Twenty-Five Years of Structural Parvovirology

Abstract
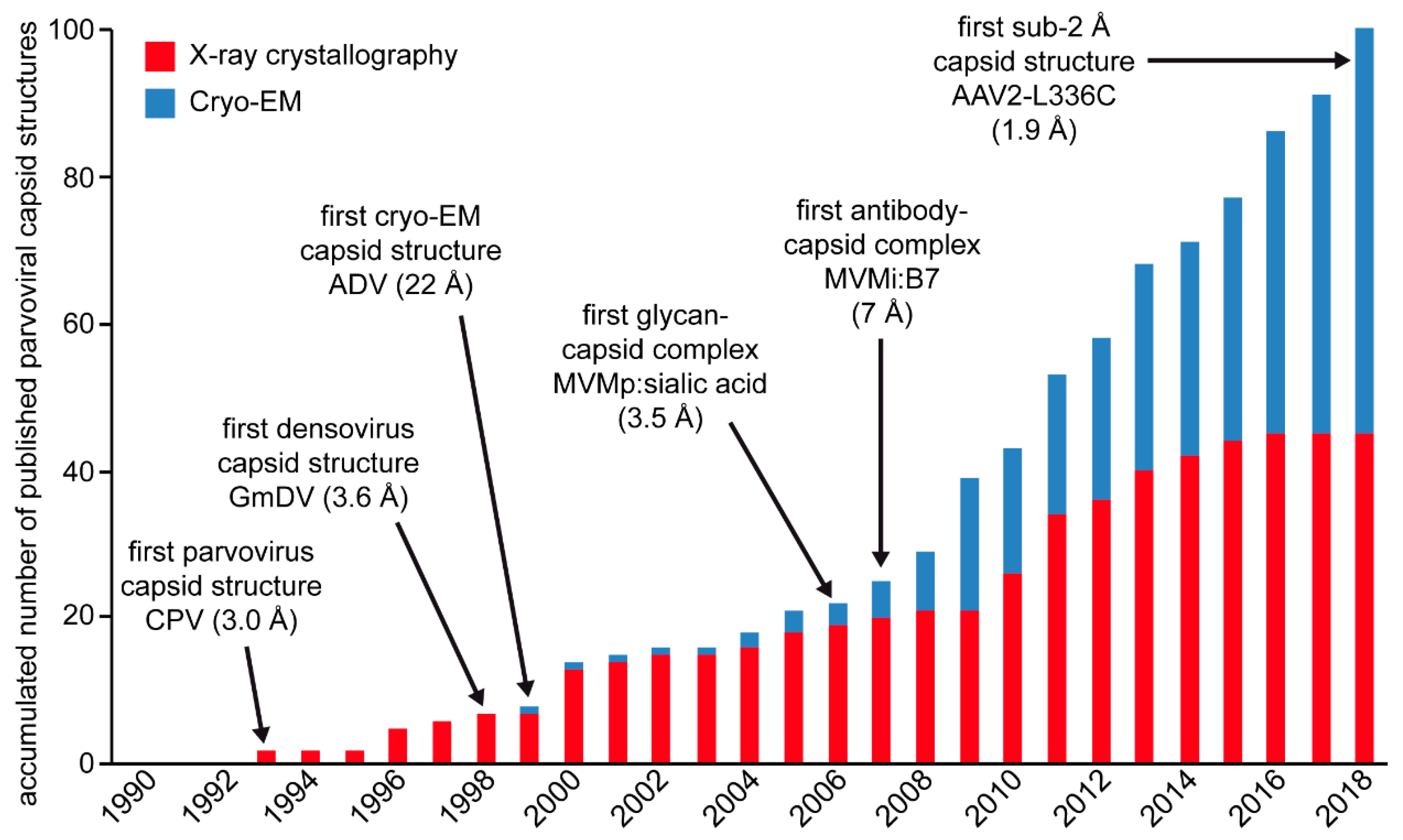
1. Introduction
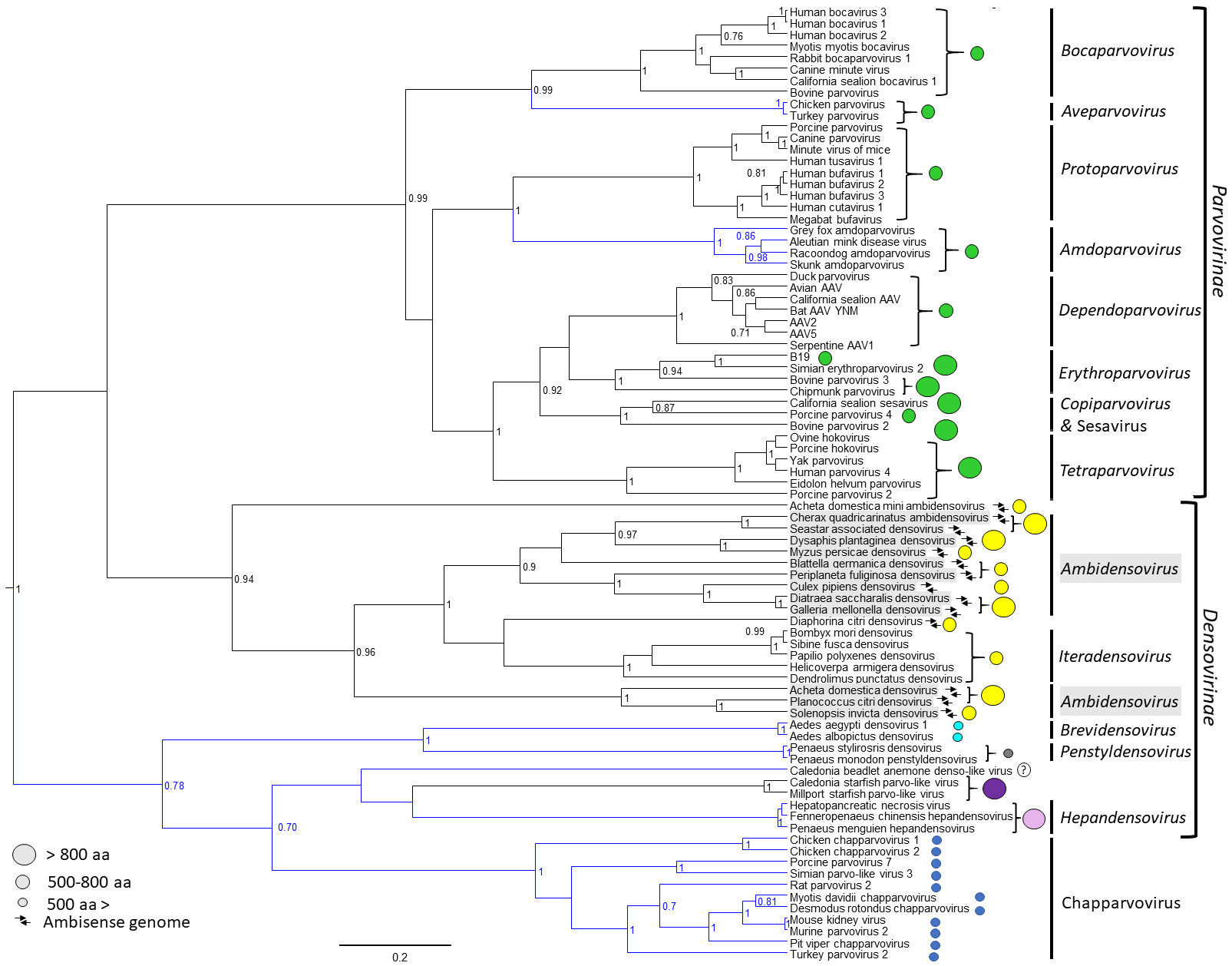
2. Parvovirinae
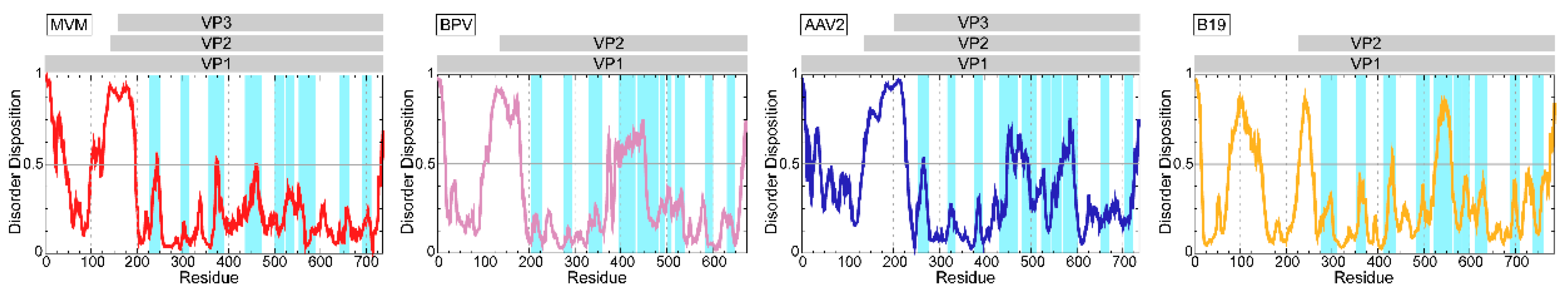
2.1. Expression of the Parvovirinae VPs Utilizes Different Transcription Strategies
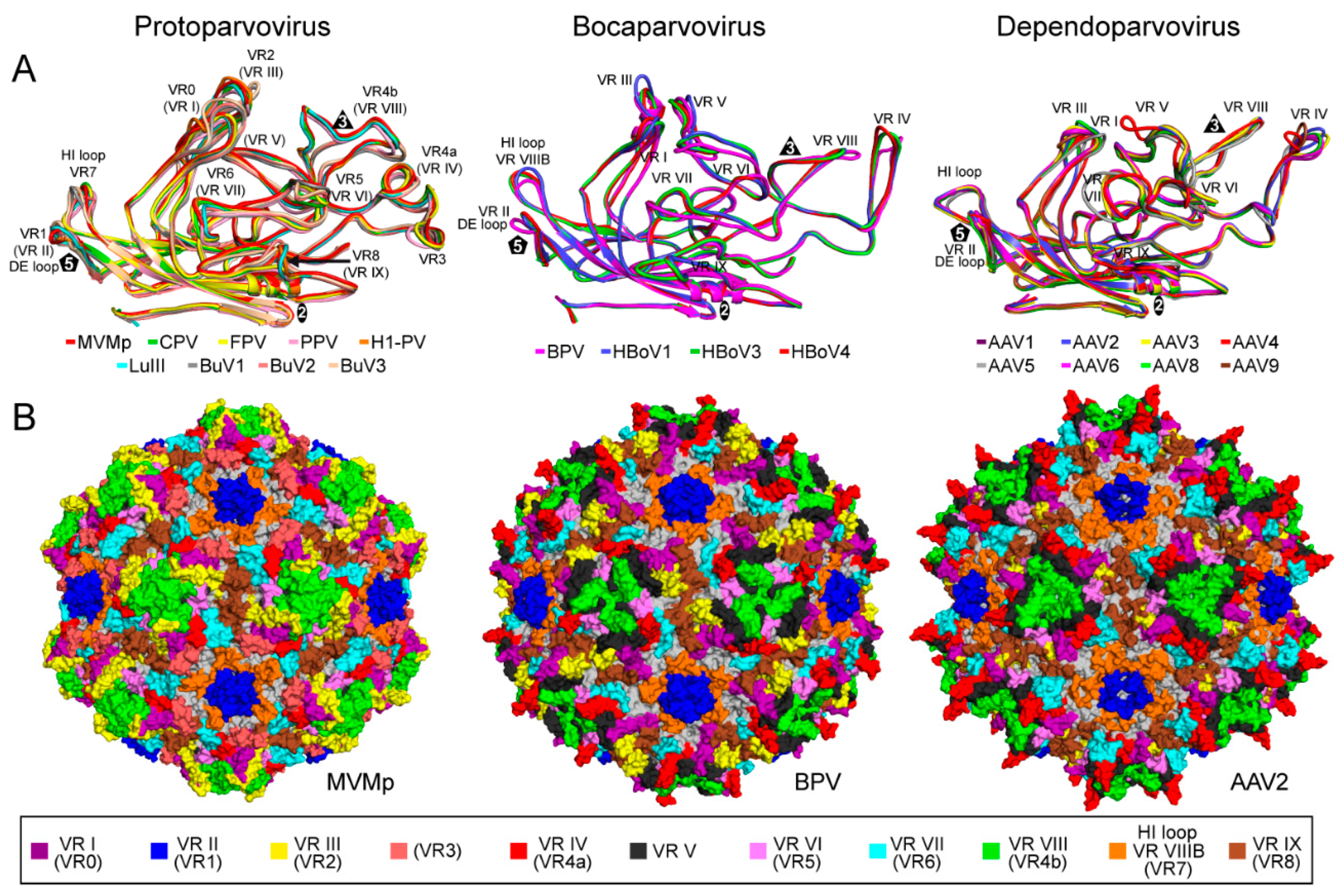
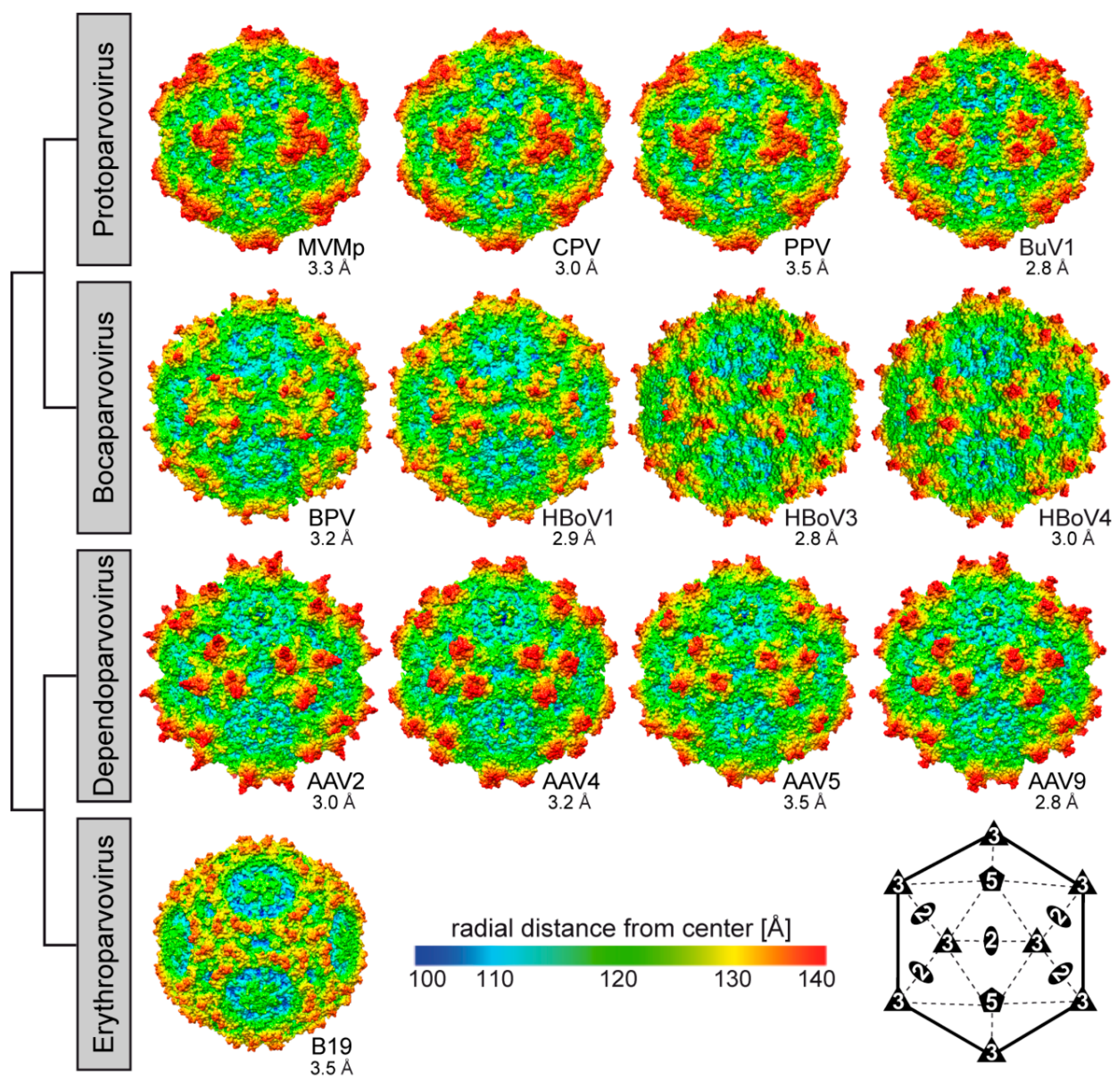
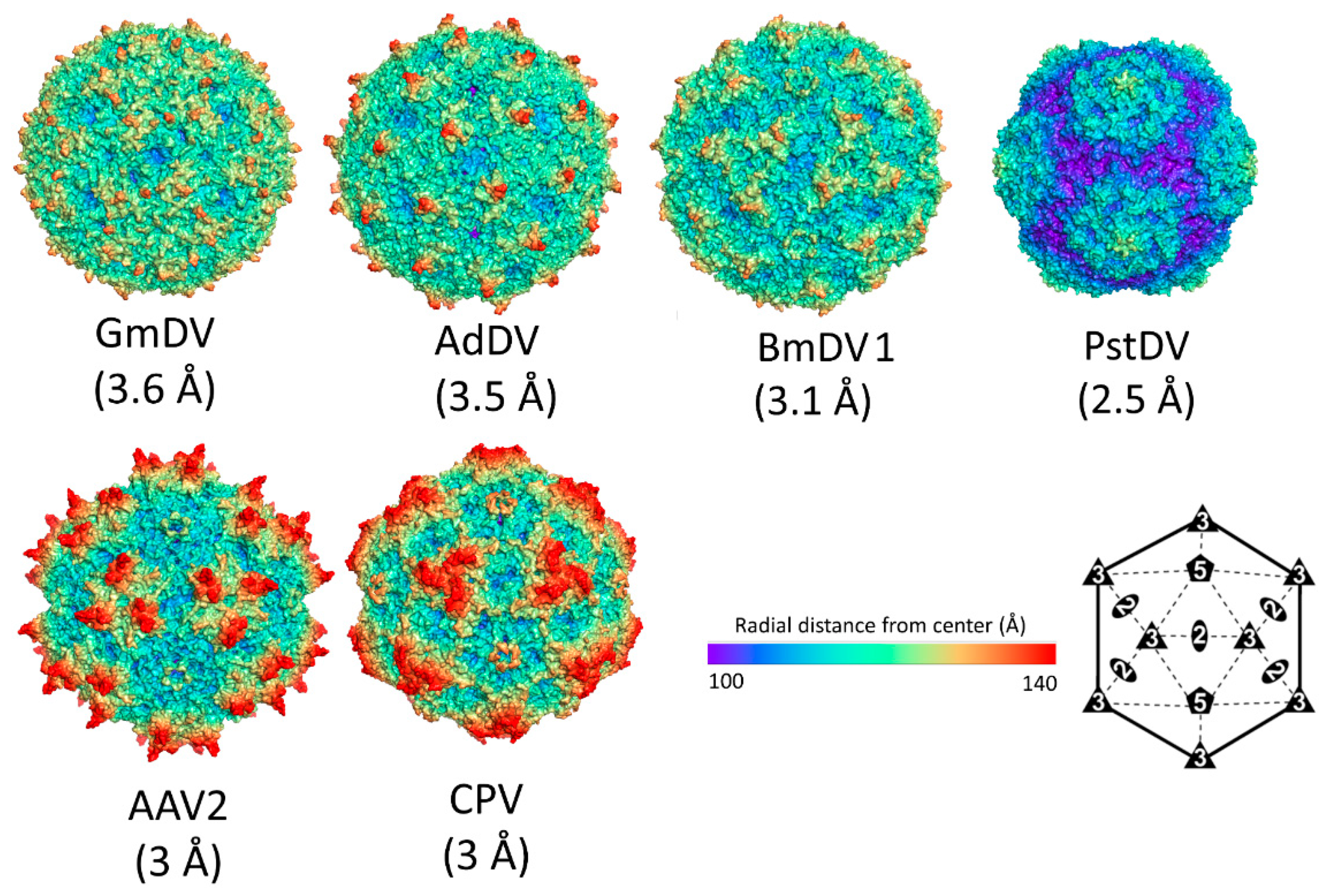
2.2. Parvovirinae Genera Display Distinct Capsid Surface Morphologies
2.3. Nucleotides Are Ordered Inside Parvovirinae Capsids Despite Lack of Icosahedral Symmetry
2.4. The Structure of Capsid Variants Provide Insight into Function
2.5. Capsid-Receptor Complex Structures
2.6. Capsid-Antibody Complex Structures
2.7. Engineering of Parvovirus Capsids to Create Biologics
3. Densovirinae
3.1. The Densovirinae Utilize Diverse Strategies for VP Expression
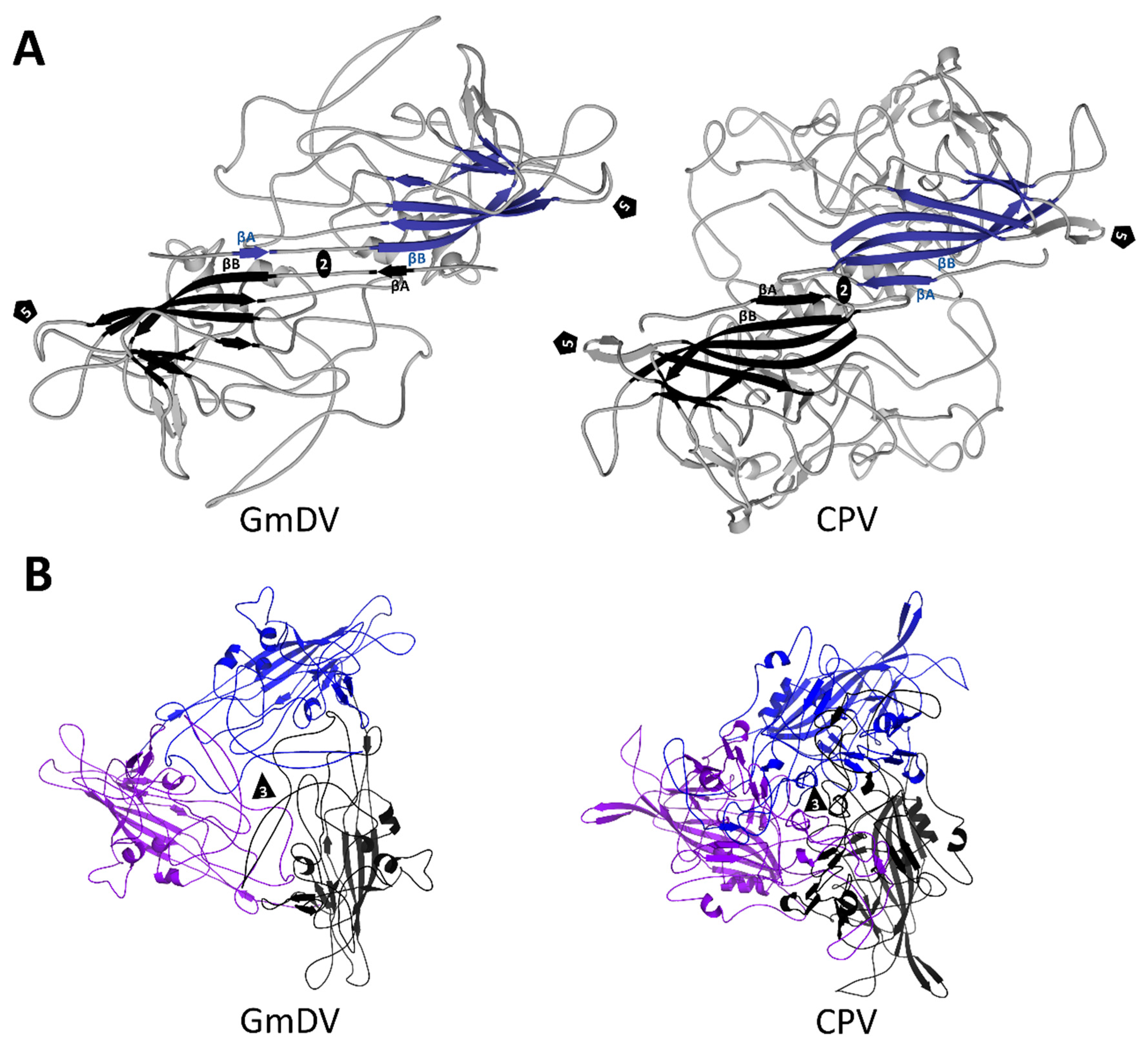
3.2. Densovirinae Capsid Structures Display Distinct Surface Morphology
3.3. Functions Associated with Densoviral Capsid Proteins
4. Summary
Funding
Conflicts of Interest
References
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Penzes, J.J.; et al. ICTV virus taxonomy profile: Parvoviridae. J. Gen. Virol. 2019, 100, 367–368. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Bu, W.; Bhatia, S.; Hare, J.; Somasundaram, T.; Azzi, A.; Chapman, M.S. The atomic structure of adeno-associated virus (AAV-2), a vector for human gene therapy. Proc. Natl. Acad. Sci. USA 2002, 99, 10405–10410. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, B.; Simpson, A.A.; Rossmann, M.G. The structure of human parvovirus B19. Proc. Natl. Acad. Sci. USA 2004, 101, 11628–11633. [Google Scholar] [CrossRef]
- Kailasan, S.; Halder, S.; Gurda, B.; Bladek, H.; Chipman, P.R.; McKenna, R.; Brown, K.; Agbandje-McKenna, M. Structure of an enteric pathogen, bovine parvovirus. J. Virol. 2015, 89, 2603–2614. [Google Scholar] [CrossRef] [PubMed]
- Tsao, J.; Chapman, M.S.; Agbandje, M.; Keller, W.; Smith, K.; Wu, H.; Luo, M.; Smith, T.J.; Rossmann, M.G.; Compans, R.W.; et al. The three-dimensional structure of canine parvovirus and its functional implications. Science 1991, 251, 1456–1464. [Google Scholar] [CrossRef]
- Simpson, A.A.; Chipman, P.R.; Baker, T.S.; Tijssen, P.; Rossmann, M.G. The structure of an insect parvovirus (Galleria mellonella densovirus) at 3.7 A resolution. Structure 1998, 6, 1355–1367. [Google Scholar] [CrossRef]
- Meng, G.; Zhang, X.; Plevka, P.; Yu, Q.; Tijssen, P.; Rossmann, M.G. The structure and host entry of an invertebrate parvovirus. J. Virol. 2013, 87, 12523–12530. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, B.; El-Far, M.; Plevka, P.; Bowman, V.D.; Li, Y.; Tijssen, P.; Rossmann, M.G. Structure of Bombyx mori densovirus 1, a silkworm pathogen. J. Virol. 2011, 85, 4691–4697. [Google Scholar] [CrossRef] [PubMed]
- Zadori, Z.; Szelei, J.; Lacoste, M.C.; Li, Y.; Gariepy, S.; Raymond, P.; Allaire, M.; Nabi, I.R.; Tijssen, P. A viral phospholipase A2 is required for parvovirus infectivity. Dev. Cell 2001, 1, 291–302. [Google Scholar] [CrossRef]
- Popa-Wagner, R.; Sonntag, F.; Schmidt, K.; King, J.; Kleinschmidt, J.A. Nuclear translocation of adeno-associated virus type 2 capsid proteins for virion assembly. J. Gen. Virol. 2012, 93, 1887–1898. [Google Scholar] [CrossRef]
- Popa-Wagner, R.; Porwal, M.; Kann, M.; Reuss, M.; Weimer, M.; Florin, L.; Kleinschmidt, J.A. Impact of vp1-specific protein sequence motifs on adeno-associated virus type 2 intracellular trafficking and nuclear entry. J. Virol. 2012, 86, 9163–9174. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.W.; Liu, W.P.; Qi, Z.Y.; Duan, Z.J.; Zheng, L.S.; Kuang, Z.Z.; Zhang, W.J.; Hou, Y.D. Phospholipase A2-like activity of human bocavirus VP1 unique region. Biochem. Biophys. Res Commun. 2008, 365, 158–163. [Google Scholar] [CrossRef]
- Buller, R.M.; Janik, J.E.; Sebring, E.D.; Rose, J.A. Herpes simplex virus types 1 and 2 completely help adenovirus-associated virus replication. J. Virol. 1981, 40, 241–247. [Google Scholar] [PubMed]
- McPherson, R.A.; Rosenthal, L.J.; Rose, J.A. Human cytomegalovirus completely helps adeno-associated virus replication. Virology 1985, 147, 217–222. [Google Scholar] [CrossRef]
- Weindler, F.W.; Heilbronn, R. A subset of herpes simplex virus replication genes provides helper functions for productive adeno-associated virus replication. J. Virol. 1991, 65, 2476–2483. [Google Scholar] [PubMed]
- Chang, L.S.; Shi, Y.; Shenk, T. Adeno-associated virus P5 promoter contains an adenovirus E1A-inducible element and a binding site for the major late transcription factor. J. Virol. 1989, 63, 3479–3488. [Google Scholar]
- Weitzman, M.D.; Linden, R.M. Adeno-associated virus biology. Methods Mol. Biol. 2011, 807, 1–23. [Google Scholar] [PubMed]
- Laughlin, C.A.; Westphal, H.; Carter, B.J. Spliced adenovirus-associated virus rna. Proc. Natl. Acad. Sci. USA 1979, 76, 5567–5571. [Google Scholar] [CrossRef]
- Qiu, J.; Soderlund-Venermo, M.; Young, N.S. Human parvoviruses. Clin. Microbiol. Rev. 2017, 30, 43–113. [Google Scholar]
- Becerra, S.P.; Rose, J.A.; Hardy, M.; Baroudy, B.M.; Anderson, C.W. Direct mapping of adeno-associated virus capsid proteins b and c: A possible ACG initiation codon. Proc. Natl. Acad. Sci. USA 1985, 82, 7919–7923. [Google Scholar] [CrossRef]
- Becerra, S.P.; Koczot, F.; Fabisch, P.; Rose, J.A. Synthesis of adeno-associated virus structural proteins requires both alternative mrna splicing and alternative initiations from a single transcript. J. Virol. 1988, 62, 2745–2754. [Google Scholar]
- Trempe, J.P.; Carter, B.J. Alternate mrna splicing is required for synthesis of adeno-associated virus Vp1 capsid protein. J. Virol. 1988, 62, 3356–3363. [Google Scholar] [PubMed]
- Qiu, J.; Pintel, D. Processing of adeno-associated virus rna. Front. Biosci. 2008, 13, 3101–3115. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Yoto, Y.; Tullis, G.E.; Pintel, D. Parvovirus RNA processing strategies. In Parvoviruses; Edward Arnold, Ltd.: London, UK, 2006; pp. 253–274. [Google Scholar]
- Wistuba, A.; Kern, A.; Weger, S.; Grimm, D.; Kleinschmidt, J.A. Subcellular compartmentalization of adeno-associated virus type 2 assembly. J. Virol. 1997, 71, 1341–1352. [Google Scholar]
- Cotmore, S.F.; D’Abramo, A.M., Jr.; Carbonell, L.F.; Bratton, J.; Tattersall, P. The NS2 polypeptide of parvovirus MVM is required for capsid assembly in murine cells. Virology 1997, 231, 267–280. [Google Scholar] [CrossRef] [PubMed]
