009 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 2.22 | 0.00 | 0.00 | 0.44 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Los Ángeles, Los Ángeles | −37.463 | −72.303 | Biobío | 312 | 2000 | 0.00 | 0.00 | 1.28 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.64 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cañicura, Quilleco | −37.541 | −72.688 | Biobío | 600 | 2018 | 0.00 | 0.17 | 3.33 | 0.00 | 0.00 | 0.00 | 2.83 | 0.00 | 0.00 | 0.50 | 0.50 | 3 | 0 | 0 | 1.06 | 0.00 | 0.00 | 0.41 |
| Hacienda San Lorenzo, Quilleco | −37.546 | −71.463 | Biobío | 450 | 2005 | 0.00 | 2.44 | 2.67 | 0.00 | 0.00 | 0.67 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Alto las Viñas, Los Ángeles | −37.559 | −72.544 | Biobío | 300 | 2004 | 0.00 | 0.00 | 8.67 | 0.00 | 0.00 | 0.00 | 2.67 | 0.00 | 0.33 | 0.00 | 0.67 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Antihuala, Los Álamos | −37.706 | −73.419 | Biobío | 300 | 2006 | 0.00 | 1.00 | 0.33 | 0.00 | 0.00 | 0.00 | 1.67 | 0.00 | 0.00 | 0.33 | 2.67 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| La Curva, Cañete | −37.796 | −73.308 | Araucanía | 600 | 2013 | 0.00 | 0.17 | 1.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 1.33 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Llacolén, Contulmo | −37.916 | −73.273 | Biobío | 675 | 2013 | 0.00 | 0.59 | 1.04 | 0.00 | 0.00 | 0.00 | 3.26 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Llanquén, Lonquimay | −38.150 | −71.300 | Araucanía | 600 | 2014 | 0.00 | 2.00 | 0.00 | 0.00 | 0.00 | 1.67 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 3 | 0 | 0 | 2.25 | 0.00 | 0.00 | 0.60 |
| Tirúa, Tirúa | −38.272 | −73.513 | Biobío | 270 | 2006 | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lautaro, Lautaro | −38.527 | −72.429 | Araucanía | 300 | 2002 | 0.00 | 0.00 | 2.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.67 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fundo La Aguada, Gorbea | −39.060 | −72.400 | Araucanía | 450 | 2005 | 0.00 | 1.56 | 0.44 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.22 | 0.22 | 0 | 2 | 0 | 0.00 | 1.29 | 0.00 | 0.82 |
| Lago Colico, Cunco | −39.064 | −71.973 | Araucanía | 183 | 2008 | 0.00 | 2.19 | 1.09 | 0.00 | 0.00 | 0.00 | 12.57 | 0.00 | 0.00 | 0.55 | 1.09 | 1 | 0 | 0 | 0.08 | 0.00 | 0.00 | 0.06 |
| Boroa Norte, Toltén | −39.286 | −73.076 | Araucanía | 310 | 2013 | 0.00 | 0.00 | 0.32 | 0.00 | 0.00 | 0.00 | 6.13 | 0.00 | 0.00 | 0.32 | 3.55 | 2 | 0 | 0 | 0.33 | 0.00 | 0.00 | 0.19 |
| Loncoche, Loncoche | −39.371 | −72.612 | Araucanía | 240 | 2003 | 0.00 | 2.08 | 1.67 | 0.00 | 0.00 | 0.00 | 2.50 | 0.00 | 0.00 | 0.00 | 0.42 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Llancahue Alto, Panguipulli | −39.527 | −71.813 | Los Ríos | 600 | 2014 | 0.00 | 3.83 | 0.00 | 0.00 | 0.00 | 0.17 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 3 | 0 | 0.00 | 0.78 | 0.00 | 0.75 |
| Fundo Miraflores, Lanco | −39.543 | −72.630 | Los Ríos | 675 | 2013 | 0.00 | 0.74 | 1.04 | 0.00 | 0.00 | 0.00 | 8.74 | 0.00 | 0.00 | 0.00 | 0.00 | 6 | 1 | 0 | 0.69 | 1.35 | 0.00 | 0.67 |
| Ñancul, Panguipulli | −39.729 | −72.402 | Los Ríos | 120 | 2001 | 0.00 | 0.00 | 3.33 | 0.00 | 0.00 | 0.00 | 17.50 | 0.00 | 0.00 | 0.83 | 1.67 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Huellelhue-Pishuinco, Valdivia | −39.806 | −73.069 | Los Ríos | 306 | 2003 | 0.00 | 1.63 | 6.54 | 0.00 | 0.00 | 0.00 | 0.65 | 0.00 | 0.00 | 0.00 | 0.33 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Campamento Vientos del Sur, Valdivia | −39.824 | −73.204 | Los Ríos | 300 | 2003 | 0.00 | 1.00 | 1.00 | 0.00 | 0.00 | 0.00 | 2.33 | 0.00 | 0.00 | 0.33 | 1.00 | 1 | 0 | 0 | 0.43 | 0.00 | 0.00 | 0.18 |
| San Carlos, Corral | −39.862 | −73.444 | Los Ríos | 136 | 2011 | 0.00 | 0.00 | 3.68 | 0.00 | 0.00 | 0.00 | 1.47 | 0.00 | 0.00 | 0.74 | 0.74 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Playa San Julián, Corral | −39.905 | −73.379 | Los Ríos | 330 | 2011 | 0.00 | 7.27 | 1.52 | 0.00 | 0.00 | 0.00 | 0.61 | 0.00 | 0.00 | 0.61 | 0.30 | 0 | 1 | 0 | 0.00 | 0.14 | 0.00 | 0.10 |
| Fundo Futangue, Lago Ranco | −40.365 | −72.315 | Los Ríos | 273 | 2002 | 0.00 | 3.30 | 2.20 | 0.73 | 0.00 | 0.00 | 3.30 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Las Quemas, Osorno | −40.630 | −73.115 | Los Lagos | 340 | 2017 | 0.00 | 0.88 | 3.53 | 0.00 | 0.29 | 0.59 | 0.00 | 0.00 | 0.00 | 0.59 | 0.00 | 0 | 1 | 2 | 0.00 | 1.13 | 0.57 | 0.51 |
| Ñilque, Puyehue | −40.725 | −72.434 | Los Lagos | 270 | 2004 | 0.00 | 2.96 | 1.85 | 0.00 | 0.00 | 0.00 | 5.56 | 0.00 | 0.00 | 0.37 | 0.00 | 0 | 1 | 0 | 0.00 | 0.34 | 0.00 | 0.09 |
| El Encanto, Puyehue | −40.760 | −72.444 | Los Lagos | 540 | 2017 | 0.00 | 2.04 | 1.67 | 0.00 | 1.11 | 0.00 | 4.07 | 0.00 | 0.00 | 0.37 | 0.37 | 0 | 0 | 1 | 0.00 | 0.00 | 0.60 | 0.10 |
| Rupanquito, Osorno | −40.772 | −72.783 | Los Lagos | 310 | 2013 | 0.00 | 0.97 | 3.23 | 0.00 | 0.00 | 0.00 | 8.39 | 0.00 | 0.32 | 0.32 | 0.65 | 2 | 0 | 0 | 0.24 | 0.00 | 0.00 | 0.14 |
| El Cabrito, Puerto Octay | −40.909 | −72.410 | Los Lagos | 330 | 2011 | 0.00 | 1.82 | 2.42 | 0.00 | 0.00 | 0.00 | 6.06 | 0.00 | 0.00 | 0.30 | 0.00 | 1 | 1 | 0 | 0.17 | 0.55 | 0.00 | 0.19 |
| Peulla, Puerto Varas | −41.086 | −72.018 | Los Lagos | 360 | 2006 | 0.00 | 2.22 | 0.83 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.83 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fundo Pichi-Juan, Puerto Varas | −41.243 | −72.722 | Los Lagos | 600 | 2013 | 0.00 | 0.17 | 1.83 | 0.00 | 0.17 | 0.17 | 5.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Las Quemas, Puerto Montt | −41.416 | −73191.000 | Los Lagos | 771 | 2002 | 0.00 | 0.13 | 1.43 | 0.00 | 0.00 | 0.13 | 0.39 | 0.00 | 0.00 | 0.00 | 0.13 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Caleta Rollizo, Cochamó | −41.430 | −72.308 | Los Lagos | 402 | 2011 | 0.00 | 0.25 | 3.98 | 0.00 | 0.00 | 0.00 | 1.99 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 1 | 0 | 0.00 | 4.02 | 0.00 | 0.16 |
