Infectious Virions of Bombyx Mori Latent Virus Are Incorporated into Bombyx Mori Nucleopolyhedrovirus Occlusion Bodies

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines, Viruses, Larvae, and Plaque Assays
2.2. Co-Infection of Bombyx mori latent virus (BmLV) and Bombyx mori nucleopolyhedrovirus (BmNPV) in B. Mori larvae
2.3. Detection of BmLV from BmNPV Occlusion Bodies (OBs)
2.4. Infection of BmLV Incorporated in BmNPV OBs
3. Results and Discussion
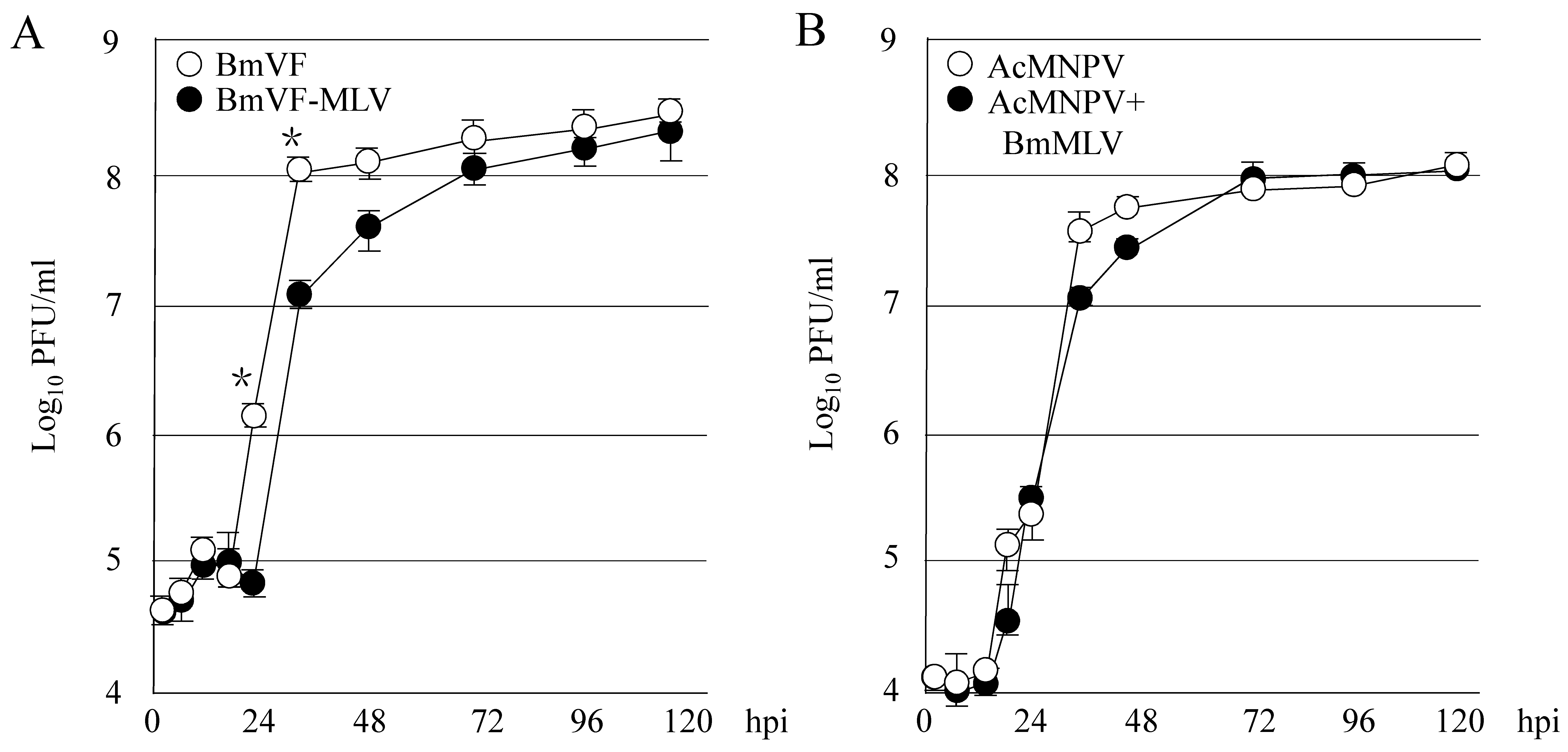
3.1. Persistent Infection with BmLV Affects the Multiplication of BmNPV in B. Mori Cultured Cells
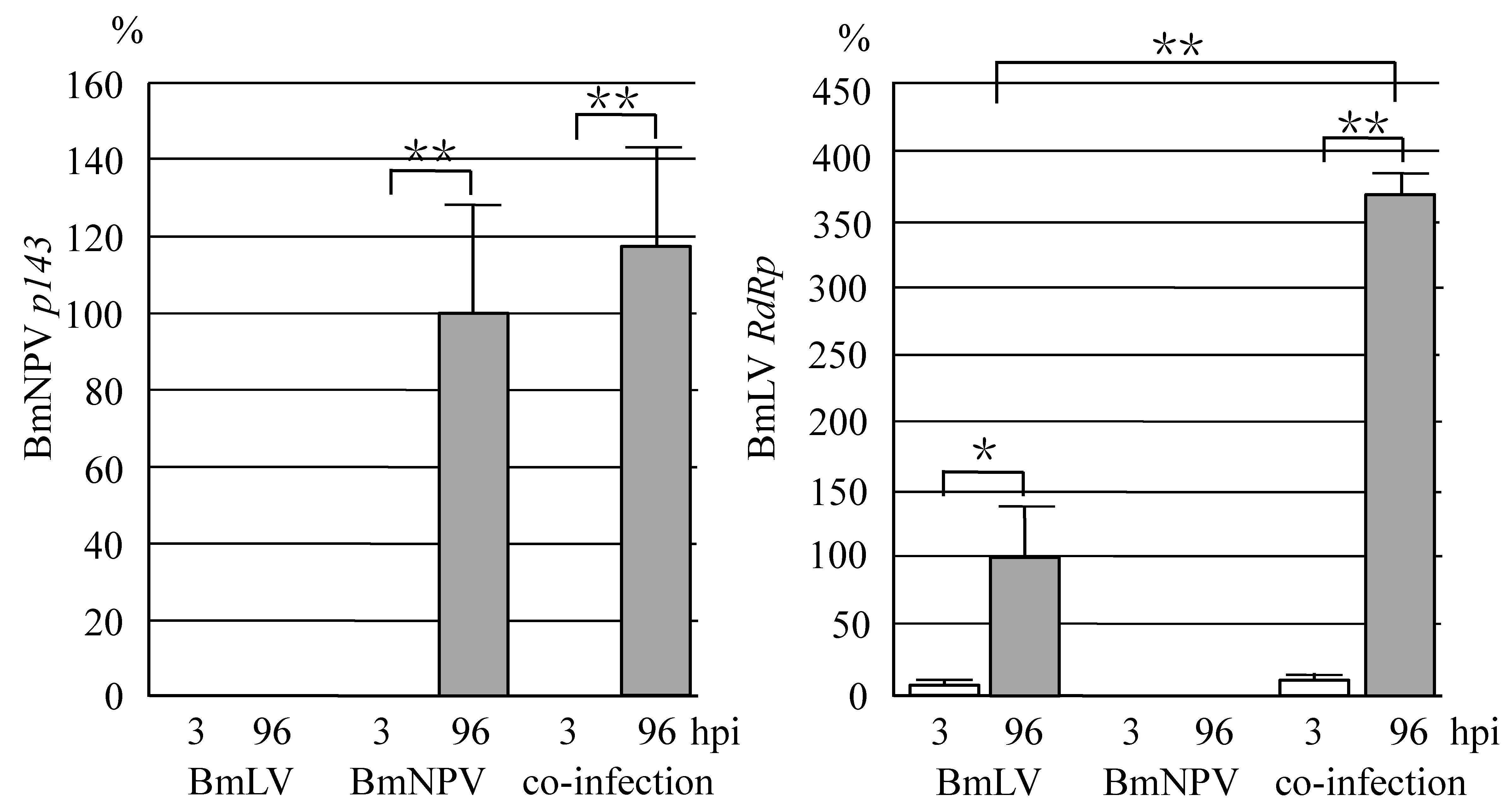
3.2. Co-Infection with BmLV and BmNPV Causes Proliferation of BmLV in B. Mori larvae
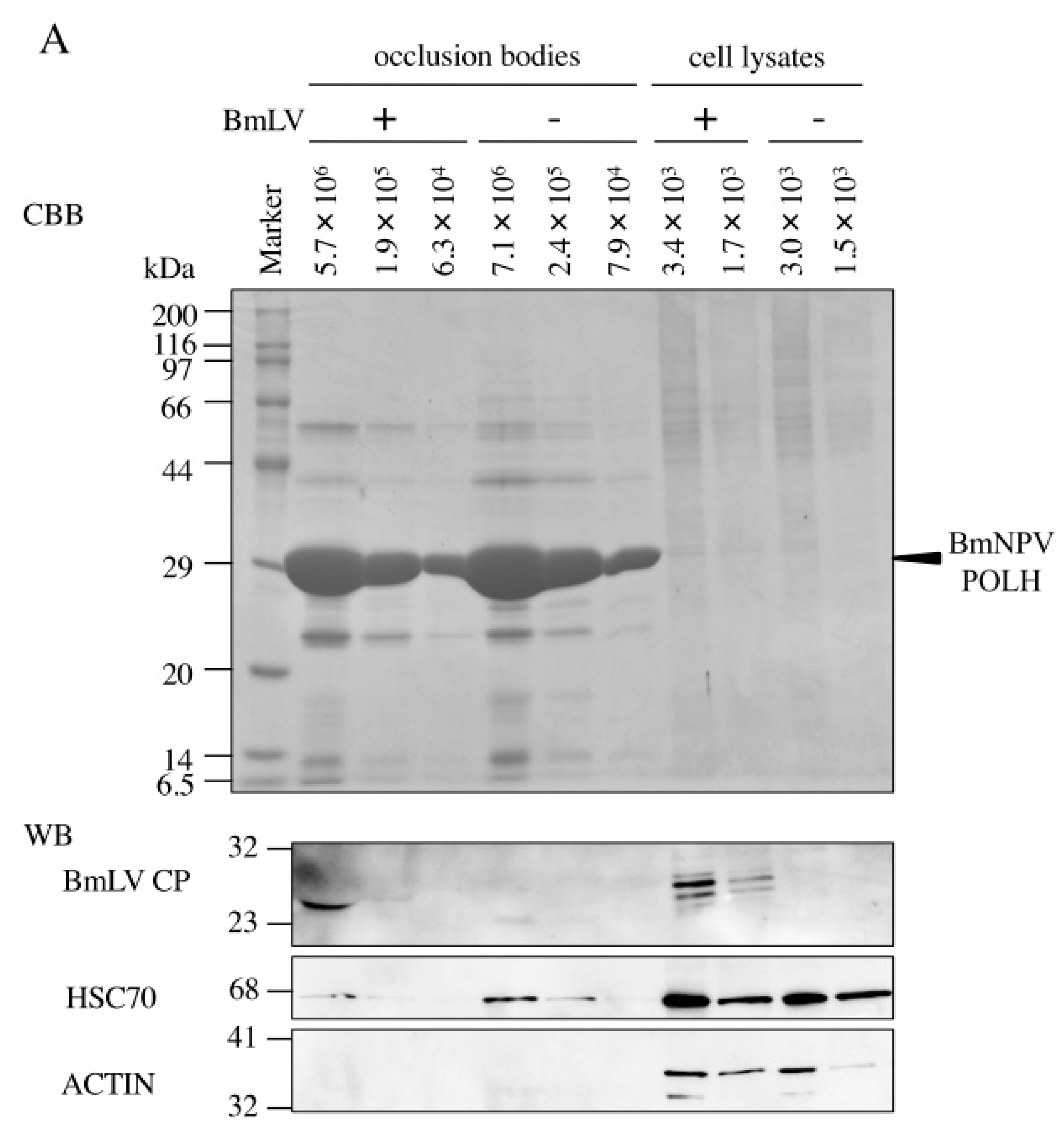
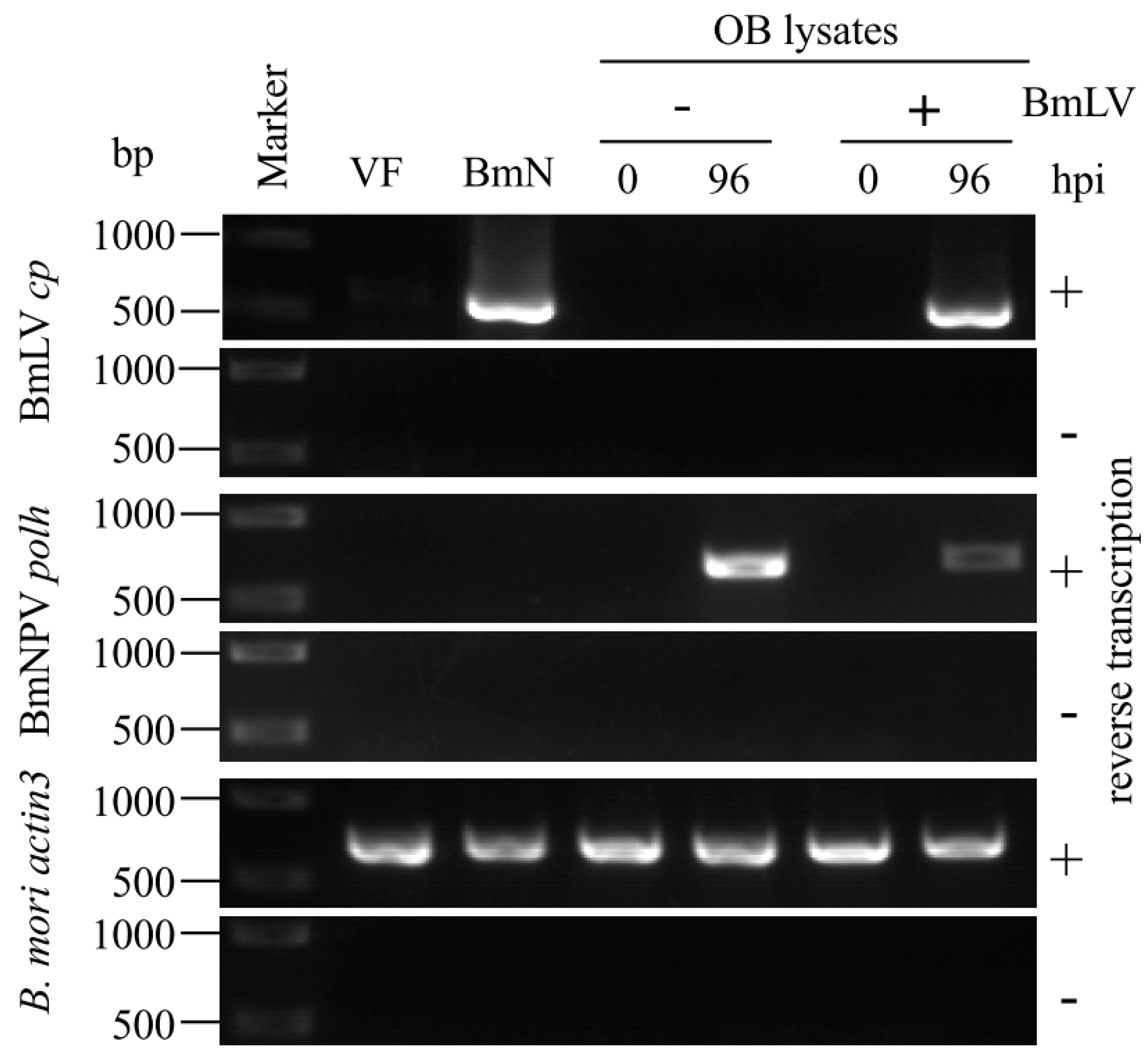
3.3. BmLV Infectious Virions are Incorporated into OBs of BmNPV
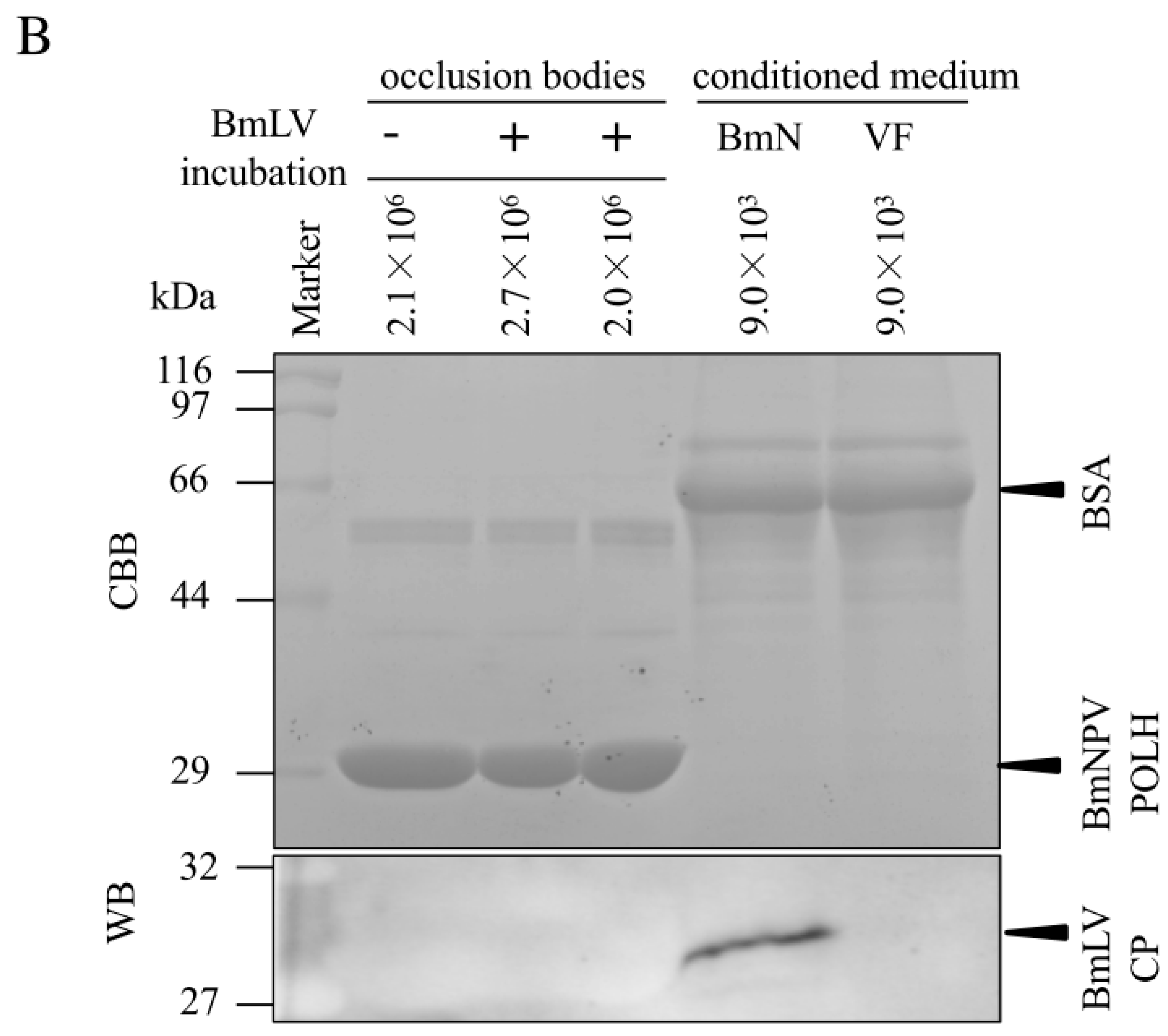
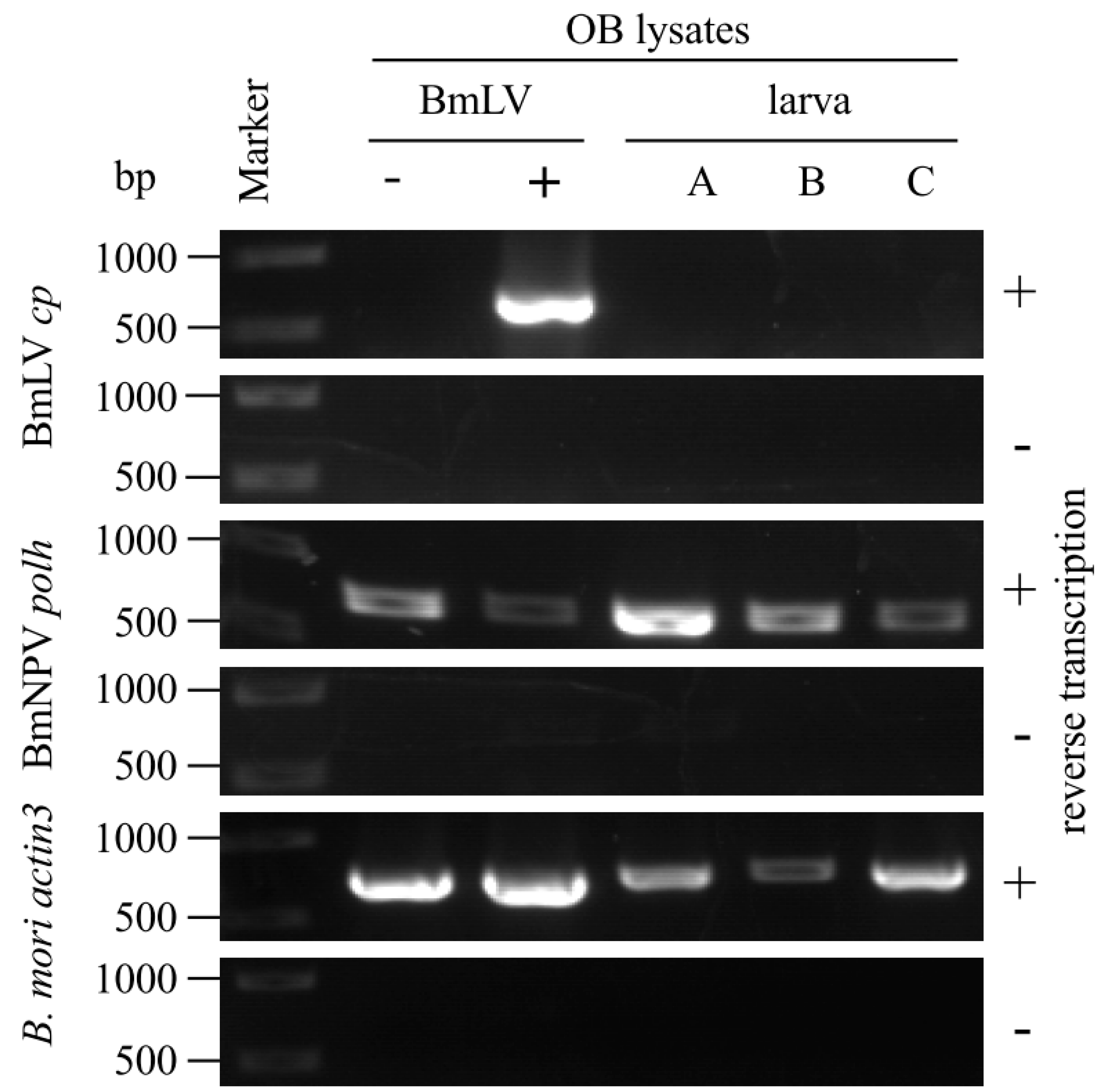
3.4. Incorporation of BmLV into BmNPV OBs Is not Maintained in B. Mori larvae