- Cotmore, S.F.; McKie, V.C.; Anderson, L.J.; Astell, C.R.; Tattersall, P. Identification of the major structural and nonstructural proteins encoded by human parvovirus B19 and mapping of their genes by procaryotic expression of isolated genomic fragments. J. Virol. 1986, 60, 548–557. [Google Scholar]
- Buller, R.M.; Rose, J.A. Characterization of adenovirus-associated virus-induced polypeptides in KB cells. J. Virol. 1978, 25, 331–338. [Google Scholar] [PubMed]
- Snijder, J.; van de Waterbeemd, M.; Damoc, E.; Denisov, E.; Grinfeld, D.; Bennett, A.; Agbandje-McKenna, M.; Makarov, A.; Heck, A.J. Defining the stoichiometry and cargo load of viral and bacterial nanoparticles by orbitrap mass spectrometry. J. Am. Chem. Soc. 2014, 136, 7295–7299. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.A.; Maizel, J.V., Jr.; Inman, J.K.; Shatkin, A.J. Structural proteins of adenovirus-associated viruses. J. Virol. 1971, 8, 766–770. [Google Scholar]
- Sonntag, F.; Schmidt, K.; Kleinschmidt, J.A. A viral assembly factor promotes AAV2 capsid formation in the nucleolus. Proc. Natl. Acad. Sci. USA 2010, 107, 10220–10225. [Google Scholar] [CrossRef]
- Bleker, S.; Pawlita, M.; Kleinschmidt, J.A. Impact of capsid conformation and rep-capsid interactions on adeno-associated virus type 2 genome packaging. J. Virol. 2006, 80, 810–820. [Google Scholar] [CrossRef]
- Clinton, G.M.; Hayashi, M. The parovivirus MVM: Particles with altered structural proteins. Virology 1975, 66, 261–267. [Google Scholar] [CrossRef]
- Weichert, W.S.; Parker, J.S.; Wahid, A.T.; Chang, S.F.; Meier, E.; Parrish, C.R. Assaying for structural variation in the parvovirus capsid and its role in infection. Virology 1998, 250, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Paradiso, P.R. Infectious process of the parvovirus h-1: Correlation of protein content, particle density, and viral infectivity. J. Virol. 1981, 39, 800–807. [Google Scholar] [PubMed]
- Farr, G.A.; Cotmore, S.F.; Tattersall, P. Vp2 cleavage and the leucine ring at the base of the fivefold cylinder control ph-dependent externalization of both the Vp1 N terminus and the genome of minute virus of mice. J. Virol. 2006, 80, 161–171. [Google Scholar] [CrossRef]
- Wu, H.; Rossmann, M.G. The canine parvovirus empty capsid structure. J. Mol. Biol. 1993, 233, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Ilyas, M.; Mietzsch, M.; Kailasan, S.; Vaisanen, E.; Luo, M.; Chipman, P.; Smith, J.K.; Kurian, J.; Sousa, D.; McKenna, R.; et al. Atomic resolution structures of human bufaviruses determined by Cryo-electron microscopy. Viruses 2018, 10, 22. [Google Scholar] [CrossRef]
- Xie, Q.; Chapman, M.S. Canine parvovirus capsid structure, analyzed at 2.9 A resolution. J. Mol. Biol. 1996, 264, 497–520. [Google Scholar] [CrossRef]
- Govindasamy, L.; Hueffer, K.; Parrish, C.R.; Agbandje-McKenna, M. Structures of host range-controlling regions of the capsids of canine and feline parvoviruses and mutants. J. Virol. 2003, 77, 12211–12221. [Google Scholar] [CrossRef]
- Simpson, A.A.; Chandrasekar, V.; Hebert, B.; Sullivan, G.M.; Rossmann, M.G.; Parrish, C.R. Host range and variability of calcium binding by surface loops in the capsids of canine and feline parvoviruses. J. Mol. Biol. 2000, 300, 597–610. [Google Scholar] [CrossRef]
- Llamas-Saiz, A.L.; Agbandje-McKenna, M.; Parker, J.S.; Wahid, A.T.; Parrish, C.R.; Rossmann, M.G. Structural analysis of a mutation in canine parvovirus which controls antigenicity and host range. Virology 1996, 225, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Organtini, L.J.; Allison, A.B.; Lukk, T.; Parrish, C.R.; Hafenstein, S. Global displacement of canine parvovirus by a host-adapted variant: Structural comparison between pandemic viruses with distinct host ranges. J. Virol. 2015, 89, 1909–1912. [Google Scholar] [CrossRef] [PubMed]
- Agbandje, M.; McKenna, R.; Rossmann, M.G.; Strassheim, M.L.; Parrish, C.R. Structure determination of feline Panleukopenia virus empty particles. Proteins 1993, 16, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.; Nam, H.J.; Govindasamy, L.; Vogel, M.; Dinsart, C.; Salome, N.; McKenna, R.; Agbandje-McKenna, M. Structural characterization of H-1 parvovirus: Comparison of infectious virions to empty capsids. J. Virol. 2013, 87, 5128–5140. [Google Scholar] [CrossRef]
- Pittman, N.; Misseldine, A.; Geilen, L.; Halder, S.; Smith, J.K.; Kurian, J.; Chipman, P.; Janssen, M.; McKenna, R.; Baker, T.S.; et al. Atomic resolution structure of the oncolytic parvovirus luiii by electron microscopy and 3d image reconstruction. Viruses 2017, 9, 321. [Google Scholar] [CrossRef] [PubMed]
- Callaway, H.M.; Subramanian, S.; Urbina, C.; Barnard, K.; Dick, R.; Bator, C.M.; Hafentein, S.L.; Gifford, R.J.; Parrish, C.R. Examination and reconstruction of three ancient endogenous parvovirus capsid protein gene remnants found in rodent genomes. J. Virol. 2019, 93, e01542-18. [Google Scholar] [CrossRef]
- Llamas-Saiz, A.L.; Agbandje-McKenna, M.; Wikoff, W.R.; Bratton, J.; Tattersall, P.; Rossmann, M.G. Structure determination of minute virus of mice. Acta Crystallogr. Sect. D Biol. Crystallogr. 1997, 53, 93–102. [Google Scholar] [CrossRef]
- Kontou, M.; Govindasamy, L.; Nam, H.J.; Bryant, N.; Llamas-Saiz, A.L.; Foces-Foces, C.; Hernando, E.; Rubio, M.P.; McKenna, R.; Almendral, J.M.; et al. Structural determinants of tissue tropism and in vivo pathogenicity for the parvovirus minute virus of mice. J. Virol. 2005, 79, 10931–10943. [Google Scholar] [CrossRef]
- Plevka, P.; Hafenstein, S.; Li, L.; D’Abrgamo, A., Jr.; Cotmore, S.F.; Rossmann, M.G.; Tattersall, P. Structure of a packaging-defective mutant of minute virus of mice indicates that the genome is packaged via a pore at a 5-fold axis. J. Virol. 2011, 85, 4822–4827. [Google Scholar] [CrossRef]
- Guerra, P.; Valbuena, A.; Querol-Audi, J.; Silva, C.; Castellanos, M.; Rodriguez-Huete, A.; Garriga, D.; Mateu, M.G.; Verdaguer, N. Structural basis for biologically relevant mechanical stiffening of a virus capsid by cavity-creating or spacefilling mutations. Sci. Rep. 2017, 7, 4101. [Google Scholar] [CrossRef] [PubMed]
- Simpson, A.A.; Hebert, B.; Sullivan, G.M.; Parrish, C.R.; Zadori, Z.; Tijssen, P.; Rossmann, M.G. The structure of porcine parvovirus: Comparison with related viruses. J. Mol. Biol. 2002, 315, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Mietzsch, M.; Kailasan, S.; Garrison, J.; Ilyas, M.; Chipman, P.; Kantola, K.; Janssen, M.E.; Spear, J.; Sousa, D.; McKenna, R.; et al. Structural insights into human bocaparvoviruses. J. Virol. 2017, 91, e00261-17. [Google Scholar] [CrossRef] [PubMed]
