Terminase Large Subunit Provides a New Drug Target for Herpesvirus Treatment

 , ,
, ,
Abstract
1. Introduction
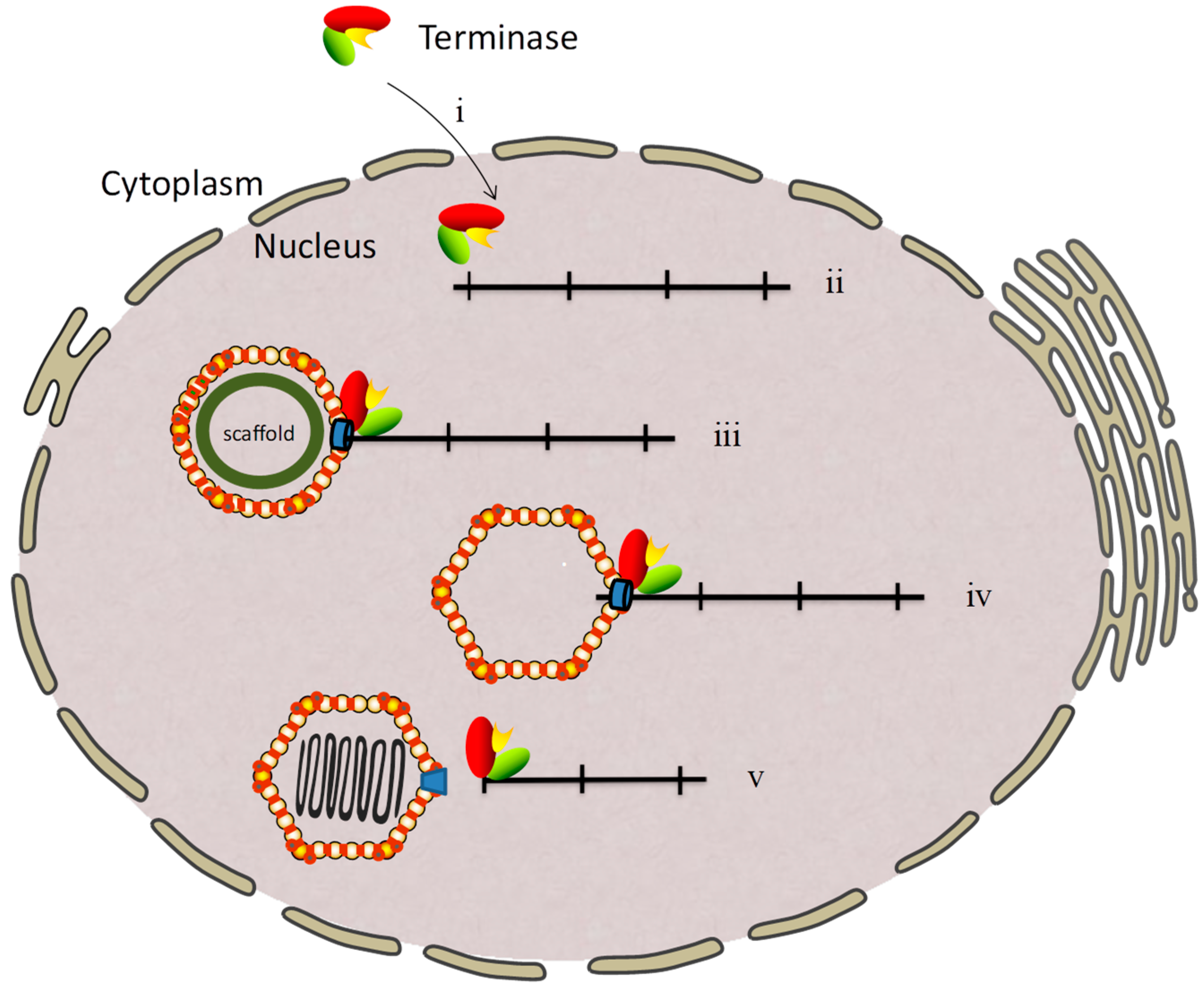
2. Genome Cleavage and Packaging of the Herpesvirus
3. Characteristics of the Terminase Large Subunit Gene
3.1. Terminase Large Subunit Coding by a Splicing Gene
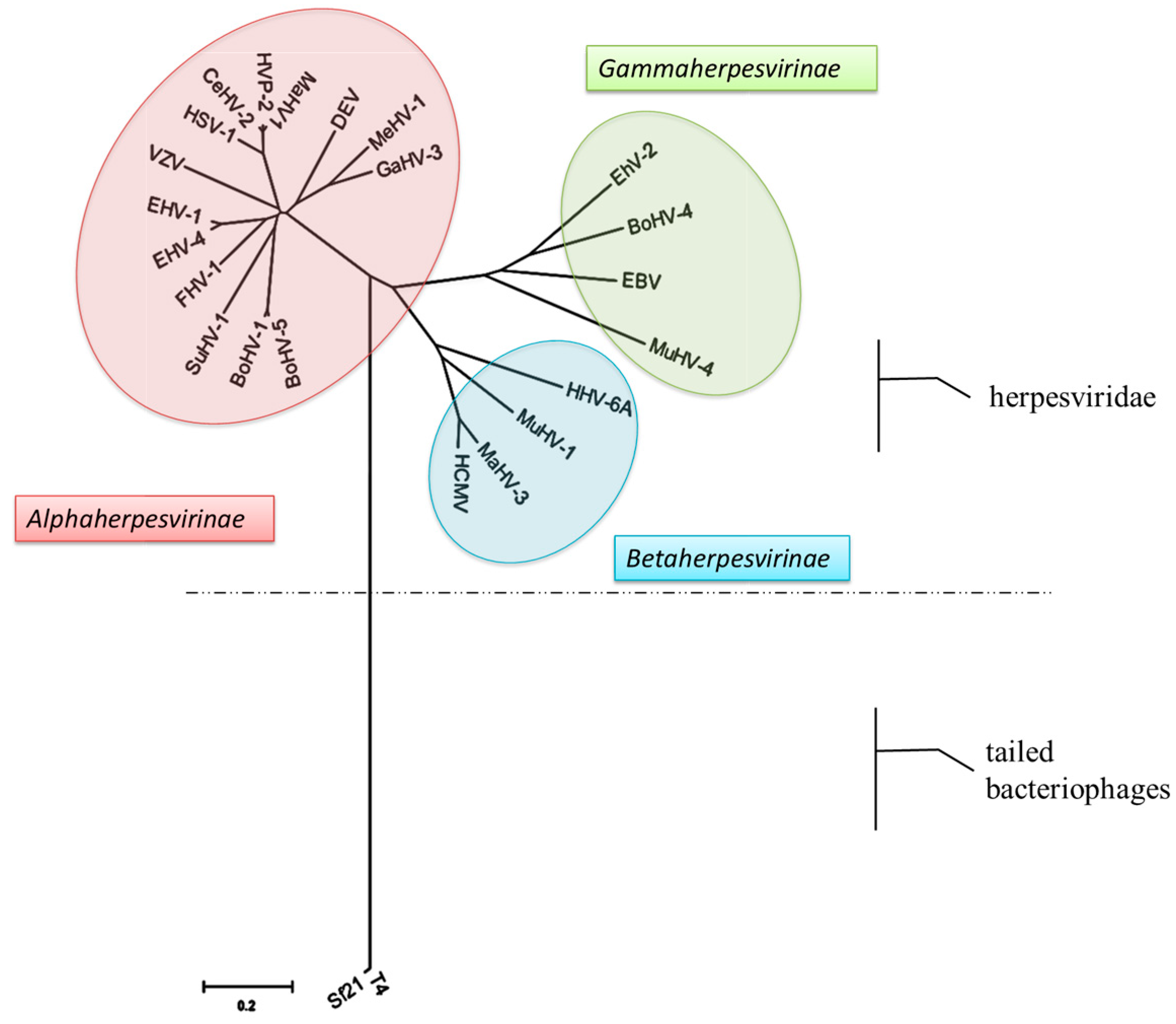
3.2. Evolutionary Analysis and Sequence Alignment of the Terminase Large Subunit
4. Structural and Functional Features of the Terminase Large Subunit
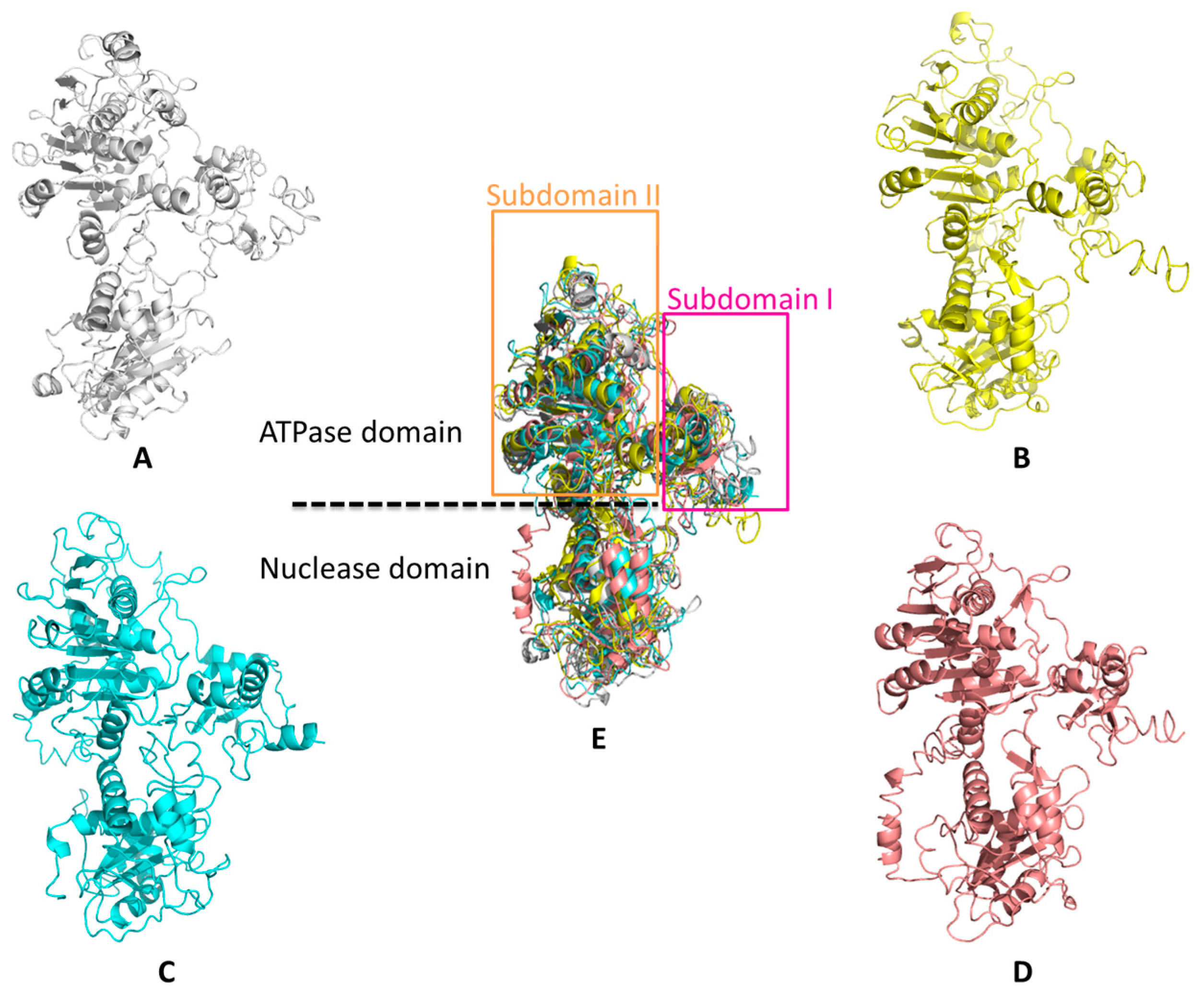
4.1. Overall Structure of the Terminase Large Subunit
4.2. ATPase Functions of the Terminase Large Subunit
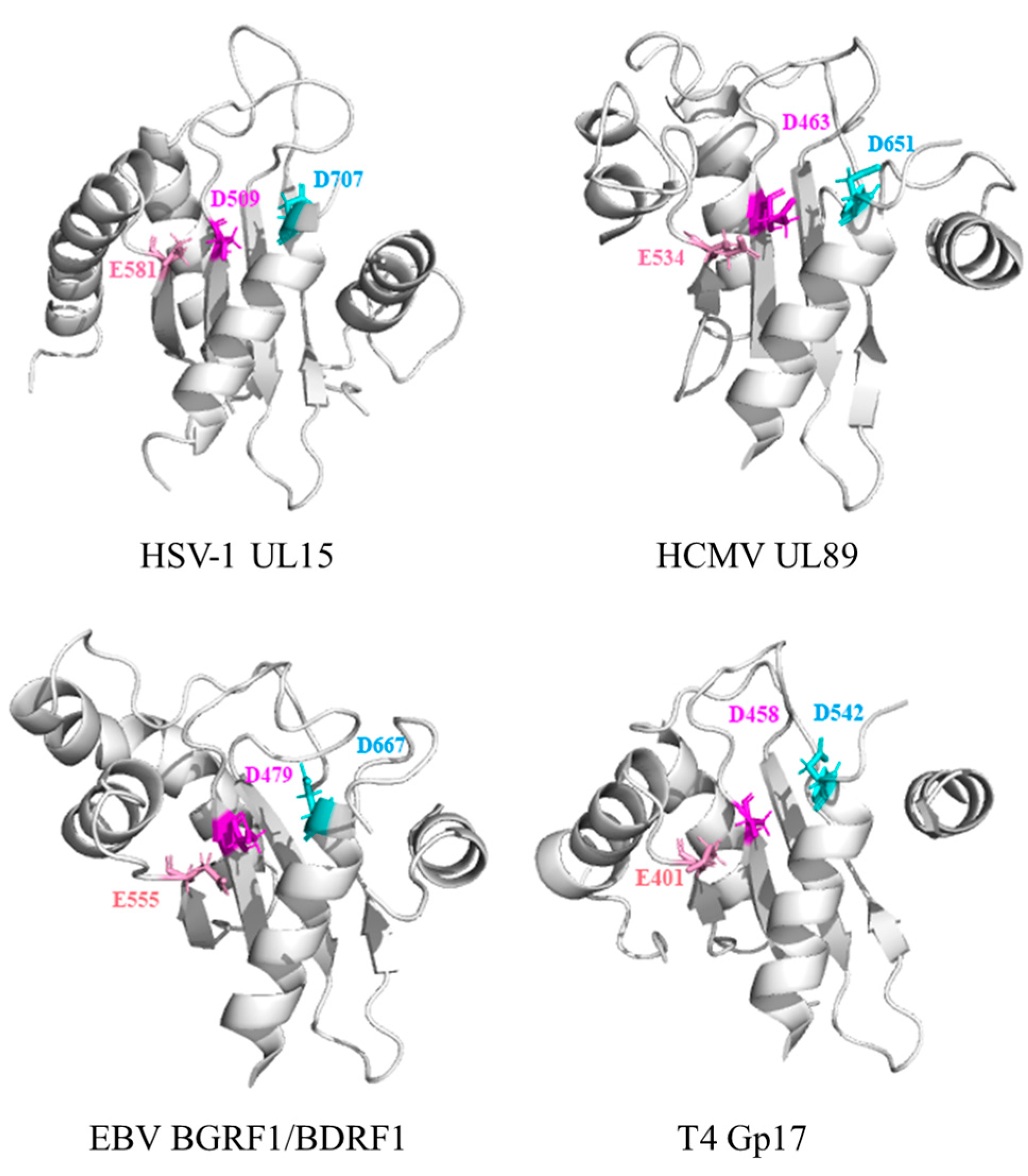
4.3. Nuclease Functions of the Terminase Large Subunit
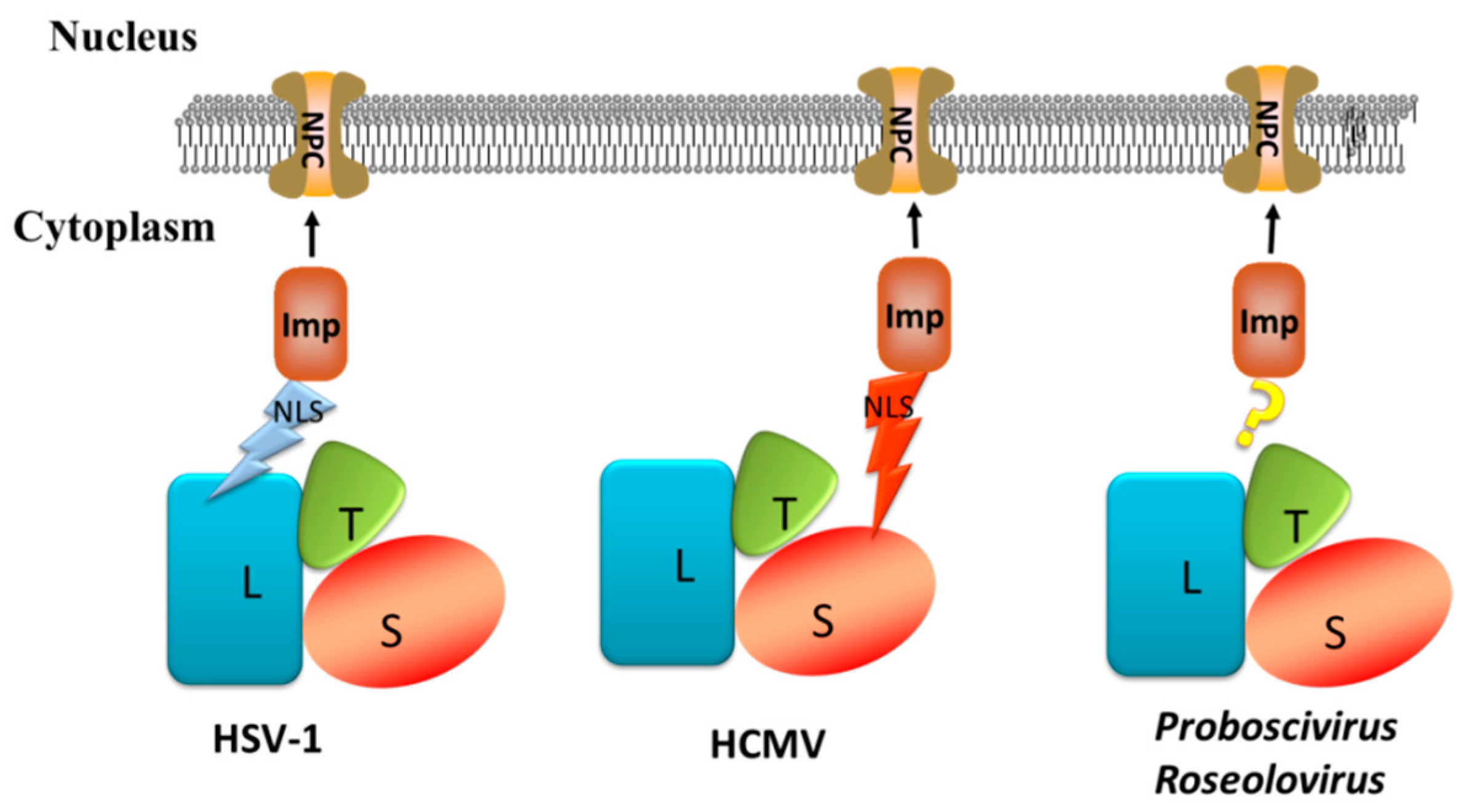
5. Three Models of the Terminase Complex Entering the Nucleus
5.1. Nuclear Import of Viral Protein
5.2. NLS of the Terminase Large Subunit Mediates HSV-1 Terminase Complex Entry into the Nucleus
5.3. NLS of the Terminase Small Subunit Mediate Terminase Complex Entry into the Nucleus
5.4. Holoenzyme Mediates Terminase Complex Entry into the Nucleus
6. Interaction between the Terminase Large Subunit and Other Viral Proteins
7. Antiviral Inhibitors Targeting the Terminase Large Subunit
7.1. Antiviral Drugs Targeting Herpesviruses
7.2. Small Molecular Inhibitors Targeting the HCMV Terminase Large Subunit
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- King, A.M.Q.; Lefkowitz, E.J.; Mushegian, A.R.; Adams, M.J.; Dutilh, B.E.; Gorbalenya, A.E.; Harrach, B.; Harrison, R.L.; Junglen, S.; Knowles, N.J.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2018). Arch. Virol. 2018, 162, 2505–2538. [Google Scholar] [CrossRef] [PubMed]
- Mcgeoch, D.J.; Rixon, F.J.; Davison, A.J. Topics in herpesvirus genomics and evolution. Virus Res. 2006, 117, 90–104. [Google Scholar] [CrossRef] [PubMed]
- Roizman, B.; Carmichael, L.E.; Deinhardt, F.; de-The, G.; Nahmias, A.J.; Plowright, W.; Rapp, F.; Sheldrick, P.; Takahashi, M.; Wolf, K. Herpesviridae. Definition, provisional nomenclature, and taxonomy. The Herpesvirus Study Group, the International Committee on Taxonomy of Viruses. Intervirology 1981, 16, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Pellett, P.E.; Roizman, B. The Family Herpesviridae: A Brief Introduction; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 2479–2499. [Google Scholar]
- Arvin, A.; Campadelli-Fiume, G.; Mocarski, E.; Moore, P.S.; Roizman, B.; Whitley, R.; Yamanishi, K. Comparative virion structures of human herpesviruses. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Cambridge University Press: Cambridge, UK, 2007; pp. 2–9. [Google Scholar]
- Zhou, Z.H.; Chen, D.H.; Jakana, J.; Rixon, F.J.; Chiu, W. Visualization of tegument-capsid interactions and DNA in intact herpes simplex virus type 1 virions. J. Virol. 1999, 73, 3210–3218. [Google Scholar] [PubMed]
- Roizman, B.; Furlong, D. The Replication of Herpesviruses; Springer: New York, NY, USA, 1974. [Google Scholar]
- Heldwein, E.E.; Lou, H.; Bender, F.C.; Cohen, G.H.; Eisenberg, R.J.; Harrison, S.C. Crystal structure of glycoprotein B from herpes simplex virus 1. Science 2006, 313, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.C.; Baines, J.D. Herpesviruses remodel host membranes for virus egress. Nat. Rev. Microbiol. 2011, 9, 382. [Google Scholar] [CrossRef] [PubMed]
- Liashkovich, I.; Hafezi, W.; Kühn, J.M.; Oberleithner, H.; Shahin, V. Nuclear delivery mechanism of herpes simplex virus type 1 genome. J. Mol. Recognit. 2011, 24, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Enquist, L.W.; Husak, P.J.; Banfield, B.W.; Smith, G.A. Infection and spread of alphaherpesviruses in the nervous system. Adv. Virus Res. 1998, 51, 237–347. [Google Scholar] [PubMed]
- Epstein, M.A. Observations on the mode of release of herpes virus from infected HeLa cells. J. Cell Biol. 1962, 12, 589–597. [Google Scholar] [CrossRef] [PubMed]
- White, C.A.; Stow, N.D.; Patel, A.H.; Hughes, M.; Preston, V.G. Herpes Simplex Virus Type 1 Portal Protein UL6 Interacts with the Putative Terminase Subunits UL15 and UL28. J. Virol. 2003, 77, 6351–6358. [Google Scholar] [CrossRef] [PubMed]
