Effects of a Covert Infection with Phthorimaea operculella granulovirus in Insect Populations of Phthorimaea operculella

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Viruses
2.3. Co-Propagation of PhopGV-CR3 and PhopGV-GR1
2.4. Virus Purification
2.5. DNA Isolation from Occlusion Body
2.6. Whole DNA Isolation from P. operculella Eggs, Larvae, Pupae and Adults
2.7. Complete egt Gene Amplification
2.8. DNA Restriction Endonuclease Digests
2.9. Mixed Infection of Phop-IT Neonate Larvae
2.10. Crowding Experiments with Phop-IT
2.11. Whole Genome Sequencing of Isolates
3. Results
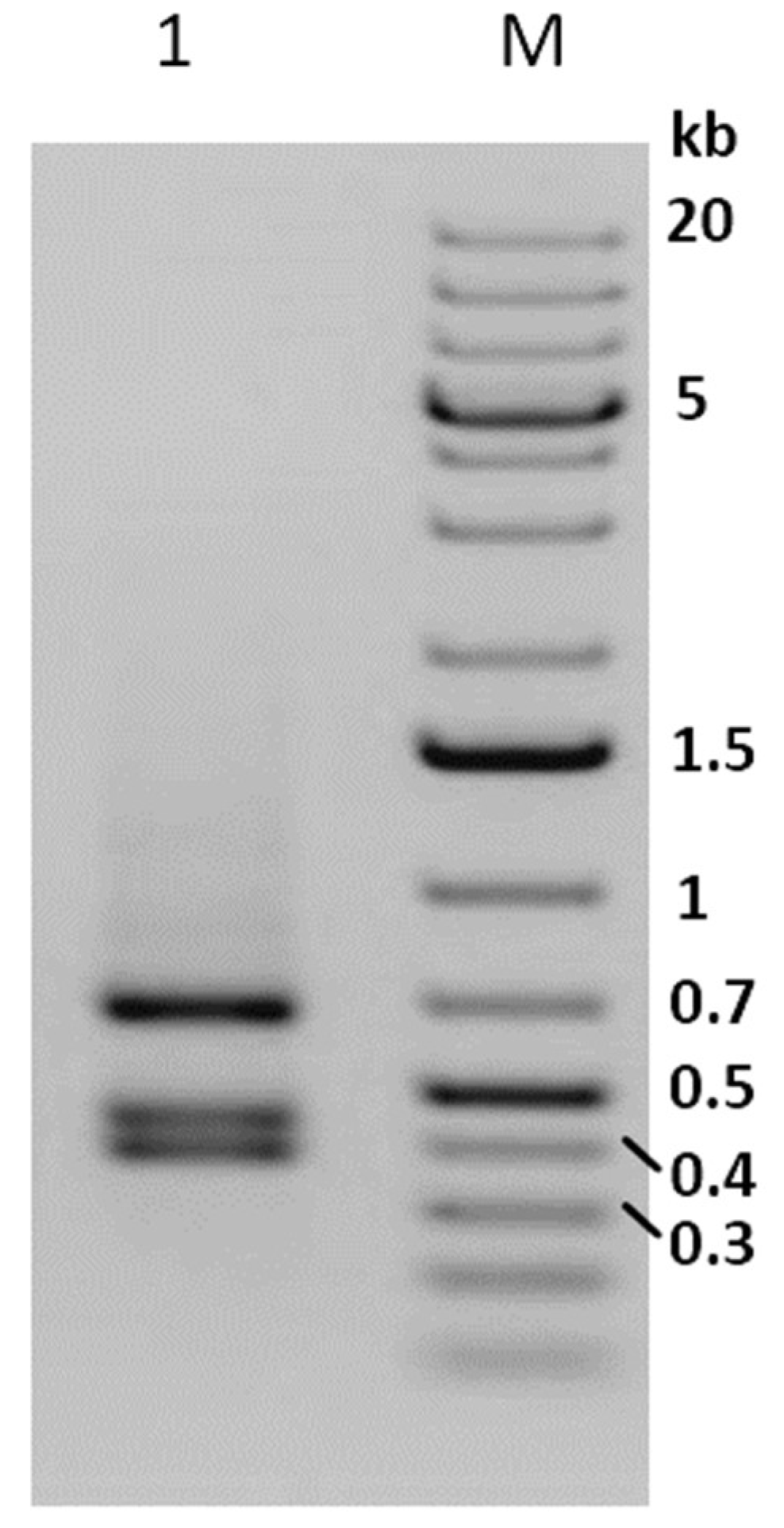
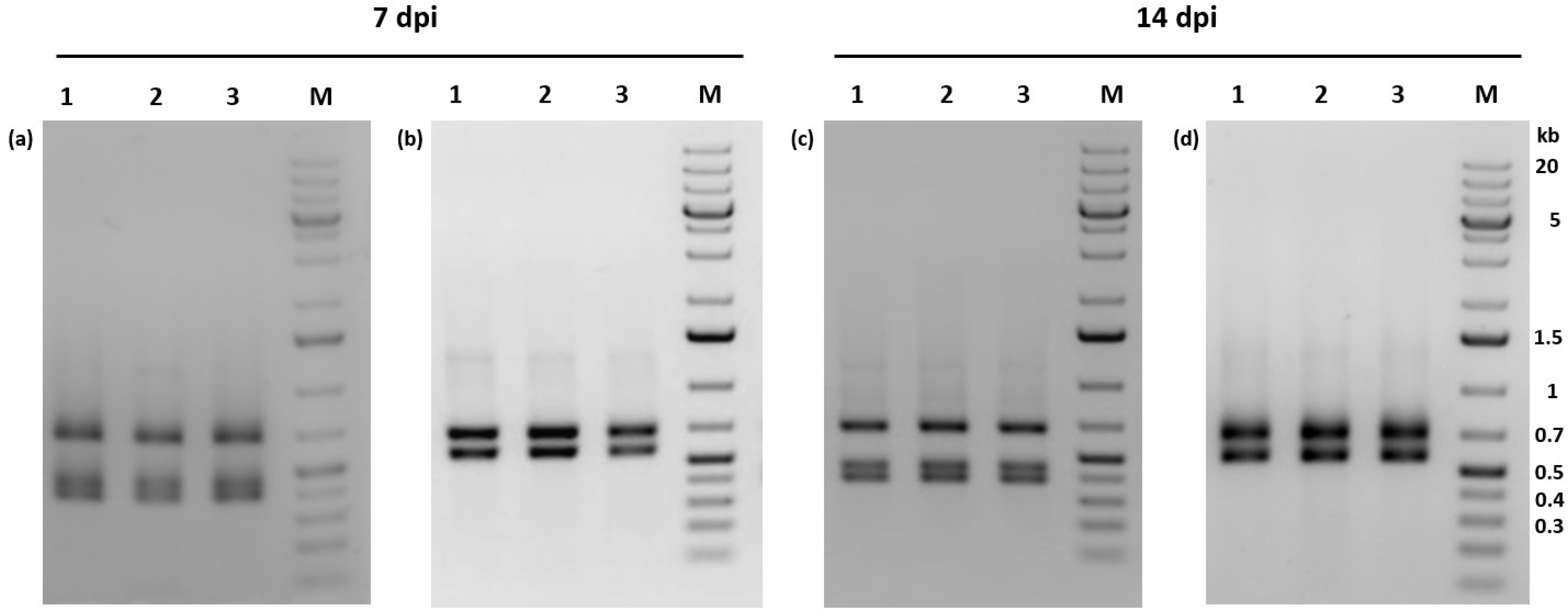
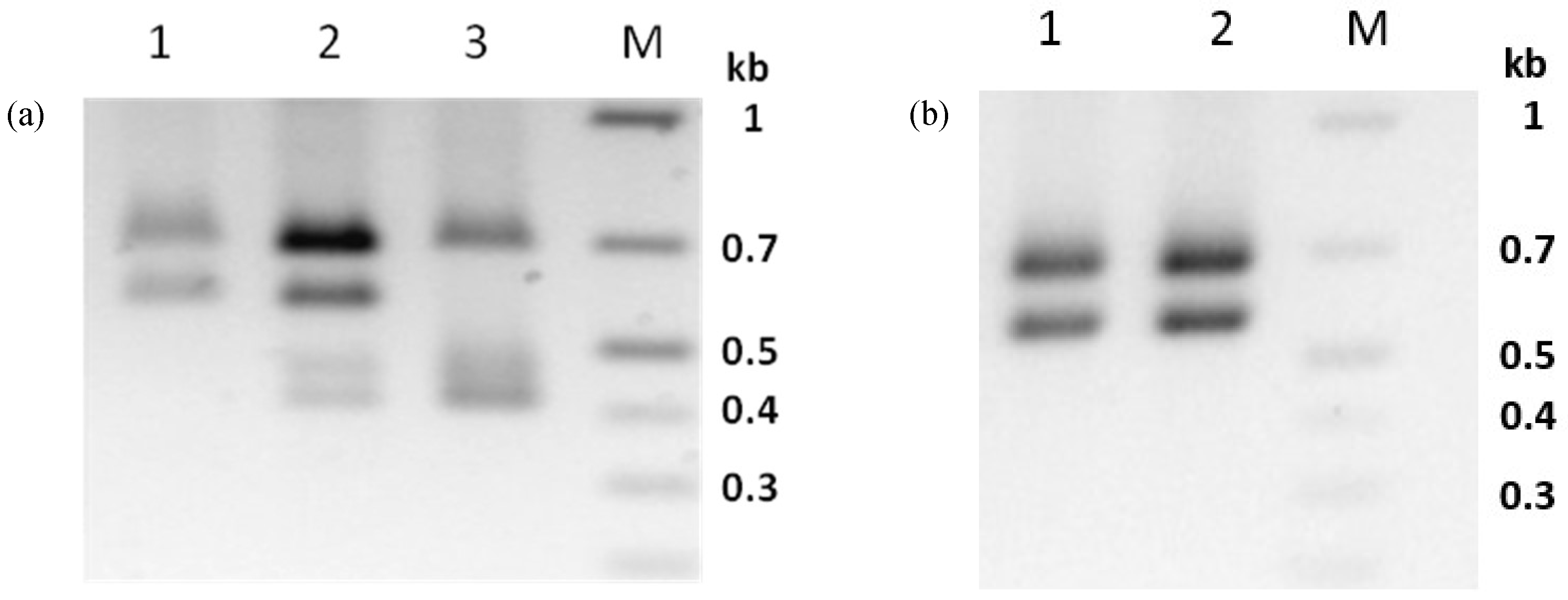
3.1. Identification of a Covert Infection in Phop-IT