- Ng, R.; Govindasamy, L.; Gurda, B.L.; McKenna, R.; Kozyreva, O.G.; Samulski, R.J.; Parent, K.N.; Baker, T.S.; Agbandje-McKenna, M. Structural characterization of the dual glycan binding adeno-associated virus serotype 6. J. Virol. 2010, 84, 12945–12957. [Google Scholar] [CrossRef] [PubMed]
- Drouin, L.M.; Lins, B.; Janssen, M.; Bennett, A.; Chipman, P.; McKenna, R.; Chen, W.; Muzyczka, N.; Cardone, G.; Baker, T.S.; et al. Cryo-electron microscopy reconstruction and stability studies of the wild type and the R432A variant of adeno-associated virus type 2 reveal that capsid structural stability is a major factor in genome packaging. J. Virol. 2016, 90, 8542–8551. [Google Scholar] [CrossRef]
- Tan, Y.Z.; Aiyer, S.; Mietzsch, M.; Hull, J.A.; McKenna, R.; Grieger, J.; Samulski, R.J.; Baker, T.S.; Agbandje-McKenna, M.; Lyumkis, D. Sub-2 A Ewald curvature corrected structure of an AAV2 capsid variant. Nat. Commun. 2018, 9, 3628. [Google Scholar] [CrossRef] [PubMed]
- Burg, M.; Rosebrough, C.; Drouin, L.M.; Bennett, A.; Mietzsch, M.; Chipman, P.; McKenna, R.; Sousa, D.; Potter, M.; Byrne, B.; et al. Atomic structure of a rationally engineered gene delivery vector, aav2.5. J. Struct. Biol. 2018, 203, 236–241. [Google Scholar] [CrossRef]
- Lerch, T.F.; Xie, Q.; Chapman, M.S. The structure of adeno-associated virus serotype 3b (aav-3b): Insights into receptor binding and immune evasion. Virology 2010, 403, 26–36. [Google Scholar] [CrossRef]
- Govindasamy, L.; Padron, E.; McKenna, R.; Muzyczka, N.; Kaludov, N.; Chiorini, J.A.; Agbandje-McKenna, M. Structurally mapping the diverse phenotype of adeno-associated virus serotype 4. J. Virol. 2006, 80, 11556–11570. [Google Scholar] [CrossRef]
- Govindasamy, L.; Dimattia, M.A.; Gurda, B.L.; Halder, S.; McKenna, R.; Chiorini, J.A.; Muzyczka, N.; Zolotukhin, S.; Agbandje-McKenna, M. Structural insights into adeno-associated virus serotype 5. J. Virol. 2013, 87, 11187–11199. [Google Scholar] [CrossRef]
- Xie, Q.; Lerch, T.F.; Meyer, N.L.; Chapman, M.S. Structure-function analysis of receptor-binding in adeno-associated virus serotype 6 (AAV-6). Virology 2011, 420, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Nam, H.J.; Lane, M.D.; Padron, E.; Gurda, B.; McKenna, R.; Kohlbrenner, E.; Aslanidi, G.; Byrne, B.; Muzyczka, N.; Zolotukhin, S.; et al. Structure of adeno-associated virus serotype 8, a gene therapy vector. J. Virol. 2007, 81, 12260–12271. [Google Scholar] [CrossRef]
- Nam, H.J.; Gurda, B.L.; McKenna, R.; Potter, M.; Byrne, B.; Salganik, M.; Muzyczka, N.; Agbandje-McKenna, M. Structural studies of adeno-associated virus serotype 8 capsid transitions associated with endosomal trafficking. J. Virol. 2011, 85, 11791–11799. [Google Scholar] [CrossRef] [PubMed]
- DiMattia, M.A.; Nam, H.J.; Van Vliet, K.; Mitchell, M.; Bennett, A.; Gurda, B.L.; McKenna, R.; Olson, N.H.; Sinkovits, R.S.; Potter, M.; et al. Structural insight into the unique properties of adeno-associated virus serotype 9. J. Virol. 2012, 86, 6947–6958. [Google Scholar] [CrossRef]
- Guenther, C.M.; Brun, M.J.; Bennett, A.D.; Ho, M.L.; Chen, W.; Zhu, B.; Lam, M.; Yamagami, M.; Kwon, S.; Bhattacharya, N.; et al. Protease-activatable adeno-associated virus vector for gene delivery to damaged heart tissue. Mol. Ther. 2019, 27, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Lerch, T.F.; O’Donnell, J.K.; Meyer, N.L.; Xie, Q.; Taylor, K.A.; Stagg, S.M.; Chapman, M.S. Structure of AAV-dj, a retargeted gene therapy vector: Cryo-electron microscopy at 4.5 A resolution. Structure 2012, 20, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.; Van Vliet, K.; Smith, J.K.; Duong, T.T.; McKenna, R.; Wilson, J.M.; Agbandje-McKenna, M. Structure of neurotropic adeno-associated virus AAVrh.8. J. Struct. Biol. 2015, 192, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Mikals, K.; Nam, H.J.; Van Vliet, K.; Vandenberghe, L.H.; Mays, L.E.; McKenna, R.; Wilson, J.M.; Agbandje-McKenna, M. The structure of AAVrh32.33, a novel gene delivery vector. J. Struct. Biol. 2014, 186, 308–317. [Google Scholar] [CrossRef]
- Kronenberg, S.; Bottcher, B.; von der Lieth, C.W.; Bleker, S.; Kleinschmidt, J.A. A conformational change in the adeno-associated virus type 2 capsid leads to the exposure of hidden Vp1 N termini. J. Virol. 2005, 79, 5296–5303. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, B.; Chipman, P.R.; Kostyuchenko, V.A.; Modrow, S.; Rossmann, M.G. Visualization of the externalized Vp2 N termini of infectious human parvovirus B19. J. Virol. 2008, 82, 7306–7312. [Google Scholar] [CrossRef]
- Xue, B.; Dunbrack, R.L.; Williams, R.W.; Dunker, A.K.; Uversky, V.N. Pondr-fit: A meta-predictor of intrinsically disordered amino acids. Biochim. Biophys. Acta 2010, 1804, 996–1010. [Google Scholar] [CrossRef] [PubMed]
- Venkatakrishnan, B.; Yarbrough, J.; Domsic, J.; Bennett, A.; Bothner, B.; Kozyreva, O.G.; Samulski, R.J.; Muzyczka, N.; McKenna, R.; Agbandje-McKenna, M. Structure and dynamics of adeno-associated virus serotype 1 Vp1-unique n-terminal domain and its role in capsid trafficking. J. Virol. 2013, 87, 4974–4984. [Google Scholar] [CrossRef] [PubMed]
- Agbandje-McKenna, M.; Kleinschmidt, J. Aav capsid structure and cell interactions. Methods Mol. Biol. 2011, 807, 47–92. [Google Scholar] [PubMed]
- Bennett, A.; Mietzsch, M.; Agbandje-McKenna, M. Understanding capsid assembly and genome packaging for adeno-associated viruses. Future Virol. 2017, 12, 283–297. [Google Scholar] [CrossRef]
- Chapman, M.S.; Agbandje-McKenna, M. Atomic structure of viral particles. In Parvoviruses; Bloom, M.E., Cotmore, S.F., Linden, R.M., Parrish, C.R., Kerr, J.R., Eds.; Edward Arnold, Ltd.: London, UK, 2006; pp. 109–123. [Google Scholar]
- DeLano, W.L. The Pymol Molecular Graphics Syste; DeLano Scientific: San Carlos, CA, USA, 2002. [Google Scholar]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. Ucsf chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Chapman, M.S.; Rossmann, M.G. Single-stranded DNA-protein interactions in canine parvovirus. Structure 1995, 3, 151–162. [Google Scholar] [CrossRef]
- Grieger, J.C.; Johnson, J.S.; Gurda-Whitaker, B.; Agbandje-McKenna, M.; Samulski, R.J. Surface-exposed adeno-associated virus Vp1-NLS capsid fusion protein rescues infectivity of noninfectious wild-type Vp2/Vp3 and Vp3-only capsids but not that of fivefold pore mutant virions. J. Virol. 2007, 81, 7833–7843. [Google Scholar] [CrossRef] [PubMed]