- Addison, C.; Rixon, F.J.; Preston, V.G. Herpes simplex virus type 1 UL28 gene product is important for the formation of mature capsids. J. Gen. Virol. 1990, 71 Pt 10, 2377–2384. [Google Scholar] [CrossRef]
- Alkobaisi, M.F.; Rixon, F.J.; Mcdougall, I.; Preston, V.G. The herpes simplex virus UL33 gene product is required for the assembly of full capsids. Virology 1991, 180, 380–388. [Google Scholar] [CrossRef]
- Lamberti, C.; Weller, S.K. The herpes simplex virus type 1 cleavage/packaging protein, UL32, is involved in efficient localization of capsids to replication compartments. J. Virol. 1998, 72, 2463–2473. [Google Scholar] [PubMed]
- Mcnab, A.R.; Desai, P.; Person, S.; Roof, L.L.; Thomsen, D.R.; Newcomb, W.W.; Brown, J.C.; Homa, F.L. The Product of the Herpes Simplex Virus Type 1 UL25 Gene Is Required for Encapsidation but Not for Cleavage of Replicated Viral DNA. J. Virol. 1998, 72, 1060–1070. [Google Scholar] [PubMed]
- Salmon, B.; Cunningham, C.; Davison, A.J.; Harris, W.J.; Baines, J.D. The Herpes Simplex Virus Type 1 UL17 Gene Encodes Virion Tegument Proteins That Are Required for Cleavage and Packaging of Viral DNA. J. Virol. 1998, 72, 3779–3788. [Google Scholar] [PubMed]
- Albright, B.S.; Kosinski, A.; Szczepaniak, R.; Cook, E.A.; Stow, N.D.; Conway, J.F.; Weller, S.K. The putative herpes simplex virus 1 chaperone protein UL32 modulates disulfide bond formation during infection. J. Virol. 2015, 89, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Beard, P.M.; Taus, N.S.; Baines, J.D. DNA cleavage and packaging proteins encoded by genes U(L)28, U(L)15, and U(L)33 of herpes simplex virus type 1 form a complex in infected cells. J. Virol. 2002, 76, 4785–4791. [Google Scholar] [CrossRef] [PubMed]
- Beilstein, F.; Higgs, M.R.; Stow, N.D. Mutational Analysis of the Herpes Simplex Virus Type 1 DNA Packaging Protein UL33. J. Virol. 2009, 83, 8938–8945. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wills, E.G.; Baines, J.D. A mutation in UL15 of herpes simplex virus 1 that reduces packaging of cleaved genomes. J. Virol. 2011, 85, 11972–11980. [Google Scholar] [CrossRef] [PubMed]
- Heming, J.D.; Huffman, J.B.; Jones, L.M.; Homa, F.L. Isolation and characterization of the herpes simplex virus 1 terminase complex. J. Virol. 2014, 88, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.R. The DNA-packaging nanomotor of tailed bacteriophages. Nat. Rev. Microbiol. 2011, 9, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.B.; Feiss, M. The Bacteriophage DNA Packaging Motor. Annu. Rev. Genet. 2008, 42, 647–681. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Zhao, Z.; Haak, J.; Wang, S.; Wu, D.; Meng, B.; Weitao, T. Common mechanisms of DNA translocation motors in bacteria and viruses using one-way revolution mechanism without rotation. Biotechnol. Adv. 2014, 32, 853–872. [Google Scholar] [CrossRef] [PubMed]
- McNulty, R.; Lokareddy, R.K.; Roy, A.; Yang, Y.; Lander, G.C.; Heck, A.J.R.; Johnson, J.E.; Cingolani, G. Architecture of the Complex Formed by Large and Small Terminase Subunits from Bacteriophage P22. J. Mol. Biol. 2015, 427, 3285–3299. [Google Scholar] [CrossRef] [PubMed]
- Heming, J.D.; Conway, J.F.; Homa, F.L. Herpesvirus Capsid Assembly and DNA Packaging. Adv. Anat. Embryol. Cell Biol. 2017, 223, 119–142. [Google Scholar] [PubMed]
- Gibson, W.; Roizman, B. Proteins specified by herpes simplex virus. 8. Characterization and composition of multiple capsid forms of subtypes 1 and 2. J. Virol. 1972, 10, 1044–1052. [Google Scholar] [PubMed]
- Newcomb, W.W.; Homa, F.L.; Thomsen, D.R.; Booy, F.P.; Trus, B.L.; Steven, A.C.; Spencer, J.V.; Brown, J.C. Assembly of the Herpes Simplex Virus Capsid: Characterization of Intermediates Observed During Cell-free Capsid Formation. J. Mol. Biol. 1996, 263, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Rixon, F.J.; Mcnab, D. Packaging-competent capsids of a herpes simplex virus temperature-sensitive mutant have properties similar to those of in vitro-assembled procapsids. J. Virol. 1999, 73, 5714–5721. [Google Scholar] [PubMed]
- Newcomb, W.W.; Trus, B.L.; Cheng, N.; Steven, A.C.; Sheaffer, A.K.; Tenney, D.J.; Weller, S.K.; Brown, J.C. Isolation of herpes simplex virus procapsids from cells infected with a protease-deficient mutant virus. J. Virol. 2000, 74, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Newcomb, W.; Homa, F.D.; Trus, B.; Cheng, N.; Steven, A.; Booy, F.; Brown, J. Assembly of the herpes simplex virus procapsid from purified components and identification of small complexes containing the major capsid and scaffolding proteins. J. Virol. 1999, 73, 4239–4250. [Google Scholar] [PubMed]