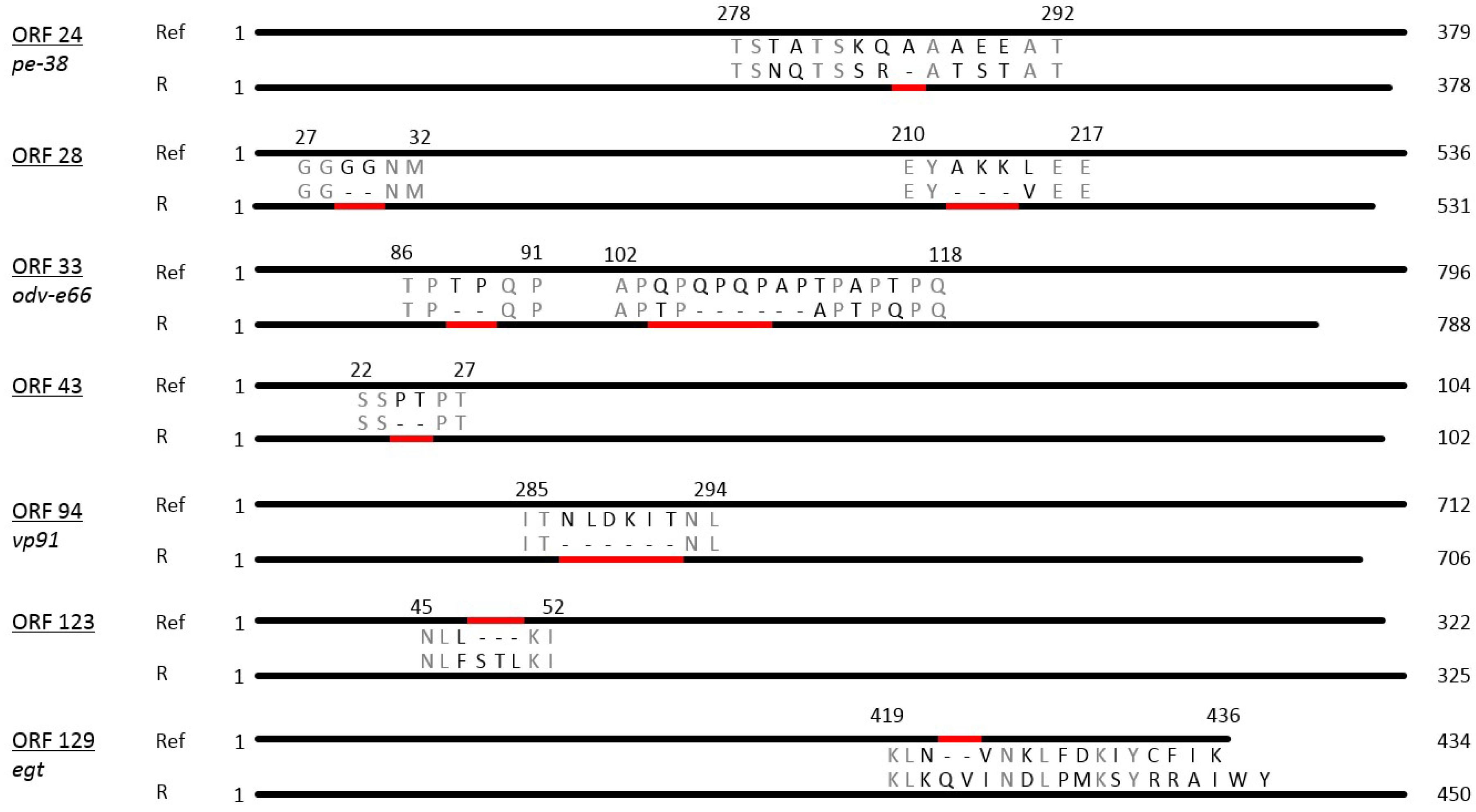
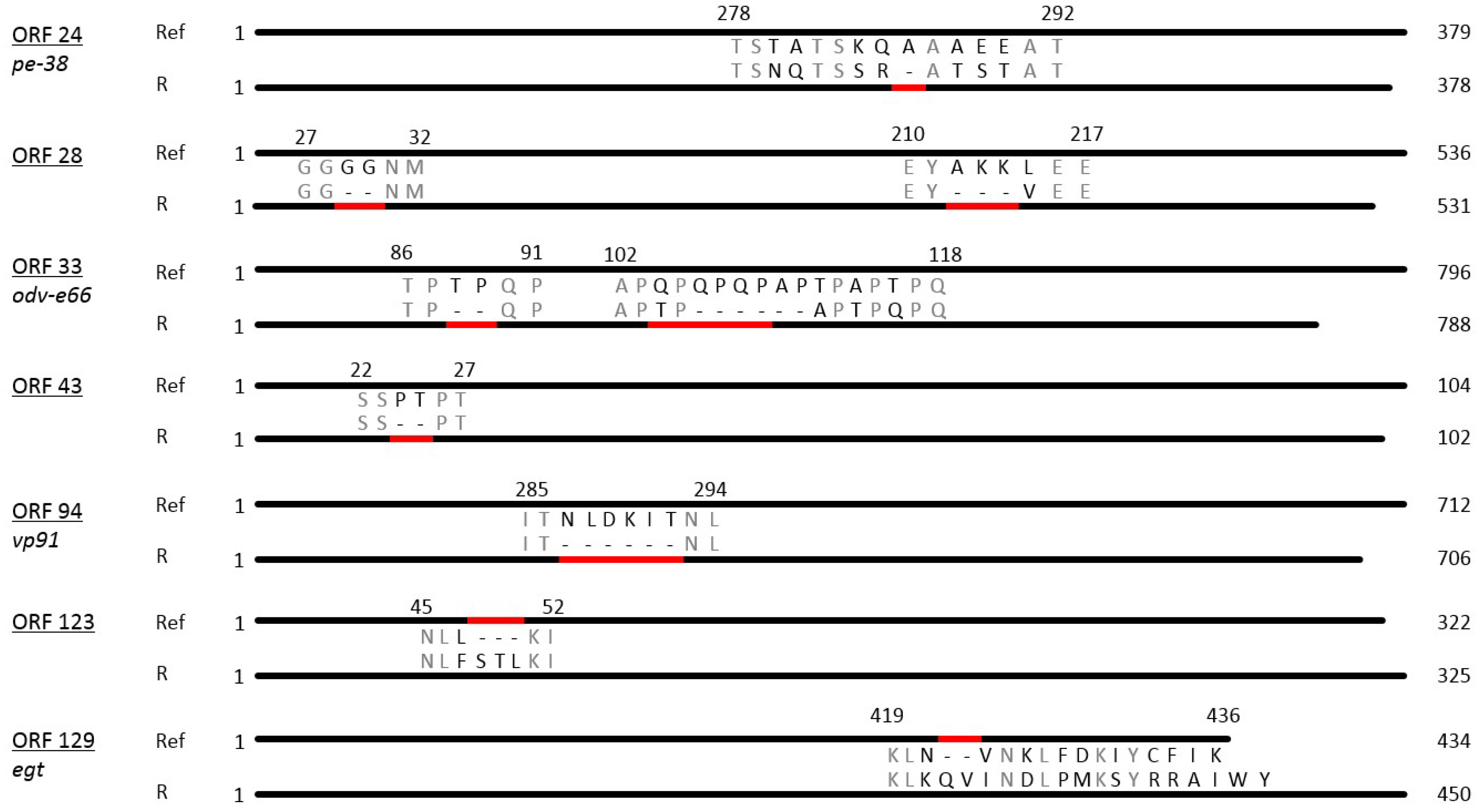
3.2. Sequence Analysis of PhopGV-R
3.3. Single Nucleotide Polymorphisms (SNPs)
3.4. Insertions and Deletions (Indels)
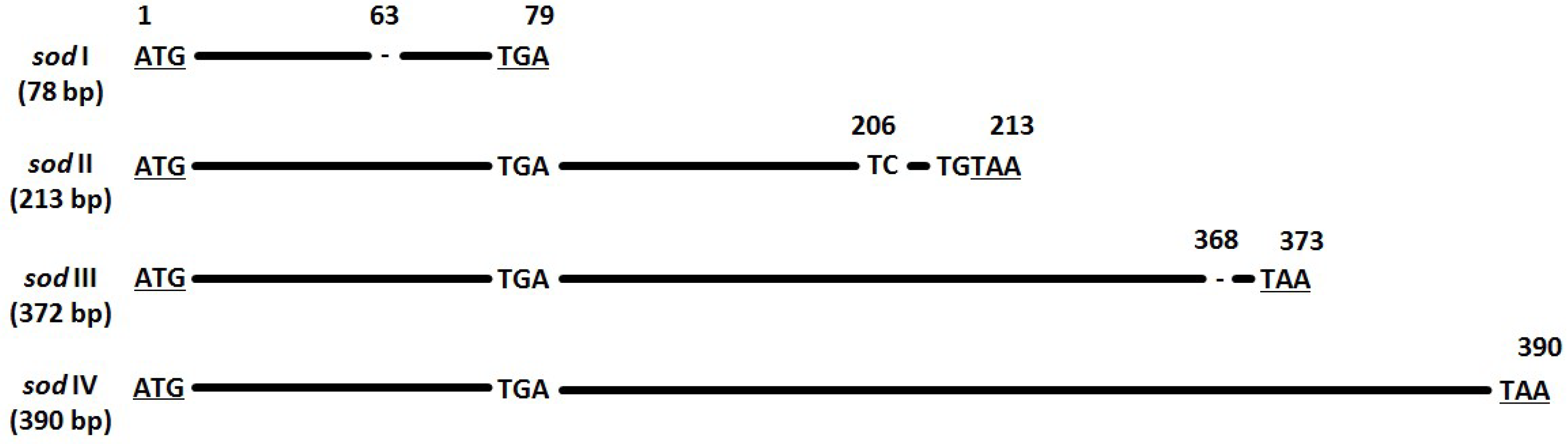
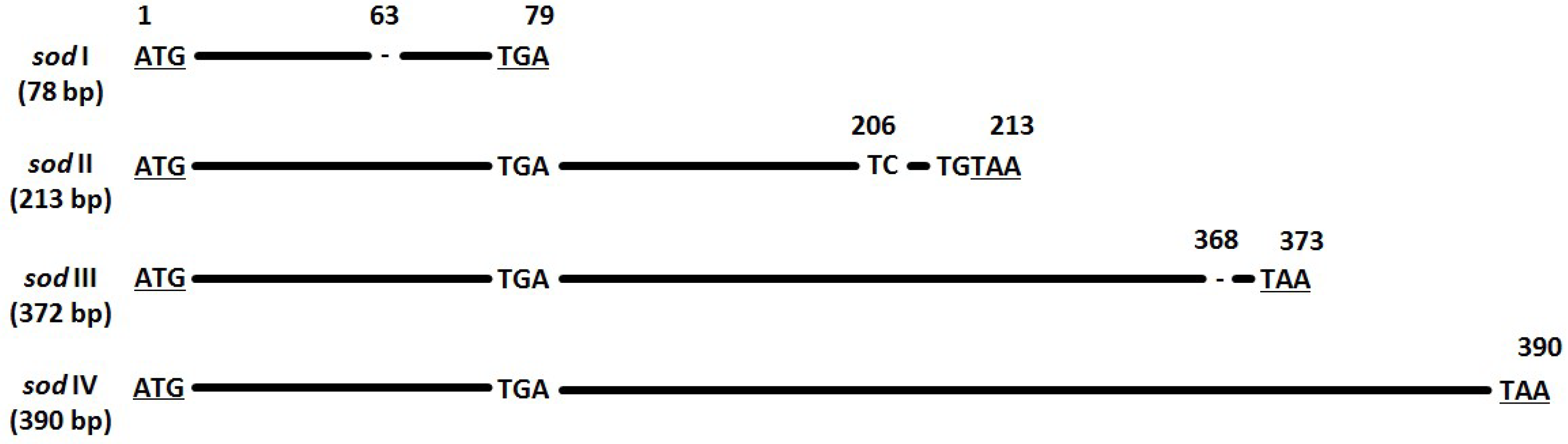
3.5. Sod Frequency
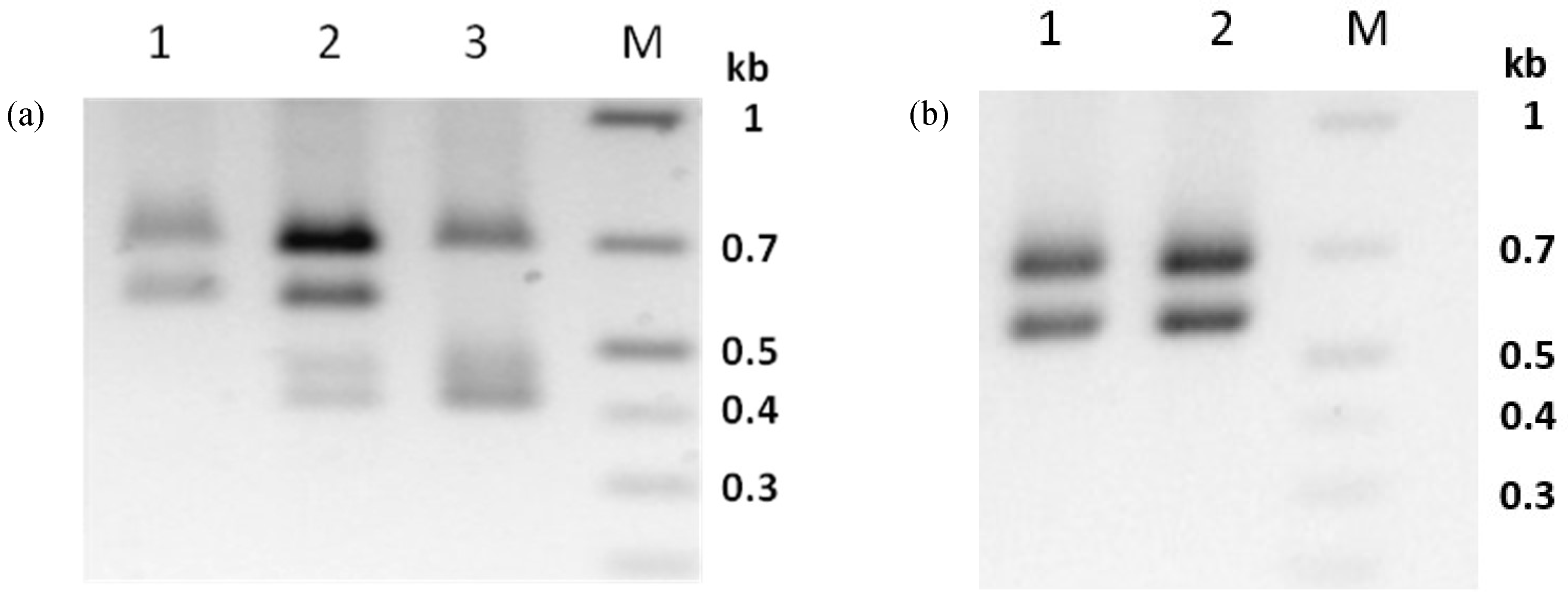
