The Nucleoprotein and Phosphoprotein of Peste des Petits Ruminants Virus Inhibit Interferons Signaling by Blocking the JAK-STAT Pathway

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells, Virus, and Antibodies
2.2. Plasmids Transfection and Luciferase Reporter Assays
2.3. Co-Immunoprecipitation Assays and Western Blotting Analysis
2.4. Indirect Immunofluorescence Assays
2.5. RNA Isolation and Quantitative Real-Time PCR (qPCR)
2.6. Statistical Analysis
3. Results
3.1. PPRV N and P Proteins Inhibit Both IFN-β- and IFN-γ-Induced IFN Response
3.2. PPRV N and P Proteins Suppress IFN-β- and IFN-γ-Induced Expression of Antiviral Genes
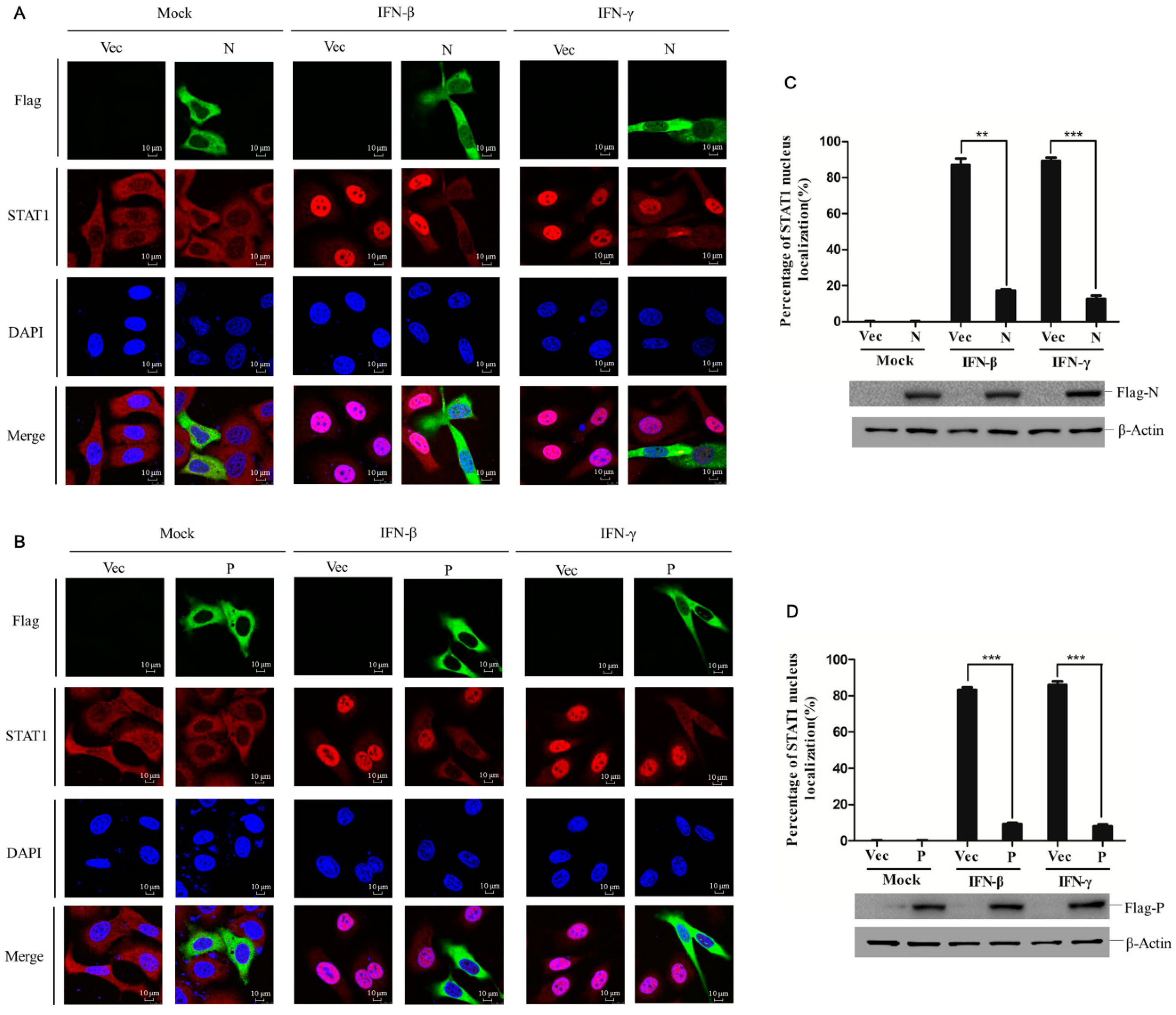
3.3. PPRV N and P Proteins Block STAT1 Nuclear Translocation
3.4. PPRV N Protein Did Not Block Phosphorylation and Dimerization of STATs
3.5. Core and Tail Domains of N Protein Play Different Suppressive Roles on IFN-β- and IFN-γ-Triggered Cellular IFN Responses
3.6. PPRV P Protein Interacts with STAT1 and Blocks STAT1 phosphorylation
3.7. Tyrosine 110 of P Protein is Critical for Blocking IFN-β- and IFN-γ-Induced STAT1 Activation
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Baron, M.D.; Diallo, A.; Lancelot, R.; Libeau, G. Peste des Petits Ruminants Virus. Adv. Virus Res. 2016, 95, 1–42. [Google Scholar] [CrossRef] [PubMed]
- Gargadennec, L.; Lalanne, A. La peste des petits ruminants. Bull. Serv. Zoo Techiques Epizoot. Afr. Occident. Fr. 1942, 5, 16–21. [Google Scholar]
- Banyard, A.C.; Parida, S.; Batten, C.; Oura, C.; Kwiatek, O.; Libeau, G. Global distribution of peste des petits ruminants virus and prospects for improved diagnosis and control. J. Gen. Virol. 2010, 91, 2885–2897. [Google Scholar] [CrossRef] [PubMed]
- Donduashvili, M.; Goginashvili, K.; Toklikishvili, N.; Tigilauri, T.; Gelashvili, L.; Avaliani, L.; Khartskhia, N.; Loitsch, A.; Bataille, A.; Libeau, G.; et al. Identification of Peste des Petits Ruminants Virus, Georgia, 2016. Emerg. Infect. Dis. 2018, 24, 1576–1578. [Google Scholar] [CrossRef] [PubMed]
- Rajko-Nenow, P.Z.; Cunliffe, T.G.; Flannery, J.T.; Ropiak, H.M.; Avaliani, L.; Donduashvili, M.; Baron, M.D.; Batten, C.A. Complete genome sequence of peste des petits ruminants virus from Georgia, 2016. Genome Announc. 2017, 5, e01091-17. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, J.; Li, L.; Liu, Y.; Wu, X.; Wang, Z. Peste des petits ruminants in China since its first outbreak in 2007: A 10-year review. Transbound. Emerg. Dis. 2018, 65, 638–648. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Maherchandani, S.; Kashyap, S.K.; Singh, S.V.; Sharma, S.; Chaubey, K.K.; Ly, H. Peste Des Petits Ruminants Virus Infection of Small Ruminants: A Comprehensive Review. Viruses 2014, 6, 2287–2327. [Google Scholar] [CrossRef]