- Higgs, M.R.; Preston, V.G.; Stow, N.D. The UL15 protein of herpes simplex virus type 1 is necessary for the localization of the UL28 and UL33 proteins to viral DNA replication centres. J. Gen. Virol. 2008, 89, 1709–1715. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Homa, F.; Baines, J.D. Putative Terminase Subunits of Herpes Simplex Virus 1 Form a Complex in the Cytoplasm and Interact with Portal Protein in the Nucleus. J. Virol. 2007, 81, 6419–6433. [Google Scholar] [CrossRef] [PubMed]
- Adelman, K.; Salmon, B.; Baines, J.D. Herpes Simplex Virus DNA Packaging Sequences Adopt Novel Structures that are Specifically Recognized by a Component of the Cleavage and Packaging Machinery. Proc. Natl. Acad. Sci. USA 2001, 98, 3086–3091. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Dang, X.; Baines, J.D. A domain of Herpes simplex virus pUL33 required to release monomeric viral genomes from cleaved concatameric DNA. J. Virol. 2017, 9, 817–854. [Google Scholar]
- Yang, K.; Wills, E.; Baines, J.D. A Herpes Simplex Virus Scaffold Peptide That Binds the Portal Vertex Inhibits Early Steps in Viral Replication. J. Virol. 2013, 87, 6876–6887. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth, W.; Luella, S.; Baines, J.D. Herpes simplex virus 1 DNA packaging proteins encoded by UL6, UL15, UL17, UL28, and UL33 are located on the external surface of the viral capsid. J. Virol. 2006, 80, 10894–10899. [Google Scholar]
- Dasgupta, A.; Wilson, D.W. ATP depletion blocks herpes simplex virus DNA packaging and capsid maturation. J. Virol. 1999, 73, 2006–2015. [Google Scholar] [PubMed]
- Sheaffer, A.K.; Newcomb, W.W.; Gao, M.; Yu, D.; Weller, S.K.; Brown, J.C.; Tenney, D.J. Herpes simplex virus DNA cleavage and packaging proteins associate with the procapsid prior to its maturation. J. Virol. 2001, 75, 687–698. [Google Scholar] [CrossRef] [PubMed]
- Hodge, P.D.; Stow, N.D. Effects of mutations within the herpes simplex virus type 1 DNA encapsidation signal on packaging efficiency. J. Virol. 2001, 75, 8977–8986. [Google Scholar] [CrossRef] [PubMed]
- Mocarski, E.S.; Roizman, B. Structure and role of the herpes simplex virus DNA termini in inversion, circularization and generation of virion DNA. Cell 1982, 31, 89–97. [Google Scholar] [CrossRef]
- Beard, P.M.; Baines, J.D. The DNA cleavage and packaging protein encoded by the UL33 gene of herpes simplex virus 1 associates with capsids. Virology 2004, 324, 475–482. [Google Scholar] [CrossRef] [PubMed]
- Taus, N.S.; Baines, J.D. Herpes Simplex Virus 1 DNA Cleavage/Packaging: The U L 28 Gene Encodes a Minor Component of B Capsids. Virology 1998, 252, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Weller, S.K. Herpes simplex virus type 1 cleavage and packaging proteins UL15 and UL28 are associated with B but not C capsids during packaging. J. Virol. 1998, 72, 7428–7439. [Google Scholar] [PubMed]
- Baines, J.D.; Cunningham, C.; Nalwanga, D.; Davison, A. The U(L)15 gene of herpes simplex virus type 1 contains within its second exon a novel open reading frame that is translated in frame with the U(L)15 gene product. J. Virol. 1997, 71, 2666–2673. [Google Scholar] [PubMed]
- Przech, A.J.; Yu, D.; Weller, S.K. Point mutations in exon I of the herpes simplex virus putative terminase subunit, UL15, indicate that the most conserved residues are essential for cleavage and packaging. J. Virol. 2003, 77, 9613–9621. [Google Scholar] [CrossRef] [PubMed]
- Salmon, B.; Nalwanga, D.; Fan, Y.; Baines, J.D. Proteolytic cleavage of the amino terminus of the U(L)15 gene product of herpes simplex virus type 1 is coupled with maturation of viral DNA into unit-length genomes. J. Virol. 1999, 73, 8338–8348. [Google Scholar] [PubMed]