3.6. Virus Propagation of PhopGV-CR3 Resulted in Double Infections
3.7. PhopGV-GR1 Suppresses Replication of PhopGV-R
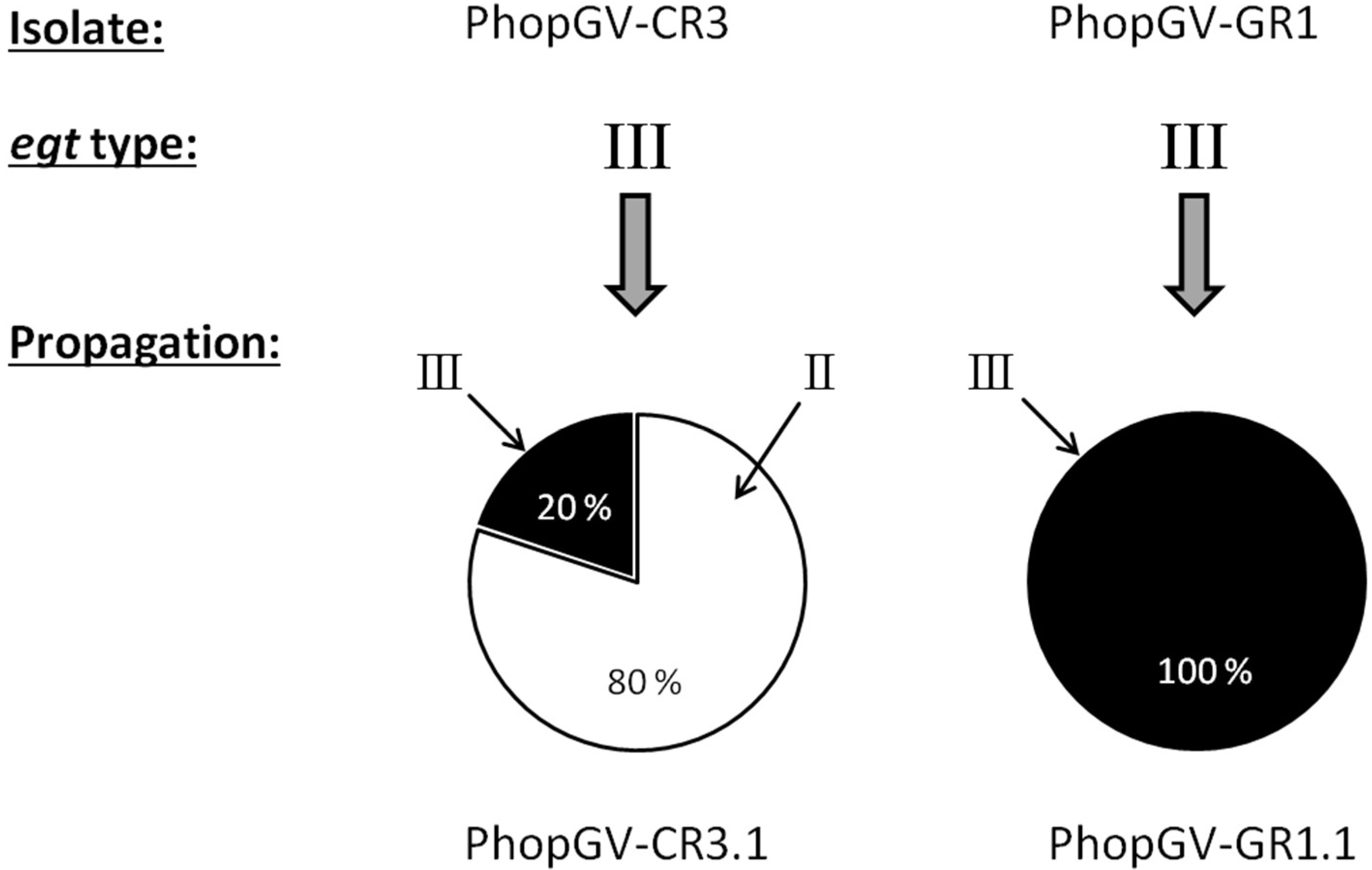
3.8. Co-Propagation of PhopGV-CR3 and PhopGV-GR1
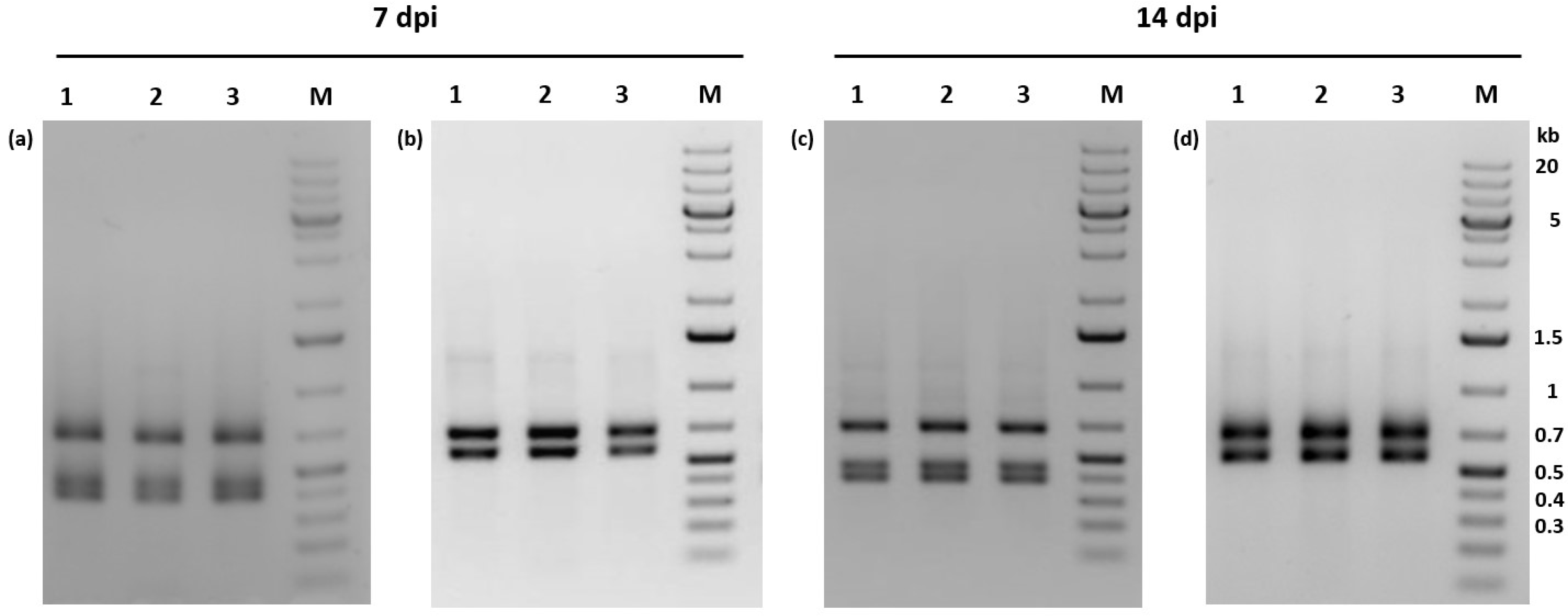
3.9. Crowding of Larvae Had No Effect on Overt Virus Infections
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andrealis, T.G. Transmission. In Epizootiology of Insect Diseases; Fuxa, J.R., Tanada, Y., Eds.; Wiley: New York, NY, USA, 1987; pp. 159–176. [Google Scholar]
- Burand, J.P. Nudiviruses. In The Insect Viruses; Miller, L.K., Ball, L.A., Eds.; Springer: Boston, MA, USA, 1998. [Google Scholar]
- Rohrmann, G. Baculovirus Molecular Biology, 2nd ed.; National Library of Medicine, NCBI: Bethesda, MD, USA, 2011.
