Annexin II as a Dengue Virus Serotype 2 Interacting Protein Mediating Virus Interaction on Vero Cells

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
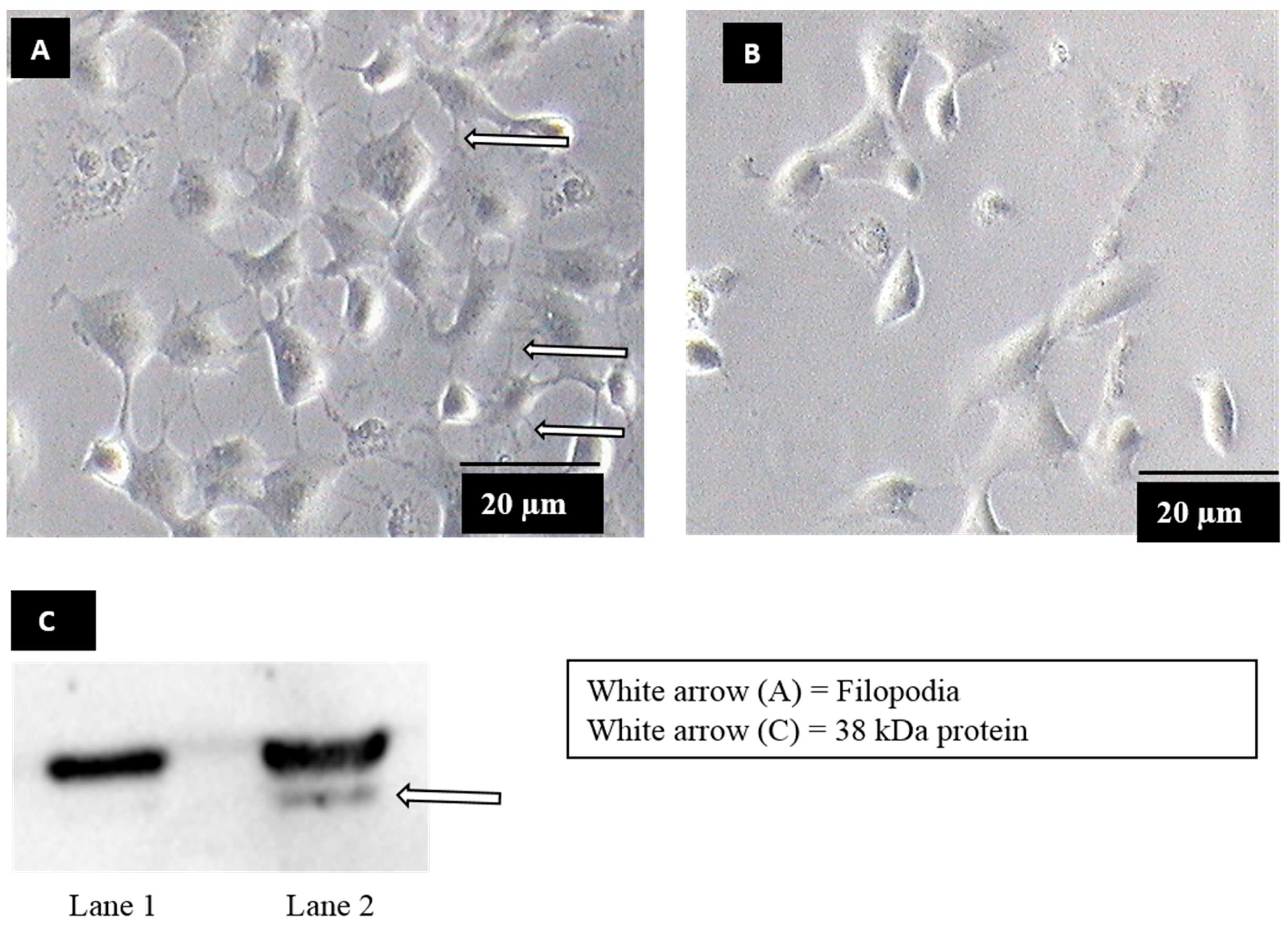
2.1. Induction of Filopodia Formation
2.2. Virus Overlay Protein Binding Assay (VOPBA) and Protein Identification
2.3. Flow Cytometry Analysis of Annexin II upon Dengue Virus Serotype 2 Exposure
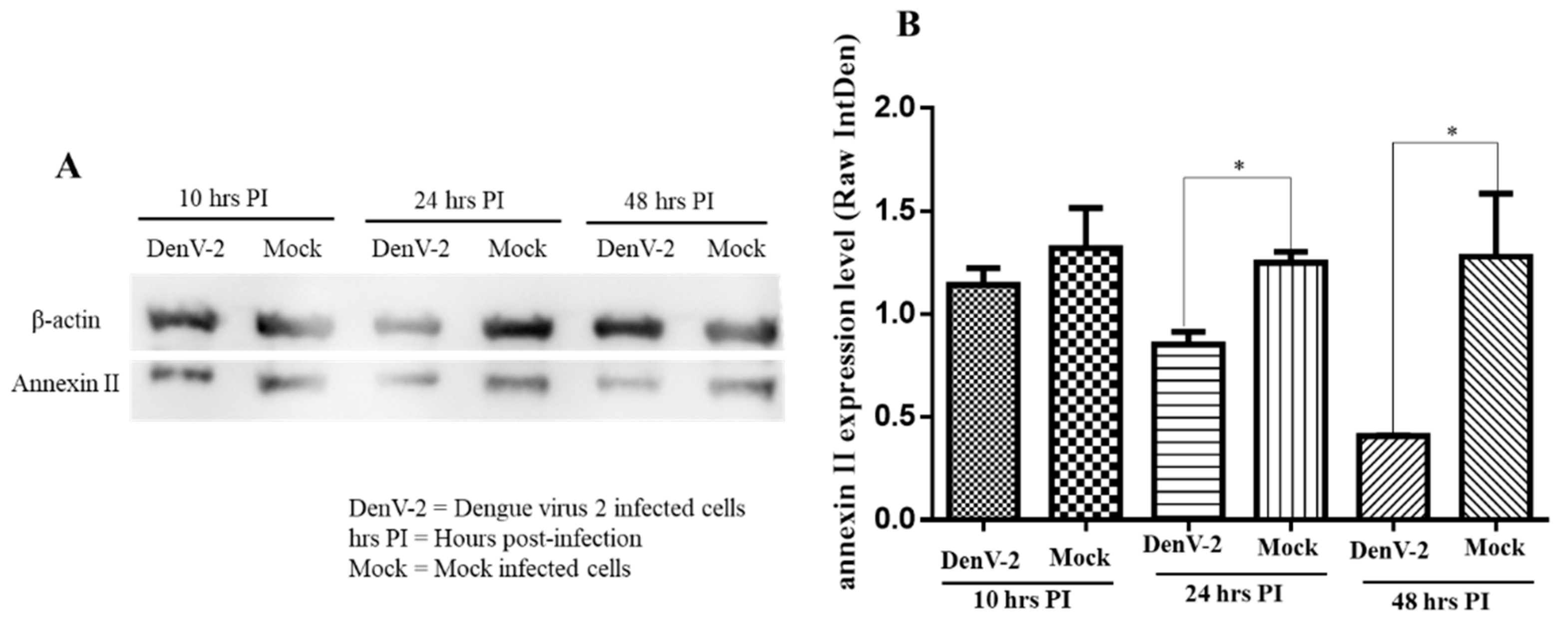
2.4. Western Blot Analysis of Annexin II upon Dengue Virus Serotype 2 Infection
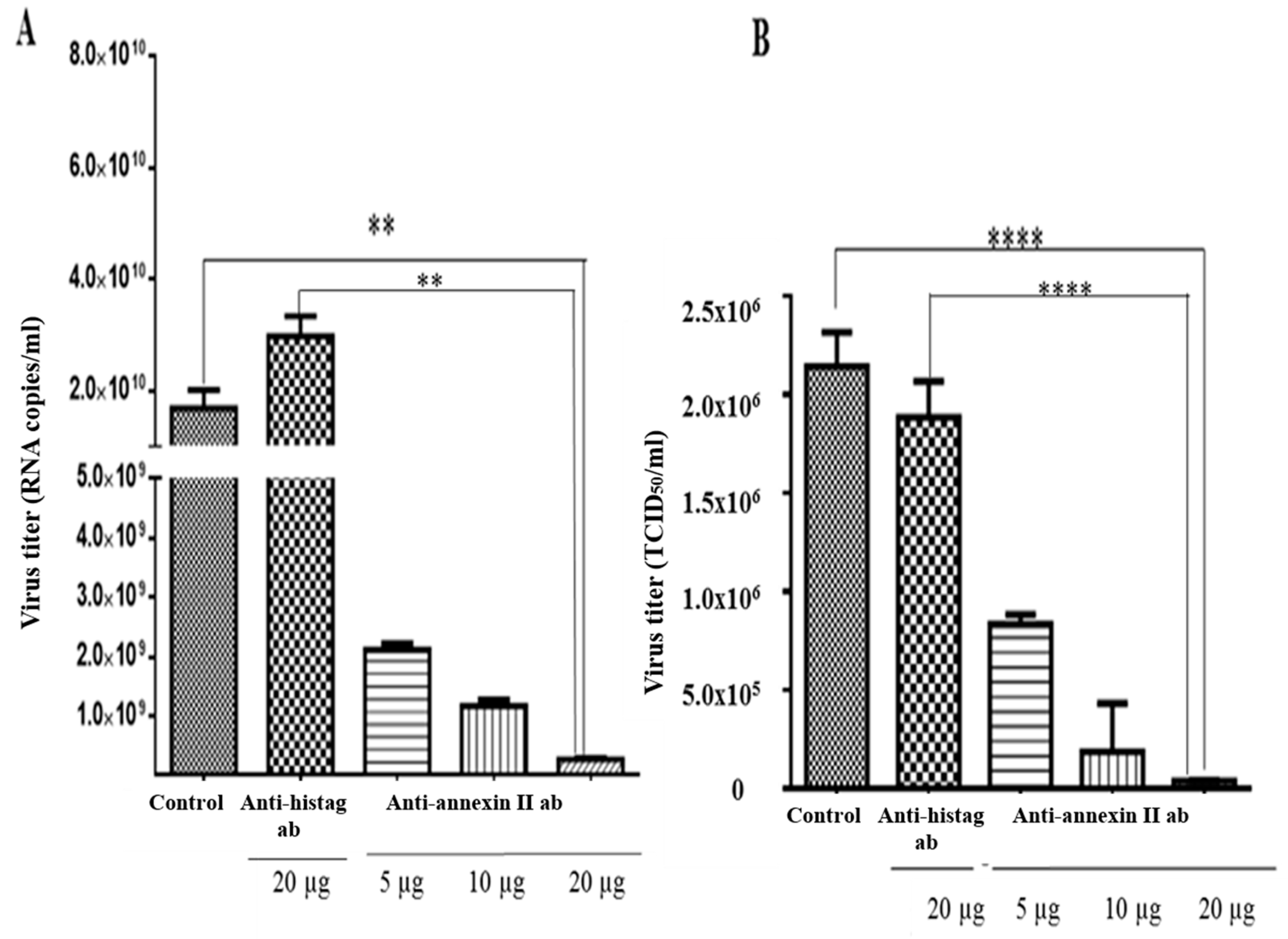
2.5. Antibody-Mediated Infections Inhibition Assay
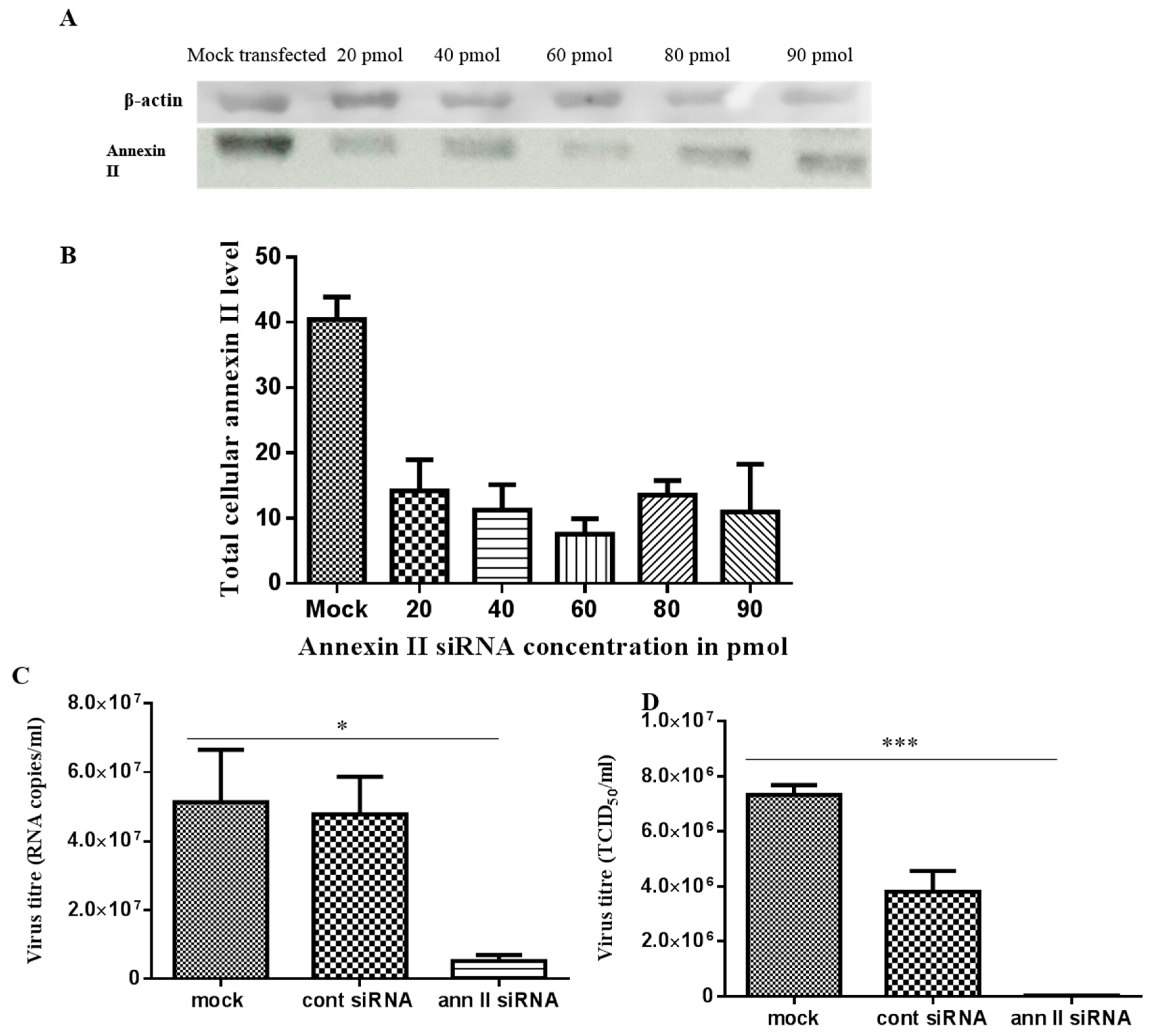
2.6. siRNA-mediated Annexin II Gene Silencing
2.7. siRNA Transfection and Dengue Virus Serotype 2 Infection
2.8. Statistical Tests
3. Results
3.1. Identification of Dengue Virus Serotype 2 Binding Protein on Filopodia
3.2. Detection of Annexin II upon Dengue Virus Serotype 2 Exposure
3.3. Dengue Virus Serotype 2 Interactions with Annexin II
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ligon, B.L. Dengue fever and dengue hemorrhagic fever: A review of the history, transmission, treatment, and prevention. Semin. Pediatric Infect. Dis. 2005, 16, 60–65. [Google Scholar] [CrossRef]
- Stanaway, J.D.; Shepard, D.S.; Undurraga, E.A.; Halasa, Y.A.; Coffeng, L.E.; Brady, O.J.; Hay, S.I.; Bedi, N.; Bensenor, I.M.; Castañeda-Orjuela, C.A.; et al. The global burden of dengue: An analysis from the Global Burden of Disease Study 2013. Lancet Infect. Dis. 2016, 16, 712–723. [Google Scholar] [CrossRef]
