Dynamics of Bacterial and Viral Communities in Paddy Soil with Irrigation and Urea Application


Abstract
1. Introduction
2. Materials and Methods
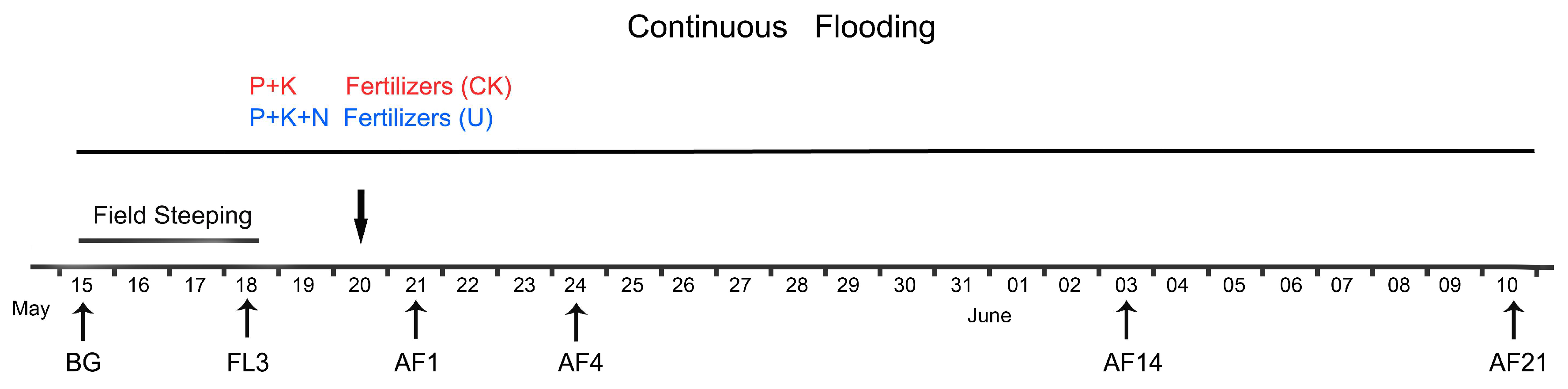
2.1. Field Description and Experimental Design
2.2. Sample Collection and Soil Analysis
2.3. Samples Processing
2.4. Epifluorescence Microscopy
2.5. Total DNA Extraction, Sequencing of 16s rDNA, and Bioinformatics Analysis
2.6. Randomly Amplified Polymorphic DNA (RAPD)-Polymerase Chain Reaction
2.7. Gaseous Emissions Measurements
2.8. Statistical Analysis
3. Results
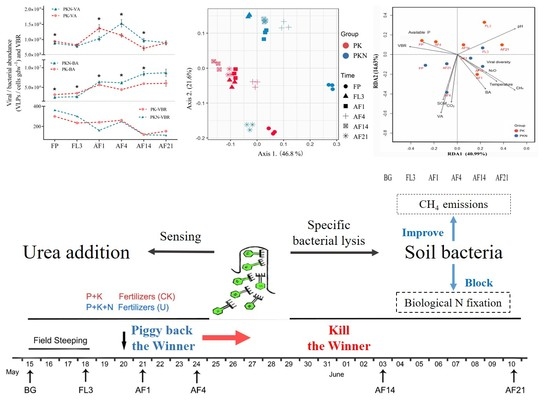
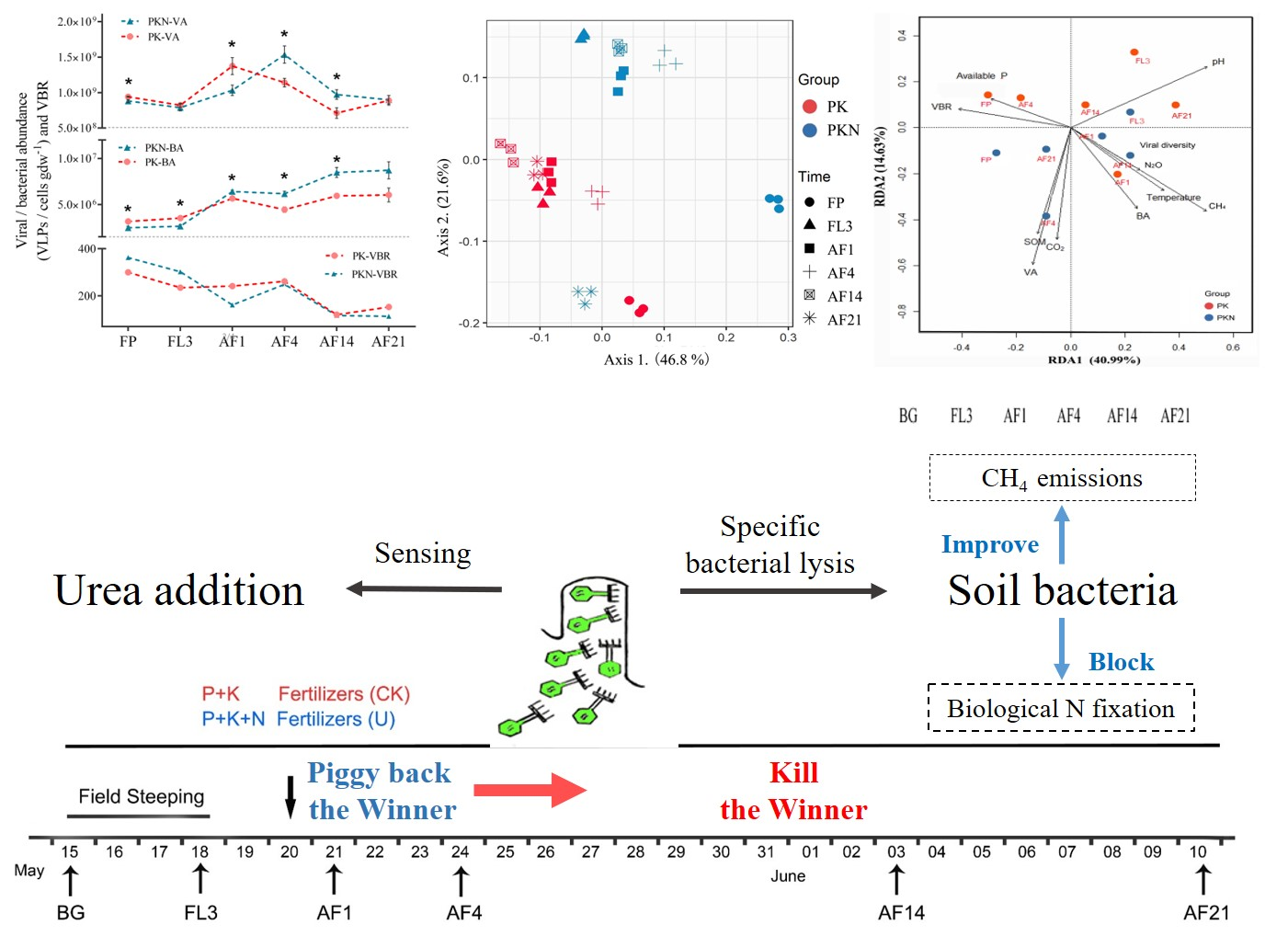
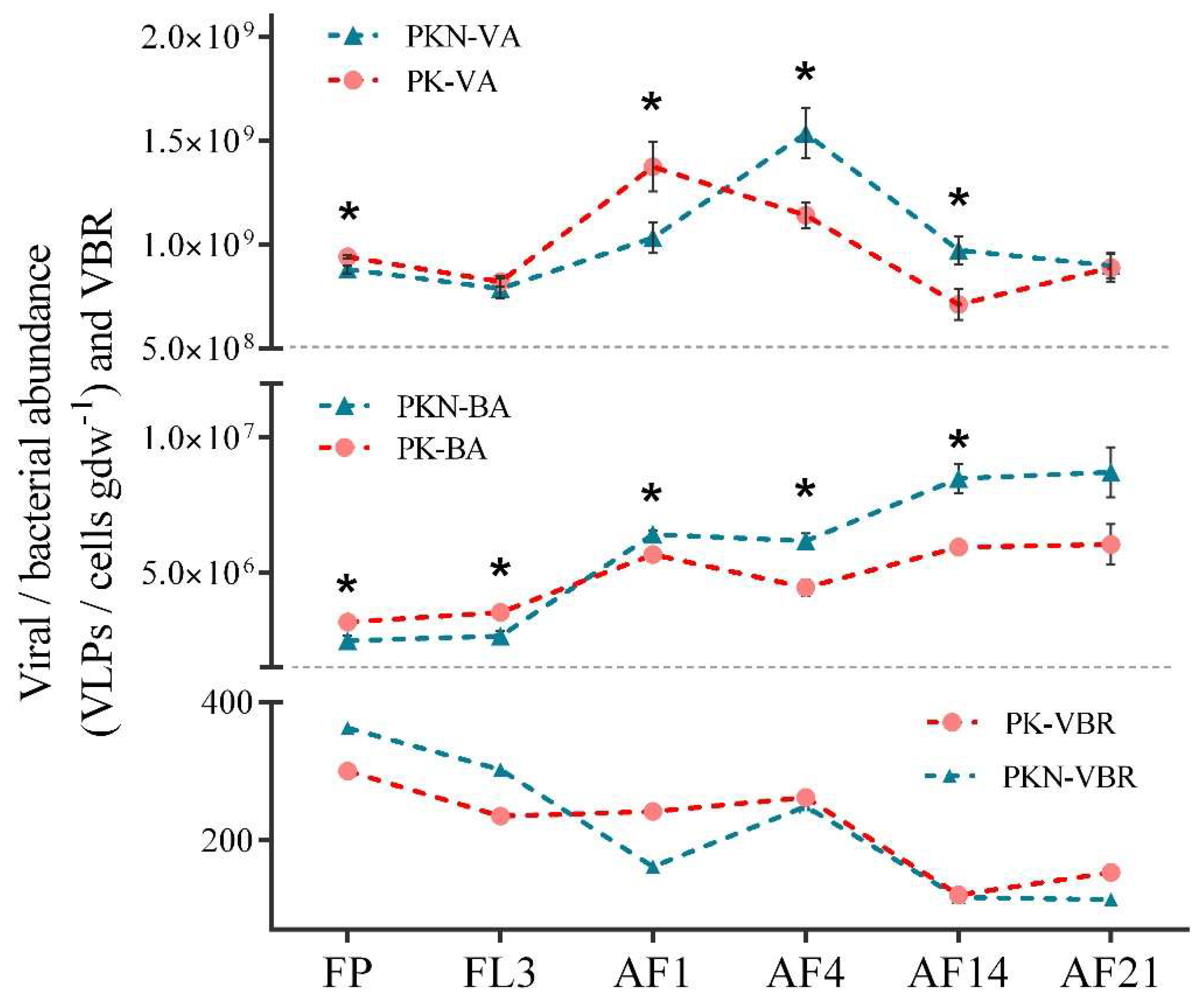
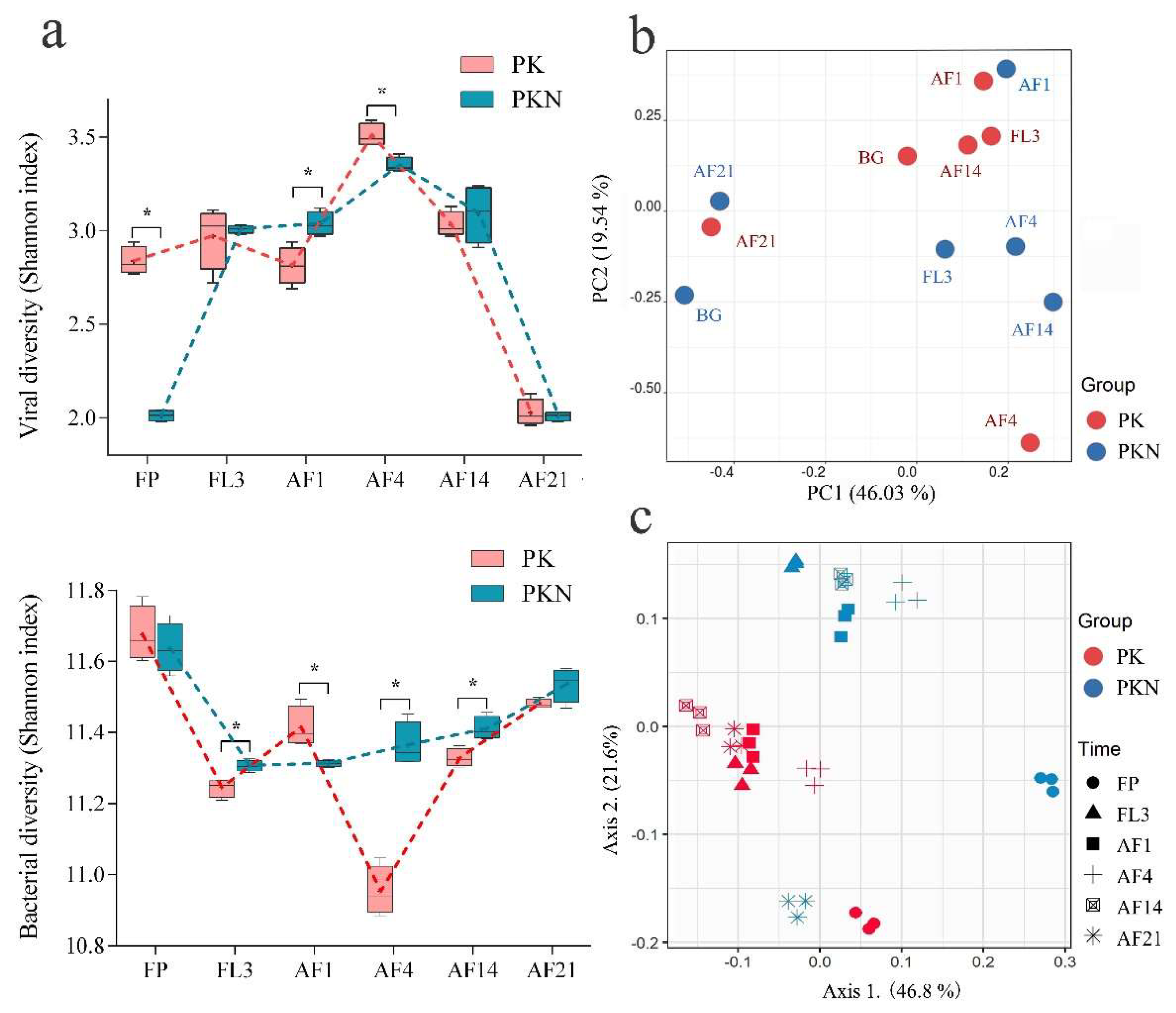
3.1. Viral and Bacterial Abundance
3.2. Viral and Bacterial Community Analyses
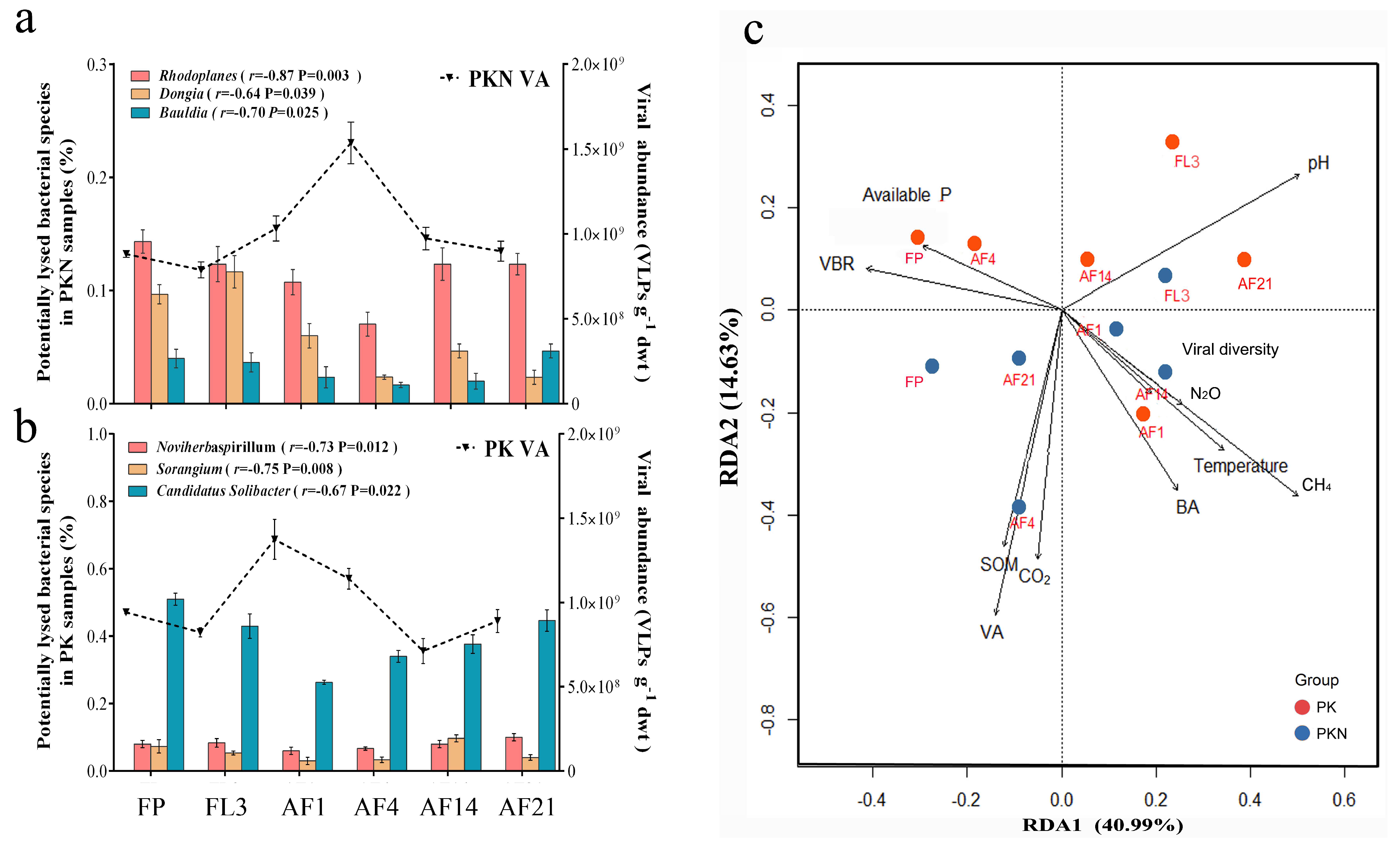
3.3. Correlations between Viral Lytic Environment and Bacterial Community
4. Discussion
4.1. Dynamic of VA, BA, and VBR
4.2. Viral and Bacterial Community
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rohwer, F.; Prangishvili, D.; Lindell, D. Roles of viruses in the environment. Environ. Microbiol. 2009, 11, 2771–2774. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Mason-Jones, K. Viruses in soil: Nano-scale undead drivers of microbial life, biogeochemical turnover and ecosystem functions. Soil Biol. Biochem. 2018, 127, 305–317. [Google Scholar] [CrossRef]
- Lindell, D.; Jaffe, J.D.; Coleman, M.L.; Futschik, M.E.; Axmann, I.M.; Rector, T.; Kettler, G.; Sullivan, M.B.; Steen, R.; Hess, W.R.; et al. Genome-wide expression dynamics of a marine virus and host reveal features of co-evolution. Nature 2007, 449, 83. [Google Scholar] [CrossRef] [PubMed]