- Farr, G.A.; Tattersall, P. A conserved leucine that constricts the pore through the capsid fivefold cylinder plays a central role in parvoviral infection. Virology 2004, 323, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Xiao, W.; Conlon, T.; Hughes, J.; Agbandje-McKenna, M.; Ferkol, T.; Flotte, T.; Muzyczka, N. Mutational analysis of the adeno-associated virus type 2 (AAV2) capsid gene and construction of aav2 vectors with altered tropism. J. Virol. 2000, 74, 8635–8647. [Google Scholar] [CrossRef]
- Hafenstein, S.; Palermo, L.M.; Kostyuchenko, V.A.; Xiao, C.; Morais, M.C.; Nelson, C.D.; Bowman, V.D.; Battisti, A.J.; Chipman, P.R.; Parrish, C.R.; et al. Asymmetric binding of transferrin receptor to parvovirus capsids. Proc. Natl. Acad. Sci. USA 2007, 104, 6585–6589. [Google Scholar] [CrossRef] [PubMed]
- Hafenstein, S.; Bowman, V.D.; Sun, T.; Nelson, C.D.; Palermo, L.M.; Chipman, P.R.; Battisti, A.J.; Parrish, C.R.; Rossmann, M.G. Structural comparison of different antibodies interacting with parvovirus capsids. J. Virol. 2009, 83, 5556–5566. [Google Scholar] [CrossRef] [PubMed]
- Hueffer, K.; Govindasamy, L.; Agbandje-McKenna, M.; Parrish, C.R. Combinations of two capsid regions controlling canine host range determine canine transferrin receptor binding by canine and feline parvoviruses. J. Virol. 2003, 77, 10099–10105. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.Y.; Halder, S.; Agbandje-McKenna, M. Parvovirus glycan interactions. Curr. Opin. Virol. 2014, 7, 108–118. [Google Scholar] [CrossRef] [PubMed]
- Pillay, S.; Meyer, N.L.; Puschnik, A.S.; Davulcu, O.; Diep, J.; Ishikawa, Y.; Jae, L.T.; Wosen, J.E.; Nagamine, C.M.; Chapman, M.S.; et al. An essential receptor for adeno-associated virus infection. Nature 2016, 530, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Mietzsch, M.; Broecker, F.; Reinhardt, A.; Seeberger, P.H.; Heilbronn, R. Differential adeno-associated virus serotype-specific interaction patterns with synthetic heparins and other glycans. J. Virol. 2014, 88, 2991–3003. [Google Scholar] [CrossRef] [PubMed]
- Akache, B.; Grimm, D.; Pandey, K.; Yant, S.R.; Xu, H.; Kay, M.A. The 37/67-kilodalton laminin receptor is a receptor for adeno-associated virus serotypes 8, 2, 3, and 9. J. Virol. 2006, 80, 9831–9836. [Google Scholar] [CrossRef]
- Qing, K.; Mah, C.; Hansen, J.; Zhou, S.; Dwarki, V.; Srivastava, A. Human fibroblast growth factor receptor 1 is a co-receptor for infection by adeno-associated virus 2. Nat. Med. 1999, 5, 71–77. [Google Scholar] [CrossRef]
- Kashiwakura, Y.; Tamayose, K.; Iwabuchi, K.; Hirai, Y.; Shimada, T.; Matsumoto, K.; Nakamura, T.; Watanabe, M.; Oshimi, K.; Daida, H. Hepatocyte growth factor receptor is a coreceptor for adeno-associated virus type 2 infection. J. Virol. 2005, 79, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Weller, M.L.; Amornphimoltham, P.; Schmidt, M.; Wilson, P.A.; Gutkind, J.S.; Chiorini, J.A. Epidermal growth factor receptor is a co-receptor for adeno-associated virus serotype 6. Nat. Med. 2010, 16, 662–664. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Miller, E.; Agbandje-McKenna, M.; Samulski, R.J. Alpha2,3 and alpha2,6 n-linked sialic acids facilitate efficient binding and transduction by adeno-associated virus types 1 and 6. J. Virol. 2006, 80, 9093–9103. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, G.; Kaludov, N.; Agbandje-McKenna, M.; Chiorini, J.A. Baav transcytosis requires an interaction with Beta-1-4 linked- glucosamine and gp96. PLoS ONE 2010, 5, e9336. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.; Taylor, K.A.; Chapman, M.S. Adeno-associated virus-2 and its primary cellular receptor--cryo-em structure of a heparin complex. Virology 2009, 385, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Levy, H.C.; Bowman, V.D.; Govindasamy, L.; McKenna, R.; Nash, K.; Warrington, K.; Chen, W.; Muzyczka, N.; Yan, X.; Baker, T.S.; et al. Heparin binding induces conformational changes in adeno-associated virus serotype 2. J. Struct. Biol. 2009, 165, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Spear, J.M.; Noble, A.J.; Sousa, D.R.; Meyer, N.L.; Davulcu, O.; Zhang, F.; Linhardt, R.J.; Stagg, S.M.; Chapman, M.S. The 2.8 A electron microscopy structure of adeno-associated virus-dj bound by a heparinoid pentasaccharide. Mol. Ther. Methods Clin. Dev. 2017, 5, 1–12. [Google Scholar] [CrossRef][Green Version]
- Afione, S.; DiMattia, M.A.; Halder, S.; Di Pasquale, G.; Agbandje-McKenna, M.; Chiorini, J.A. Identification and mutagenesis of the adeno-associated virus 5 sialic acid binding region. J. Virol. 2015, 89, 1660–1672. [Google Scholar] [CrossRef]
- Lerch, T.F.; Chapman, M.S. Identification of the heparin binding site on adeno-associated virus serotype 3B (AAV-3B). Virology 2012, 423, 6–13. [Google Scholar] [CrossRef]
- Huang, L.Y.; Patel, A.; Ng, R.; Miller, E.B.; Halder, S.; McKenna, R.; Asokan, A.; Agbandje-McKenna, M. Characterization of the adeno-associated virus 1 and 6 sialic acid binding site. J. Virol. 2016, 90, 5219–5230. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Bueno, A.; Rubio, M.P.; Bryant, N.; McKenna, R.; Agbandje-McKenna, M.; Almendral, J.M. Host-selected amino acid changes at the sialic acid binding pocket of the parvovirus capsid modulate cell binding affinity and determine virulence. J. Virol. 2006, 80, 1563–1573. [Google Scholar] [CrossRef]
- Zhang, R.; Cao, L.; Cui, M.; Sun, Z.; Hu, M.; Zhang, R.; Stuart, W.; Zhao, X.; Yang, Z.; Li, X.; et al. Adeno-associated virus 2 bound to its cellular receptor AAVR. Nat. Microbiol. 2019, 4, 675–682. [Google Scholar] [CrossRef]
- Rohrer, C.; Gartner, B.; Sauerbrei, A.; Bohm, S.; Hottentrager, B.; Raab, U.; Thierfelder, W.; Wutzler, P.; Modrow, S. Seroprevalence of parvovirus B19 in the german population. Epidemiol. Infect. 2008, 136, 1564–1575. [Google Scholar] [CrossRef]
- Boutin, S.; Monteilhet, V.; Veron, P.; Leborgne, C.; Benveniste, O.; Montus, M.F.; Masurier, C. Prevalence of serum igg and neutralizing factors against adeno-associated virus (AAV) types 1, 2, 5, 6, 8, and 9 in the healthy population: Implications for gene therapy using AAV vectors. Hum. Gene Ther. 2010, 21, 704–712. [Google Scholar] [CrossRef]
- Guo, L.; Wang, Y.; Zhou, H.; Wu, C.; Song, J.; Li, J.; Paranhos-Baccala, G.; Vernet, G.; Wang, J.; Hung, T. Differential seroprevalence of human bocavirus species 1-4 in Beijing, China. PLoS ONE 2012, 7, e39644. [Google Scholar] [CrossRef] [PubMed]
- Vaisanen, E.; Mohanraj, U.; Kinnunen, P.M.; Jokelainen, P.; Al-Hello, H.; Barakat, A.M.; Sadeghi, M.; Jalilian, F.A.; Majlesi, A.; Masika, M.; et al. Global distribution of human protoparvoviruses. Emerg. Infect. Dis. 2018, 24, 1292–1299. [Google Scholar] [CrossRef] [PubMed]