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martelli, G.P.; Sabanadzovic, S.; Abou Ghanem-Sabanadzovic, N.; Saldarelli, P. Maculavirus, a new genus of plant viruses. Arch. Virol. 2002, 147, 1847–1853. [Google Scholar] [CrossRef]
- Katsuma, S.; Tanaka, S.; Omuro, N.; Takabuchi, L.; Daimon, T.; Imanishi, S.; Yamashita, S.; Iwanaga, M.; Mita, K.; Maeda, S.; et al. Novel Macula-like virus identified in Bombyx mori cultured cells. J. Virol. 2005, 79, 5577–5584. [Google Scholar] [CrossRef] [PubMed]
- King, A.M.Q.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Commettee on Taxonomy of Viruses (2018). Arch. Virol. 2018, 163, 2601–26131. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, M. Persistent virus in the silkworm cell lines: Bombyx mori macula-like virus (BmMLV). Sanshi-Konchu Biotec 2012, 81, 139–148. (In Japanese) [Google Scholar] [CrossRef]
- Iwanaga, M.; Hitotsuyama, T.; Katsuma, S.; Ishihara, G.; Daimon, T.; Shimada, T.; Imanishi, S.; Kawasaki, H. Infection study of Bombyx mori macula-like virus (BmMLV) using a BmMLV-negative cell line