- Moi, M.L.; Takasaki, T.; Kurane, I. Human antibody response to dengue virus: Implications for dengue vaccine design. Trop. Med. Health 2016, 44. [Google Scholar] [CrossRef] [PubMed]
- Mongkolsapaya, J.; Dejnirattisai, W.; Xu, X.; Vasanawathana, S.; Tangthawornchaikul, N.; Chairunsri, A.; Sawasdivorn, S.; Duangchinda, T.; Dong, T.; Rowland-Jones, S.; et al. Original antigenic sin and apoptosis in the pathogenesis of dengue hemorrhagic fever. Nat. Med. 2003, 9, 921–927. [Google Scholar] [CrossRef] [PubMed]
- Azeredo, E.L.; Zagne, S.M.; Alvarenga, A.R.; Nogueira, R.M.; Kubelka, C.F.; Oliveira-Pinto, L.M. Activated peripheral lymphocytes with increased expression of cell adhesion molecules and cytotoxic markers are associated with dengue fever disease. Memórias Do Inst. Oswaldo Cruz 2006, 101, 437–449. [Google Scholar] [CrossRef]
- Guy, B.; Briand, O.; Lang, J.; Saville, M.; Jackson, N. Development of the Sanofi Pasteur tetravalent dengue vaccine: One more step forward. Vaccine 2015, 33, 7100–7111. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.J.; Rothman, A.L. Trials and Tribulations on the Path to Developing a Dengue Vaccine. Am. J. Prev. Med. 2015, 49, S334–S344. [Google Scholar] [CrossRef] [PubMed]
- de Chassey, B.; Meyniel-Schicklin, L.; Vonderscher, J.; André, P.; Lotteau, V. Virus-host interactomics: New insights and opportunities for antiviral drug discovery. Genome Med. 2014, 6. [Google Scholar] [CrossRef] [PubMed]
- Haqqani, A.A.; Tilton, J.C. Entry inhibitors and their use in the treatment of HIV-1 infection. Antivir. Res. 2013, 98, 158–170. [Google Scholar] [CrossRef]