- Gibbs, E.P.J.; Taylor, W.P.; Lawman, M.J.P.; Bryant, J. Classification of peste des petits ruminants virus as the fourth member of the genus Morbillivirus. Intervirology 1979, 11, 268–274. [Google Scholar] [CrossRef]
- Platanias, L.C. Mechanisms of type-I- and type-II-interferon-mediated signalling. Nat. Rev. Immunol. 2005, 5, 375–386. [Google Scholar] [CrossRef]
- Der, S.; Zhou, A.M.; Silverman, R.H.; William, B.R.G. Identification of genes differentially regulated by IFN-alpha, beta, gamma using oligonucleotide microarrays. Proc. Natl. Acad. Sci. USA 1998, 95, 15623–15628. [Google Scholar] [CrossRef]
- Stark, G.R.; Kerr, I.M.; Williams, B.R.G.; Silverman, R.H.; Schreiber, R.D. How Cells Respond to Interferons. Annu. Rev. Biochem. 1998, 67, 227–264. [Google Scholar] [CrossRef] [PubMed]
- Ihle, J.N. The Janus protein tyrosine kinase family and its role in cytokine signaling. Adv. Immunol. 1995, 60, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.; Shuai, K.; Prezioso, V.; Darnell, J. Interferon-dependent tyrosine phosphorylation of a latent cytoplasmic transcription factor. Science 1992, 257, 809–813. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, S.A.; Salditt-Georgieff, M.; Darnell, J.E. Tyrosine-phosphorylated Stat1 and Stat2 plus a 48-kDa protein all contact DNA in forming interferon-stimulated-gene factor 3. Proc. Natl. Acad. Sci. USA 1995, 92, 3829–3833. [Google Scholar] [CrossRef] [PubMed]
- Darnell, J.; Kerr, I.; Stark, G. Jak-STAT pathways and transcriptional activation in response to IFNs and other extracellular signaling proteins. Science 1994, 264, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Pestka, S.; Kotenko, S.V.; Muthukumaran, G.; Izotova, L.S.; Cook, J.R.; Garotta, G. The interferon gamma (IFN-γ) receptor: A paradigm for the multichain cytokine receptor. Cytokine Growth Factor Rev. 1997, 8, 189–206. [Google Scholar] [CrossRef]
- Schroder, K.; Hertzog, P.J.; Ravasi, T.; Hume, D.A. Interferon-gamma: An overview of signals, mechanisms and functions. J. Leukoc. Biol. 2004, 75, 163–189. [Google Scholar] [CrossRef] [PubMed]
- Shuai, K.; Horvath, C.M.; Huang, L.H.T.; Qureshi, S.A.; Cowburn, D.; Darnell, J.E. Interferon activation of the transcription factor Stat91 involves dimerization through SH2-phosphotyrosyl peptide interactions. Cell 1994, 76, 821–828. [Google Scholar] [CrossRef]
- Andrejeva, J.; Young, D.F.; Goodbourn, S.; Randall, R.E. Degradation of STAT1 and STAT2 by the V proteins of simian virus 5 and human parainfluenza virus type 2, respectively: Consequences for virus replication in the presence of alpha/beta and gamma interferons. J. Virol. 2002, 76, 2159–2167. [Google Scholar] [CrossRef]