- Tuomi, P.; Fagerbakke, K.M.; Bratbak, G.; Heldal, M. Nutritional enrichment of a microbial community: The effects on activity, elemental composition, community structure and virus production.pdf. FEMS Microbiol. Ecol. 1995, 16, 123–134. [Google Scholar] [CrossRef]
- Wommack, K.E.; Colwell, R.R. Virioplankton: Viruses in Aquatic ecosystems. Microbiol. Mol. Biol. Rev. 2000, 64, 69–114. [Google Scholar] [CrossRef] [PubMed]
- Jover, L.F.; Effler, T.C.; Buchan, A.; Wilhelm, S.W.; Weitz, J.S. The elemental composition of virus particles: implications for marine biogeochemical cycles. Nat. Rev. Microbiol. 2014, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- Maurice, C.F.; Bouvier, C.; de Wit, R.; Bouvier, T. Linking the lytic and lysogenic bacteriophage cycles to environmental conditions, host physiology and their variability in coastal lagoons. Environ. Microbiol. 2013, 15, 2463–2475. [Google Scholar] [CrossRef]
- Knowles, B.; Silveira, C.B.; Bailey, B.A.; Barott, K.; Cantu, V.A.; Cobian-Guemes, A.G.; Coutinho, F.H.; Dinsdale, E.A.; Felts, B.; Furby, K.A.; et al. Lytic to temperate switching of viral communities. Nature 2016, 531, 466–470. [Google Scholar] [CrossRef]
- Liang, X.; Zhuang, J.; Löffler, F.E.; Zhang, Y.; DeBruyn, J.M.; Wilhelm, S.W.; Schaeffer, S.M.; Radosevich, M. Viral and bacterial community responses to stimulated Fe(III)-bioreduction during simulated subsurface bioremediation. Environ. Microbiol. 2019. [Google Scholar] [CrossRef]
- Pratama, A.A.; van Elsas, J.D. The ‘neglected’ soil virome—potential role and impact. Trends Microbiol. 2018, 26, 649–662. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.E.; Fuhrmann, J.J.; Wommack, K.E.; Radosevich, M. Viruses in Soil Ecosystems: An Unknown Quantity Within an Unexplored Territory. Annu. Rev. Virol. 2017, 4, 201–219. [Google Scholar] [CrossRef]
- Zablocki, O.; Adriaenssens, E.M.; Cowan, D.; Löffler, F.E. Diversity and ecology of Viruses in hyperarid desert soils. Appl. Environ. Microbiol. 2016, 82, 770–777. [Google Scholar] [CrossRef]