- Sharp, C.P.; Vermeulen, M.; Nebie, Y.; Djoko, C.F.; LeBreton, M.; Tamoufe, U.; Rimoin, A.W.; Kayembe, P.K.; Carr, J.K.; Servant-Delmas, A.; et al. Changing epidemiology of human parvovirus 4 infection in sub-Saharan Africa. Emerg. Infect. Dis. 2010, 16, 1605–1607. [Google Scholar] [CrossRef]
- Jose, A.; Mietzsch, M.; Smith, K.; Kurian, J.; Chipman, P.; McKenna, R.; Chiorini, J.; Agbandje-McKenna, M. High resolution structural characterization of a new aav5 antibody epitope toward engineering antibody resistant recombinant gene delivery vectors. J. Virol. 2018, 93, e01394-18. [Google Scholar] [CrossRef]
- von Kietzell, K.; Pozzuto, T.; Heilbronn, R.; Grossl, T.; Fechner, H.; Weger, S. Antibody-mediated enhancement of parvovirus B19 uptake into endothelial cells mediated by a receptor for complement factor C1Q. J. Virol. 2014, 88, 8102–8115. [Google Scholar] [CrossRef]
- Kanno, H.; Wolfinbarger, J.B.; Bloom, M.E. Aleutian mink disease parvovirus infection of mink macrophages and human macrophage cell line u937: Demonstration of antibody-dependent enhancement of infection. J. Virol. 1993, 67, 7017–7024. [Google Scholar] [PubMed]
- Organtini, L.J.; Lee, H.; Iketani, S.; Huang, K.; Ashley, R.E.; Makhov, A.M.; Conway, J.F.; Parrish, C.R.; Hafenstein, S. Near-atomic resolution structure of a highly neutralizing Fab bound to canine parvovirus. J. Virol. 2016, 90, 9733–9742. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, B.; Lopez-Bueno, A.; Mateu, M.G.; Chipman, P.R.; Nelson, C.D.; Parrish, C.R.; Almendral, J.M.; Rossmann, M.G. Minute virus of mice, a parvovirus, in complex with the fab fragment of a neutralizing monoclonal antibody. J. Virol. 2007, 81, 9851–9858. [Google Scholar] [CrossRef]
- Kailasan, S.; Garrison, J.; Ilyas, M.; Chipman, P.; McKenna, R.; Kantola, K.; Soderlund-Venermo, M.; Kucinskaite-Kodze, I.; Zvirbliene, A.; Agbandje-McKenna, M. Mapping antigenic epitopes on the human bocavirus capsid. J. Virol. 2016, 90, 4670–4680. [Google Scholar] [CrossRef]
- Tseng, Y.S.; Gurda, B.L.; Chipman, P.; McKenna, R.; Afione, S.; Chiorini, J.A.; Muzyczka, N.; Olson, N.H.; Baker, T.S.; Kleinschmidt, J.; et al. Adeno-associated virus serotype 1 (AAV1)- and AAV5-antibody complex structures reveal evolutionary commonalities in parvovirus antigenic reactivity. J. Virol. 2015, 89, 1794–1808. [Google Scholar] [CrossRef] [PubMed]
- Gurda, B.L.; DiMattia, M.A.; Miller, E.B.; Bennett, A.; McKenna, R.; Weichert, W.S.; Nelson, C.D.; Chen, W.J.; Muzyczka, N.; Olson, N.H.; et al. Capsid antibodies to different adeno-associated virus serotypes bind common regions. J. Virol. 2013, 87, 9111–9124. [Google Scholar] [CrossRef]
- McCraw, D.M.; O’Donnell, J.K.; Taylor, K.A.; Stagg, S.M.; Chapman, M.S. Structure of adeno-associated virus-2 in complex with neutralizing monoclonal antibody A20. Virology 2012, 431, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.D.; Wong, K.; Lewis, J.; Tseng, Y.S.; Smith, J.K.; Chipman, P.; McKenna, R.; Samulski, R.J.; Kleinschmidt, J.; Agbandje-McKenna, M. AAV6 K531 serves a dual function in selective receptor and antibody ADK6 recognition. Virology 2018, 518, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Gurda, B.L.; Raupp, C.; Popa-Wagner, R.; Naumer, M.; Olson, N.H.; Ng, R.; McKenna, R.; Baker, T.S.; Kleinschmidt, J.A.; Agbandje-McKenna, M. Mapping a neutralizing epitope onto the capsid of adeno-associated virus serotype 8. J. Virol. 2012, 86, 7739–7751. [Google Scholar] [CrossRef] [PubMed]
- Giles, A.R.; Govindasamy, L.; Somanathan, S.; Wilson, J.M. Mapping an adeno-associated virus 9-specific neutralizing epitope to develop next-generation gene delivery vectors. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Sun, Y.; Klose, T.; Liu, Y.; Modrow, S.; Rossmann, M.G. Structure of parvovirus B19 decorated by Fabs from a human antibody. J. Virol. 2019. [Google Scholar] [CrossRef]
- Daya, S.; Berns, K.I. Gene therapy using adeno-associated virus vectors. Clin. Microbiol. Rev. 2008, 21, 583–593. [Google Scholar] [CrossRef]
- Fakhiri, J.; Schneider, M.A.; Puschhof, J.; Stanifer, M.; Schildgen, V.; Holderbach, S.; Voss, Y.; El Andari, J.; Schildgen, O.; Boulant, S.; et al. Novel chimeric gene therapy vectors based on adeno-associated virus (AAV) and four different mammalian bocaviruses (BOV). Mol. Ther. Methods Clin. Dev. 2019, 12, 202–222. [Google Scholar] [CrossRef]
- Gray, J.T.; Zolotukhin, S. Design and construction of functional AAV vectors. Methods Mol. Biol. 2011, 807, 25–46. [Google Scholar] [PubMed]
- Zhong, L.; Li, B.; Jayandharan, G.; Mah, C.S.; Govindasamy, L.; Agbandje-McKenna, M.; Herzog, R.W.; Weigel-Van Aken, K.A.; Hobbs, J.A.; Zolotukhin, S.; et al. Tyrosine-phosphorylation of AAV2 vectors and its consequences on viral intracellular trafficking and transgene expression. Virology 2008, 381, 194–202. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Zhao, W.; Wu, J.; Li, B.; Zolotukhin, S.; Govindasamy, L.; Agbandje-McKenna, M.; Srivastava, A. A dual role of EGFR protein tyrosine kinase signaling in ubiquitination of AAV2 capsids and viral second-strand DNA synthesis. Mol. Ther. 2007, 15, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Zhong, L.; Li, B.; Mah, C.S.; Govindasamy, L.; Agbandje-McKenna, M.; Cooper, M.; Herzog, R.W.; Zolotukhin, I.; Warrington, K.H., Jr.; Weigel-Van Aken, K.A.; et al. Next generation of adeno-associated virus 2 vectors: Point mutations in tyrosines lead to high-efficiency transduction at lower doses. Proc. Natl. Acad. Sci. USA 2008, 105, 7827–7832. [Google Scholar] [CrossRef] [PubMed]
- Aslanidi, G.V.; Rivers, A.E.; Ortiz, L.; Song, L.; Ling, C.; Govindasamy, L.; Van Vliet, K.; Tan, M.; Agbandje-McKenna, M.; Srivastava, A. Optimization of the capsid of recombinant adeno-associated virus 2 (AAV2) vectors: The final threshold? PLoS ONE 2013, 8, e59142. [Google Scholar] [CrossRef]
- Gabriel, N.; Hareendran, S.; Sen, D.; Gadkari, R.A.; Sudha, G.; Selot, R.; Hussain, M.; Dhaksnamoorthy, R.; Samuel, R.; Srinivasan, N.; et al. Bioengineering of AAV2 capsid at specific serine, threonine, or lysine residues improves its transduction efficiency in vitro and in vivo. Hum. Gene Ther. Methods 2013, 24, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Hurlbut, G.D.; Ziegler, R.J.; Nietupski, J.B.; Foley, J.W.; Woodworth, L.A.; Meyers, E.; Bercury, S.D.; Pande, N.N.; Souza, D.W.; Bree, M.P.; et al. Preexisting immunity and low expression in primates highlight translational challenges for liver-directed AAV8-mediated gene therapy. Mol. Ther. 2010, 18, 1983–1994. [Google Scholar] [CrossRef]