- Cory, J.S. Insect virus transmission: Different routes to persistence. Curr. Opin. Insect Sci. 2015, 8, 130–135. [Google Scholar] [CrossRef]
- Valles, S.M.; Chen, Y.; Firth, A.E.; Guérin, D.M.A.; Hashimoto, Y.; Herrero, S.; de Miranda, J.R.; Ryabov, E.; ICTV Report Consortium. ICTV Virus Taxonomy Profile: Dicistroviridae. J. Gen. Virol. 2017, 98, 355–356. [Google Scholar] [CrossRef] [PubMed]
- Kukan, B. Vertical transmission of nucleopolyhedrovirus in insects. J. Invertebr. Pathol. 1999, 74, 103–111. [Google Scholar] [CrossRef]
- Mocarski, E.S.; Grakoui, A. Persistent and Latent Viral Infection. In Desk Encyclopedia of General Virology; Mahy, B.W.J., van Regenmortel, M.H.V., Eds.; Academic Press, Elsevier: Oxford, UK, 2010. [Google Scholar]
- Herniou, E.A.; Olszewski, J.A.; Cory, J.S.; O’Reilly, D.R. The genome sequence and evolution of baculoviruses. Annu. Rev. Entomol. 2003, 48, 211–243. [Google Scholar] [CrossRef]
- Sait, S.M.; Begon, M.; Thompson, D.J. The effects of a sublethal baculovirus infection in the Indian meal moth, Plodia interpunctella. J. Anim. Ecol. 1994, 63, 541–550. [Google Scholar] [CrossRef]
- Williams, T.; Virto, C.; Murillo, R.; Caballero, P. Covert Infection of Insects by Baculoviruses. Front. Microbiol. 2017, 8, 1337. [Google Scholar] [CrossRef] [PubMed]
- Murillo, R.; Hussey, M.S.; Possee, R.D. Evidence for covert baculovirus infections in a Spodoptera exigua laboratory culture. J. Gen. Virol. 2011, 92, 1061–1070. [Google Scholar] [CrossRef] [PubMed]
- Erlandson, M. Genetic variation in field populations of Baculovirus: Mechanisms for generating variation and its potential role in baculovirus epizootiology. Virol. Sin. 2009, 24, 458–469. [Google Scholar] [CrossRef]
- Burden, J.P.; Nixon, C.P.; Hodgkinson, A.E.; Possee, R.D.; Sait, S.M.; King, L.A.; Hails, R.S. Covert infections as a mechanism for long-term persistence of baculoviruses. Ecol. Lett. 2003, 6, 524–531. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Sun, J.Z.; Weidner, E.H.; LaMotte, L.R. Stressors and rearing diseases of Trichoplusia ni: Evidence of vertical transmission of NPV and CPV. J. Invertebr. Pathol. 1999, 74, 149–155. [Google Scholar] [CrossRef]
- Hughes, D.S.; Possee, R.D.; King, L.A. Evidence for the presence of a low-level persistent baculovirus infection of Mamestra brassica insects. J. Gen. Virol. 1997, 194, 604–615. [Google Scholar]
- Jukes, M.D.; Motsoeneng, B.M.; Knox, C.M.; Hill, M.P.; Moore, S.D. The comparative analysis of complete genome sequences from two South African betabaculoviruses: Phthorimaea operculella granulovirus and Plutella xylostella granulovirus. Arch. Virol. 2016, 161, 2917–2920. [Google Scholar] [CrossRef] [PubMed]
- Jukes, M.D.; Knox, C.M.; Hill, M.P.; Moore, S.D. The isolation and genetic characterization of a South African strain of Phthorimaea operculella granulovirus, PhopGV-SA. Virus Res. 2014, 183, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Opoku-Debrah, J.K.; Hill, M.P.; Knox, C.; Moore, S.D. Overcrowding of false codling moth, Thaumatotibia leucotreta (Meyrick) leads to the isolation of five new Cryptophlebia leucotreta granulovirus (CrleGV-SA) isolates. J. Invertebr. Pathol. 2013, 112, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Fuxa, J.R.; Richter, A.R.; Ameen, A.O.; Hammock, B.D. Vertical transmission of TnSNPV, TnCPV, AcMNPV, and possibly recombinant NPV in Trichoplusia ni. J. Invertebr. Pathol. 2002, 79, 44–50. [Google Scholar] [CrossRef]
- Hughes, D.S.; Possee, R.D.; King, L.A. Activation and detection of a latent baculovirus resembling Mamestra brassica nuclear polyhedrosis virus in M. brassica insects. Virology 1993, 194, 604–615. [Google Scholar] [CrossRef] [PubMed]
- Eldered, B.D.; Rehill, B.J.; Haynes, K.J.; Dwyer, G. Induced plant defences, host-pathogen interactions, and forest insect outbreaks. Proc. Natl. Acad. Sci. USA 2013, 110, 14978–14983. [Google Scholar] [CrossRef]
- Cory, J.S.; Myers, J.M. The ecology and evolution of insect baculoviruses. Annu. Rev. Ecol. Syst. 2003, 34, 239–272. [Google Scholar] [CrossRef]
- Anderson, R.; May, R.M. Infectious diseases and population cycles of forest insects. Science 1980, 210, 658–661. [Google Scholar] [CrossRef]
- Myers, J.H. Can a general hypothesis explain population cycles of forest Lepidoptera? Adv. Ecol. Res. 1988, 18, 179–184. [Google Scholar]
- Tanada, Y.; Fuxa, J.R. The pathogen population. In Epizootiology of Insect Diseases; Fuxa, J.R., Tanada, Y., Eds.; Wiley: New York, NY, USA, 1987. [Google Scholar]
- Anderson, R.; May, R.M. The population dynamics of microparasites and their invertebrate hosts. Philos. Trans. R. Soc. 1981, 291, 451–524. [Google Scholar] [CrossRef]
- Vilaplana, L.; Wilson, K.; Redman, E.M.; Cory, J.S. Pathogen persistence in migratory insects: High levels of vertically-transmitted virus infection in field populations of the African armyworm. Evol. Ecol. 2010, 24, 147–160. [Google Scholar] [CrossRef]
- Cooper, D.; Cory, J.; Theilmann, D.A. Nucleopolyhedroviruses of forest and western tent caterpillars: Cross-infectivity and evidence for activation of latent virus in high density field populations. Ecol. Entomol. 2003, 28, 41–50. [Google Scholar] [CrossRef]
- Steinhaus, E.A. Crowding as apossible stress factor in insect disease. Ecology 1958, 39, 503–514. [Google Scholar] [CrossRef]
- Ebert, D. The epidemiology and evolution of symbionts with mixed-mode transmission. Annu. Rev. Ecol. Evol. Syst. 2013, 44, 623–643. [Google Scholar] [CrossRef]
- Croizier, L.; Taha, A.; Croizier, G.; Lopez Ferber, M. Determination of the complete genome sequence of the potato tuber moth, Phthorimaea operculella granulovirus. Unpublished work. 2002. [Google Scholar]
- Carpio, C.; Dangles, O.; Dupas, S.; Léry, X.; López-Ferber, M.; Orbe, K.; Páez, D.; Rebaudo, F.; Santillán, A.; Yangari, B.; Zeddam, J.-L. Development of a viral biopesticide for the control of the Guatemala potato tuber moth Tecia solanivora. J. Invertebr. Pathol. 2013, 112, 184–191. [Google Scholar] [CrossRef]
- Lacey, L.A.; Kroschel, J. Microbial Control of the Potato Tuber Moth (Lepidoptera: Gelechiidae). Available online: http://www.globalsciencebooks.info/Online/GSBOnline/images/0906/FVCSB_3(SI1)/FVCSB_3(SI1)46-54o.pdf (accessed on 11 December 2018).
