Potent HIV-1-Specific CD8 T Cell Responses Induced in Mice after Priming with a Multiepitopic DNA-TMEP and Boosting with the HIV Vaccine MVA-B

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Synthetic TMEP-B Gene
2.2. Frequencies of TMEP-B Epitopes in the General HIV-1-Infected Population and in Long Term Non-Progressors (LTNPs)
2.3. Cells and Viruses
2.4. DNA Vectors
2.5. Transfection Assay and Expression of TMEP-B Protein by Western Blot Analysis
2.6. Ethics Statement
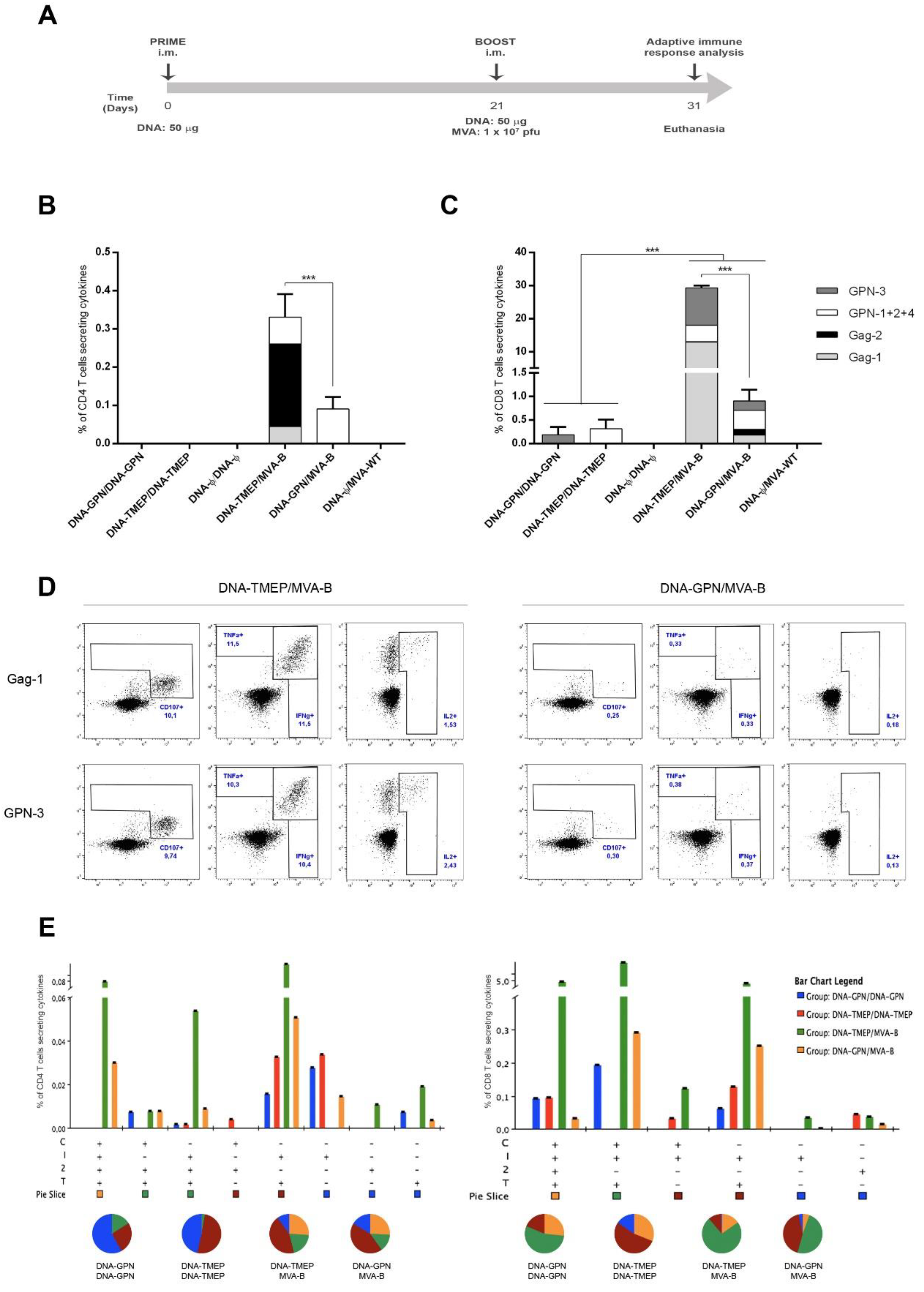
2.7. Mouse Immunization Schedule
2.8. Peptides
2.9. Intracellular Cytokine Staining (ICS) Assay
2.10. Antibody Measurement by Enzyme-Linked Immunosorbent Assay (ELISA)
2.11. Data Analysis and Statistics
3. Results
3.1. Design of the T Cell Multiepitopic Peptide TMEP-B Immunogen
3.2. Expression of TMEP-B Protein in Human Cells
3.3. Cellular Immune Profile Induced by the DNA-TMEP Immunogen
3.3.1. Magnitude, Breadth, and Functional Profile of the T Cell Responses Induced by DNA-TMEP in Homologous or Heterologous Prime/Boost Immunization Regimen with MVA-B
3.3.2. Characterization of HIV-1-Specific Tfh Cell Response Induced by DNA-TMEP in Homologous or Heterologous Prime/Boost Immunization Regimen
3.3.3. Effect of DNA Prime on the Env-Specific T Cell Immune Responses in Animals Receiving MVA-B Boost
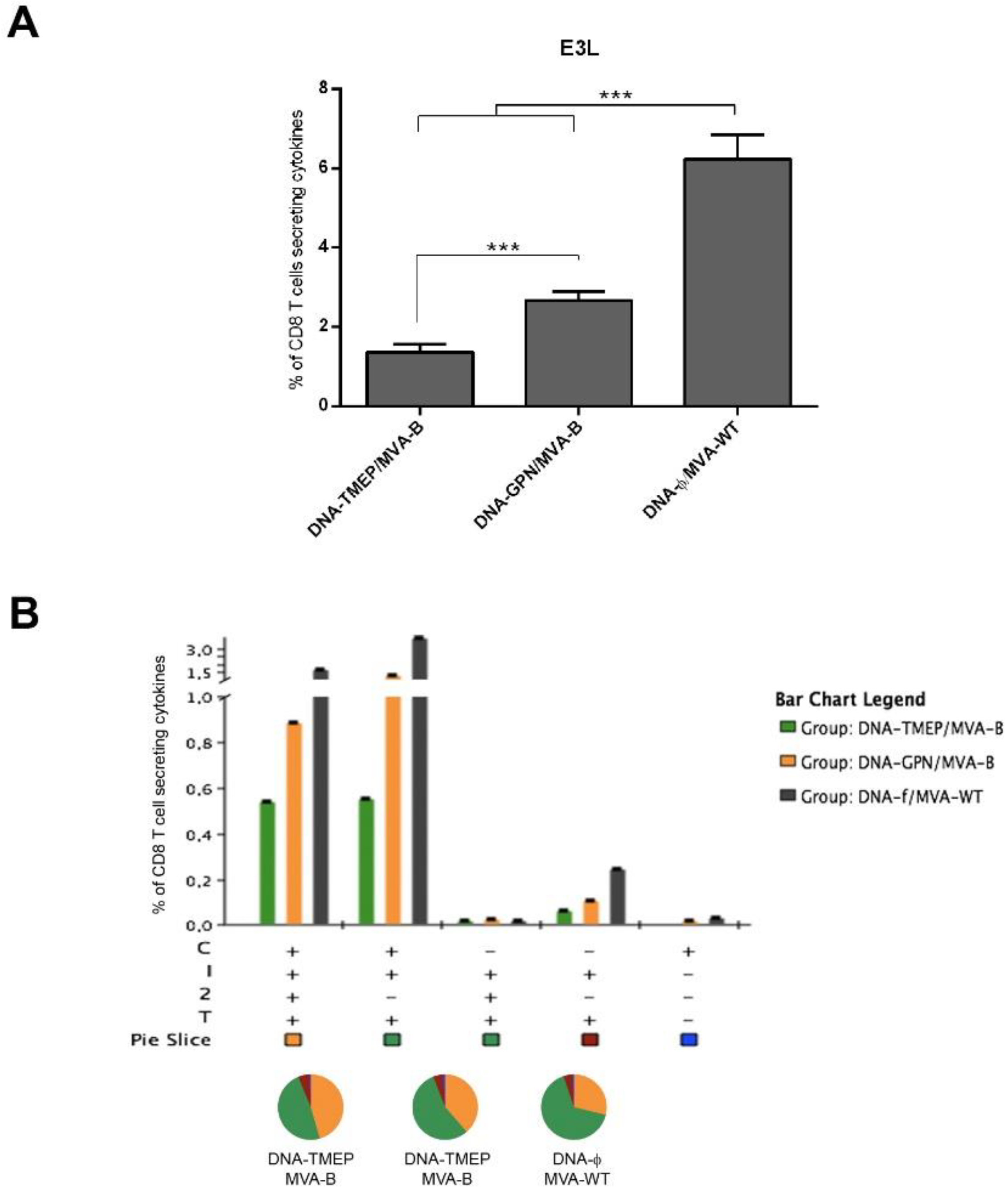
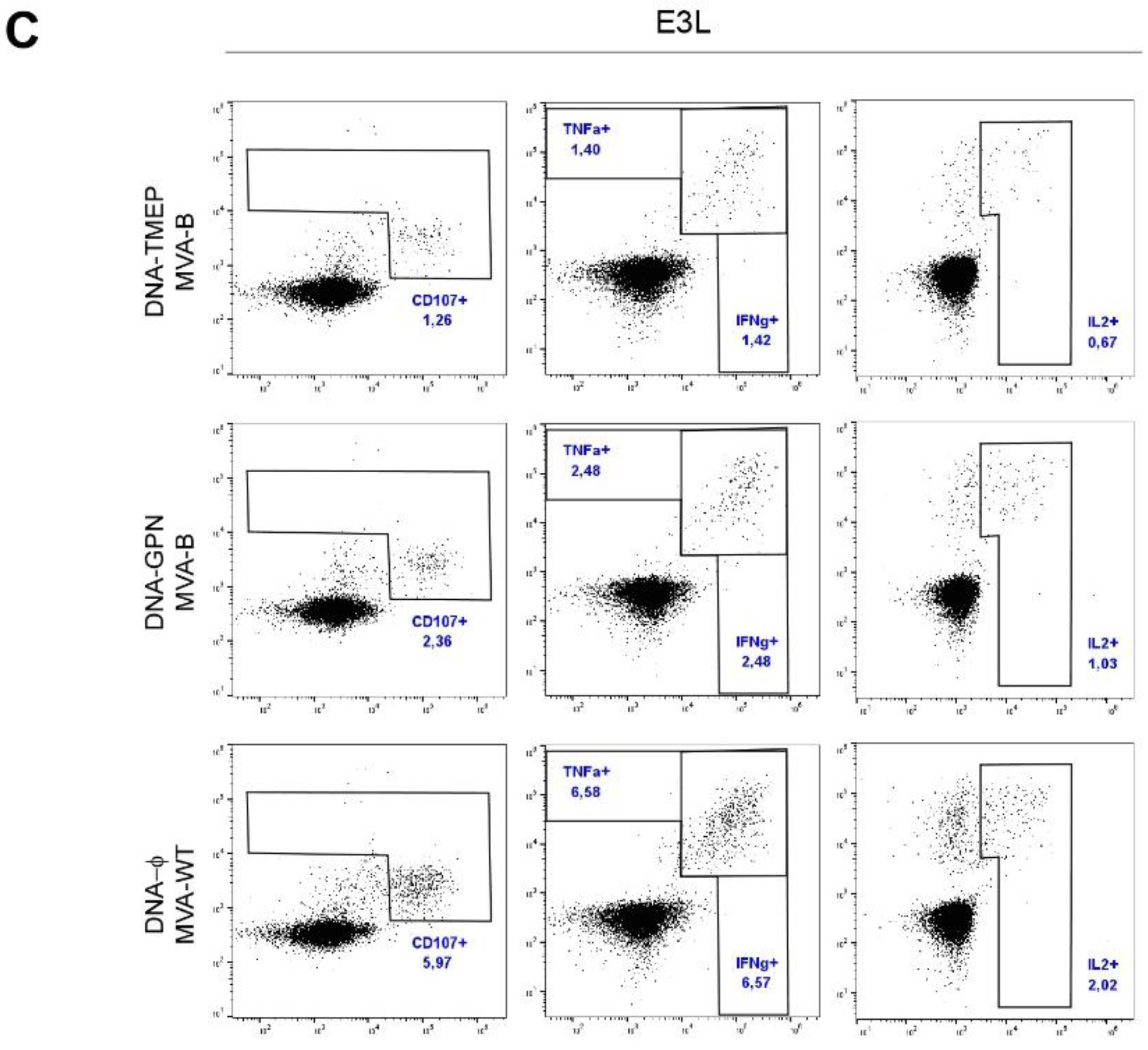
3.3.4. T Cell Immune Response against VACV E3 Peptide Induced by DNA/MVA Heterologous Combination
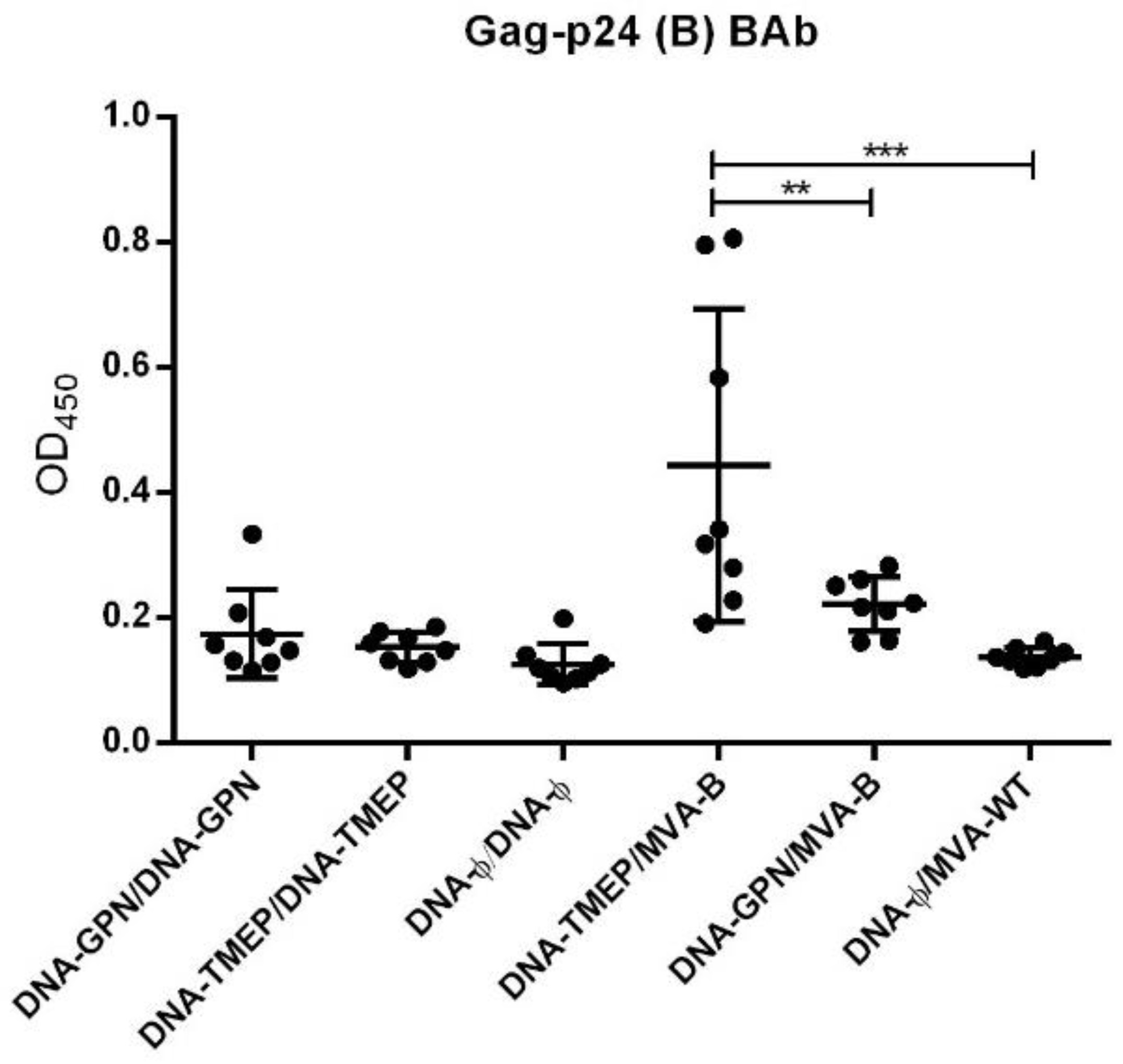
3.4. Humoral Immune Profile against HIV-1 Gag-P24