- Caignard, G.; Guerbois, M.; Labernardière, J.L.; Jacob, Y.; Jones, L.M.; Wild, F.; Tangy, F.; Vidalain, P.O. Measles virus V protein blocks Jak1-mediated phosphorylation of STAT1 to escape IFN-α/β signaling. Virology 2007, 368, 351–362. [Google Scholar] [CrossRef]
- Precious, B.; Young, D.F.; Andrejeva, L.; Goodbourn, S.; Randall, R.E. In vitro and in vivo specificity of ubiquitination and degradation of STAT1 and STAT2 by the V proteins of the paramyxoviruses simian virus 5 and human parainfluenza virus type 2. J. Gen. Virol. 2005, 86, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Nanda, S.K.; Baron, M.D. Rinderpest virus blocks type I and type II interferon action: Role of structural and nonstructural proteins. J. Virol. 2006, 80, 7555–7568. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.L.; Garcia-Sastre, A.; Palese, P.; Basler, C.F. Nipah virus V and W proteins have a common STAT1-binding domain yet inhibit STAT1 activation from the cytoplasmic and nuclear compartments, respectively. J. Virol. 2004, 78, 5633–5641. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.S.; Yang, X.; Nian, X.F.; Zhang, Z.D.; Dou, Y.X.; Zhang, X.H.; Luo, X.N.; Su, J.H.; Zhu, Q.Y.; Cai, X.P. Identification of amino-acid residues in the V protein of peste des petits ruminants essential for interference and suppression of STAT-mediated interferon signaling. Virology 2015, 483, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Takayama, I.; Sato, H.; Watanabe, A.; Omi-Furutani, M.; Sugai, A.; Kanki, K.; Yoneda, M.; Kai, C.K. The nucleocapsid protein of measles virus blocks host interferon response. Virology 2012, 424, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Sugai, A.; Sato, H.; Takayama, I.; Yoneda, M.; Kai, C. Nipah and Hendra Virus Nucleoproteins Inhibit Nuclear Accumulation of Signal Transducer and Activator of Transcription 1 (STAT1) and STAT2 by Interfering with Their Complex Formation. J. Virol. 2017, 91, 15. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, C.; Du, X.; Wang, G.; Cao, W.; Yang, F.; Feng, H.; Zhang, X.; Shi, Z.; Liu, H.; et al. Foot-and-mouth disease virus infection inhibits LGP2 protein expression to exaggerate inflammatory response and promote viral replication. Cell Death Dis. 2017, 8, e2747. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, G.; Yang, F.; Cao, W.; Mao, R.; Du, X.; Zhang, X.; Li, C.; Li, D.; Zhang, K.; et al. Foot-and-Mouth Disease Virus Viroporin 2B Antagonizes RIG-I-Mediated Antiviral Effects by Inhibition of Its Protein Expression. J. Virol. 2016, 90, 11106–11121. [Google Scholar] [CrossRef] [PubMed]
- Li, P.F.; Zhang, X.L.; Cao, W.J.; Yang, F.; Du, X.L.; Shi, Z.W.; Zhang, M.T.; Liu, X.T.; Zhu, Z.X.; Zheng, H.X. RIG-I is responsible for activation of type I interferon pathway in Seneca Valley virus-infected porcine cells to suppress viral replication. Virol. J. 2018, 15, 9. [Google Scholar] [CrossRef]