- Thepparit, C.; Smith, D.R. Serotype-Specific Entry of Dengue Virus into Liver Cells: Identification of the 37-Kilodalton/67-Kilodalton High-Affinity Laminin Receptor as a Dengue Virus Serotype 1 Receptor. J. Virol. 2004, 78, 12647–12656. [Google Scholar] [CrossRef] [PubMed]
- Tio, P.H.; Jong, W.W.; Cardosa, M.J. Two dimensional VOPBA reveals laminin receptor (LAMR1) interaction with dengue virus serotypes 1, 2 and 3. Virol. J. 2005, 2, 25. [Google Scholar] [CrossRef]
- Chen, Y.C.; Wang, S.Y.; King, C.C. Bacterial lipopolysaccharide inhibits dengue virus infection of primary human monocytes/macrophages by blockade of virus entry via a CD14-dependent mechanism. J. Virol. 1999, 73, 2650–2657. [Google Scholar] [PubMed]
- Martinez-Barragan, J.J.; del Angel, R.M. Identification of a Putative Coreceptor on Vero Cells That Participates in Dengue 4 Virus Infection. J. Virol. 2001, 75, 7818–7827. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Oliveira, C.; Freire, J.M.; Conceição, T.M.; Higa, L.M.; Castanho, M.A.R.B.; Da Poian, A.T. Receptors and routes of dengue virus entry into the host cells. FEMS Microbiol. Rev. 2015, 39, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Lidke, D.S.; Ozbun, M.A. Virus activated filopodia promote human papillomavirus type 31 uptake from the extracellular matrix. Virology 2008, 381, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Ewers, H.; Smith, A.E.; Sbalzarini, I.F.; Lilie, H.; Koumoutsakos, P.; Helenius, A. Single-particle tracking of murine polyoma virus-like particles on live cells and artificial membranes. Proc. Natl. Acad. Sci. USA 2005, 102, 15110–15115. [Google Scholar] [CrossRef]