- Selvarajan Sigamani, S.; Zhao, H.; Kamau, Y.N.; Baines, J.D.; Tang, L. The Structure of the Herpes Simplex Virus DNA-Packaging Terminase pUL15 Nuclease Domain Suggests an Evolutionary Lineage among Eukaryotic and Prokaryotic Viruses. J. Virol. 2013, 87, 7140–7148. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Jenkins, H.T.; Chechik, M.; Blagova, E.V.; Lopatina, A.; Klimuk, E.; Minakhin, L.; Severinov, K.; Greive, S.J.; Antson, A.A. Viral genome packaging terminase cleaves DNA using the canonical RuvC-like two-metal catalysis mechanism. Nucleic Acids Res. 2017, 45, 3580–3590. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Lin, Z.; Lynn, A.Y.; Varnado, B.; Beutler, J.A.; Murelli, R.P.; Le Grice, S.F.J.; Tang, L. Two distinct modes of metal ion binding in the nuclease active site of a viral DNA-packaging terminase: Insight into the two-metal-ion catalytic mechanism. Nucleic Acids Res. 2015, 43, 11003–11016. [Google Scholar] [CrossRef] [PubMed]
- DelToro, D.; Ortiz, D.; Ordyan, M.; Sippy, J.; Oh, C.S.; Keller, N.; Feiss, M.; Catalano, C.E.; Smith, D.E. Walker-A Motif Acts to Coordinate ATP Hydrolysis with Motor Output in Viral DNA Packaging. J. Mol. Biol. 2016, 428, 2709–2729. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.S.; Rao, V.B. Functional analysis of the bacteriophage T4 DNA-packaging ATPase motor. J. Biol. Chem. 2006, 281, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Weller, S. Genetic analysis of the UL 15 gene locus for the putative terminase of herpes simplex virus type 1. Virology 1998, 243, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Yang, W. Nucleases: Diversity of structure, function and mechanism. Q. Rev. Biophys. 2011, 44, 1–93. [Google Scholar] [CrossRef] [PubMed]
- Nadal, M.; Mas, P.J.; Blanco, A.G.; Arnan, C.; Solà, M.; Hart, D.J.; Coll, M. Structure and inhibition of herpesvirus DNA packaging terminase nuclease domain. Proc. Natl. Acad. Sci. USA 2010, 107, 16078–16083. [Google Scholar] [CrossRef] [PubMed]
- Ghoshkumar, M.; Alam, T.I.; Draper, B.; Stack, J.D.; Rao, V.B. Regulation by interdomain communication of a headful packaging nuclease from bacteriophage T4. Nucleic Acids Res. 2011, 39, 2742–2755. [Google Scholar] [CrossRef] [PubMed]
- Yeo, A.; Feiss, M. Specific interaction of terminase, the DNA packaging enzyme of bacteriophage lambda, with the portal protein of the prohead. J. Mol. Biol. 1995, 245, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Fujisawa, H.T.M. Structural and functional domains of the large subunit of the bacteriophage t3 dna packaging enzyme—Importance of the c-terminal region in prohead binding. J. Mol. Biol. 1995, 245, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Couvreux, A.; Hantz, S.; Marquant, R.; Champier, G.; Alain, S.; Morellet, N.; Bouaziz, S. Insight into the structure of the pUL89 C-terminal domain of the human cytomegalovirus terminase complex. Proteins Struct. Funct. Bioinform. 2010, 78, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Cingolani, G. Structure of p22 headful packaging nuclease. J. Biol. Chem. 2012, 287, 28196–28205. [Google Scholar] [CrossRef] [PubMed]
- Smits, C.; Chechik, M.; Kovalevskiy, O.V.; Shevtsov, M.B.; Foster, A.W.; Alonso, J.C.; Antson, A.A. Structural basis for the nuclease activity of a bacteriophage large terminase. EMBO Rep. 2009, 10, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Nowotny, M.; Gaidamakov, S.A.; Ghirlando, R.; Cerritelli, S.M.; Crouch, R.J.; Yang, W. Structure of Human RNase H1 Complexed with an RNA/DNA Hybrid: Insight into HIV Reverse Transcription. Mol. Cell 2007, 28, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Kondabagil, K.; Draper, B.; Alam, T.I.; Bowman, V.D.; Zhang, Z.; Hegde, S.; Fokine, A.; Rossmann, M.G.; Rao, V.B. The structure of the phage T4 DNA packaging motor suggests a mechanism dependent on electrostatic forces. Cell 2008, 135, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Black, L.W.; Rao, V.B. Structure, assembly, and DNA packaging of the bacteriophage T4 head. Adv. Virus Res. 2012, 82, 119–153. [Google Scholar] [PubMed]
- Bottoms, C.A.; Smith, P.E.; Tanner, J.J. A structurally conserved water molecule in Rossmann dinucleotide-binding domains. Protein Sci. 2010, 11, 2125–2137. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Li, H.; Han, Z.; Shao, Y.; Wang, Y.; Kong, X. Identification of a spliced gene from duck enteritis virus encoding a protein homologous to UL15 of herpes simplex virus 1. Virol. J. 2011, 8, 156. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.E.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. EMBO J. 1982, 1, 945–951. [Google Scholar] [CrossRef] [PubMed]