- Segobola, J.; Adriaenssens, E.; Tsekoa, T.; Rashamuse, K.; Cowan, D. Exploring viral diversity in a unique South African soil habitat. Sci. Rep. 2018, 8, 111. [Google Scholar] [CrossRef]
- Emerson, J.B.; Roux, S.; Brum, J.R.; Bolduc, B.; Woodcroft, B.J.; Jang, H.B.; Singleton, C.M.; Solden, L.M.; Naas, A.E.; Boyd, J.A.; et al. Host-linked soil viral ecology along a permafrost thaw gradient. Nat. Microbiol. 2018, 3, 870–880. [Google Scholar] [CrossRef]
- Trubl, G.; Jang, H.B.; Roux, S.; Emerson, J.B.; Solonenko, N.; Vik, D.R.; Solden, L.; Ellenbogen, J.; Runyon, A.T.; Bolduc, B.; et al. Soil Viruses are underexplored players in ecosystem carbon processing. mSystems 2018, 3, e00018–e00076. [Google Scholar] [CrossRef]
- Adriaenssens, E.M.; Kramer, R.; Van Goethem, M.W.; Makhalanyane, T.P.; Hogg, I.; Cowan, D.A. Environmental drivers of viral community composition in Antarctic soils identified by viromics. Microbiome 2017, 5, 83. [Google Scholar] [CrossRef]
- Zablocki, O.; van Zyl, L.; Adriaenssens, E.M.; Rubagotti, E.; Tuffin, M.; Cary, S.C.; Cowan, D. High-Level diversity of tailed phages, eukaryote-associated viruses, and virophage-like elements in the metaviromes of Antarctic soils. Appl. Environ. Microbiol. 2014, 80, 6888–6897. [Google Scholar] [CrossRef]
- Wang, X.; Fan, J.; Xing, Y.; Xu, G.; Wang, H.; Deng, J.; Wang, Y.; Zhang, F.; Li, P.; Li, Z. The Effects of mulch and Nitrogen fertilizer on the soil environment of crop plants. Adv. Agron. 2019, 153, 121–173. [Google Scholar] [CrossRef]
- Chen, L.; Xun, W.; Sun, L.; Zhang, N.; Shen, Q.; Zhang, R. Effect of different long-term fertilization regimes on the viral community in an agricultural soil of Southern China. Eur. J. Soil Biol. 2014, 62, 121–126. [Google Scholar] [CrossRef]