| Paso El León, Cochamó | −41.532 | −71.831 | Los Lagos | 300 | 2006 | 0.00 | 4.00 | 1.00 | 0.00 | 0.00 | 0.00 | 12.33 | 0.00 | 0.00 | 0.00 | 0.00 | 1 | 0 | 0 | 0.08 | 0.00 | 0.00 | 0.06 |
| Parque Nacional Chiloé, Chonchi | −42.622 | −74.106 | Los Lagos | 675 | 2013 | 0.00 | 0.00 | 4.30 | 1.04 | 0.00 | 0.00 | 1.04 | 0.00 | 0.00 | 0.15 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cerro Negro, Coyhaique | −45.583 | −71.940 | Aysén | 360 | 2004 | 0.00 | 0.56 | 0.28 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0 | 0 | 0 | 0.00 | 0.00 | 0.00 | 0.00 |
References
- Morse, S.S. Factors in the emergence of infectious diseases. Emerg. Infect. Dis. 1995, 1, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.J.; Fooks, A.R.; van der Poel, W.H.M. Public health threat of new, reemerging, and neglected zoonoses in the industrialized world. Emerg. Infect. Dis. 2010, 16, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, C.B.; Schmaljohn, C.S. Replication of hantaviruses. Curr. Top. Microbiol. Immunol. 2001, 256, 15–32. [Google Scholar] [PubMed]
- Schmaljohn, C.; Hjelle, B. Hantaviruses: A global disease problem. Emerg. Infect. Dis. 1997, 3, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Hjelle, B.; Torres-Pérez, F. Hantaviruses in the Americas and their role as emerging pathogens. Viruses 2010, 2, 2559–2586. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, C.B.; Figueiredo, L.T.M.; Vapalahti, O. A global perspective on hantavirus ecology, epidemiology, and disease. Clin. Microbiol. Rev. 2010, 23, 412–441. [Google Scholar] [CrossRef]
- Nichol, S.T.; Spiropoulou, C.F.; Morzunov, S.; Rollin, P.E.; Ksiazek, T.G.; Feldmann, H.; Sánchez, A.; Childs, J.; Zaki, S.; Peters, C.J. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science 1993, 262, 914–917. [Google Scholar] [CrossRef]
- Childs, J.E.; Ksiazek, T.G.; Spiropoulou, C.F.; Krebs, J.W.; Morzunov, S.; Maupin, G.O.; Gage, K.L.; Rollin, P.E.; Sarisky, J.; Enscore, R.E. Serologic and genetic identification of Peromyscus maniculatus as the primary rodent reservoir for a new hantavirus in the southwestern United States. J. Infect. Dis. 1994, 169, 1271–1280. [Google Scholar] [CrossRef]
- Arai, S.; Ohdachi, S.D.; Asakawa, M.; Kang, H.J.; Mocz, G.; Arikawa, J.; Okabe, N.; Yanagihara, R. Molecular phylogeny of a newfound hantavirus in the Japanese shrew mole (Urotrichus talpoides). Proc. Natl. Acad. Sci. USA 2008, 105, 16296–16301. [Google Scholar] [CrossRef]
- Klempa, B.; Fichet-Calvet, E.; Lecompte, E.; Auste, B.; Aniskin, V.; Meisel, H.; Barriere, P.; Koivogui, L.; ter Meulen, J.; Kruger, D.H. Novel hantavirus sequences in Shrew, Guinea. Emerg. Infect. Dis. 2007, 13, 520–522. [Google Scholar] [CrossRef]
- Kang, H.J.; Kosoy, M.Y.; Shrestha, S.K.; Shrestha, M.P.; Pavlin, J.A.; Gibbons, R.V.; Yanagihara, R. Genetic diversity of Thottapalayam virus, a hantavirus harbored by the Asian house shrew (Suncus murinus) in Nepal. Am. J. Trop. Med. Hyg. 2011, 85, 540–545. [Google Scholar] [CrossRef] [PubMed]
- Arai, S.; Bennett, S.N.; Sumibcay, L.; Cook, J.A.; Song, J.-W.W.; Hope, A.; Parmenter, C.; Nerurkar, V.R.; Yates, T.L.; Yanagihara, R. Phylogenetically distinct hantaviruses in the masked shrew (Sorex cinereus) and dusky shrew (Sorex monticolus) in the United States. Am. J. Trop. Med. Hyg. 2008, 78, 348–351. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.-P.; Lin, X.-D.; Wang, W.; Tian, J.-H.; Cong, M.-L.; Zhang, H.-L.; Wang, M.-R.; Zhou, R.-H.; Wang, J.-B.; Li, M.-H.; et al. Phylogeny and origins of hantaviruses harbored by bats, insectivores, and rodents. PLoS Pathogens 2013, 9, e1003159. [Google Scholar] [CrossRef] [PubMed]
- MacNeil, A.; Ksiazek, T.G.; Rollin, P.E. Hantavirus pulmonary syndrome, United States, 1993–2009. Emerg. Infect. Dis. 2011, 17, 1195–1201. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.P.; Bellomo, C.M.; Cacace, M.L.; Suárez, P.; Bogni, L.; Padula, P.J. Hantavirus pulmonary syndrome in Argentina, 1995–2008. Emerg. Infect. Dis. 2010, 16, 1853–1860. [Google Scholar] [CrossRef] [PubMed]
- Ferrés, M.; Vial, P.; Marco, C.; Yañez, L.; Godoy, P.; Castillo, C.; Hjelle, B.; Delgado, I.; Lee, S.J.; Mertz, G.J. Prospective evaluation of household contacts of persons with hantavirus cardiopulmonary syndrome in Chile. J. Infect. Dis. 2007, 195, 1563–1571. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Annual, US. Hantavirus Disease and HPS Case Fatality; Centers for Disease Control and Prevention: Atlanta, GA, USA, 1993–2018. [Google Scholar]
- Levis, S.; Morzunov, S.P.; Rowe, J.E.; Enria, D.; Pini, N.; Calderón, G.; Sabattini, M.; St Jeor, S.C. Genetic diversity and epidemiology of hantaviruses in Argentina. J. Infect. Dis. 1998, 177, 529–538. [Google Scholar] [CrossRef]
- Medina, R.A.; Torres-Pérez, F.; Galeno, H.; Navarrete, M.; Vial, P.A.; Palma, R.E.; Ferrés, M.; Cook, J.A.; Hjelle, B. Ecology, genetic diversity, and phylogeographic structure of Andes virus in humans and rodents in Chile. J. Virol. 2009, 83, 2446–2459. [Google Scholar] [CrossRef]
- Torres-Pérez, F.; Navarrete-Droguett, J.; Aldunate, R.; Yates, T.L.; Mertz, G.J.; Vial, P.A.; Ferrés, M.; Marquet, P.A.; Palma, R.E. Peridomestic small mammals associated with confirmed cases of human hantavirus disease on southcentral Chile. Am. J. Trop. Med. Hyg. 2004, 70, 305–309. [Google Scholar] [CrossRef]
- Padula, P.; Figueroa, R.; Navarrete, M.; Pizarro, E.; Cadiz, R.; Bellomo, C.; Jofre, C.; Zaror, L.; Rodriguez, E.; Murúa, R. Transmission study of Andes hantavirus infection in wild sigmodontine rodents. J. Virol. 2004, 78, 11972–11979. [Google Scholar] [CrossRef]