- Draper, B.; Rao, V.B. An ATP hydrolysis sensor in the DNA packaging motor from bacteriophage T4 suggests an inchworm-type translocation mechanism. J. Mol. Biol. 2007, 369, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Leffers, G.; Rao, V.B. Biochemical characterization of an ATPase activity associated with the large packaging subunit gp17 from bacteriophage T4. J. Biol. Chem. 2000, 275, 37127–37136. [Google Scholar] [CrossRef] [PubMed]
- Baumann, R.G.; Black, L.W. Isolation and Characterization of T4 Bacteriophage gp17 Terminase, a Large Subunit Multimer with Enhanced ATPase Activity. J. Biol. Chem. 2003, 278, 4618–4627. [Google Scholar] [CrossRef] [PubMed]
- Franklin, J.L.; Haseltine, D.; Davenport, L.; Mosig, G. The largest (70 kDa) product of the bacteriophage T4 DNA terminase gene 17 binds to single-stranded DNA segments and digests them towards junctions with double-stranded DNA. J. Mol. Biol. 1998, 277, 541–557. [Google Scholar] [CrossRef] [PubMed]
- Champier, G.; Hantz, S.; Couvreux, A.; Stuppfler, S.; Mazeron, M.C.; Bouaziz, S.; Denis, F.; Alain, S. New functional domains of human cytomegalovirus pUL89 predicted by sequence analysis and three-dimensional modelling of the catalytic site DEXDc. Antivir. Ther. 2007, 12, 217–232. [Google Scholar] [PubMed]
- Hwang, J.S.; Bogner, E. ATPase activity of the terminase subunit pUL56 of human cytomegalovirus. J. Biol. Chem. 2002, 277, 6943–6948. [Google Scholar] [CrossRef] [PubMed]
- Scholz, B.; Rechter, S.; Drach, J.C.; Townsend, L.B.; Bogner, E. Identification of the ATP-binding site in the terminase subunit pUL56 of human cytomegalovirus. Nucleic Acids Res. 2003, 31, 1426–1433. [Google Scholar] [CrossRef] [PubMed]
- Kanamaru, S.; Kondabagil, K.; Rossmann, M.G.; Rao, V.B. The functional domains of bacteriophage t4 terminase. J. Biol. Chem. 2004, 279, 40795–40801. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Du, W.; Huang, L.; Wei, Y.; Wu, H.; Feng, L.; Liu, C. The Pseudorabies Virus DNA Polymerase Accessory Subunit UL42 Directs Nuclear Transport of the Holoenzyme. Front. Microbiol. 2016, 7, 124. [Google Scholar] [CrossRef] [PubMed]
- Nakada, R.; Hirano, H.; Matsuura, Y. Structure of importin-α bound to a non-classical nuclear localization signal of the influenza A virus nucleoprotein. Sci. Rep. 2015, 5, 15055. [Google Scholar] [CrossRef] [PubMed]
- Bennett, S.M.; Zhao, L.; Bosard, C.; Imperiale, M.J. Role of a nuclear localization signal on the minor capsid Proteins VP2 and VP3 in BKPyV nuclear entry. Virology 2015, 474, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Webel, R.; Solbak, S.M.Ø.; Held, C.; Milbradt, J.; Groß, A.; Eichler, J.; Wittenberg, T.; Jardin, C.; Sticht, H.; Fossen, T. Nuclear import of isoforms of the cytomegalovirus kinase pUL97 is mediated by differential activity of NLS1 and NLS2 both acting through classical importin-α binding. J. Gen. Virol. 2012, 93, 1756–1768. [Google Scholar] [CrossRef] [PubMed]
- Marfori, M.; Mynott, A.; Ellis, J.J.; Mehdi, A.M.; Saunders, N.F.W.; Curmi, P.M.; Forwood, J.K.; Bodén, M.; Kobe, B. Molecular basis for specificity of nuclear import and prediction of nuclear localization. Biochim. Biophys. Acta 2011, 1813, 1562–1577. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, G.; Petosa, C.; Weis, K.; Muller, C.W. Structure of importin-beta bound to the IBB domain of importin-alpha. Nature 1999, 399, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, G.; Bednenko, J.; Gillespie, M.T.; Gerace, L. Molecular basis for the recognition of a nonclassical nuclear localization signal by importin beta. Mol. Cell 2002, 10, 1345–1353. [Google Scholar] [CrossRef]
- Pumroy, R.A.; Cingolani, G. Diversification of importin-α isoforms in cellular trafficking and disease states. Biochem. J. 2015, 466, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Lott, K.; Bhardwaj, A.; Mitrousis, G.; Pante, N.; Cingolani, G. The importin beta binding domain modulates the avidity of importin beta for the nuclear pore complex. J. Biol. Chem. 2010, 285, 13769–13780. [Google Scholar] [CrossRef] [PubMed]
- Lott, K.; Cingolani, G. The importin beta binding domain as a master regulator of nucleocytoplasmic transport. Biochim. Biophys. Acta 2011, 1813, 1578–1592. [Google Scholar] [CrossRef] [PubMed]