and an infectious clone. J. Virol. Methods 2012, 179, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Innami, K.; Aizawa, T.; Tsukui, T.; Katsuma, S.; Imanishi, S.; Kawasaki, H.; Iwanaga, M. Infection studies of nontarget mammalian cell lines with Bombyx mori macula-like virus. J. Virol. Methods 2016, 229, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Katsuma, S.; Kawamoto, M.; Shoji, K.; Aizawa, T.; Kiuchi, T.; Izumi, N.; Ogawa, M.; Mashiko, T.; Kawasaki, H.; Sugano, S.; et al. Transcriptome profiling reveals infection strategy of an insect maculavirus. DNA Res. 2018, 25, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herniou, E.A.; Luque, T.; Chen, X.; Vlak, J.M.; Winstanley, D.; Cory, J.S.; O’Reilly, D.R. Use of whole genome sequence data to infer baculovirus phylogeny. J. Virol. 2001, 75, 8117–8126. [Google Scholar] [CrossRef]
- Braunagel, S.C.; Summers, M.D. Autographa californica nuclear polyhedrosis virus, PDV, and ECV viral envelopes and nucleocapsids: Structural proteins, antigens, lipid and fatty acid profiles. Virology 1994, 202, 315–328. [Google Scholar] [CrossRef]