- Povolny, D. Genitalia of some nearctic and neotropic members of the tribe Gnorimoschemini (Lepidoptera, Gelechiidae). Acta Entomol. Musei Natl. Pragae 1967, 37, 51–127. [Google Scholar]
- Larem, A.; Wennmann, J.T.; Ben-Tiba, S.; Gueli-Alletti, G.; Jehle, J.A. Elucidating the genetic diversity of Phthorimaea operculella granulovirus (PhopGV). J. Gen. Virol. 2019, 100, 679–690. [Google Scholar] [CrossRef]
- Burand, J.P.; Park, E.J. Effect of Nuclear Polyhedrosis Virus Infection of the Development and Pupation of Gypsy Moth Larvae. J. Invertebr. Pathol. 1992, 60, 171–175. [Google Scholar] [CrossRef]
- O’Reilly, D.R.; Miller, L.K. A baculovirus blocks insect molting by producing ecdysteroid UDP-glycosyl transferase. Science 1989, 245, 1110–1112. [Google Scholar]
- Gómez-Bonilla, Y.; López-Ferber, M.; Caballero, P.; Léry, X.; Muñoz, D. Costa Rican soils contain highly insecticidal granulovirus strains against Phthorimaea opercullella and Tecia solanivora. J. Appl. Entomol. 2011, 136, 530–538. [Google Scholar] [CrossRef]
- Gómez Valderrama, J.A.; Barrera, G.; López-Ferber, M.; Belaich, M.; Ghiringhelli, P.D.; Villamizar, L. Potential of betabaculoviruses to control the tomato leafminer Tuta absoluta (Meyrick). J. Appl. Entomol. 2017, 1–11. [Google Scholar] [CrossRef]
- Lacey, L.A.; Hoffmann, D.F.; Federici, B. Histopathology and effect on development of the PhopGV on larvae of the potato tuber-moth, Phthorimaea operculella (Lepidoptera: Gelechiidae). J. Invertebr. Pathol. 2011, 108, 52–55. [Google Scholar] [CrossRef]
- Briese, D.T. The incidence of parasitism and disease in field populations of the potato moth Phthorimaea operculella (Zeller) in Australia. J. Aust. Entomol. Soc. 1981, 20, 319–326. [Google Scholar] [CrossRef]
- Reed, E.M. A granulosis virus of potato moth. Aust. J. Sci. 1969, 31, 300–301. [Google Scholar]
- Smith, I.R.; Crook, N.E. In vivo purification of Baculovirus Genotypes. Virology 1988, 166, 240–244. [Google Scholar] [CrossRef]
- Arends, H.M.; Jehle, J.A. Homologous recombination between the inverted terminal repeats of defective transposon TCp3.2 causes an inversion in the genome of Cydia pomonella granulovirus. J. Gen. Virol. 2002, 83, 1573–1578. [Google Scholar] [CrossRef]
- Larem, A. Diversity and Host Interaction of Phthorimaea operculella granulovirus. Ph.D. Thesis, TUPrints, Darmstadt, Germany, 2019; 147p. [Google Scholar]
- Javed, M.A.; Biswas, S.; Willis, L.G.; Harris, S.; Pritchard, C.; van Oers, M.M.; Donly, B.C.; Erlandson, M.A.; Hegedus, D.D.; Theilmann, D.A. Autographa californica Multiple Nucleopolyhedrovirus AC83 is a Per Os Infectivity Factor (PIF) Protein Required for Occlusion-Derived Virus (ODV) and Budded Virus Nucleocapsid Assembly as well as Assembly of the PIF Complex in ODV Envelopes. J. Virol. 2017, 91, 5. [Google Scholar] [CrossRef]
- Ferrelli, M.L.; Berretta, F.M.; Belaich, M.; Ghiringhelli, P.; Sciocco-Cap, A.; Romanowski, V. The Baculoviral Genome. In Viral Genomes—Molecular Structure, Diversity, Gene Expression Mechanisms and Host-VirusIinteractions; Garcia, L.M., Romanowski, V., Eds.; IntechOpen: London, UK, 2012; pp. 3–32. [Google Scholar] [CrossRef]
- Russel, R.L.; Rohrmann, G.F. Characterization of P91, a protein associated with virions of an Orygia pseudotsugata baculovirus. Virology 1997, 233, 210–223. [Google Scholar] [CrossRef]
- Fukai, T.; Ushio-Fukai, M. Superoxide Dismutases: Role in Redox Signaling, Vascular Function, and Diseases. Antioxid. Redox Signal. 2011, 15, 1583–1606. [Google Scholar] [CrossRef]
- Tomalski, M.D.; Eldridge, R.; Miller, L.K. A baculovirus homolog of a Cu/Zn superoxide dismutase gene. Viruses 1991, 184, 149–161. [Google Scholar]
- Larem, A. Gel Electrophoresis of PCR Products; Julius Kuehn Institute: Darmstadt, Germany, 2018; unpublished. [Google Scholar]
- Burden, J.P.; Possee, R.D.; Sait, S.M.; King, L.A.; Hails, R.S. Phenotypic and genotypic characterisation of persistent baculovirus infections in populations of the cabbage moth (Mamestra brassicae) within the British Isles. Arch. Virol. 2006, 151, 635–649. [Google Scholar] [CrossRef]
- Kelly, D.C.; Lescott, T.; Ayres, M.D. Induction of a non-occluded baculovirus persistently infecting Heliothis zea by Heliothis armigera and Trichoplusia ni nuclear poyhedrosis virus. Virology. 1981, 112, 174–180. [Google Scholar] [CrossRef]
- Beperet, I.; Irons, S.L.; Simón, O.; King, L.A.; Williams, T.; Possee, R.D.; López-Ferber, M.; Caballero, P. Superinfection exclusion in alphabaculovirus infections is concomitant with actin reorganization. J. Virol. 2014, 88, 3548–3556. [Google Scholar] [CrossRef]
- Lively, C.M.; Clay, K.; Wade, M.J. Competitive co-existence of vertical and horizontally transmitted parasites. Evol. Ecol. Res. 2005, 7, 1183–1190. [Google Scholar]
- Espinel-Correal, C.; Léry, X.; Villamizar, L.; Gómez, J.; Zeddam, J.L.; Cotes, A.M.; López-Ferber, M. Genetic and Biological Analysis of Colombian Phthorimaea operculella Granulovirus Isolated form Tecia solanivora (Lepidoptera: Gelechiidae). Appl. Environ. Microbiol. 2010, 7617–7625. [Google Scholar] [CrossRef]