- Pelkmans, L. Local Actin Polymerization and Dynamin Recruitment in SV40-Induced Internalization of Caveolae. Science 2002, 296, 535–539. [Google Scholar] [CrossRef]
- Favoreel, H.W.; Enquist, L.; Feierbach, B. Actin and Rho GTPases in herpesvirus biology. Trends Microbiol. 2007, 15, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Zamudio-Meza, H.; Castillo-Alvarez, A.; Gonzalez-Bonilla, C.; Meza, I. Cross-talk between Rac1 and Cdc42 GTPases regulates formation of filopodia required for dengue virus type-2 entry into HMEC-1 cells. J. Gen. Virol. 2009, 90, 2902–2911. [Google Scholar] [CrossRef]
- Low, F.C.Y.; AbuBakar, S.; Othman, F.; Chee, H.Y. Ultrastructural aspects of sylvatic dengue virus infection in Vero cell. Malays. J. Pathol. 2019. accepted. [Google Scholar]
- Chee, H.-Y.; AbuBakar, S. Identification of a 48kDa tubulin or tubulin-like C6/36 mosquito cells protein that binds dengue virus 2 using mass spectrometry. Biochem. Biophys. Res. Commun. 2004, 320, 11–17. [Google Scholar] [CrossRef]
- Bringans, S.; Eriksen, S.; Kendrick, T.; Gopalakrishnakone, P.; Livk, A.; Lock, R.; Lipscombe, R. Proteomic analysis of the venom ofHeterometrus longimanus (Asian black scorpion). Proteomics 2008, 8, 1081–1096. [Google Scholar] [CrossRef] [PubMed]
- Idris, F.; Muharram, S.H.; Zaini, Z.; Diah, S. Effectiveness of physical inactivation methods of dengue virus: Heat- versus UV-inactivation. bioRxiv 2018. [Google Scholar] [CrossRef]
- Tang, L.I.; Ling, A.P.; Koh, R.Y.; Chye, S.M.; Voon, K.G. Screening of anti-dengue activity in methanolic extracts of medicinal plants. BMC Complementary Altern. Med. 2012, 12. [Google Scholar] [CrossRef]