- Martinez, V.P.; Bellomo, C.; San Juan, J.; Pinna, D.; Forlenza, R.; Elder, M.; Padula, P.J. Person-to-person transmission of Andes virus. Emerg. Infect. Dis. 2005, 11, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Lázaro, M.E.; Cantoni, G.E.; Calanni, L.M.; Resa, A.J.; Herrero, E.R.; Iacono, M.A.; Enria, D.A.; Cappa, S.M.G. Clusters of hantavirus infection, southern Argentina. Emerg. Infect. Dis. 2007, 13, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Valdebenito, C.; Calvo, M.; Vial, C.; Mansilla, R.; Marco, C.; Palma, R.E.; Vial, P.A.; Valdivieso, F.; Mertz, G.; Ferrés, M. Person-to-person household and nosocomial transmission of Andes hantavirus, Southern Chile, 2011. Emerg. Infect. Dis. 2014, 20, 1637–1644. [Google Scholar] [CrossRef]
- Padula, P.J.; Edelstein, A.; Miguel, S.D.L.; López, N.M.; Rossi, C.M.; Rabinovich, R.D. Hantavirus pulmonary syndrome outbreak in Argentina: Molecular evidence for person-to-person transmission of Andes virus. Virology 1998, 241, 323–330. [Google Scholar] [CrossRef] [PubMed]
- Piudo, L.; Monteverde, M.J.; Walker, R.S.; Douglass, R.J. Rodent community structure and Andes virus infection in Sylvan and Peridomestic habitats in Northwestern Patagonia, Argentina. Vector Borne Zoonotic Dis. 2011, 11, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.L.; Holmes, E.C.; Larrous, F.; van der Poel, W.H.; Tjornehoj, K.; Alonso, W.J.; Bourhy, H. Phylogeography, population dynamics, and molecular evolution of European bat lyssaviruses. J. Virol. 2005, 79, 10487–10497. [Google Scholar] [CrossRef]
- Vadell, M.V.; Bellomo, C.; San Martín, A.; Padula, P.; Gómez Villafañe, I. Hantavirus ecology in rodent populations in three protected areas of Argentina. Trop. Med. Int. Heal. 2011, 16, 1342–1352. [Google Scholar] [CrossRef] [PubMed]
- Toro, J.; Vega, J.D.; Khan, A.S.; Mills, J.N.; Padula, P.; Terry, W.; Yadon, Z.; Valderrama, R.; Ellis, B.A.; Pavletic, C.; et al. An outbreak of hantavirus pulmonary syndrome, Chile, 1997. Emerg. Infect. Dis. 1998, 4, 687–694. [Google Scholar] [CrossRef]
- Ortiz, J.C.; Venegas, W.; Sandoval, J.A.; Chandía, P.; Torres-Pérez, F. Hantavirus in rodents of the VIII Region of Chile. Rev. Chile Hist. Nat. 2004, 77, 251–256. [Google Scholar]
- Mills, J.N.; Johnson, J.M.; Ksiazek, T.G.; Ellis, B.A.; Rollin, P.E.; Yates, T.L.; Minn, M.O.; Johnson, M.R.; Campbell, M.L.; Miyashiro, J.; et al. A survey of hantavirus antibody in small-mammal populations in selected United States National Parks. Am. J. Trop. Med. Hyg. 1998, 58, 525–532. [Google Scholar] [CrossRef]
- Sikes, R.S.; Gannon, W.L. Guidelines of the American Society of Mammalogists for the use of wild mammals in research. J. Mammal. 2011, 92, 235–253. [Google Scholar] [CrossRef]
- Teta, P.; Pardiñas, U.F.J. Variación morfológica cualitativa y cuantitativa en Abrothrix longipilis (Cricetidae, Sigmodontinae). Mastozoología Neotropical 2014, 21, 291–309. [Google Scholar]
- D’Elía, G.; Teta, P.; Upham, N.S.; Pardiñas, U.F.J.; Patterson, B.D. Description of a new soft-haired mouse, genus Abrothrix (Sigmodontinae), from the temperate Valdivian rainforest. J. Mammal. 2015, 96, 839–853. [Google Scholar] [CrossRef]
- Yee, J.; Wortman, I.A.; Nofchissey, R.A.; Goade, D.; Bennett, S.G.; Webb, J.P.; Irwin, W.; Hjelle, B. Rapid and simple method for screening wild rodents for antibodies to Sin Nombre hantavirus. J. Wildl. Dis. 2003, 39, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Torres-Pérez, F.; Palma, R.E.; Hjelle, B.; Ferrés, M.; Cook, J.A. Andes virus infections in the rodent reservoir and in humans vary across contrasting landscapes in Chile. Infect. Genet. Evol. 2010, 10, 820–825. [Google Scholar] [CrossRef] [PubMed]
- Meserve, P.L.; Le Boulengé, E. Population dynamics and ecology of small mammals in the northern Chilean semi-arid region. Fieldiana Zool. New Ser. 1987, 39, 413–431. [Google Scholar]
- Guthmann, N.; Lozada, M.; Monjeau, J.A.; Heinemann, K.M. Population dynamics of five sigmodontine rodents of northwestern Patagonia. Acta Theriologica 1997, 42, 143–152. [Google Scholar] [CrossRef]
- Ihaka, R.; Gentleman, R. R: A Language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar]
- Kallio, E.R.; Klingström, J.; Gustafsson, E.; Manni, T.; Vaheri, A.; Henttonen, H.; Vapalahti, O.; Lundkvist, Å. Prolonged survival of Puumala hantavirus outside the host: Evidence for indirect transmission via the environment. J. Gen. Virol. 2006, 87, 2127–2134. [Google Scholar] [CrossRef]
- Arai, S.; Kikuchi, F.; Bawm, S.; Sơn, N.; Lin, K.; Tú, V.; Aoki, K.; Tsuchiya, K.; Tanaka-Taya, K.; Morikawa, S.; et al. Molecular phylogeny of Mobatviruses (Hantaviridae) in Myanmar and Vietnam. Viruses 2019, 11, 228. [Google Scholar] [CrossRef]
- Torres-Pérez, F.; Boric-Bargetto, D.; Palma Vásquez, R.E. Hantavirus in Chile: New rodents with potential epidemiological importance. Rev. Med. Chile 2016, 144, 818. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liphardt, S.W.; Kang, H.J.; Dizney, L.J.; Ruedas, L.A.; Cook, J.A.; Yanagihara, R. Complex history of codiversification and host switching of a newfound soricid-borne Orthohantavirus in North America. Viruses 2019, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Palma, R.E.; Torres-Pérez, F.; Boric-Bargetto, D. The ecology and evolutionary history of Oligoryzomys longicaudatus in southern South American. In The Quintessential Naturalist: Honoring the Life and Legacy of Oliver P. Pearson; Kelt, D., Lessa, E.P., Salazar-Bravo, J., Patton, J.L., Eds.; University of California Publications in Zoology: Los Angeles, CA, USA, 2007; Volume 134, pp. 671–693. [Google Scholar]
- Della Valle, M.G.; Edelstein, A.; Miguel, S.; Martinez, V.; Cortez, J.; Cacace, M.L.; Jurgelenas, G.; Estani, S.S.; Padula, P. Andes virus associated with hantavirus pulmonary syndrome in northern Argentina and determination of the precise site of infection. Am. J. Trop. Med. Hyg. 2002, 66, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Cautivo, K.; Schountz, T.; Acuña-Retamar, M.; Ferrés, M.; Torres-Pérez, F. Rapid enzyme-linked immunosorbent assay for the detection of hantavirus-specific antibodies in divergent small mammals. Viruses 2014, 6, 2028–2037. [Google Scholar] [CrossRef] [PubMed]