- Velazquez, V.M.; Meadows, A.S.; Pineda, R.J.; Camboni, M.; McCarty, D.M.; Fu, H. Effective depletion of pre-existing anti-aav antibodies requires broad immune targeting. Mol. Ther. Methods Clin. Dev. 2017, 4, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Mingozzi, F.; High, K.A. Immune responses to AAV vectors: Overcoming barriers to successful gene therapy. Blood 2013, 122, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Arruda, V.R.; Favaro, P.; Finn, J.D. Strategies to modulate immune responses: A new frontier for gene therapy. Mol. Ther. 2009, 17, 1492–1503. [Google Scholar] [CrossRef]
- Wang, Z.; Storb, R.; Halbert, C.L.; Banks, G.B.; Butts, T.M.; Finn, E.E.; Allen, J.M.; Miller, A.D.; Chamberlain, J.S.; Tapscott, S.J. Successful regional delivery and long-term expression of a dystrophin gene in canine muscular dystrophy: A preclinical model for human therapies. Mol. Ther. 2012, 20, 1501–1507. [Google Scholar] [CrossRef]
- Mingozzi, F.; Anguela, X.M.; Pavani, G.; Chen, Y.; Davidson, R.J.; Hui, D.J.; Yazicioglu, M.; Elkouby, L.; Hinderer, C.J.; Faella, A.; et al. Overcoming preexisting humoral immunity to aav using capsid decoys. Sci. Transl. Med. 2013, 5, 194ra192. [Google Scholar] [CrossRef] [PubMed]
- Tse, L.V.; Klinc, K.A.; Madigan, V.J.; Castellanos Rivera, R.M.; Wells, L.F.; Havlik, L.P.; Smith, J.K.; Agbandje-McKenna, M.; Asokan, A. Structure-guided evolution of antigenically distinct adeno-associated virus variants for immune evasion. Proc. Natl. Acad. Sci. USA 2017, 114, E4812–E4821. [Google Scholar] [CrossRef]
- Tseng, Y.S.; Agbandje-McKenna, M. Mapping the aav capsid host antibody response toward the development of second generation gene delivery vectors. Front. Immunol. 2014, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Buning, H.; Ried, M.U.; Perabo, L.; Gerner, F.M.; Huttner, N.A.; Enssle, J.; Hallek, M. Receptor targeting of adeno-associated virus vectors. Gene Ther. 2003, 10, 1142–1151. [Google Scholar] [CrossRef][Green Version]
- Yu, C.Y.; Yuan, Z.; Cao, Z.; Wang, B.; Qiao, C.; Li, J.; Xiao, X. A muscle-targeting peptide displayed on AAV2 improves muscle tropism on systemic delivery. Gene Ther. 2009, 16, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Dalkara, D.; Byrne, L.C.; Klimczak, R.R.; Visel, M.; Yin, L.; Merigan, W.H.; Flannery, J.G.; Schaffer, D.V. In vivo-directed evolution of a new adeno-associated virus for therapeutic outer retinal gene delivery from the vitreous. Sci. Transl. Med. 2013, 5, 189ra176. [Google Scholar] [CrossRef]
- Judd, J.; Wei, F.; Nguyen, P.Q.; Tartaglia, L.J.; Agbandje-McKenna, M.; Silberg, J.J.; Suh, J. Random insertion of mcherry into vp3 domain of adeno-associated virus yields fluorescent capsids with no loss of infectivity. Mol. Ther. Nucleic Acids 2012, 1, e54. [Google Scholar] [CrossRef] [PubMed]
- Asuri, P.; Bartel, M.A.; Vazin, T.; Jang, J.H.; Wong, T.B.; Schaffer, D.V. Directed evolution of adeno-associated virus for enhanced gene delivery and gene targeting in human pluripotent stem cells. Mol. Ther. 2012, 20, 329–338. [Google Scholar] [CrossRef]
- Bowles, D.E.; McPhee, S.W.; Li, C.; Gray, S.J.; Samulski, J.J.; Camp, A.S.; Li, J.; Wang, B.; Monahan, P.E.; Rabinowitz, J.E.; et al. Phase 1 gene therapy for Duchenne muscular dystrophy using a translational optimized aav vector. Mol. Ther. 2012, 20, 443–455. [Google Scholar] [CrossRef]
- Bergoin, M.; Tijssen, P. Densoviruses: A highly diverse group of arthropod parvoviruses. In Insect Virology; Asgari, S., Johnson, K.N., Eds.; Horizon Scientific Press: Norfolk, UK, 2010; pp. 59–82. [Google Scholar]
- Palinski, R.M.; Mitra, N.; Hause, B.M. Discovery of a novel parvovirinae virus, porcine parvovirus 7, by metagenomic sequencing of porcine rectal swabs. Virus Genes 2016, 52, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Baquerizo-Audiot, E.; Abd-Alla, A.; Jousset, F.X.; Cousserans, F.; Tijssen, P.; Bergoin, M. Structure and expression strategy of the genome of Culex pipiens densovirus, a mosquito densovirus with an ambisense organization. J. Virol. 2009, 83, 6863–6873. [Google Scholar] [CrossRef]
- Tijssen, P.; Penzes, J.J.; Yu, Q.; Pham, H.T.; Bergoin, M. Diversity of small, single-stranded DNA viruses of invertebrates and their chaotic evolutionary past. J. Invertebr. Pathol. 2016, 140, 83–96. [Google Scholar] [CrossRef]
- Kim, E.; Magen, A.; Ast, G. Different levels of alternative splicing among eukaryotes. Nucleic Acids Res. 2007, 35, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Roediger, B.; Lee, Q.; Tikoo, S.; Cobbin, J.C.A.; Henderson, J.M.; Jormakka, M.; O’Rourke, M.B.; Padula, M.P.; Pinello, N.; Henry, M.; et al. An atypical parvovirus drives chronic tubulointerstitial nephropathy and kidney fibrosis. Cell 2018, 175, 530–543.e524. [Google Scholar] [CrossRef] [PubMed]
- Tijssen, P.; Li, Y.; El-Far, M.; Szelei, J.; Letarte, M.; Zadori, Z. Organization and expression strategy of the ambisense genome of densonucleosis virus of Galleria mellonella. J. Virol. 2003, 77, 10357–10365. [Google Scholar] [CrossRef]
- Yang, B.; Dong, X.; Cai, D.; Wang, X.; Liu, Z.; Hu, Z.; Wang, H.; Cao, X.; Zhang, J.; Hu, Y. Characterization of the promoter elements and transcription profile of Periplaneta fuliginosa densovirus nonstructural genes. Virus Res. 2008, 133, 149–156. [Google Scholar] [CrossRef]
- Liu, K.; Li, Y.; Jousset, F.X.; Zadori, Z.; Szelei, J.; Yu, Q.; Pham, H.T.; Lepine, F.; Bergoin, M.; Tijssen, P. The Acheta domesticus densovirus, isolated from the european house cricket, has evolved an expression strategy unique among parvoviruses. J. Virol. 2011, 85, 10069–10078. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Tijssen, P. Gene expression of five different iteradensoviruses: Bombyx mori densovirus, Casphalia extranea densovirus, Papilio polyxenes densovirus, Sibine fusca densovirus, and Danaus plexippus densovirus. J. Virol. 2014, 88, 12152–12157. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zadori, Z.; Bando, H.; Dubuc, R.; Fediere, G.; Szelei, J.; Tijssen, P. Genome organization of the densovirus from Bombyx mori (BmDNV-1) and enzyme activity of its capsid. J. Gen. Virol. 2001, 82, 2821–2825. [Google Scholar] [CrossRef]
- Pham, H.T.; Jousset, F.X.; Perreault, J.; Shike, H.; Szelei, J.; Bergoin, M.; Tijssen, P. Expression strategy of Aedes albopictus densovirus. J. Virol. 2013, 87, 9928–9932. [Google Scholar] [CrossRef] [PubMed]
- Pham, H.T. Molecular Biology of Single-Stranded DNA Viruses in Shrimps and Crickets. Ph.D. Thesis, Université du Québec, Quebec City, QC, Canada, 2015. [Google Scholar]
- Sukhumsirichart, W.; Attasart, P.; Boonsaeng, V.; Panyim, S. Complete nucleotide sequence and genomic organization of hepatopancreatic parvovirus (HPV) of penaeus monodon. Virology 2006, 346, 266–277. [Google Scholar] [CrossRef]