- Cory, J.S.; Green, B.M.; Paul, R.K. Genotypic and phenotypic diversity of a baculovirus population within an individual insect host. J. Invertebr. Pathol. 2005, 89, 101–111. [Google Scholar] [CrossRef]
- Smith, P.H.; Vlak, J.M. Biological activity of Spodoptera exigua nuclear polyhedrovirus against S. exigua larvae. J. Invertebr. Pathol. 1988, 51, 107–114. [Google Scholar] [CrossRef]
- Moore, J. Parasites and the Behaviour of Animals; Oxford University Press: Oxford, UK, 2002. [Google Scholar]
- Earn, D.J.D.; Rohani, P.; Grenfell, B.J. Persistence, chaos and synchrony in ecology and epidemiology. Proc. R. Soc. Lond. B. Biol. Sci. 1998, 265, 7–10. [Google Scholar] [CrossRef]
- Swinton, J.; Harwood, J.; Grenfell, B.J. Persistence thresholds for phocine distemper virus infection in harbour seal Phoca vitulina metapopulations. J. Anim. Ecol. 1998, 67, 54–68. [Google Scholar] [CrossRef]
- Keeling, M.J.; Grenfell, B.T. Disease extinction and community size: Modelling the persistence of measles. Science 1997, 275, 65–67. [Google Scholar] [CrossRef]
- Jones, K.A.; Moawad, G.; McKinley, D.J.; Grzywacz, D. The effect of natural sunlight on Spodoptera littoralis nuclear polyhedrosis virus. Biocontrol Sci. Technol. 1993, 3, 189–194. [Google Scholar] [CrossRef]
- Biever, K.D.; Hostetter, D.L. Field persistence of Trichoplusia ni Lepidoptera Noctuidae single embedded nuclear polyhedrosis virus on cabbage foliage. Environ. Entomol. 1985, 14, 579–581. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate (PhopGV-) | Origin | Reads Assembled | Coverage/nt | Consensus Seq. Length | Accession No. | Source/Reference |
|---|---|---|---|---|---|---|
| R | Darmstadt | 1,163,983 | 1284 | 119,080 | MK033575 | JKI |
| CR3.1 | Costa Rica | 936,612 | 1187 | 118,355 | MK033565 | UPNA/[38] |
| GR1.1 | Greece | 1,647,866 | 2076 | 119,061 | MK033567 | Hellafarm |
| ORF | Encoding Gene | Direction | Position | Change (nt) | Change (aa) | Present | |
|---|---|---|---|---|---|---|---|
| CR3.1 | GR1.1 | ||||||
| 13 | p49 | reverse | 9514 | A -> G | Tyr -> His | + | |
| 24 | pe38 | reverse | 16,657 | T -> G | Lys -> Ans | − | |
| 27 | efp | forward | 21,377 | C -> A | Thr -> Lys | − | |
| 41 | MP-nase | reverse | 35,905 | C -> T | Asp -> Asn | + | |
| 54 | sod | reverse | 45,394 | G -> A | Leu -> Phe | − | |
| 54 | sod | reverse | 45,586 | A -> T | Cys -> Ser | + | + |
| 55 | p74 | reverse | 47,389 | C -> A | Arg -> Ile | − | |
| 66 | lef-1 | reverse | 56,063 | C -> T | Cys -> Tyr | − | |
| 66 | lef-1 | reverse | 56,420 | A -> G | Val -> Ala | − | |
| 82 | helicase-1 | forward | 68,953 | C -> G | Thr -> Arg | + | |
| 87 | lef-4 | reverse | 75,799 | G -> A | Ala -> Val | − | |
| 87 | lef-4 | reverse | 76,009 | C -> A | Ser -> Ile | + | + |
| 87 | lef-4 | reverse | 76,284 | C -> T | Met -> Ile | + | + |
| 87 | lef-4 | reverse | 76,364 | T -> C | Tyr -> Ala | − | |
| 88 | vp39 | forward | 77,124 | T -> G | Ser -> Ala | + | |
| 99 | vlf-1 | forward | 85,078 | T -> G | Tyr -> Asp | + | + |
| 99 | vlf-1 | forward | 85,232 | A -> C | Asn -> Thr | − | |
| 103 | DNApol | reverse | 87,530 | G -> C | Gln -> Glu | + | + |
| 105 | lef-3 | reverse | 93422 | G -> C | Asn -> Lys | + | + |
| 105 | lef-3(*) | reverse | 94,066 | A -> T | Leu -> Met | − | |
| 121 | lef-8 | reverse | 111,135 | C -> T | Gly -> Arg | + | |
| Larvae/Disc | 5 | 10 | 15 | 20 | 30 | 40 |
|---|---|---|---|---|---|---|
| N | 8 | 8 | 8 | 8 | 8 | 8 |
| n | 100 | 180 | 285 | 380 | 540 | 680 |
| % Larvae (s) | 1.1 ± 4.6 | 1.7 ± 3.9 | 1.8 ± 3.8 | 2.1 ± 3.8 | 3.2 ± 4.4 | 4.1 ± 4.2 |
| % Larvae (not s). | 9 ± 16.5 | 4.4 ± 12.5 | 4.6 ± 5.9 | 2.6 ± 4.5 | 2.8 ± 3.2 | 7.2 ± 9.8 |
| % Pupae | 67 ± 29.2a | 70.6 ± 21a | 68.1 ± 15.3a | 59 ± 15.8ab | 55.7 ± 10.3ab | 50.5 ± 11.5bc |
| % Living | 77 ± 25.4a | 76.1 ± 17.9ab | 73.7 ± 15.8ab | 63.7 ± 16.2ab | 61.8 ± 9.9ab | 59 ± 11.5b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Larem, A.; Ben Tiba, S.; Fritsch, E.; Undorf-Spahn, K.; Wennmann, J.T.; Jehle, J.A. Effects of a Covert Infection with Phthorimaea operculella granulovirus in Insect Populations of Phthorimaea operculella. Viruses 2019, 11, 337. https://doi.org/10.3390/v11040337
Larem A, Ben Tiba S, Fritsch E, Undorf-Spahn K, Wennmann JT, Jehle JA. Effects of a Covert Infection with Phthorimaea operculella granulovirus in Insect Populations of Phthorimaea operculella. Viruses. 2019; 11(4):337. https://doi.org/10.3390/v11040337
Chicago/Turabian StyleLarem, Andreas, Saoussen Ben Tiba, Eva Fritsch, Karin Undorf-Spahn, Jörg T. Wennmann, and Johannes A. Jehle. 2019. "Effects of a Covert Infection with Phthorimaea operculella granulovirus in Insect Populations of Phthorimaea operculella" Viruses 11, no. 4: 337. https://doi.org/10.3390/v11040337
APA StyleLarem, A., Ben Tiba, S., Fritsch, E., Undorf-Spahn, K., Wennmann, J. T., & Jehle, J. A. (2019). Effects of a Covert Infection with Phthorimaea operculella granulovirus in Insect Populations of Phthorimaea operculella. Viruses, 11(4), 337. https://doi.org/10.3390/v11040337