- Rohrmann, G.F. The baculovirus replication cycle: Effects on cells and insects. In Baculovirus Molecular Biology, 3rd ed.; National Center for Biotechnology Information: Bethesda, MD, USA, 2013. [Google Scholar]
- Jakubowska, A.K.; Murillo, R.; Carballo, A.; Williams, T.; van Lent, J.W.; Caballero, P.; Herrero, S. Iflavirus increases its infectivity and physical stability in association with baculovirus. Peer J. 2016, 4, e1687. [Google Scholar] [CrossRef]
- Carballo, A.; Murillo, R.; Jakubowska, A.; Herrero, S.; Williams, T.; Caballero, P. Co-infection with iflaviruses influences the insecticidal properties of Spodoptera exigua multiple nucleopolyhedrovirus occlusion bodies: Implications for the production and biosecurity of baculovirus insecticides. PLoS ONE 2017, 12, e0177301. [Google Scholar] [CrossRef]
- Iwanaga, M.; Takaya, K.; Katsuma, S.; Ote, M.; Tanaka, S.; Kamita, S.G.; Kang, W.; Shimada, T.; Kobayashi, M. Expression profiling of baculovirus genes in permissive and nonpermissive cell lines. Biochem. Biophys. Res. Commun. 2004, 323, 599–614. [Google Scholar] [CrossRef]