- Williamson, K.E.; Radosevich, M.; Wommack, K.E. Abundance and diversity of viruses in six delaware soils. Appl. Environ. Microbiol. 2005, 71, 3119–3125. [Google Scholar] [CrossRef]
- Swanson, M.M.; Fraser, G.; Daniell, T.J.; Torrance, L.; Gregory, P.J.; Taliansky, M. Viruses in soils: morphological diversity and abundance in the rhizosphere. An. Appl. Biol. 2009, 155, 51–60. [Google Scholar] [CrossRef]
- Zhang, Y.; Liao, J.; Li, F.; Huang, Y.; Hu, R.; Yuan, Z. Fixed ammonium content of chief paddy soil types in Hunan Province and its influencing factors. Chin. J. Appl. Ecol. 2002, 13, 693–697. [Google Scholar]
- Wang, A.; Su, Y.; Li, Y.; Hu, L.; Wu, J. Response of the turnover of soil organic carbon to the soil moisture in paddy and upland Soil. Sci. Agric. Sin. 2012, 45, 266–274. [Google Scholar]
- Kimura, M.; Jia, Z.-J.; Nakayama, N.; Asakawa, S. Ecology of viruses in soils: Past, present and future perspectives. Soil Sci. Plant Nutr. 2008, 54, 1–32. [Google Scholar] [CrossRef]
- Srinivasiah, S.; Lovett, J.; Ghosh, D.; Roy, K.; Fuhrmann, J.J.; Radosevich, M.; Wommack, K.E. Dynamics of autochthonous soil viral communities parallels dynamics of host communities under nutrient stimulation. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef]
- Dong, D.; Kou, Y.; Yang, W.; Chen, G.; Xu, H. Effects of urease and nitrification inhibitors on nitrous oxide emissions and nitrifying/denitrifying microbial communities in a rainfed maize soil: A 6-year field observation. Soil Tillage Res. 2018, 180, 82–90. [Google Scholar] [CrossRef]
- Lu, R. Analysis of Soil Agrochemistry; Chinese Agricultural Science and Technology Press: Beijing, China, 1999. (In Chinese) [Google Scholar]
- Williamson, K.E.; Wommack, K.E.; Radosevich, M. Sampling Natural viral communities from soil for culture-independent analyses. Appl. Environ. Microbiol. 2003, 69, 6628–6633. [Google Scholar] [CrossRef]