- Fontes, M.R.; Teh, T.; Kobe, B. Structural basis of recognition of monopartite and bipartite nuclear localization sequences by mammalian importin-alpha. J. Mol. Biol. 2000, 297, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Conti, E.; Uy, M.; Leighton, L.; Blobel, G.; Kuriyan, J. Crystallographic analysis of the recognition of a nuclear localization signal by the nuclear import factor karyopherin alpha. Cell 1998, 94, 193–204. [Google Scholar] [CrossRef]
- Koslowski, K.M.; Shaver, P.R.; Ii, J.T.C.; Wilson, T.; Yamanaka, G.; Sheaffer, A.K.; Tenney, D.J.; Pederson, N.E. Physical and Functional Interactions between the Herpes Simplex Virus UL15 and UL28 DNA Cleavage and Packaging Proteins. J. Virol. 1999, 73, 1704–1707. [Google Scholar] [PubMed]
- Sankhala, R.S.; Lokareddy, R.K.; Cingolani, G. Divergent Evolution of Nuclear Localization Signal Sequences in Herpesvirus Terminase Subunits. J. Biol. Chem. 2016, 291, 11420–11433. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.M.; Yang, V.W. Two Potent Nuclear Localization Signals in the Gut-enriched Krüppel-like Factor Define a Subfamily of Closely Related Krüppel Proteins. J. Biol. Chem. 1997, 272, 18504–18507. [Google Scholar] [CrossRef] [PubMed]
- Giesen, K.; Radsak, K.; Bogner, E. The potential terminase subunit of human cytomegalovirus, pUL56, is translocated into the nucleus by its own nuclear localization signal and interacts with importin alpha. J. Gen. Virol. 2000, 81, 2231–2244. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.B.; Zhu, Y.; McVoy, M.A.; Parris, D.S. Changes in subcellular localization reveal interactions between human cytomegalovirus terminase subunits. Virol. J. 2012, 9, 315. [Google Scholar] [CrossRef] [PubMed]
- Neuber, S.; Wagner, K.; Goldner, T.; Lischka, P.; Steinbrueck, L.; Messerle, M.; Borst, E.M. Mutual Interplay between the Human Cytomegalovirus Terminase Subunits pUL51, pUL56, and pUL89 Promotes Terminase Complex Formation. J. Virol. 2017, 91, e2316–e2384. [Google Scholar] [CrossRef] [PubMed]
- Borst, E.M.; Kleine-Albers, J.; Gabaev, I.; Babic, M.; Wagner, K.; Binz, A.; Degenhardt, I.; Kalesse, M.; Jonjic, S.; Bauerfeind, R. The human cytomegalovirus UL51 protein is essential for viral genome cleavage-packaging and interacts with the terminase subunits pUL56 and pUL89. J. Virol. 2013, 87, 1720–1732. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.G.; Yang, K.; Baines, J.D.; Homa, F.L. Linker Insertion Mutations in the Herpes Simplex Virus Type 1 UL28 Gene: Effects on UL28 Interaction with UL15 and UL33 and Identification of a Second-Site Mutation in the UL15 Gene That Suppresses a Lethal UL28 Mutation. J. Virol. 2006, 80, 12312–12323. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Baines, J.D. The putative terminase subunit of herpes simplex virus 1 encoded by UL28 is necessary and sufficient to mediate interaction between pUL15 and pUL33. J. Virol. 2006, 80, 5733–5739. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Poon, A.P.; Roizman, B.; Baines, J.D. Temperature-Sensitive Mutations in the Putative Herpes Simplex Virus Type 1 Terminase Subunits pUL15 and pUL33 Preclude Viral DNA Cleavage/Packaging and Interaction with pUL28 at the Nonpermissive Temperature. J. Virol. 2008, 82, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Lokareddy, R.K.; Sankhala, R.S.; Roy, A.; Afonine, P.V.; Motwani, T.; Teschke, C.M.; Parent, K.N.; Cingolani, G. Portal protein functions akin to a DNA-sensor that couples genome-packaging to icosahedral capsid maturation. Nat. Commun. 2017, 8, 14310. [Google Scholar] [CrossRef] [PubMed]
- Mcelwee, M.; Vijayakrishnan, S.; Rixon, F.; Bhella, D. Structure of the herpes simplex virus portal-vertex. PLoS Biol. 2018, 16, e2006191. [Google Scholar] [CrossRef] [PubMed]
- Liu, L. Fields Virology, 6th Edition. Clin. Infect. Dis. 2014, 59, 613. [Google Scholar] [CrossRef]
- Fujiwara, S.; Matsuda, G.; Imadome, K. Humanized mouse models of epstein-barr virus infection and associated diseases. Pathogens 2013, 2, 153–176. [Google Scholar] [CrossRef] [PubMed]
- Beese, L.S.; Steitz, T.A. Structural basis for the 3’-5’ exonuclease activity of Escherichia coli DNA polymerase I: A two metal ion mechanism. EMBO J. 1991, 10, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Lurain, N.S.; Chou, S. Antiviral drug resistance of human cytomegalovirus. Clin. Microbiol. Rev. 2010, 23, 689–712. [Google Scholar] [CrossRef] [PubMed]