- Rojas, J.M.; Sevilla, N.; Martin, V. PPRV-induced immunosuppression at the interface of virus-host interaction. Br. J. Virol. 2016, 3, 140. [Google Scholar] [CrossRef]
- Kingston, R.L.; Baase, W.A.; Gay, L.S. Characterization of nucleocapsid binding by the measles virus and mumps virus phosphoproteins. J. Virol. 2004, 78, 8630–8640. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, C.; Spehner, D.; Drillien, R.; Neubert, W.J.; Homann, H.E. The conserved N-terminal region of Sendai virus nucleocapsid protein NP is required for nuclocapsid assembly. J. Virol. 1993, 167, 5803–5812. [Google Scholar] [CrossRef]
- Longhi, S.; Receveur-Brechot, V.; Karlin, D.; Johansson, K.; Darbon, H.; Bhella, D.; Yeo, R.; Finet, S.; Canard, B. The C-terminal domain of the measles virus nucleoprotein is intrinsically disordered and folds upon binding to the C-terminal moiety of the phosphoprotein. J. Biol. Chem. 2003, 278, 18638–18648. [Google Scholar] [CrossRef] [PubMed]
- Chinnakannan, S.K.; Holzer, B.; Bernardo, B.S.; Nanda, S.K.; Baron, M.D. Different functions of the common P/V/W and V-specific domains of rinderpest virus V protein in blocking IFN signalling. J. Gen. Virol. 2014, 95, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Devaux, P.; von Messling, V.; Songsungthong, W.; Springfeld, C.; Cattaneo, R. Tyrosine 110 in the measles virus phosphoprotein is required to block STAT1 phosphorylation. Virology 2007, 360, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Palosaari, H.; Parisien, J.P.; Rodriguez, J.J.; Ulane, C.M.; Horvath, C.M. STAT protein interference and suppression of cytokine signal transduction by measles virus V protein. J. Virol. 2003, 77, 7635–7644. [Google Scholar] [CrossRef]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Bernardo, B.S.; Goodbourn, S.; Baron, M.D. Control of the induction of type I interferon by Peste des petits ruminants virus. PLoS ONE 2017, 12, 28. [Google Scholar] [CrossRef]
- Ding, Z.; Fang, L.R.; Jing, H.Y.; Zeng, S.L.; Wang, D.; Liu, L.Z.; Zhang, H.; Luo, R.; Chen, H.C.; Xiao, S.B. Porcine Epidemic Diarrhea Virus Nucleocapsid Protein Antagonizes Beta Interferon Production by Sequestering the Interaction between IRF3 and TBK1. J. Virol. 2014, 88, 8936–8945. [Google Scholar] [CrossRef]
- Ding, Z.; Fang, L.; Yuan, S.; Zhao, L.; Wang, X.; Long, S.; Wang, M.; Wang, D.; Foda, M.; Xiao, S. The nucleocapsid proteins of mouse hepatitis virus and severe acute respiratory syndrome coronavirus share the same IFN-β antagonizing mechanism: Attenuation of PACT-mediated RIG-I/MDA5 activation. Oncotarget 2017, 8, 49655–49670. [Google Scholar] [CrossRef]