- Kuenzi, A.J.; Douglass, R.J.; Bond, C.W.; Calisher, C.H.; Mills, J.N. Long-term dynamics of Sin Nombre viral RNA and antibody in deer mice in Montana. J. Wildl. Dis. 2005, 41, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Pedreros, A.; Rutherford, P.; Gil, C. Hantavirus risk maps for Conguillio National Park, Southern Chile. Rev. Chil. Hist. Nat. 2007, 80, 363–379. [Google Scholar]
- Calderón, G.; Pini, N.; Bolpe, J.; Levis, S.; Mills, J.; Segura, E.; Guthmann, N.; Cantoni, G.; Becker, J.; Fonollat, A.; et al. Hantavirus reservoir hosts associated with peridomestic habitats in Argentina. Emerg. Infect. Dis. 1999, 5, 792–797. [Google Scholar] [CrossRef] [PubMed]
- Rosa, E.S.T.; Mills, J.N.; Padula, P.J.; Elkhoury, M.R.; Ksiazek, T.G.; Mendes, W.S.; Santos, E.D.; Araújo, G.C.B.; Martinez, V.P.; Rosa, J.F.; et al. Newly recognized hantaviruses associated with hantavirus pulmonary syndrome in northern Brazil: Partial genetic characterization of viruses and serologic implication of likely reservoirs. Vector Borne Zoonotic Dis. 2005, 5, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Luong, L.T.; Vigliotti, B.A.; Campbell, S.; Comer, J.A.; Mills, J.N.; Hudson, P.J. Dynamics of hantavirus infection in Peromyscus leucopus of central Pennsylvania. Vector Borne Zoonotic Dis. 2011, 11, 1459–1464. [Google Scholar] [CrossRef]
- Armién, A.G.; Armién, B.; Koster, F.; Pascale, J.M.; Avila, M.; Gonzalez, P.; de la Cruz, M.; Zaldivar, Y.; Mendoza, Y.; Gracia, F.; et al. Hantavirus infection and habitat associations among rodent populations in agroecosystems of Panama: Implications for human disease risk. Am. J. Trop. Med. Hyg. 2009, 81, 59–66. [Google Scholar] [CrossRef]
- Salazar-Bravo, J.; Armién, B.; Suzén, G.; Armién, A.; Ruedas, L.A.; Avila, M.; Zaldívar, Y.; Pascale, J.M.; Gracia, F.; Yates, T.L. Serosurvey of wild rodents for hantaviruses in Panama, 2000–2002. J. Wildl. Dis. 2004, 40, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Berl, J.L.; Kuenzi, A.J.; Flaherty, E.A.; Swihart, R.K. Interspecific comparison of hantavirus prevalence in Peromyscus populations from a fragmented agro-ecosystem in Indiana, USA. J. Wildl. Dis. 2017, 54, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Vigueras-Galván, A.L.; López-Pérez, A.M.; García-Peña, G.E.; Rico-Chávez, O.; Sarmiento-Silva, R.E.; Suzán, G. Current situation and perspectives on hantaviruses in Mexico. Viruses 2019, 11, 642. [Google Scholar] [CrossRef] [PubMed]
- Hjelle, B.; Glass, G.E. Outbreak of hantavirus infection in the four corners region of the United States in the wake of the 1997–1998 El Niño-southern oscillation. J. Infect. Dis. 2000, 181, 1569–1573. [Google Scholar] [CrossRef] [PubMed]
- Glass, G.E.; Shields, T.; Cai, B.; Yates, T.L.; Parmenter, R. Persistently highest risk areas for hantavirus pulmonary syndrome: Potential sites for refugia. Ecol. Appl. 2007, 17, 129–139. [Google Scholar] [CrossRef]
- Madhav, N.K.; Wagoner, K.D.; Douglass, R.J.; Mills, J.N. Delayed density-dependent prevalence of Sin Nombre virus antibody in Montana deer mice (Peromyscus maniculatus) and implications for human disease risk. Vector Borne Zoonotic Dis. 2007, 7, 353–364. [Google Scholar] [CrossRef] [PubMed]
- Adler, F.R.; Pearce-Duvet, J.M.C.; Dearing, M.D. How host population dynamics translate into time-lagged prevalence: An investigation of Sin Nombre virus in deer mice. Bull. Math. Biol. 2008, 70, 236–252. [Google Scholar] [CrossRef] [PubMed]
- Lehmer, E.M.; Clay, C.A.; Pearce-Duvet, J.; St Jeor, S.; Dearing, M.D. Differential regulation of pathogens: The role of habitat disturbance in predicting prevalence of Sin Nombre virus. Oecologia 2008, 155, 429–439. [Google Scholar] [CrossRef]
- Suzán, G.; Marce, E.; Giermakowski, J.T.; Mills, J.N.; Ceballos, G.; Ostfeld, R.S.; Armién, B.; Pascale, J.M.; Yates, T.L. Experimental evidence for reduced rodent diversity causing increased hantavirus prevalence. PLoS ONE 2009, 4, e5461. [Google Scholar] [CrossRef]
- Suzán, G.; Armién, A.; Mills, J.N.; Marce, E.; Ceballos, G.; Ávila, M.; Salazar-Bravo, J.; Ruedas, L.; Armién, B.; Yates, T.L. Epidemiological considerations of rodent community composition in fragmented landscapes in Panama. J. Mammal. 2008, 89, 684–690. [Google Scholar] [CrossRef]
- Murúa, R.; Navarrete, M.; Cádiz, R.; Figueroa, R.; Padula, P.; Zaror, L.; Mansilla, R.; González, L.; Muñoz-Pedreros, A. Síndrome pulmonar por hantavirus: Situación de los roedores reservorios y la población humana en la Décima Región, Chile. Rev. Med. Chil. 2003, 131, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Geoghegan, J.L.; Holmes, E.C. The phylogenomics of evolving virus virulence. Nat. Rev. Genet. 2018, 19, 756–769. [Google Scholar] [CrossRef] [PubMed]
- Baró, M.; Vergara, J.; Navarrete, M. Hantavirus in Chile: Review and cases analysis since 1975. Rev. Med. Chil. 1999, 127, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Pavletic, C. Hantavirus: Geographical distribution among the wild rodents in Chile. Rev. Chil. Infectol. 2000, 17, 186–196. [Google Scholar]
- Polop, F.J.; Provensal, M.C.; Pini, N.; Levis, S.C.; Priotto, J.W.; Enria, D.; Calderón, G.E.; Costa, F.; Polop, J.J. Temporal and spatial host abundance and prevalence of Andes hantavirus in Southern Argentina. Ecohealth 2010, 7, 176–184. [Google Scholar] [CrossRef]
- Yanagihara, R.; Gu, S.H.; Arai, S.; Kang, H.J.; Song, J.W. Hantaviruses: Rediscovery and new beginnings. Virus Res. 2014, 187, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H.; Wagoner, K.D.; Amman, B.R.; Root, J.J.; Douglass, R.J.; Kuenzi, A.J.; Abbott, K.D.; Parmenter, C.; Yates, T.L.; Ksiazek, T.G.; et al. Demographic factors associated with prevalence of antibody to Sin Nombre Virus in deer mice in the Western United States. J. Wildl. Dis. 2007, 43, 1–11. [Google Scholar] [CrossRef]