- Andrei, G.; Clercq, E.D.; Snoeck, R. Drug Targets in Cytomegalovirus Infection. Infect. Disord. Drug Targets 2009, 9, 201–222. [Google Scholar] [CrossRef] [PubMed]
- Biron, K.K. Antiviral drugs for cytomegalovirus diseases. Antivir. Res. 2006, 71, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.; Smith, J.L.; Chen, M.S. Effect of incorporation of cidofovir into DNA by human cytomegalovirus DNA polymerase on DNA elongation. Antimicrob. Agents Chemother. 1997, 41, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Drew, W.L. Cytomegalovirus resistance testing: Pitfalls and problems for the clinician. Clin. Infect. Dis. 2010, 50, 733–736. [Google Scholar] [CrossRef] [PubMed]
- Chou, S. Approach to drug-resistant cytomegalovirus in transplant recipients. Curr. Opin. Infect. Dis. 2015, 28, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Chou, S. Antiviral drug resistance in human cytomegalovirus. Transpl. Infect. Dis. 2010, 1, 105–114. [Google Scholar] [CrossRef]
- Lea, A.P.; Bryson, H.M. Cidofovir. Drugs 1996, 52, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, A.J.; Bryson, H.M. Foscarnet. Drugs 1994, 48, 199–226. [Google Scholar] [CrossRef] [PubMed]
- Komazin, G.; Townsend, L.B.; Drach, J.C. Role of a Mutation in Human Cytomegalovirus Gene UL104 in Resistance to Benzimidazole Ribonucleosides. J. Virol. 2004, 78, 710–715. [Google Scholar] [CrossRef] [PubMed]
- Zou, R.; Drach, J.C.; Townsend, L.B. Design, synthesis, and antiviral evaluation of 2-substituted 4,5-dichloro- and 4,6-dichloro-1-beta-D-ribofuranosylbenzimidazoles as potential agents for human cytomegalovirus infections. J. Med. Chem. 2010, 28, 802–810. [Google Scholar]
- Scheffczik, H.; Savva, C.G.W.; Holzenburg, A.; Kolesnikova, L.; Bogner, E. The terminase subunits pUL56 and pUL89 of human cytomegalovirus are DNA-metabolizing proteins with toroidal structure. Nucleic Acids Res. 2002, 30, 1695–1703. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.R.; Harvey, R.J.; Stanat, S.C.; Hemphill, M.L.; Miller, T.; Drach, J.C.; Townsend, L.B.; Biron, K.K. Inhibition of human cytomegalovirus DNA maturation by a benzimidazole ribonucleoside is mediated through the UL89 gene product. J. Virol. 1998, 72, 717–725. [Google Scholar] [PubMed]
- Reefschlaeger, J.; Bender, W.; Hallenberger, S.; Weber, O.; Eckenberg, P.; Goldmann, S.; Haerter, M.; Buerger, I.; Trappe, J.; Herrington, J.A.; et al. Novel non-nucleoside inhibitors of cytomegaloviruses (BAY 38-4766): In vitro and in vivo antiviral activity and mechanism of action. J. Antimicrob. Chemother. 2001, 48, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Kankanala, J.; Wang, Y.; Geraghty, R.J.; Wang, Z. Hydroxypyridonecarboxylic Acids as Inhibitors of Human Cytomegalovirus pUL89 Endonuclease. Chemmedchem 2018, 13, 1658–1663. [Google Scholar] [CrossRef] [PubMed]
- Underwood, M.R.; Ferris, R.G.; Selleseth, D.W.; Davis, M.G.; Drach, J.C.; Townsend, L.B.; Biron, K.K.; Boyd, F.L. Mechanism of action of the ribopyranoside benzimidazole GW275175X against human cytomegalovirus. Antimicrob. Agents Chemother. 2004, 48, 1647–1651. [Google Scholar] [CrossRef] [PubMed]
- Marty, F.M.; Ljungman, P.; Chemaly, R.F.; Maertens, J.; Dadwal, S.S.; Duarte, R.F.; Haider, S.; Ullmann, A.J.; Katayama, Y.; Brown, J. Letermovir Prophylaxis for Cytomegalovirus in Hematopoietic-Cell Transplantation. N. Engl. J. Med. 2017, 377, 2433–2444. [Google Scholar] [CrossRef] [PubMed]
- Frange, P.; Leruez-Ville, M. Maribavir, brincidofovir and letermovir: Efficacy and safety of new antiviral drugs for treating cytomegalovirus infections. Méd. Mal. Infect. 2018, 48, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Frietsch, J.J.; Michel, D.; Stamminger, T.; Hunstig, F.; Birndt, S.; Schnetzke, U.; Scholl, S.; Hochhaus, A.; Hilgendorf, I. In Vivo Emergence of UL56 C325Y Cytomegalovirus Resistance to Letermovir in a Patient with Acute Myeloid Leukemia after Hematopoietic Cell Transplantation. Mediterr. J. Hematol Infect. Dis. 2019, 11, e2019001. [Google Scholar] [CrossRef] [PubMed]
- Rao, V.B.; Feiss, M. Mechanisms of DNA Packaging by Large Double-Stranded DNA Viruses. Annu. Rev. Virol. 2015, 2, 351–378. [Google Scholar] [CrossRef] [PubMed]