- Najioullah, F.; Viron, F.; Césaire, R. Evaluation of four commercial real-time RT-PCR kits for the detection of dengue viruses in clinical samples. Virol. J. 2014, 11, 164. [Google Scholar] [CrossRef]
- Sakoonwatanyoo, P.; Boonsanay, V.; Smith, D.R. Growth and Production of the Dengue Virus in C6/36 Cells and Identification of a Laminin-Binding Protein as a Candidate Serotype 3 and 4 Receptor Protein. Intervirology 2006, 49, 161–172. [Google Scholar] [CrossRef]
- Kozma, R.; Ahmed, S.; Best, A.; Lim, L. The Ras-related protein Cdc42Hs and bradykinin promote formation of peripheral actin microspikes and filopodia in Swiss 3T3 fibroblasts. Mol. Cell. Biol. 1995, 15, 1942–1952. [Google Scholar] [CrossRef] [PubMed]
- Petrovic, N.; Schacke, W.; Gahagan, J.R.; O’Conor, C.A.; Winnicka, B.; Conway, R.E.; Mina-Osorio, P.; Shapiro, L.H. CD13/APN regulates endothelial invasion and filopodia formation. Blood 2007, 110, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Gerke, V.; Moss, S.E. Annexins: From Structure to Function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Chu, J.J.H.; Leong, P.W.H.; Ng, M.L. Analysis of the endocytic pathway mediating the infectious entry of mosquito-borne flavivirus West Nile into Aedes albopictus mosquito (C6/36) cells. Virology 2006, 349, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Eash, S.; Atwood, W.J. Involvement of Cytoskeletal Components in BK Virus Infectious Entry. J. Virol. 2005, 79, 11734–11741. [Google Scholar] [CrossRef]