- Williamson, K.E.; Radosevich, M.; Smith, D.W.; Wommack, K.E. Incidence of lysogeny within temperate and extreme soil environments. Environ. Microbiol. 2007, 9, 2563–2574. [Google Scholar] [CrossRef]
- Patel, A.; Noble, R.T.; Steele, J.A.; Schwalbach, M.S.; Hewson, I.; Fuhrman, J.A. Virus and prokaryote enumeration from planktonic aquatic environments by epifluorescence microscopy with SYBR Green I. Nat. Protoc. 2007, 2, 269–276. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef]
- Edwards, J.; Johnson, C.; Santos-Medellin, C.; Lurie, E.; Podishetty, N.K.; Bhatnagar, S.; Eisen, J.A.; Sundaresan, V. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Nat. Acad. Sci. USA 2015, 112, E911–E920. [Google Scholar] [CrossRef]
- Srinivasiah, S.; Lovett, J.; Polson, S.; Bhavsar, J.; Ghosh, D.; Roy, K.; Fuhrmann, J.J.; Radosevich, M.; Wommack, K.E. Direct assessment of viral diversity in soils by random PCR amplification of polymorphic DNA.pdf. Appl. Environ. Microbiol. 2013, 79, 5450–5457. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R package for reproducible interactive analysis and Graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Cobian Guemes, A.G.; Youle, M.; Cantu, V.A.; Felts, B.; Nulton, J.; Rohwer, F. Viruses as winners in the game of life. Annu. Rev. Virol. 2016, 3, 197–214. [Google Scholar] [CrossRef]
- Suttle, C.A. Marine viruses—major players in the global ecosystem. Nat. Rev. Microbiol. 2007, 5, 801–812. [Google Scholar] [CrossRef]
- Burroughs, N.J.; Marsh, P.; Wellington, E.M.H. Mathematical analysis of growth and interaction dynamics of streptomycetes and a bacteriophage in soil. Appl. Environ. Microbiol. 2000, 66, 3868–3877. [Google Scholar] [CrossRef]