- Iwanaga, M.; Tsukui, K.; Uchiyama, K.; Katsuma, S.; Imanishi, S.; Kawasaki, H. Expression of recombinant proteins by BEVS in a macula-like virus-free silkworm cell line. J. Invertebr. Pathol. 2014, 123, 34–37. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, S.; Kokusho, R.; Kakemizu, H.; Izaku, T.; Katsuma, S.; Iwashita, Y.; Kawasaki, H.; Iwanaga, M. Characterization of a Bombyx mori nucleopolyhedrovirus variant isolated in Laos. J. Insect Biotech. Sericol. 2017, 86, 85–94. [Google Scholar] [CrossRef]
- Uchiyama, K.; Fujimoto, H.; Katsuma, S.; Imanishi, S.; Kato, A.; Kawasaki, H.; Iwanaga, M. Inactivation of Bombyx mori macula-like virus under physical conditions. In Vitro Cell. Dev. Biol. Anim. 2016, 52, 265–270. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, D.R.; Miller, L.K.; Luckow, V.A. Cotransfection and recombinant virus identification. In Baculovirus Expression Vectors: A Laboratory Manual; W. H. Freeman & Co.: New York, NY, USA, 1992; pp. 139–179. [Google Scholar]
- Iwanaga, M.; Shibano, Y.; Ohsawa, T.; Fujita, T.; Katsuma, S.; Kawasaki, H. Involvement of HSC70-4 and other inducible HSPs in Bombyx mori nucleopolyhedrovirus infection. Virus Res. 2014, 179, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Okano, K.; Shimada, T.; Mita, K.; Maeda, S. Comparative expressed-sequence-tag analysis of differential gene expression profiles in BmNPV-infected BmN cells. Virology 2001, 282, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Iwanaga, M.; Shimada, T.; Kobayashi, M.; Kang, W. Identification of differentially expressed host genes in Bombyx mori nucleopolyhedrovirus infected cells by using subtractive hybridization. Appl. Entomol. Zool. 2007, 42, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Kanthong, N.; Khemnu, N.; Sriurairatana, S.; Pattanakitsakul, S.-N.; Malaise, P.; Flegel, T.W. Mosquito cells accommodate balanced, persistent co-infections with a densovirus and Dengue virus. Dev. Comp. Immunol. 