- Lehmann, M.J.; Sherer, N.M.; Marks, C.B.; Pypaert, M.; Mothes, W. Actin- and myosin-driven movement of viruses along filopodia precedes their entry into cells. J. Cell Biol. 2005, 170, 317–325. [Google Scholar] [CrossRef]
- Mercer, J.; Helenius, A. Vaccinia Virus Uses Macropinocytosis and Apoptotic Mimicry to Enter Host Cells. Science 2008, 320, 531–535. [Google Scholar] [CrossRef]
- Sharma-Walia, N.; Naranatt, P.P.; Krishnan, H.H.; Zeng, L.; Chandran, B. Kaposi’s Sarcoma-Associated Herpesvirus/Human Herpesvirus 8 Envelope Glycoprotein gB Induces the Integrin-Dependent Focal Adhesion Kinase-Src-Phosphatidylinositol 3-Kinase-Rho GTPase Signal Pathways and Cytoskeletal Rearrangements. J. Virol. 2004, 78, 4207–4223. [Google Scholar] [CrossRef]
- Coyne, C.B.; Bergelson, J.M. Virus-Induced Abl and Fyn Kinase Signals Permit Coxsackievirus Entry through Epithelial Tight Junctions. Cell 2006, 124, 119–131. [Google Scholar] [CrossRef]
- Deora, A.B.; Kreitzer, G.; Jacovina, A.T.; Hajjar, K.A. An Annexin 2 Phosphorylation Switch Mediates p11-dependent Translocation of Annexin 2 to the Cell Surface. J. Biol. Chem. 2004, 279, 43411–43418. [Google Scholar] [CrossRef]
- Dziduszko, A.; Ozbun, M.A. Annexin A2 and S100A10 Regulate Human Papillomavirus Type 16 Entry and Intracellular Trafficking in Human Keratinocytes. J. Virol. 2013, 87, 7502–7515. [Google Scholar] [CrossRef]
- Derry, M.C.; Sutherland, M.R.; Restall, C.M.; Waisman, D.M.; Pryzdial, E.L.G. Annexin 2-mediated enhancement of cytomegalovirus infection opposes inhibition by annexin 1 or annexin 5. J. Gen. Virol. 2007, 88, 19–27. [Google Scholar] [CrossRef]