- Beckett Stephen, J.; Williams Hywel, T.P. Coevolutionary diversification creates nested-modular structure in phage–bacteria interaction networks. Interface Focus 2013, 3, 20130033. [Google Scholar] [CrossRef]
- Weitz, J.S. Quantitative Viral Ecology: Dynamics of Viruses and Their Microbial Hosts; Princeton University Press: Oxfordshire, UK, 2015; pp. 98–99. [Google Scholar]
- Korytowski, D.A.; Smith, H.L. Permanence and Stability of a kill the winner model in marine ecology. Bull. Math. Biol. 2017, 79, 995–1004. [Google Scholar] [CrossRef]
- Kadavy, D.R.; Shaffer, J.J.; Lott, S.E.; Wolf, T.A.; Bolton, C.E.; Gallimore, W.H.; Martin, E.L.; Nickerson, K.W.; Kokjohn, T.A. Influence of infected cell growth state on bacteriophage reactivation levels. Appl. Environ. Microbiol. 2000, 66, 5206–5212. [Google Scholar] [CrossRef]
- Liang, X.; Radosevich, M. Commentary: A host-produced quorum-sensing autoinducer controls a phage lysis-lysogeny decision. Front. Microbiol. 2019. (to be published). [Google Scholar] [CrossRef]
- Hamilton, J.T.G.; McRoberts, W.C.; Keppler, F.; Kalin, R.M.; Harper, D.B. Chloride methylation by plant pectin: An efficient environmentally significant process. Science 2003, 301, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, K.S.; Lauber, C.L.; Knight, R.; Bradford, M.A.; Fierer, N. Consistent effects of nitrogen fertilization on soil bacterial communities in contrasting systems. Ecology 2010, 91, 3463–3470. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Liu, X.; Song, L.; Lin, X.; Zhang, H.; Shen, C.; Chu, H. Nitrogen fertilization directly affects soil bacterial diversity and indirectly affects bacterial community composition. Soil Biol. Biochem. 2016, 92, 41–49. [Google Scholar] [CrossRef]
- Emerson, J.B.; Adams, R.I.; Román, C.M.B.; Brooks, B.; Coil, D.A.; Dahlhausen, K.; Ganz, H.H.; Hartmann, E.M.; Hsu, T.; Justice, N.B.; et al. Schrödinger’s microbes: Tools for distinguishing the living from the dead in microbial ecosystems. Microbiome 2017, 5, 86. [Google Scholar] [CrossRef]