- Kaufmann, B.; Bowman, V.D.; Li, Y.; Szelei, J.; Waddell, P.J.; Tijssen, P.; Rossmann, M.G. Structure of Penaeus stylirostris densovirus, a shrimp pathogen. J. Virol. 2010, 84, 11289–11296. [Google Scholar] [CrossRef]
- Bruemmer, A.; Scholari, F.; Lopez-Ferber, M.; Conway, J.F.; Hewat, E.A. Structure of an insect parvovirus (Junonia coenia Densovirus) determined by cryo-electron microscopy. J. Mol. Biol. 2005, 347, 791–801. [Google Scholar] [CrossRef]
- Chen, S.; Cheng, L.; Zhang, Q.; Lin, W.; Lu, X.; Brannan, J.; Zhou, Z.H.; Zhang, J. Genetic, biochemical, and structural characterization of a new densovirus isolated from a chronically infected aedes albopictus C6/36 cell line. Virology 2004, 318, 123–133. [Google Scholar] [CrossRef]
- Holm, L.; Rosenstrom, P. Dali server: Conservation mapping in 3d. Nucleic Acids Res. 2010, 38, W545–W549. [Google Scholar] [CrossRef]
- Harrison, S.C.; Olson, A.J.; Schutt, C.E.; Winkler, F.K.; Bricogne, G. Tomato bushy stunt virus at 2.9 a resolution. Nature 1978, 276, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Rossmann, M.G.; Abad-Zapatero, C.; Hermodson, M.A.; Erickson, J.W. Subunit interactions in southern bean mosaic virus. J. Mol. Biol. 1983, 166, 37–73. [Google Scholar] [CrossRef]
- Multeau, C.; Froissart, R.; Perrin, A.; Castelli, I.; Casartelli, M.; Ogliastro, M. Four amino acids of an insect Densovirus capsid determine midgut tropism and virulence. J. Virol. 2012, 86, 5937–5941. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, Y.; Gosselin Grenet, A.S.; Castelli, I.; Cermenati, G.; Ravallec, M.; Fiandra, L.; Debaisieux, S.; Multeau, C.; Lautredou, N.; Dupressoir, T.; et al. Densovirus crosses the insect midgut by transcytosis and disturbs the epithelial barrier function. J. Virol. 2013, 87, 12380–12391. [Google Scholar] [CrossRef] [PubMed]
- Kozlov, E.N.; Martynova, E.U.; Popenko, V.I.; Schal, C.; Mukha, D.V. Intracellular localization of blattella germanica Densovirus (BgDV1) capsid proteins. Viruses 2018, 10, 370. [Google Scholar] [CrossRef]
- Owens, L. Bioinformatical analysis of nuclear localisation sequences in penaeid densoviruses. Mar. Genomics 2013, 12, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Yuan, H.; Yang, X.; Graham, R.I.; Liu, K.; Wu, K. Structural proteins of helicoverpa armigera Densovirus 2 enhance transcription of viral genes through transactivation. Arch. Virol. 2017, 162, 1745–1750. [Google Scholar] [CrossRef] [PubMed]
- Aydemir, F.; Salganik, M.; Resztak, J.; Singh, J.; Bennett, A.; Agbandje-McKenna, M.; Muzyczka, N. Mutants at the 2-fold interface of adeno-associated virus type 2 (AAV2) structural proteins suggest a role in viral transcription for aav capsids. J. Virol. 2016, 90, 7196–7204. [Google Scholar] [CrossRef] [PubMed]
- McKenna, R.; Olson, N.H.; Chipman, P.R.; Baker, T.S.; Booth, T.F.; Christensen, J.; Aasted, B.; Fox, J.M.; Bloom, M.E.; Wolfinbarger, J.B.; et al. Three-dimensional structure of aleutian mink disease parvovirus: Implications for disease pathogenicity. J. Virol. 1999, 73, 6882–6891. [Google Scholar]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Empty/Full | Structure Determination Method | Year | Resolution in Å | PDB-ID | Reference |
|---|---|---|---|---|---|---|
| Protoparvovirus | ||||||
| BuV1 | Empty | Cryo-EM | 2018 | 2.8 | 6BWX | Ilyas et al. [39] |
| BuV2 | Empty | Cryo-EM | 2018 | 3.8 | 6BX0 | Ilyas et al. [39] |
| BuV3 | Empty | Cryo-EM | 2018 | 3.3 | 6BX1 | Ilyas et al. [39] |
| CPV | Empty | X-Ray Crystallography | 1993 | 3.0 | 2CAS | Wu et al. [38] |
| CPV | Full | X-Ray Crystallography | 1996 | 2.9 | 4DPV | Xie et al. [40] |
| CPV-N93D | Full | X-Ray Crystallography | 2003 | 3.3 | 1P5Y | Govindasamy et al. [41] |
| CPV-N93R | Full | X-Ray Crystallography | 2003 | 3.3 | 1P5W | Govindasamy et al. [41] |
| CPV-d-A300D | Empty | X-Ray Crystallography | 2000 | 3.3 | 1C8D | Simpson et al. [42] |
| CPV-d-A300D | Full | X-Ray Crystallography | 1996 | 3.3 | 1IJS | Llamas-Saiz et al. [43] |
| CPV-d pH5.5 | Empty | X-Ray Crystallography | 2000 | 3.5 | 1C8H | Simpson et al. [42] |
| CPV2a | Full | X-Ray Crystallography | 2014 | 3.3 | 4QYK | Organtini et al. [44] |
| FPV | Empty | X-Ray Crystallography | 1993 | 3.3 | 1FPV | Agbandje et al. [45] |
| FPV | Empty | X-Ray Crystallography | 2000 | 3.0 | 1C8F | Simpson et al. [42] |
| FPV low pH | Empty | X-Ray Crystallography | 2000 | 3.0 | 1C8G | Simpson et al. [42] |
| FPV no CaCl2 | Empty | X-Ray Crystallography | 2000 | 3.0 | 1C8E | Simpson et al. [42] |
| H-1PV | Full | X-Ray Crystallography | 2013 | 2.7 | 4G0R | Halder et al. [46] |
| H-1PV | Empty | X-Ray Crystallography | 2013 | 3.2 | 4GBT | Halder et al. [46] |
| LuIII | Empty | Cryo-EM | 2017 | 3.2 | 6B9Q | Pittman et al. [47] |
| M. Spretus EVE | Empty | Cryo-EM | 2018 | 3.9 | 6NF9 | Callaway et al. [48] |
| MVMi | Full | X-Ray Crystallography | 1997 | 3.5 | 1MVM | Llamas-Saiz et al. [49] |
| MVMi | Empty | X-Ray Crystallography | 2005 | 3.5 | 1Z1C | Kontou et al. [50] |
| MVMi-L172W | Empty | X-Ray Crystallography | 2011 | 4.2 | 2XGK | Plevka et al. [51] |
| MVMp | Full | X-Ray Crystallography | 2005 | 3.3 | 1Z14 | Kontou et al. [50] |
| MVMp-N170A | Empty | X-Ray Crystallography | 2015 | 3.8 | 4ZPY | Guerra et al. [52] |
| PPV | Empty | X-Ray Crystallography | 2001 | 3.5 | 1K3V | Simpson et al. [53] |
| Bocaparvovirus | ||||||
| BPV | Empty | X-Ray Crystallography | 2015 | 3.2 | 4QC8 | Kailasan et al. [5] |
| HBoV1 | Empty | Cryo-EM | 2017 | 2.9 | 5URF | Mietzsch et al. [54] |
| HBoV3 | Empty | Cryo-EM | 2017 | 2.8 | 5US7 | Mietzsch et al. [54] |
| HBoV4 | Empty | Cryo-EM | 2017 | 3.0 | 5US9 | Mietzsch et al. [54] |
| Dependoparvovirus | ||||||
| AAV1 | Full | X-Ray Crystallography | 2011 | 2.5 | 3NG9 | Ng et al. [55] |
| AAV2 | Full | X-Ray Crystallography | 2002 | 3.0 | 1LP3 | Xie et al. [3] |
| AAV2 | Empty | Cryo-EM | 2016 | 3.8 | 5IPI | Drouin et al. [56] |
| AAV2-L336C | Empty | Cryo-EM | 2018 | 1.9 | 6E9D | Tan et al. [57] |
| AAV2-R432A | Empty | Cryo-EM | 2016 | 3.7 | 5IPK | Drouin et al. [56] |
| AAV2.5 | Full | Cryo-EM | 2018 | 2.8 | 6CBE | Burg et al. [58] |
| AAV3 | Full | X-Ray Crystallography | 2010 | 2.6 | 3KIC | Lerch et al. [59] |
| AAV4 | Full | X-Ray Crystallography | 2007 | 3.2 | 2G8G | Govindasamy et al. [60] |
| AAV5 | Empty | X-Ray Crystallography | 2010 | 3.5 | 3NTT | Govindasamy et al. [61] |
| AAV6 | Empty | X-Ray Crystallography | 2010 | 3.0 | 3OAH | Ng et al. [55] |
| AAV6 | Full | X-Ray Crystallography | 2011 | 3.0 | 4V86 | Xie et al. [62] |
| AAV8 | Empty | X-Ray Crystallography | 2007 | 2.6 | 2QA0 | Nam et al. [63] |