2008, 32, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Volkman, L.E.; Summers, M.D. Autographa californica nuclear polyhedrosis virus: Comparative infectivity of the occluded, alkali-liberated, and nonoccluded forms. J. Invertebr. Pathol. 1977, 30, 102–103. [Google Scholar] [CrossRef]
- De Mirand, J.R.; Cornman, R.S.; Evans, J.D.; Semberg, E.; Haddad, N.; Neumann, P.; Gauthier, L. Genome characterization, prevalence and distribution of a macula-like virus from Apis mellifera and Varroa destructor. Viruses 2015, 7, 3586–3602. [Google Scholar] [CrossRef] [PubMed]
- Parmentier, L.; Smagghe, G.; de Graaf, D.C.; Meeus, I. Varroa destructor Macula-like virus, Lake Sinai virus and other new RNA viruses in wild bumblebee hosts (Bombus pascuorum, Bombus lapidarius and Bombus pratorum). J. Invertebr. Pathol. 2016, 134, 6–11. [Google Scholar] [CrossRef] [PubMed]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsukui, K.; Yagisawa, C.; Fujimoto, S.; Ogawa, M.; Kokusho, R.; Nozawa, M.; Kawasaki, H.; Katsuma, S.; Iwanaga, M. Infectious Virions of Bombyx Mori Latent Virus Are Incorporated into Bombyx Mori Nucleopolyhedrovirus Occlusion Bodies. Viruses 2019, 11, 316. https://doi.org/10.3390/v11040316