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Garcia, F.; Bernaldo de Quiros, J.C.; Gomez, C.E.; Perdiguero, B.; Najera, J.L.; Jimenez, V.; Garcia-Arriaza, J.; Guardo, A.C.; Perez, I.; Diaz-Brito, V.; et al. Safety and immunogenicity of a modified pox vector-based HIV/AIDS vaccine candidate expressing Env, Gag, Pol and Nef proteins of HIV-1 subtype B (MVA-B) in healthy HIV-1-uninfected volunteers: A phase I clinical trial (RISVAC02). Vaccine 2011, 29, 8309–8316. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.E.; Najera, J.L.; Perdiguero, B.; Garcia-Arriaza, J.; Sorzano, C.O.; Jimenez, V.; Gonzalez-Sanz, R.; Jimenez, J.L.; Munoz-Fernandez, M.A.; Lopez Bernaldo de Quiros, J.C.; et al. The HIV/AIDS vaccine candidate MVA-B administered as a single immunogen in humans triggers robust, polyfunctional, and selective effector memory T cell responses to HIV-1 antigens. J. Virol. 2011, 85, 11468–11478. [Google Scholar] [CrossRef] [PubMed]
- Guardo, A.C.; Gomez, C.E.; Diaz-Brito, V.; Pich, J.; Arnaiz, J.A.; Perdiguero, B.; Garcia-Arriaza, J.; Gonzalez, N.; Sorzano, C.O.S.; Jimenez, L.; et al. Safety and vaccine-induced HIV-1 immune responses in healthy volunteers following a late MVA-B boost 4 years after the last immunization. PLoS ONE 2017, 12, e0186602. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.E.; Perdiguero, B.; Garcia-Arriaza, J.; Cepeda, V.; Sanchez-Sorzano, C.O.; Mothe, B.; Jimenez, J.L.; Munoz-Fernandez, M.A.; Gatell, J.M.; Lopez Bernaldo de Quiros, J.C.; et al. A phase I randomized therapeutic MVA-B vaccination improves the magnitude and quality of the T cell immune responses in HIV-1-infected subjects on HAART. PLoS ONE 2015, 10, e0141456. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Climent, N.; Plana, M.; Rosas, M.; Jimenez, J.L.; Munoz-Fernandez, M.A.; Puertas, M.C.; Carrillo, J.; Gonzalez, N.; Leon, A.; et al. Safety and immunogenicity of a modified vaccinia Ankara-based HIV-1 vaccine (MVA-B) in HIV-1-infected patients alone or in combination with a drug to reactivate latent HIV-1. J. Antimicrob. Chemother. 2015, 70, 1833–1842. [Google Scholar] [CrossRef] [PubMed]
- Gori, A.; Longhi, R.; Peri, C.; Colombo, G. Peptides for immunological purposes: Design, strategies and applications. Amino Acids 2013, 45, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Borthwick, N.J.; Gilmour, J.; Hayes, P.; Dorrell, L.; Hanke, T. Control of HIV-1 replication in vitro by vaccine-induced human CD8+ T cells through conserved subdominant pol epitopes. Vaccine 2016, 34, 1215–1224. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Borthwick, N.; Ahmed, T.; Ondondo, B.; Hayes, P.; Rose, A.; Ebrahimsa, U.; Hayton, E.J.; Black, A.; Bridgeman, A.; Rosario, M.; et al. Vaccine-elicited human T cells recognizing conserved protein regions inhibit HIV-1. Mol. Ther. 2014, 22, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Valentin, A.; Dayton, F.; Kulkarni, V.; Alicea, C.; Rosati, M.; Chowdhury, B.; Gautam, R.; Broderick, K.E.; Sardesai, N.Y.; et al. DNA prime-boost vaccine regimen to increase breadth, magnitude, and cytotoxicity of the cellular immune responses to subdominant Gag epitopes of simian immunodeficiency virus and HIV. J. Immunol. 2016, 197, 3999–4013. [Google Scholar] [CrossRef] [PubMed]
- Letourneau, S.; Im, E.J.; Mashishi, T.; Brereton, C.; Bridgeman, A.; Yang, H.; Dorrell, L.; Dong, T.; Korber, B.; McMichael, A.J.; et al. Design and pre-clinical evaluation of a universal HIV-1 vaccine. PLoS ONE 2007, 2, e984. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Hu, X.; Llano, A.; Rosati, M.; Olvera, A.; Kulkarni, V.; Valentin, A.; Alicea, C.; Pilkington, G.R.; Sardesai, N.Y.; et al. A human immune data-informed vaccine concept elicits strong and broad T-cell specificities associated with HIV-1 control in mice and macaques. J. Transl. Med. 2015, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Ondondo, B.; Murakoshi, H.; Clutton, G.; Abdul-Jawad, S.; Wee, E.G.; Gatanaga, H.; Oka, S.; McMichael, A.J.; Takiguchi, M.; Korber, B.; et al. Novel conserved-region T-cell mosaic vaccine with high global HIV-1 coverage is recognized by protective responses in untreated infection. Mol. Ther. 2016, 24, 832–842. [Google Scholar] [CrossRef] [PubMed]
- Yang, O.O.; Ali, A.; Kasahara, N.; Faure-Kumar, E.; Bae, J.Y.; Picker, L.J.; Park, H. Short conserved sequences of HIV-1 are highly immunogenic and shift immunodominance. J. Virol. 2015, 89, 1195–1204. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hayton, E.J.; Rose, A.; Ibrahimsa, U.; Del Sorbo, M.; Capone, S.; Crook, A.; Black, A.P.; Dorrell, L.; Hanke, T. Safety and tolerability of conserved region vaccines vectored by plasmid DNA, simian adenovirus and modified vaccinia virus Ankara administered to human immunodeficiency virus type 1-uninfected adults in a randomized, single-blind phase I trial. PLoS ONE 2014, 9, e101591. [Google Scholar] [CrossRef] [PubMed]
- Moyo, N.; Borthwick, N.J.; Wee, E.G.; Capucci, S.; Crook, A.; Dorrell, L.; Hanke, T. Long-term follow up of human T-cell responses to conserved HIV-1 regions elicited by DNA/simian adenovirus/MVA vaccine regimens. PLoS ONE 2017, 12, e0181382. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.E.; Perdiguero, B.; Cepeda, M.V.; Mingorance, L.; Garcia-Arriaza, J.; Vandermeeren, A.; Sorzano, C.O.; Esteban, M. High, broad, polyfunctional, and durable T cell immune responses induced in mice by a novel hepatitis C virus (HCV) vaccine candidate (MVA-HCV) based on modified vaccinia virus Ankara expressing the nearly full-length HCV genome. J. Virol. 2013, 87, 7282–7300. [Google Scholar] [CrossRef] [PubMed]
- Kuiken, C.; Korber, B.; Shafer, R.W. HIV sequence databases. AIDS Rev. 2003, 5, 52–61. [Google Scholar] [PubMed]
- Kent, W.J. BLAT—The BLAST-like alignment tool. Genome Res. 2002, 12, 656–664. [Google Scholar] [CrossRef] [PubMed]
- Ward, G.A.; Stover, C.K.; Moss, B.; Fuerst, T.R. Stringent chemical and thermal regulation of recombinant gene expression by vaccinia virus vectors in mammalian cells. Proc. Natl. Acad. Sci. USA 1995, 92, 6773–6777. [Google Scholar] [CrossRef] [PubMed]
- Gomez, C.E.; Najera, J.L.; Jimenez, E.P.; Jimenez, V.; Wagner, R.; Graf, M.; Frachette, M.J.; Liljestrom, P.; Pantaleo, G.; Esteban, M. Head-to-head comparison on the immunogenicity of two HIV/AIDS vaccine candidates based on the attenuated poxvirus strains MVA and NYVAC co-expressing in a single locus the HIV-1BX08 gp120 and HIV-1(IIIB) Gag-Pol-Nef proteins of clade B. Vaccine 2007, 25, 2863–2885. [Google Scholar] [CrossRef] [PubMed]
- Tscharke, D.C.; Woo, W.P.; Sakala, I.G.; Sidney, J.; Sette, A.; Moss, D.J.; Bennink, J.R.; Karupiah, G.; Yewdell, J.W. Poxvirus CD8+ T-cell determinants and cross-reactivity in BALB/c mice. J. Virol. 2006, 80, 6318–6323. [Google Scholar] [CrossRef] [PubMed]
- Najera, J.L.; Gomez, C.E.; Garcia-Arriaza, J.; Sorzano, C.O.; Esteban, M. Insertion of vaccinia virus C7L host range gene into NYVAC-B genome potentiates immune responses against HIV-1 antigens. PLoS ONE 2010, 5, e11406. [Google Scholar] [CrossRef] [PubMed]