- Breiner, K.M.; Urban, S.; Glass, B.; Schaller, H. Envelope Protein-Mediated Down-Regulation of Hepatitis B Virus Receptor in Infected Hepatocytes. J. Virol. 2001, 75, 143–150. [Google Scholar] [CrossRef]
- Fongsaran, C.; Jirakanwisal, K.; Kuadkitkan, A.; Wikan, N.; Wintachai, P.; Thepparit, C.; Ubol, S.; Phaonakrop, N.; Roytrakul, S.; Smith, D.R. Involvement of ATP synthase β subunit in chikungunya virus entry into insect cells. Arch. Virol. 2014, 159, 3353–3364. [Google Scholar] [CrossRef]
- Yang, S.-L.; Chou, Y.-T.; Wu, C.-N.; Ho, M.-S. Annexin II binds to capsid protein VP1 of enterovirus 71 and enhances viral infectivity. J. Virol. 2011, 85, 11809–11820. [Google Scholar] [CrossRef]
- Ryzhova, E.V.; Vos, R.M.; Albright, A.V.; Harrist, A.V.; Harvey, T.; Gonzalez-Scarano, F. Annexin 2: A Novel Human Immunodeficiency Virus Type 1 Gag Binding Protein Involved in Replication in Monocyte-Derived Macrophages. J. Virol. 2006, 80, 2694–2704. [Google Scholar] [CrossRef]
- Saxena, V.; Lai, C.-K.; Chao, T.-C.; Jeng, K.-S.; Lai, M.M.C. Annexin A2 Is Involved in the Formation of Hepatitis C Virus Replication Complex on the Lipid Raft. J. Virol. 2012, 86, 4139–4150. [Google Scholar] [CrossRef]
- LeBouder, F.; Morello, E.; Rimmelzwaan, G.F.; Bosse, F.; Pechoux, C.; Delmas, B.; Riteau, B. Annexin II Incorporated into Influenza Virus Particles Supports Virus Replication by Converting Plasminogen into Plasmin. J. Virol. 2008, 82, 6820–6828. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/N | Ref. XP | Mass | Score | Matches | Sequence | emPAI | Protein Identity | Species |
|---|---|---|---|---|---|---|---|---|