- Glass, G.E.; Livingstone, W.; Mills, J.N.; Hlady, W.G.; Fine, J.B.; Biggler, W.; Coke, T.; Frazier, D.; Atherley, S.; Rollin, P.E.; et al. Black creek canal virus infection in Sigmodon hispidus in Southern Florida. Am. J. Trop. Med. Hyg. 1998, 59, 699–703. [Google Scholar] [CrossRef]
- Yahnke, C.J.; Meserve, P.L.; Ksiazek, T.G.; Mills, J.N. Patterns of infection with Laguna Negra virus in wild populations of Calomys laucha in the central Paraguayan chaco. Am. J. Trop. Med. Hyg. 2001, 65, 768–776. [Google Scholar] [CrossRef]
- Cantoni, G.; Padula, P.; Calderón, G.; Mills, J.; Herrero, E.; Sandoval, P.; Martinez, V.; Pini, N.; Larrieu, E. Seasonal variation in prevalence of antibody to hantaviruses in rodents from southern Argentina. Trop. Med. Int. Heal. 2001, 6, 811–816. [Google Scholar] [CrossRef]
- Piudo, L.; Monteverde, M.J.; Walker, R.S.; Douglass, R.J. Oligoryzomys longicaudatus characteristics’ associated with the presence of Andes virus (Hantavirus). Rev. Chil. Infectología 2012, 29, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Gieseke, J. Modern Infectious Diseases Epidemiology.; Edward Arnold: London, UK, 1994. [Google Scholar]
- Juan, E.E.; Provensal, M.C.; Steinmann, A.R. Space use and social mating system of the hantavirus host, Oligoryzomys longicaudatus. Ecohealth 2018, 15, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Polop, F.; Levis, S.; Pini, N.; Enría, D.; Polop, J.; Provensal, M.C. Factors associated with hantavirus infection in a wild host rodent from Cholila, Chubut Province, Argentina. Mamm. Biol. 2018, 88, 107–113. [Google Scholar] [CrossRef]
- Mills, J.N.; Amman, B.R.; Glass, G.E. Ecology of hantaviruses and their hosts in north america. Vector Borne Zoonotic Dis. 2010, 10, 563–574. [Google Scholar] [CrossRef] [PubMed]
- Glass, G.E.; Childs, J.E.; Korch, G.W.; Leduc, J.W. Association of intraspecific wounding with hantaviral infection in wild rats (Rattus norvegicus). Epidemiol. Infect. 1988, 101, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Mills, J.N.; Yates, T.L.; Ksiazek, T.G.; Peters, C.J.; Childs, J.E. Long-term studies of hantavirus reservoir populations in the southwestern United States: Rationale, potential, and methods. Emerg. Infect. Dis. 1999, 5, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Douglass, R.J.; Calisher, C.H.; Wagoner, K.D.; Mills, J.N. Sin Nombre virus infection of deer mice in Montana: Characteristics of newly infected mice, incidence, and temporal pattern of infection. J. Wildl. Dis. 2007, 43, 12–22. [Google Scholar] [CrossRef]
- Bagamian, K.H.; Towner, J.S.; Kuenzi, A.J.; Douglass, R.J.; Rollin, P.E.; Waller, L.A.; Mills, J.N. Transmission ecology of Sin Nombre hantavirus in naturally infected North American deermouse populations in outdoor enclosures. PLoS ONE 2012, 7, e47731. [Google Scholar] [CrossRef]
- Calisher, C.H.; Sweeney, W.; Mills, J.N.; Beaty, B.J. Natural history of Sin Nombre virus in Western Colorado. Emerg. Infect. Dis. 1999, 5, 126–134. [Google Scholar] [CrossRef]
- Hinson, E.R.; Shone, S.M.; Zink, M.C.; Glass, G.E.; Klein, S.L. Wounding: The primary mode of Seoul virus transmission among male Norway rats. Am. J. Trop. Med. Hyg. 2004, 70, 310–317. [Google Scholar] [CrossRef]
- Klein, S.L.; Zink, M.C.; Glass, G.E. Seoul virus infection increases aggressive behaviour in male Norway rats. Anim. Behav. 2004, 67, 421–429. [Google Scholar] [CrossRef]
- Gonzalez, L.A.; Gaete, H.; Jofre, C.; Gonzalez, L.A.; Gaete, H.; Jofre, C. Variación estacional de los patrones conductuales en Oryzomys longicaudatus y Akodon olivaceus en encuentros intraespecíficos e interespecíficos. Bol. Soc. Biol. Concepc. 1990, 61, 63–70. [Google Scholar]
- Larrieu, E.; Cantoni, G.; Herrero, E.; Perez, A.; Talmon, G.; Vazquez, G.; Arellano, O.; Padula, P. Hantavirus antibodies in rodents and human cases with pulmonary syndrome, Rio Negro, Argentina. Medicina 2008, 68, 373–379. [Google Scholar] [PubMed]
- Astorga, F.; Escobar, L.E.; Poo-Muñoz, D.; Escobar-Dodero, J.; Rojas-Hucks, S.; Alvarado-Rybak, M.; Duclos, M.; Romero-Alvarez, D.; Molina-Burgos, B.E.; Peñafiel-Ricaurte, A.; et al. Distributional ecology of Andes hantavirus: A macroecological approach. Int. J. Health Geogr. 2018, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Linard, C.; Tersago, K.; Leirs, H.; Lambin, E.F. Environmental conditions and Puumala virus transmission in Belgium. Int. J. Heal. Geogr. 2007, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Torres-Pérez, F.; Palma, R.E.; Hjelle, B.; Holmes, E.C.; Cook, J.A. Spatial but not temporal co-divergence of a virus and its mammalian host. Mol. Ecol. 2011, 20, 4109–4122. [Google Scholar] [CrossRef] [PubMed]
- Veblen, T.T. Temperate forests of the Southern Andean Region. In The Physical Geography of South America; Veblen, T.T., Orme, A.R., Young, K.R., Eds.; Oxford University Press: Oxford, UK, 2007; pp. 217–231. [Google Scholar]
- Armesto, J.J.; Arollo, M.T.K.; Hinojosa, F. The Mediterranean environment of central Chile. In The Physical Geography of South America; Veblen, T.T., Young, K.R., Orme, A.R., Eds.; Oxford University Press: New York, NY, USA, 2007; pp. 184–199. [Google Scholar]
- Palma, R.E.; Boric-Bargetto, D.; Torres-Pérez, F.; Hernández, C.E.; Yates, T.L. Glaciation effects on the phylogeographic structure of Oligoryzomys longicaudatus (Rodentia: Sigmodontinae) in the Southern Andes. PLoS ONE 2012, 7, e32206. [Google Scholar] [CrossRef]
- Lazo-Cancino, D.; Musleh, S.S.; Hernandez, C.E.; Palma, E.; Rodriguez-Serrano, E. Does silvoagropecuary landscape fragmentation affect the genetic diversity of the sigmodontine rodent Oligoryzomys longicaudatus? PeerJ 2017, 5, e3842. [Google Scholar] [CrossRef]
- Palma, R.E.; Cancino, R.A.; Rodríguez-Serrano, E. Molecular systematics of Abrothrix longipilis (Rodentia: Cricetidae: Sigmodontinae) in Chile. J. Mammal. 2010, 91, 1102–1111. [Google Scholar] [CrossRef]