- Carini, P.; Marsden, P.J.; Leff, J.W.; Morgan, E.E.; Strickland, M.S.; Fierer, N. Relic DNA is abundant in soil and obscures estimates of soil microbial diversity. Nat. Microbiol. 2016, 2, 16242. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gao, Y.; Zhao, F. Phage–bacteria interaction network in human oral microbiome. Environ. Microbiol. 2016, 18, 2143–2158. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Nishiyama, M.; Otsuka, S.; Matsumoto, S. Effects of inoculation of phototrophic purple bacteria on grain yield of rice and nitrogenase activity of paddy soil in a pot experiment. Soil Sci. Plant Nutr. 2005, 51, 361–367. [Google Scholar] [CrossRef]
- Sun, M.; Xiao, T.; Ning, Z.; Xiao, E.; Sun, W. Microbial community analysis in rice paddy soils irrigated by acid mine drainage contaminated water. Appl. Microbiol. Biotechnol. 2015, 99, 2911–2922. [Google Scholar] [CrossRef]
- Kumar, P.A.; Srinivas, T.N.R.; Manasa, P.; Madhu, S.; Shivaji, S. Lutibaculum baratangense gen. nov., sp. nov., a proteobacterium isolated from a mud volcano. Int. J. Syst. Evol. Microbiol. 2012, 62, 2025–2031. [Google Scholar] [CrossRef]
- Black, M.; Moolhuijzen, P.; Chapman, B.; Barrero, R.; Howieson, J.; Hungria, M.; Bellgard, M. The Genetics of symbiotic nitrogen fixation: Comparative genomics of 14 rhizobia strains by resolution of Protein clusters. Genes 2012, 3, 138–166. [Google Scholar] [CrossRef]
- Qian, W.; Ma, B.; Li, X.; Zhang, Q.; Peng, Y. Long-term effect of pH on denitrification: High pH benefits achieving partial-denitrification. Biores. Technol. 2019, 278, 444–449. [Google Scholar] [CrossRef]
- Oshiki, M.; Segawa, T.; Ishii, S. Nitrogen cycle evaluation (nice) chip for simultaneous analysis of multiple n cycle-associated genes. Appl. Environ. Microbiol. 2018, 84, e02615–e02617. [Google Scholar] [CrossRef]