| 1 | V9HW65 | 38552 | 2672 | 125 (72) | 28 (21) | 18.63 | Human annexin II | Homo sapiens |
| A0A096NM87 | 38580 | 2672 | 125 (72) | 28 (21) | 18.63 | Papan annexin II | Papio anubis | |
| X008014586.1 | 38580 | 2672 | 125 (72) | 28 (21) | 18.63 | Annexin II | Cholocebus sabaeus | |
| 2 | A0A096NCGO | 35959 | 2085 | 82 (52) | 21 (16) | 31.54 | Papan Glyceraldehyde-3-phosphate dehydrogenase | Papio anubus |
| 3 | F6ZXWS | 31457 | 2076 | 85 (51) | 20 (15) | 58.60 | Glyceraldehyde-3-phosphate dehydrogenase | Mecaca malutta |
| 4 | A0A0D9SDK6 | 38721 | 1829 | 77 (49) | 15 (10) | 2.65 | Annexin II | Chlorocebus sabaeus |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aliyu, I.A.; Ling, K.-H.; Md Hashim, N.F.; Lam, J.-Y.; Chee, H.-Y. Annexin II as a Dengue Virus Serotype 2 Interacting Protein Mediating Virus Interaction on Vero Cells. Viruses 2019, 11, 335. https://doi.org/10.3390/v11040335
Aliyu IA, Ling K-H, Md Hashim NF, Lam J-Y, Chee H-Y. Annexin II as a Dengue Virus Serotype 2 Interacting Protein Mediating Virus Interaction on Vero Cells. Viruses. 2019; 11(4):335. https://doi.org/10.3390/v11040335
Chicago/Turabian StyleAliyu, Isah Abubakar, King-Hwa Ling, Nur Fariesha Md Hashim, Jia-Yong Lam, and Hui-Yee Chee. 2019. "Annexin II as a Dengue Virus Serotype 2 Interacting Protein Mediating Virus Interaction on Vero Cells" Viruses 11, no. 4: 335. https://doi.org/10.3390/v11040335
APA StyleAliyu, I. A., Ling, K.-H., Md Hashim, N. F., Lam, J.-Y., & Chee, H.-Y. (2019). Annexin II as a Dengue Virus Serotype 2 Interacting Protein Mediating Virus Interaction on Vero Cells. Viruses, 11(4), 335. https://doi.org/10.3390/v11040335