- Rodríguez-Serrano, E.; Cancino, R.A.; Palma, R.E. Molecular phylogeography of Abrothrix olivaceus (Rodentia: Sigmodontinae) in Chile. J. Mammal. 2006, 87, 971–980. [Google Scholar] [CrossRef]
- Belmar-Lucero, S.; Godoy, P.; Ferrés, M.; Vial, P.; Palma, R.E. Range expansion of Oligoryzomys longicaudatus (Rodentia, Sigmodontinae) in Patagonian Chile, and first record of hantavirus in the region. Rev. Chil. Hist. Nat. 2009, 82, 265–275. [Google Scholar] [CrossRef]
- Carver, S.; Mills, J.N.; Parmenter, C.A.; Parmenter, R.R.; Richardson, K.S.; Harris, R.L.; Douglass, R.J.; Kuenzi, A.J.; Luis, A.D. Toward a mechanistic understanding of environmentally forced zoonotic disease emergence: Sin nombre hantavirus. Bioscience 2015, 65, 651–666. [Google Scholar] [CrossRef] [PubMed]
- Guterres, A.; de Lemos, E.R.S. Hantaviruses and a neglected environmental determinant. One Heal. 2018, 5, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Langlois, J.P.; Fahrig, L.; Merriam, G.; Artsob, H. Landscape structure influences continental distribution of hantavirus in deer mice. Landsc. Ecol. 2001, 16, 255–266. [Google Scholar] [CrossRef]
- Rynkiewicz, E.C.; Pedersen, A.B.; Fenton, A. An ecosystem approach to understanding and managing within-host parasite community dynamics. Trends Parasitol. 2015, 31, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Barrera, K.; Murua, R. Nuevo desafío en Salud Pública: Presencia de reservorios de Hanta, Oligoryzomys longicaudatus y Rattus spp., en aéreas de borde en praderas del sur de Chile. Sustain. Agri. Food Environ. Res. 2015, 3, 33–46. [Google Scholar]
- Monteverde, M.J.; Hodara, K. Movimientos de roedores intra-e inter-ambiente y riesgo de exposición al hantavirus Andes en Patagonia Norte, Argentina. Ecol. Austral 2017, 27, 279–289. [Google Scholar] [CrossRef]
- Belay, E.D.; Kile, J.C.; Hall, A.J.; Barton-behravesh, C.; Parsons, M.B.; Salyer, S.; Walke, H. Zoonotic disease programs for enhancing global health security. Emerg. Infect. Dis. 2017, 23, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Plowright, R.K.; Parrish, C.R.; Mccallum, H.; Hudson, P.J.; Ko, A.I.; Graham, A.L.; Lloyd-smith, J.O. Pathways to zoonotic spillover. Nat. Publ. Gr. 2017, 15, 502–510. [Google Scholar] [CrossRef] [PubMed]




| Relative Abundance | Relative Seropositivity | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Locality, County | Administrative Region | Al | Ah | Ao | As | Am | Lm | Ol | Pd | Mm | Rn | Rr | Ol | Ah | Ao | Total |
| Chiñigue, Ovalle | Coquimbo | 1.00 | 0.00 | 2.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.67 | 0.67 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fundo Chuico Blanco, Hijuelas | Valparaíso | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.37 | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Llaillay, Llaillay | Valparaíso | 0.00 | 0.00 | 0.42 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.53 | 0.42 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lo Mardones, Quillota | Valparaíso | 0.00 | 0.00 | 2.06 | 0.00 | 0.00 | 0.00 | 0.48 | 0.00 | 0.63 | 0.95 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| La Palma, Quillota | Valparaíso | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.83 | 0.17 | 3.00 | 0.00 | 0.00 | 0.60 |
| Población Prat, Villa Alemana | Valparaíso | 0.00 | 0.00 | 4.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 |
| Laguna Verde, Valparaíso | Valparaíso | 0.83 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Las Docas, Valparaíso | Valparaíso | 3.23 | 0.00 | 0.40 | 0.00 | 0.00 | 0.00 | 1.41 | 1.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Alto el Manzano, TilTil | Metropolitan | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| San Antonio de Naltahua, Isla de Maipo | Metropolitan | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 |
| La Reina, La Reina | Metropolitan | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lo Espejo, Lo Espejo | Metropolitan | 0.00 | 0.00 | 0.50 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.17 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| El Canelo, San José de Maipo | Metropolitan | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.39 | 0.39 | 0.00 | 0.39 | 0.78 | 0.00 | 0.00 | 0.00 | 0.00 |
| La Florida Alto, La Florida | Metropolitan | 0.37 | 0.00 | 3.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Mallarauco, Melipilla | Metropolitan | 0.00 | 0.00 | 1.67 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 2.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| San Antonio, San Antonio | Valparaíso | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.44 | 0.00 | 1.33 | 0.00 | 0.89 | 0.00 | 0.00 | 0.00 | 0.00 |
| Pomaire, Melipilla | Metropolitan | 0.00 | 0.00 | 0.89 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.22 | 0.22 | 0.00 | 0.00 | 0.00 | 0.00 |
| Constructora Inca, Melipilla | Metropolitan | 0.00 | 0.00 | 2.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.32 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Condominio Puerta del Sol, Talagante | Metropolitan | 0.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Talagante, Talagante | Metropolitan | 0.00 | 0.00 | 6.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.42 | 2.08 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Chocalán, Melipilla | Metropolitan | 0.00 | 0.00 | 3.67 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fundo La Ventolera, Santo Domingo | Valparaíso | 4.00 | 0.00 | 6.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| El Ingenio, San José de Maipo | Metropolitan | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Hijuelas, Hijuelas | Valparaíso | 0.00 | 0.00 | 0.95 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.95 | 0.48 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Abrantes, Paine | Metropolitan | 0.00 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.33 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Ucúquer, Litueche | O’Higgins | 0.00 | 0.00 | 3.11 | 0.00 | 0.00 | 0.00 | 1.33 | 1.33 | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.32 | 0.15 |