- Lindau, C.W.; DeLaune, R.D.; Patrick, W.H.; Bollich, P.K. Fertilizer effects on dinitrogen, nitrous oxide, and methane emissions from lowland rice. Soil Sci. Soc. Am. J. 1990, 54, 1789–1794. [Google Scholar] [CrossRef]
- Galagan, J.E.; Nusbaum, C.; Roy, A.; Endrizzi, M.G.; Macdonald, P.; FitzHugh, W.; Calvo, S.; Engels, R.; Smirnov, S.; Atnoor, D.; et al. The genome of M. acetivorans reveals extensive metabolic and physiological diversity. Genome Res. 2002, 12, 532–542. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil properties | PK | PKN |
|---|---|---|
| SOM2 (g/kg dw) | 14.62 | 13.86 |
| Total N (g/kg dw) | 0.84 | 0.78 |
| Total P (g/kg dw) | 0.74 | 0.63 |
| Available P (mg/kg dw) | 18.62 | 12.53 |
| Total K (g/kg dw) | 19.75 | 19.93 |
| Available K (mg/kg dw) | 209.64 | 218.53 |
| pH | 6.86 | 7.15 |
| SWC3 (%) | 11.40 | 11.48 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Sun, H.; Yang, W.; Chen, G.; Xu, H. Dynamics of Bacterial and Viral Communities in Paddy Soil with Irrigation and Urea Application. Viruses 2019, 11, 347. https://doi.org/10.3390/v11040347
Li Y, Sun H, Yang W, Chen G, Xu H. Dynamics of Bacterial and Viral Communities in Paddy Soil with Irrigation and Urea Application. Viruses. 2019; 11(4):347. https://doi.org/10.3390/v11040347
Chicago/Turabian StyleLi, Yuting, Hao Sun, Weichao Yang, Guanxiong Chen, and Hui Xu. 2019. "Dynamics of Bacterial and Viral Communities in Paddy Soil with Irrigation and Urea Application" Viruses 11, no. 4: 347. https://doi.org/10.3390/v11040347
APA StyleLi, Y., Sun, H., Yang, W., Chen, G., & Xu, H. (2019). Dynamics of Bacterial and Viral Communities in Paddy Soil with Irrigation and Urea Application. Viruses, 11(4), 347. https://doi.org/10.3390/v11040347