| AAV8 pH7.5 | Full | X-Ray Crystallography | 2011 | 2.7 | 3RA4 | Nam et al. [64] |
| AAV8 pH6.0 | Full | X-Ray Crystallography | 2011 | 2.7 | 3RA9 | Nam et al. [64] |
| AAV8 pH5.5 | Full | X-Ray Crystallography | 2011 | 2.7 | 3RA8 | Nam et al. [64] |
| AAV8 pH4.0 | Full | X-Ray Crystallography | 2011 | 2.7 | 3RA2 | Nam et al. [64] |
| AAV8 pH4/7.5 | Full | X-Ray Crystallography | 2011 | 3.2 | 3RAA | Nam et al. [64] |
| AAV9 | Empty | X-Ray Crystallography | 2011 | 2.8 | 3UX1 | Dimattia et al. [65] |
| AAV9-L001 | Full | Cryo-EM | 2019 | 3.2 | 6NXE | Guenther et al. [66] |
| AAV-DJ | Empty | Cryo-EM | 2012 | 4.5 | 3J1Q | Lerch et al. [67] |
| AAVrh.8 | Full | X-Ray Crystallography | 2014 | 3.5 | 4RSO | Halder et al. [68] |
| AAVrh.32.33 | Full | X-Ray Crystallography | 2013 | 3.5 | 4IOV | Mikals et al. [69] |
| Erythroparvovirus | ||||||
| B19 | Empty | X-Ray Crystallography | 2004 | 3.5 | 1S58 | Kaufmann et al. [4] |

| Virus | Receptor | Structure Determination Method | Year | Resolution in Å | Reference |
|---|---|---|---|---|---|
| AAV2 | AAVR | Cryo-EM | 2019 | 2.8 | Zhang et al. [102] |
| AAV-DJ | heparinoid pentasaccharide | Cryo-EM | 2017 | 2.8 | Xie et al. [97] |
| AAV1 | SIA | X-Ray Crystallography | 2016 | 3.0 | Huang et al. [100] |
| AAV5 | SIA | X-Ray Crystallography | 2015 | 3.5 | Afione et al. [98] |
| AAV3 | sucrose octasulfate | X-Ray Crystallography | 2012 | 6.5 | Lerch et al. [99] |
| AAV2 | heparin | Cryo-EM | 2009 | 8.3 | O’Donnell et al. [95] |
| AAV2 | heparin | Cryo-EM | 2009 | 18.0 | Levy et al. [96] |
| CPV | transferrin receptor | Cryo-EM | 2007 | 25.0 | Hafenstein et al. [83] |
| MVMp | SIA | X-Ray Crystallography | 2006 | 3.5 | López-Bueno et al. [101] |
| Virus | Antibody Name | Year | Binding Region | Neutralizing for Infection | Resolution in Å | Reference |
|---|---|---|---|---|---|---|
| Protoparvovirus | ||||||
| CPV | Fab-E | 2012 | side of 3-fold protrusions across 2-fold axis | Yes | 4.1 | Organtini et al. [111] |
| CPV | Fab-14 | 2009 | 3-fold protrusions | Yes | 12.4 | Hafenstein et al. [84] |
| FPV | Fab-6 | 2009 | 3-fold protrusions | Yes | 18.0 | Hafenstein et al. [84] |
| FPV | Fab-8 | 2009 | 2/5-fold wall | Yes | 8.5 | Hafenstein et al. [84] |
| FPV | Fab-15 | 2009 | 2/5-fold wall | Yes | 10.5 | Hafenstein et al. [84] |
| FPV | Fab-16 | 2009 | 2/5-fold wall | Yes | 13.0 | Hafenstein et al. [84] |
| FPV | Fab-B | 2009 | 3-fold protrusions | Yes | 14.0 | Hafenstein et al. [84] |
| FPV | Fab-E | 2009 | side of 3-fold protrusions across 2-fold axis | Yes | 12.0 | Hafenstein et al. [84] |
| FPV | Fab-F | 2009 | side of 3-fold protrusions across 2-fold axis | Yes | 14.0 | Hafenstein et al. [84] |
| MVMi | B7 | 2007 | center of 3-fold symmetry axis | Yes | 7.0 | Kaufmann et al. [112] |
| Bocaparvovirus | ||||||
| HBoV1 | 4C2 | 2016 | 3-fold protrusions | unknown | 16.0 | Kailasan et al. [113] |
| HBoV1 | 9G12 | 2016 | 3-fold protrusions | unknown | 8.5 | Kailasan et al. [113] |
| HBoV1 | 12C1 | 2016 | 3-fold protrusions | unknown | 11.9 | Kailasan et al. [113] |
| HBoV1 | 15C6 | 2016 | around 5-fold symmetry axis | unknown | 18.6 | Kailasan et al. [113] |
| HBoV2 | 15C6 | 2016 | around 5-fold symmetry axis | unknown | 17.8 | Kailasan et al. [113] |
| HBoV4 | 15C6 | 2016 | around 5-fold symmetry axis | unknown | 9.5 | Kailasan et al. [113] |
| Dependoparvovirus | ||||||
| AAV1 | ADK1a | 2015 | 3-fold protrusions | Yes | 11.0 | Tseng et al. [114] |
| AAV1 | ADK1b | 2015 | 2/5-fold wall | Yes | 11.0 | Tseng et al. [114] |
| AAV1 | 4E4 | 2013 | side of 3-fold protrusions across 2-fold axis | Yes | 12.0 | Gurda et al. [115] |
| AAV1 | 5H7 | 2013 | center of 3-fold symmetry axis | Yes | 23.0 | Gurda et al. [115] |
| AAV2 | C37-B | 2013 | 3-fold protrusions | Yes | 11.0 | Gurda et al. [115] |
| AAV2 | A20 | 2012 | 2/5-fold wall | Yes | 8.5 | McCraw et al. [116] |
| AAV5 | ADK5a | 2015 | 2/5-fold wall | Yes | 11.0 | Tseng et al. [114] |
| AAV5 | ADK5b | 2015 | 2/5-fold wall to 5-fold symmetry axis | Yes | 12.0 | Tseng et al. [114] |
| AAV5 | HL2476 | 2018 | 3-fold protrusions | Yes | 3.1 | Jose et al. [108] |
| AAV5 | 3C5 | 2013 | 2/5-fold wall in a tangential orientation | No | 16.0 | Gurda et al. [115] |
| AAV6 | 5H7 | 2013 | center of 3-fold symmetry axis | unknown | 15.0 | Gurda et al. [115] |
| AAV6 | ADK6 | 2018 | 3-fold protrusions & 2/5-fold wall | Yes | 13.0 | Bennett et al. [117] |
| AAV8 | ADK8 | 2011 | 3-fold protrusions | Yes | 18.7 | Gurda et al. [118] |
| AAV9 | PAV9.1 | 2018 | center of 3-fold symmetry axis | Yes | 4.2 | Giles et al. [119] |
| Erythroparvovirus | ||||||
| B19 | human Fab | 2018 | around 5-fold symmetry axis | Yes | 3.2 | Sun et al. [120] |
| Virus | Empty / Full | Structure Determination Method | Year | Resolution in Å | PDB-ID | Reference |
|---|---|---|---|---|---|---|
| Ambidensovirus | ||||||
| AdDV | Full | X-Ray Crystallography | 2013 | 3.5 | 4MGU | Meng et al. [8] |
| GmDV | Full | X-Ray Crystallography | 1998 | 3.6 | 1DNV | Simpson et al. [7] |
| JcDV | Empty | Cryo-EM | 2005 | 8.7 | N/A | Bruemmer et al. [158] |
| Brevidensovirus | ||||||
| AalDV2 | Full | Cryo-EM | 2004 | 15.6 | N/A | Chen et al. [159] |
| Iteradensovirus | ||||||
| BmDV1 | Empty | X-Ray Crystallography | 2011 | 3.1 | 3P0S | Kaufmann et al. [9] |
| Penstyldensovirus | ||||||
| PstDV | Empty | X-Ray Crystallography | 2010 | 2.5 | 3N7X | Kaufmann et al. [157] |

| Inner Radius (Å) | Inner Surface Area (nm2) | Inner Volume (nm3) | Genome Size (nt) | Genus | |
|---|---|---|---|---|---|
| GmDV | 98.7 | 1223.2 | 4022.6 | 6039 | Ambidensovirus |
| AdDV | 91.7 | 1056.2 | 3227.8 | 5425 | Ambidensovirus |
| BmDV1 | 98.7 | 1224 | 4027.5 | 5076 | Iteradensovirus |
| PstDV | 87.6 | 963.3 | 2811.5 | 3914 | Penstyldensovirus |
| CPV | 92.9 | 1084.8 | 3359.5 | 5323 | Protoparvovirus |
| AAV2 | 89.9 | 1014.9 | 3040.4 | 4679 | Dependoparvovirus |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mietzsch, M.; Pénzes, J.J.; Agbandje-McKenna, M. Twenty-Five Years of Structural Parvovirology. Viruses 2019, 11, 362. https://doi.org/10.3390/v11040362
Mietzsch M, Pénzes JJ, Agbandje-McKenna M. Twenty-Five Years of Structural Parvovirology. Viruses. 2019; 11(4):362. https://doi.org/10.3390/v11040362
Chicago/Turabian StyleMietzsch, Mario, Judit J. Pénzes, and Mavis Agbandje-McKenna. 2019. "Twenty-Five Years of Structural Parvovirology" Viruses 11, no. 4: 362. https://doi.org/10.3390/v11040362
APA StyleMietzsch, M., Pénzes, J. J., & Agbandje-McKenna, M. (2019). Twenty-Five Years of Structural Parvovirology. Viruses, 11(4), 362. https://doi.org/10.3390/v11040362