Tsukui K, Yagisawa C, Fujimoto S, Ogawa M, Kokusho R, Nozawa M, Kawasaki H, Katsuma S, Iwanaga M. Infectious Virions of Bombyx Mori Latent Virus Are Incorporated into Bombyx Mori Nucleopolyhedrovirus Occlusion Bodies. Viruses. 2019; 11(4):316. https://doi.org/10.3390/v11040316
Chicago/Turabian StyleTsukui, Keita, Chihiro Yagisawa, Shota Fujimoto, Moe Ogawa, Ryuhei Kokusho, Mitsuyoshi Nozawa, Hideki Kawasaki, Susumu Katsuma, and Masashi Iwanaga. 2019. "Infectious Virions of Bombyx Mori Latent Virus Are Incorporated into Bombyx Mori Nucleopolyhedrovirus Occlusion Bodies" Viruses 11, no. 4: 316. https://doi.org/10.3390/v11040316
APA StyleTsukui, K., Yagisawa, C., Fujimoto, S., Ogawa, M., Kokusho, R., Nozawa, M., Kawasaki, H., Katsuma, S., & Iwanaga, M. (2019). Infectious Virions of Bombyx Mori Latent Virus Are Incorporated into Bombyx Mori Nucleopolyhedrovirus Occlusion Bodies. Viruses, 11(4), 316. https://doi.org/10.3390/v11040316