- Bataille, D.; Epstein, A.L. Herpes simplex virus type 1 replication and recombination. Biochimie 1995, 77, 787–795. [Google Scholar] [CrossRef]
- Oliveira, L.; Cuervo, A.P. Direct interaction of the bacteriophage SPP1 packaging ATPase with the portal protein. J. Biol. Chem. 2010, 285, 7366–7373. [Google Scholar] [CrossRef] [PubMed]
- Dhar, A.; Feiss, M. Bacteriophage lambda terminase: Alterations of the high-affinity ATPase affect viral DNA packaging. J. Mol. Biol. 2005, 347, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Hilbert, B.J.; Hayes, J.A.; Stone, N.P.; Xu, R.G.; Kelch, B.A. The large terminase DNA packaging motor grips DNA with its ATPase domain for cleavage by the flexible nuclease domain. Nucleic Acids Res. 2017, 45, 3591–3605. [Google Scholar] [CrossRef] [PubMed]
- Ligat, G.; Cazal, R.; Hantz, S.; Alain, S. The human cytomegalovirus terminase complex as an antiviral target: A close-up view. FEMS Microbiol. Rev. 2018, 42, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Gentry, B.G.; Bogner, E.; Drach, J.C. Targeting the terminase: An important step forward in the treatment and prophylaxis of human cytomegalovirus infections. Antivir. Res. 2019, 161, 116–124. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subfamily | Virus Name | Abbreviation | GenBank Accession Number | Coding Region | Exon/Intron | Number of Amino Acids |
|---|---|---|---|---|---|---|
| Aphaherpesvirus | Herpes simplex virus 1 | HSV-1 | AER38023.1 | UL15 | 2/2 | 735 |
| Varicella-zoster virus | VZV | NP_040165.1 | ORF42 | 2/2 | 747 | |
| Macacine alphaherpesvirus 1 | MaHV1 | ARS02909.1 | UL15 | 2/2 | 738 | |
| Cercopithecine alphaherpesvirus 2 | CeHV-2 | YP_164457.1 | UL15 | 2/2 | 735 | |
| Papiine alphaherpesvirus 2 | HVP-2 | AHM96136.1 | UL15 | 2/2 | 735 | |
| Felid alphaherpesvirus 1 | FHV-1 | YP_003331564.1 | UL15 | 2/2 | 734 | |
| Suid alphaherpesvirus 1 | SuHV-1 | YP_068358.1 | UL15 | 2/2 | 735 | |
| Bovine alphaherpesvirus 5 | BoHV-5 | YP_003662508.1 | UL15 | 2/2 | 737 | |
| Bovine alphaherpesvirus 1 | BoHV-1 | APW77369.1 | UL15 | 2/2 | 735 | |
| Equid alphaherpesvirus 4 | EHV-4 | NP_045262.1 | ORF44 | 2/2 | 734 | |
| Equid alphaherpesvirus 1 | EHV-1 | BAM75895.1 | ORF44 | 2/2 | 734 | |
| Meleagrid alphaherpesvirus 1 | MeHV-1 | NP_073308.1 | HVT022 | 2/2 | 738 | |
| Duck enteritis virus | DEV | YP_003084405.1 | UL15 | 2/2 | 739 | |
| Betaherpesvirinae | Human cytomegalovirus | HCMV | YP_081537.1 | UL89 | 2/4 | 674 |
| Murid betaherpesvirus 1 | MuHV-1 | CCE56594.1 | M89 | 2/5 | 678 | |
| Human betaherpesvirus 6A | HHV-6A | APO38446.1 | U60 | 2/4 | 666 | |
| Macacine betaherpesvirus 3 | MaHV-3 | AAT07420.1 | grh124 | 2/5 | 671 | |
| Gammaherpesvirinae | Epstein–Barr virus | EBV | YP_401690.1 | BGRF1/BDRF1 | 2/4 | 690 |
| Bovine gammaherpesvirus 4 | BoHV-4 | AEL29773.1 | ORF29 | 2/4 | 682 | |
| Murid gammaherpesvirus 4 | MuHV-4 | AAF19294.1 | 29 | 2/4 | 679 | |
| Equid gammaherpesvirus 2 | EhV-2 | NP_042630.2 | ORF29 | 2/4 | 686 |
| Active Sites of Catalytic Domain | HSV-1 pUL15 | HCMV pUL89 | EBV BGRF1/BDRF1 | T4 Gp17 |
|---|---|---|---|---|
| Asp | 509 | 463 | 479 | 401 |
| Glu | 581 | 534 | 555 | 458 |
| Asp | 707 | 651 | 667 | 542 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Yang, Q.; Wang, M.; Jia, R.; Chen, S.; Zhu, D.; Liu, M.; Wu, Y.; Zhao, X.; Zhang, S.; et al. Terminase Large Subunit Provides a New Drug Target for Herpesvirus Treatment. Viruses 2019, 11, 219. https://doi.org/10.3390/v11030219
Yang L, Yang Q, Wang M, Jia R, Chen S, Zhu D, Liu M, Wu Y, Zhao X, Zhang S, et al. Terminase Large Subunit Provides a New Drug Target for Herpesvirus Treatment. Viruses. 2019; 11(3):219. https://doi.org/10.3390/v11030219
Chicago/Turabian StyleYang, Linlin, Qiao Yang, Mingshu Wang, Renyong Jia, Shun Chen, Dekang Zhu, Mafeng Liu, Ying Wu, Xinxin Zhao, Shaqiu Zhang, and et al. 2019. "Terminase Large Subunit Provides a New Drug Target for Herpesvirus Treatment" Viruses 11, no. 3: 219. https://doi.org/10.3390/v11030219
APA StyleYang, L., Yang, Q., Wang, M., Jia, R., Chen, S., Zhu, D., Liu, M., Wu, Y., Zhao, X., Zhang, S., Liu, Y., Yu, Y., Zhang, L., Chen, X., & Cheng, A. (2019). Terminase Large Subunit Provides a New Drug Target for Herpesvirus Treatment. Viruses, 11(3), 219. https://doi.org/10.3390/v11030219