- Roederer, M.; Nozzi, J.L.; Nason, M.C. Spice: Exploration and analysis of post-cytometric complex multivariate datasets. Cytometry A 2011, 79, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Song, W.T.; Li, Y.; Chen, W.J.; Yang, D.; Zhong, G.C.; Zhou, H.Z.; Ren, C.Y.; Yu, H.T.; Ling, H. Improved expression of secretory and trimeric proteins in mammalian cells via the introduction of a new trimer motif and a mutant of the tpa signal sequence. Appl. Microbiol. Biotechnol. 2011, 91, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Llano, A.; Ibarrondo, J.; Daniels, M.; Miranda, C.; Zamarreno, J.; Bach, V.; Zuniga, R.; Perez-Alvarez, S.; Berger, C.T.; et al. Definition of the viral targets of protective HIV-1-specific T cell responses. J. Transl. Med. 2011, 9, 208. [Google Scholar] [CrossRef] [PubMed]
- Murakoshi, H.; Akahoshi, T.; Koyanagi, M.; Chikata, T.; Naruto, T.; Maruyama, R.; Tamura, Y.; Ishizuka, N.; Gatanaga, H.; Oka, S.; et al. Clinical control of HIV-1 by cytotoxic t cells specific for multiple conserved epitopes. J. Virol. 2015, 89, 5330–5339. [Google Scholar] [CrossRef] [PubMed]
- Rolland, M.; Manocheewa, S.; Swain, J.V.; Lanxon-Cookson, E.C.; Kim, M.; Westfall, D.H.; Larsen, B.B.; Gilbert, P.B.; Mullins, J.I. HIV-1 conserved-element vaccines: Relationship between sequence conservation and replicative capacity. J. Virol. 2013, 87, 5461–5467. [Google Scholar] [CrossRef] [PubMed]
- Mothe, B.; Llano, A.; Ibarrondo, J.; Zamarreno, J.; Schiaulini, M.; Miranda, C.; Ruiz-Riol, M.; Berger, C.T.; Herrero, M.J.; Palou, E.; et al. Ctl responses of high functional avidity and broad variant cross-reactivity are associated with HIV control. PLoS ONE 2012, 7, e29717. [Google Scholar] [CrossRef] [PubMed]