- Hu, Y.; Li, W.; Gao, T.; Cui, Y.; Jin, Y.W.; Li, P.; Ma, Q.J.; Liu, X.; Cao, C. The Severe Acute Respiratory Syndrome Coronavirus Nucleocapsid Inhibits Type I Interferon Production by Interfering with TRIM25-Mediated RIG-I Ubiquitination. J. Virol. 2017, 91, 15. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.J.; Huang, Q.F.; Liu, X.Y.; Da, D.; Liang, Y.Y.; Ly, H. Arenaviral Nucleoproteins Suppress PACT-Induced Augmentation of RIG-I Function to Inhibit Type I Interferon Production. J. Virol. 2018, 92, 11. [Google Scholar] [CrossRef] [PubMed]
- Liston, P.; Batal, R.; DiFlumeri, C.; Briedis, D.J. Protein interaction domains of the measles virus nucleocapsid protein (NP). Arch. Virol. 1997, 142, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Vidy, A.; Chelbi-Alix, M.; Blondel, D. Rabies virus P protein interacts with STAT1 and inhibits interferon signal transduction pathways. J. Virol. 2005, 79, 14411–14420. [Google Scholar] [CrossRef] [PubMed]
- Devaux, P.; Priniski, L.; Cattaneo, R. The measles virus phosphoprotein interacts with the linker domain of STAT1. Virology 2013, 444, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Chinnakannan, S.K.; Nanda, S.K.; Baron, M.D. Morbillivirus v proteins exhibit multiple mechanisms to block type 1 and type 2 interferon signalling pathways. PLoS ONE 2013, 8, e57063. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.M.; Lu, H.; Weber, M.J.; Thorner, M.O. Differential tyrosine phosphorylation of JAK1, JAK2, and STAT1 by growth hormone and interferon-gamma in IM-9 cells. J. Biol. Chem. 1994, 269, 27532–27539. [Google Scholar]
- Ohno, S.; Ono, N.; Takeda, M.; Takeuchi, K.; Yanagi, Y. Dissection of measles virus V protein in relation to its ability to block alpha/beta interferon signal transduction. J. Gen. Virol. 2004, 85, 2991–2999. [Google Scholar] [CrossRef]
- Rothlisberger, A.; Wiener, D.; Schweizer, M.; Peterhans, E.; Zurbriggen, A.; Plattet, P. Two domains of the V protein of virulent canine distemper virus selectively inhibit STAT1 and STAT2 nuclear import. J. Virol. 2010, 84, 6328–6343. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers (5′ → 3′) |
|---|---|
| JAK1 | Forward: ATTTGCGGCCGCATGGCTTTCTGTGCTAAAATG |
| Reverse: CGCGATATCTTTTAAAAGTGCTTCAAATCCTTC | |
| JAK2 | Forward: ATTTGCGGCCGCATGGGGATGGCTTGCCTTACG |
| Reverse: GCGGATATCTCCAGCCATGTTATCCCTTACTTG | |
| STAT1 | Forward: ATTTGCGGCCGCATGTCTCAGTGGTACGAACTT |
| Reverse: GCGGATATCTACTGTGTTCATCATACTGTC | |
| STAT2 | Forward: ATTTGCGGCCGCATGGCGCAGTGGGAAATGCTGC |
| Reverse: GCGGATATCGAAGTCAGAAGGCATCAAGGGTCC | |
| TYK2 | Forward: ATTTGCGGCCGCATGCCTCTGCGCCACTGGGG |
| Reverse: GCGGATATCGCACACGCTGAACACTGAAGGGGC | |
| IRF9 | Forward: ATTTGCGGCCGCATGGCATCAGGCAGGGCACGCT |
| Reverse: GCGGATATCCACCAGGGACAGAATGGCTGCCTG | |
| N | Forward: TTGCGGCCGCGATGGCGACTCTCCTTAAAAGCT |
| Reverse: TTGTCGACTCAGCCGAGGAGATCCTTGTCGTT | |
| P | Forward: TTGCGGCCGCGATGGCAGAAGAACAAGCATACCAT |