| Quilamuta-Las Cabras, Las Cabras | O’Higgins | 0.33 | 0.00 | 1.00 | 0.00 | 0.00 | 0.00 | 2.67 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Coya, Machalí | O’Higgins | 0.00 | 0.00 | 0.69 | 0.00 | 0.00 | 0.00 | 0.69 | 1.38 | 0.00 | 3.45 | 0.00 | 1.45 | 0.00 | 0.00 | 0.16 |
| Sn Vicente de Tagua Tagua, Sn Vicente de Tagua Tagua | O’Higgins | 0.00 | 0.00 | 0.28 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.42 | 0.00 | 0.14 | 0.00 | 0.00 | 0.00 | 0.00 |
| Los Maitenes, Lolol | O’Higgins | 0.00 | 0.00 | 0.46 | 0.00 | 0.00 | 0.00 | 0.31 | 0.00 | 0.15 | 0.31 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Las Peñas, San Fernando 2002 | O’Higgins | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.28 | 0.00 | 0.00 | 0.00 | 0.00 | 3.60 | 0.00 | 0.00 | 1.20 |
| Las Peñas, San Fernando 2018 | O’Higgins | 0.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.50 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| El Sauce, Chimbarongo | O’Higgins | 0.00 | 0.00 | 2.31 | 0.00 | 0.00 | 0.00 | 0.77 | 0.00 | 0.00 | 0.00 | 0.26 | 1.30 | 0.00 | 0.00 | 0.30 |
| Lipimávida, Vichuquén | Maule | 0.44 | 0.00 | 0.22 | 0.00 | 0.00 | 0.00 | 1.78 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Duao, Licantén | Maule | 1.00 | 0.00 | 8.00 | 0.00 | 0.00 | 0.00 | 12.33 | 0.00 | 0.00 | 0.67 | 0.00 | 0.32 | 0.00 | 0.00 | 0.18 |
| Escuela Quilpoco, Rauco | Maule | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.37 | 0.00 | 0.37 | 1.11 | 0.00 | 2.70 | 0.00 | 0.00 | 0.54 |
| El Pumaitén, Romeral | Maule | 0.67 | 0.00 | 3.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 |
| Hualañé, Hualañé 2010 | Maule | 0.00 | 0.00 | 0.22 | 0.00 | 0.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| El Trapiche, Curicó | Maule | 0.00 | 0.00 | 3.00 | 0.00 | 0.00 | 0.00 | 1.00 | 0.00 | 0.33 | 0.17 | 0.50 | 1.00 | 0.00 | 0.00 | 0.20 |
| Hualañé, Hualañé 2012 | Maule | 0.00 | 0.00 | 3.06 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Palquibudi, Rauco | Maule | 0.00 | 0.00 | 0.74 | 0.00 | 0.00 | 0.00 | 0.37 | 0.00 | 0.00 | 0.37 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Los Queñes, Romeral | Maule | 0.00 | 0.00 | 0.28 | 0.00 | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.28 | 1.39 | 3.60 | 0.00 | 3.60 | 1.64 |
| Las Lomas, San Clemente | Maule | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 |
| Canal Melado, Longaví | Maule | 0.00 | 0.27 | 1.87 | 0.00 | 0.00 | 0.00 | 0.80 | 0.00 | 0.00 | 0.00 | 0.00 | 1.25 | 0.00 | 0.00 | 0.34 |
| Retupel, Cauquenes | Maule | 0.00 | 0.00 | 2.33 | 0.00 | 0.00 | 0.00 | 1.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.60 | 0.00 | 0.00 | 0.25 |
| Bullileo, Parral | Maule | 0.00 | 0.42 | 0.00 | 0.00 | 0.00 | 0.00 | 1.25 | 0.00 | 0.00 | 0.42 | 1.25 | 0.00 | 0.00 | 0.00 | 0.00 |
| San Miguel de Ablemo, San Carlos | Ñuble | 0.00 | 0.00 | 7.10 | 0.00 | 0.00 | 0.00 | 3.09 | 0.00 | 0.00 | 0.93 | 1.54 | 0.00 | 0.00 | 0.00 | 0.00 |
| Tres Esquinas, Coihueco | Biobío | 0.00 | 0.00 | 0.15 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Agua Tendida, Tomé | Biobío | 0.00 | 3.67 | 5.67 | 0.00 | 0.00 | 0.00 | 3.33 | 0.67 | 0.00 | 0.33 | 0.33 | 0.60 | 0.27 | 0.00 | 0.21 |
| Chillán-Pinto, Chillán | Ñuble | 0.00 | 1.33 | 7.67 | 0.00 | 0.00 | 0.00 | 3.00 | 0.00 | 0.33 | 0.33 | 0.67 | 0.33 | 0.00 | 0.00 | 0.08 |
| Lloicura, Tomé | Biobío | 0.00 | 1.63 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 |
| Peñablanca, Quillón | Ñuble | 0.00 | 0.35 | 1.39 | 0.00 | 0.00 | 0.00 | 2.08 | 0.00 | 0.00 | 1.04 | 0.69 | 0.00 | 0.00 | 0.00 | 0.00 |
| Vilumanque, Concepción | Biobío | 0.00 | 8.72 | 8.26 | 0.00 | 0.00 | 0.00 | 11.93 | 0.00 | 0.00 | 0.00 | 0.46 | 0.00 | 0.00 | 0.00 | 0.00 |
| Chaimávida, Concepción | Biobío | 0.00 | 0.44 | 1.11 | 0.00 | 0.00 | 0.00 | 2.89 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Forestal Millalemu, El Carmen | Ñuble | 0.00 | 0.67 | 0.89 | 0.00 | 0.00 | 0.00 | 4.22 | 0.00 | 0.00 | 0.00 | 0.22 | 0.00 | 0.00 | 0.00 | 0.00 |
| Talcamávida, Hualqui | Biobío | 0.00 | 1.56 | 0.67 | 0.00 | 0.00 | 0.00 | 1.56 | 0.00 | 0.44 | 0.00 | 0.22 | 0.00 | 0.00 | 0.00 | 0.00 |
| Forestal Millalemu, Tucapel | Biobío | 0.00 | 1.33 | 7.00 | 0.00 | 0.00 | 0.00 | 3.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.67 | 0.00 | 0.00 | 0.17 |
| Santa Juana, Santa Juana | Biobío | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 2.22 | 0.00 | 0.00 | 0.44 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Los Ángeles, Los Ángeles | Biobío | 0.00 | 0.00 | 1.28 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.64 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cañicura, Quilleco | Biobío | 0.00 | 0.17 | 3.33 | 0.00 | 0.00 | 0.00 | 2.83 | 0.00 | 0.00 | 0.50 | 0.50 | 1.06 | 0.00 | 0.00 | 0.41 |
| Hacienda San Lorenzo, Quilleco | Biobío | 0.00 | 2.44 | 2.67 | 0.00 | 0.00 | 0.67 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Alto las Viñas, Los Ángeles | Biobío | 0.00 | 0.00 | 8.67 | 0.00 | 0.00 | 0.00 | 2.67 | 0.00 | 0.33 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 |
| Antihuala, Los Álamos | Biobío | 0.00 | 1.00 | 0.33 | 0.00 | 0.00 | 0.00 | 1.67 | 0.00 | 0.00 | 0.33 | 2.67 | 0.00 | 0.00 | 0.00 | 0.00 |
| La Curva, Cañete | Araucanía | 0.00 | 0.17 | 1.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Llacolén, Contulmo | Biobío | 0.00 | 0.59 | 1.04 | 0.00 | 0.00 | 0.00 | 3.26 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Llanquén, Lonquimay | Araucanía | 0.00 | 2.00 | 0.00 | 0.00 | 0.00 | 1.67 | 1.33 | 0.00 | 0.00 | 0.00 | 0.00 | 2.25 | 0.00 | 0.00 | 0.60 |