| Reverse: GCGATATCTTACGGCTGCTTGGCAAGAATGGCTGTTA | |
| C | Forward: CTGAATTCAATGTCAACAAGGGACTGGAA |
| Reverse: GCCTCTAGACTAATTTTTCGACATCTGTTGACCT | |
| M | Forward: GCAAGCTTATGACCGAGATCTACGA |
| Reverse: TGGATCCTTACAGGATCTTGAACAGGCCTTGAT | |
| H | Forward: TTGCGGCCGCGATGTCCGCACAAAGGGAAAGGAT |
| Reverse: GCTGGTACCTCAGACTGGATTACATGTTACCTCTATACC | |
| F | Forward: TTGCGGCCGCGATGACACGGGTCGCAACCTTAGTATTT |
| Reverse: TGGATCCCTACAGTGATCTCACGTACGACT | |
| P-Y110H | Forward: AACTCTCAAGTACAGCGTCACTATGTTTATAGCCACGGG |
| Reverse: CCCGTGGCTATAAACATAGTGACGCTGTACTTGAGAGTT | |
| P-Y110F | Forward: AACTCTCAAGTACAGCGTTTCTATGTTTATAGCCACGGG |
| Reverse: CCCGTGGCTATAAACATAGAAACGCTGTACTTGAGAGTT | |
| N-core | Forward: TTGCGGCCGCGATGGCGACTCTCCTTAAAAGCT |
| Reverse: GCGGATATCCTACTTTCCTGCAGATCTTCTG | |
| N-tail | Forward: ATTTGCGGCCGCGTCAGCTCTGTAATCGCGGC |
| Reverse: TTGTCGACTCAGCCGAGGAGATCCTTGTCGTT |
| Gene | Primers (5′→3′) |
|---|---|
| GAPDH | Forward: CGGGAAGCTTGTGATCAATGG |
| Reverse: GGCAGTGATGGCATGGACTG | |
| ISG54 | Forward: GGTCTCTTCAGCATTTATTGGTG |
| Reverse: TGCCGTAGGCTGCTCTCCA | |
| ISG15 | Forward: TGGACAAATGCGACGAACC |
| Reverse: CCCGCTCACTTGCTGCTT | |
| MXA | Forward: TCTTCATGCTCCAGACGTAC |
| Reverse: CCAGCTGTAGGTGTCCTTG | |
| OAS1 | Forward: TGTCCAAGGTGGTAAAGGGTG |
| Reverse: CCGGCGATTTAACTGATCCTG | |
| IRF1 | Forward: GAGGAGGTGAAAGACCAGAGCA |
| Reverse: TAGCATCTCGGCTGGACTTCGA | |
| STAT1 | Forward: ATGGCAGTCTGGCGGCTGAATT |
| Reverse: CCAAACCAGGCTGGCACAATTG |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, P.; Zhu, Z.; Zhang, X.; Dang, W.; Li, L.; Du, X.; Zhang, M.; Wu, C.; Xue, Q.; Liu, X.; et al. The Nucleoprotein and Phosphoprotein of Peste des Petits Ruminants Virus Inhibit Interferons Signaling by Blocking the JAK-STAT Pathway. Viruses 2019, 11, 629. https://doi.org/10.3390/v11070629
Li P, Zhu Z, Zhang X, Dang W, Li L, Du X, Zhang M, Wu C, Xue Q, Liu X, et al. The Nucleoprotein and Phosphoprotein of Peste des Petits Ruminants Virus Inhibit Interferons Signaling by Blocking the JAK-STAT Pathway. Viruses. 2019; 11(7):629. https://doi.org/10.3390/v11070629
Chicago/Turabian StyleLi, Pengfei, Zixiang Zhu, Xiangle Zhang, Wen Dang, Linlin Li, Xiaoli Du, Miaotao Zhang, Chunyan Wu, Qinghong Xue, Xiangtao Liu, and et al. 2019. "The Nucleoprotein and Phosphoprotein of Peste des Petits Ruminants Virus Inhibit Interferons Signaling by Blocking the JAK-STAT Pathway" Viruses 11, no. 7: 629. https://doi.org/10.3390/v11070629
APA StyleLi, P., Zhu, Z., Zhang, X., Dang, W., Li, L., Du, X., Zhang, M., Wu, C., Xue, Q., Liu, X., Zheng, H., & Nan, Y. (2019). The Nucleoprotein and Phosphoprotein of Peste des Petits Ruminants Virus Inhibit Interferons Signaling by Blocking the JAK-STAT Pathway. Viruses, 11(7), 629. https://doi.org/10.3390/v11070629