| Tirúa, Tirúa | Biobío | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.00 | 1.11 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Lautaro, Lautaro | Araucanía | 0.00 | 0.00 | 2.00 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.67 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fundo La Aguada, Gorbea | Araucanía | 0.00 | 1.56 | 0.44 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.22 | 0.22 | 0.00 | 1.29 | 0.00 | 0.82 |
| Lago Colico, Cunco | Araucanía | 0.00 | 2.19 | 1.09 | 0.00 | 0.00 | 0.00 | 12.57 | 0.00 | 0.00 | 0.55 | 1.09 | 0.08 | 0.00 | 0.00 | 0.06 |
| Boroa Norte, Toltén | Araucanía | 0.00 | 0.00 | 0.32 | 0.00 | 0.00 | 0.00 | 6.13 | 0.00 | 0.00 | 0.32 | 3.55 | 0.33 | 0.00 | 0.00 | 0.19 |
| Loncoche, Loncoche | Araucanía | 0.00 | 2.08 | 1.67 | 0.00 | 0.00 | 0.00 | 2.50 | 0.00 | 0.00 | 0.00 | 0.42 | 0.00 | 0.00 | 0.00 | 0.00 |
| Llancahue Alto, Panguipulli | Los Ríos | 0.00 | 3.83 | 0.00 | 0.00 | 0.00 | 0.17 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.78 | 0.00 | 0.75 |
| Fundo Miraflores, Lanco | Los Ríos | 0.00 | 0.74 | 1.04 | 0.00 | 0.00 | 0.00 | 8.74 | 0.00 | 0.00 | 0.00 | 0.00 | 0.69 | 1.35 | 0.00 | 0.67 |
| Ñancul, Panguipulli | Los Ríos | 0.00 | 0.00 | 3.33 | 0.00 | 0.00 | 0.00 | 17.50 | 0.00 | 0.00 | 0.83 | 1.67 | 0.00 | 0.00 | 0.00 | 0.00 |
| Huellelhue-Pishuinco, Valdivia | Los Ríos | 0.00 | 1.63 | 6.54 | 0.00 | 0.00 | 0.00 | 0.65 | 0.00 | 0.00 | 0.00 | 0.33 | 0.00 | 0.00 | 0.00 | 0.00 |
| Campamento Vientos del Sur, Valdivia | Los Ríos | 0.00 | 1.00 | 1.00 | 0.00 | 0.00 | 0.00 | 2.33 | 0.00 | 0.00 | 0.33 | 1.00 | 0.43 | 0.00 | 0.00 | 0.18 |
| San Carlos, Corral | Los Ríos | 0.00 | 0.00 | 3.68 | 0.00 | 0.00 | 0.00 | 1.47 | 0.00 | 0.00 | 0.74 | 0.74 | 0.00 | 0.00 | 0.00 | 0.00 |
| Playa San Julián, Corral | Los Ríos | 0.00 | 7.27 | 1.52 | 0.00 | 0.00 | 0.00 | 0.61 | 0.00 | 0.00 | 0.61 | 0.30 | 0.00 | 0.14 | 0.00 | 0.10 |
| Fundo Futangue, Lago Ranco | Los Ríos | 0.00 | 3.30 | 2.20 | 0.73 | 0.00 | 0.00 | 3.30 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Las Quemas, Osorno | Los Lagos | 0.00 | 0.88 | 3.53 | 0.00 | 0.29 | 0.59 | 0.00 | 0.00 | 0.00 | 0.59 | 0.00 | 0.00 | 1.13 | 0.57 | 0.51 |
| Ñilque, Puyehue | Los Lagos | 0.00 | 2.96 | 1.85 | 0.00 | 0.00 | 0.00 | 5.56 | 0.00 | 0.00 | 0.37 | 0.00 | 0.00 | 0.34 | 0.00 | 0.09 |
| El Encanto, Puyehue | Los Lagos | 0.00 | 2.04 | 1.67 | 0.00 | 1.11 | 0.00 | 4.07 | 0.00 | 0.00 | 0.37 | 0.37 | 0.00 | 0.00 | 0.60 | 0.10 |
| Rupanquito, Osorno | Los Lagos | 0.00 | 0.97 | 3.23 | 0.00 | 0.00 | 0.00 | 8.39 | 0.00 | 0.32 | 0.32 | 0.65 | 0.24 | 0.00 | 0.00 | 0.14 |
| El Cabrito, Puerto Octay | Los Lagos | 0.00 | 1.82 | 2.42 | 0.00 | 0.00 | 0.00 | 6.06 | 0.00 | 0.00 | 0.30 | 0.00 | 0.17 | 0.55 | 0.00 | 0.19 |
| Peulla, Puerto Varas | Los Lagos | 0.00 | 2.22 | 0.83 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.83 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fundo Pichi-Juan, Puerto Varas | Los Lagos | 0.00 | 0.17 | 1.83 | 0.00 | 0.17 | 0.17 | 5.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Las Quemas, Puerto Montt | Los Lagos | 0.00 | 0.13 | 1.43 | 0.00 | 0.00 | 0.13 | 0.39 | 0.00 | 0.00 | 0.00 | 0.13 | 0.00 | 0.00 | 0.00 | 0.00 |
| Caleta Rollizo, Cochamó | Los Lagos | 0.00 | 0.25 | 3.98 | 0.00 | 0.00 | 0.00 | 1.99 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4.02 | 0.00 | 0.16 |
| Paso El León, Cochamó | Los Lagos | 0.00 | 4.00 | 1.00 | 0.00 | 0.00 | 0.00 | 12.33 | 0.00 | 0.00 | 0.00 | 0.00 | 0.08 | 0.00 | 0.00 | 0.06 |
| Parque Nacional Chiloé, Chonchi | Los Lagos | 0.00 | 0.00 | 4.30 | 1.04 | 0.00 | 0.00 | 1.04 | 0.00 | 0.00 | 0.15 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Cerro Negro, Coyhaique | Aysén | 0.00 | 0.56 | 0.28 | 0.00 | 0.00 | 0.00 | 0.56 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Species/Trait | O. longicaudatus | A. hirta | A. olivacea | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (+) | (−) | Sum | chi-Square | df | p | (+) | (−) | Sum | p | (+) | (−) | Sum | p | ||
| Sex | Male | 32 | 319 | 351 | 8.93 | 1 | 0.0028 | 8 | 129 | 137 | 0.394 | 4 | 376 | 380 | 0.419 |
| Female | 9 | 271 | 280 | 4 | 93 | 97 | 1 | 255 | 256 | ||||||
| Sum | 41 | 590 | 631 | 12 | 222 | 234 | 5 | 631 | 636 | ||||||
| Age | Adult | 41 | 499 | 540 | 7.39 | 1 | 0.0066 | 11 | 187 | 198 | 0.423 | 3 | 556 | 559 | 0.114 |
| Juvenile | 0 | 91 | 91 | 1 | 35 | 36 | 2 | 75 | 77 | ||||||
| Sum | 41 | 590 | 631 | 12 | 222 | 234 | 5 | 631 | 636 | ||||||
| Wound | Wounds | 26 | 101 | 127 | 51.11 | 1 | <0.0001 | 5 | 62 | 67 | 0.236 | 2 | 89 | 91 | 0.152 |
| Without wounds | 15 | 489 | 504 | 7 | 160 | 167 | 3 | 542 | 545 | ||||||
| Sum | 41 | 590 | 631 | 12 | 222 | 234 | 5 | 631 | 636 | ||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres-Pérez, F.; Palma, R.E.; Boric-Bargetto, D.; Vial, C.; Ferrés, M.; Vial, P.A.; Martínez-Valdebenito, C.; Pavletic, C.; Parra, A.; Marquet, P.A.; et al. A 19 Year Analysis of Small Mammals Associated with Human Hantavirus Cases in Chile. Viruses 2019, 11, 848. https://doi.org/10.3390/v11090848
Torres-Pérez F, Palma RE, Boric-Bargetto D, Vial C, Ferrés M, Vial PA, Martínez-Valdebenito C, Pavletic C, Parra A, Marquet PA, et al. A 19 Year Analysis of Small Mammals Associated with Human Hantavirus Cases in Chile. Viruses. 2019; 11(9):848. https://doi.org/10.3390/v11090848
Chicago/Turabian StyleTorres-Pérez, Fernando, R. Eduardo Palma, Dusan Boric-Bargetto, Cecilia Vial, Marcela Ferrés, Pablo A. Vial, Constanza Martínez-Valdebenito, Carlos Pavletic, Alonso Parra, Pablo A. Marquet, and et al. 2019. "A 19 Year Analysis of Small Mammals Associated with Human Hantavirus Cases in Chile" Viruses 11, no. 9: 848. https://doi.org/10.3390/v11090848
APA StyleTorres-Pérez, F., Palma, R. E., Boric-Bargetto, D., Vial, C., Ferrés, M., Vial, P. A., Martínez-Valdebenito, C., Pavletic, C., Parra, A., Marquet, P. A., & Mertz, G. J. (2019). A 19 Year Analysis of Small Mammals Associated with Human Hantavirus Cases in Chile. Viruses, 11(9), 848. https://doi.org/10.3390/v11090848