- Yusim, K.; Korber, B.T.; Brander, C.; Barouch, D.; de Boer, R.; Haynes, B.F.; Koup, R.; Moore, J.P.; Walker, B.D.; Watkins, D. HIV Molecular Immunology 2015; Los Alamos National Lab. (LANL): Los Alamos, NM, USA, 2016.
- Kaufmann, D.E.; Bailey, P.M.; Sidney, J.; Wagner, B.; Norris, P.J.; Johnston, M.N.; Cosimi, L.A.; Addo, M.M.; Lichterfeld, M.; Altfeld, M.; et al. Comprehensive analysis of human immunodeficiency virus type 1-specific CD4 responses reveals marked immunodominance of gag and nef and the presence of broadly recognized peptides. J. Virol. 2004, 78, 4463–4477. [Google Scholar] [CrossRef] [PubMed]
- Laher, F.; Ranasinghe, S.; Porichis, F.; Mewalal, N.; Pretorius, K.; Ismail, N.; Buus, S.; Stryhn, A.; Carrington, M.; Walker, B.D.; et al. HIV controllers exhibit enhanced frequencies of major histocompatibility complex class II tetramer+ Gag-specific CD4+ T cells in chronic clade C HIV-1 infection. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Schieffer, M.; Jessen, H.K.; Oster, A.F.; Pissani, F.; Soghoian, D.Z.; Lu, R.; Jessen, A.B.; Zedlack, C.; Schultz, B.T.; Davis, I.; et al. Induction of Gag-specific CD4 T cell responses during acute HIV infection is associated with improved viral control. J. Virol. 2014, 88, 7357–7366. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Rahman, T.; Chakravorty, R. Characterization of the protective HIV-1 CTL epitopes and the corresponding HLA class I alleles: A step towards designing CTL based HIV-1 vaccine. Adv. Virol. 2014, 2014, 321974. [Google Scholar] [CrossRef] [PubMed]
- Vinuesa, C.G.; Linterman, M.A.; Yu, D.; MacLennan, I.C. Follicular helper T cells. Annu. Rev. Immunol. 2016, 34, 335–368. [Google Scholar] [CrossRef] [PubMed]
- Havenar-Daughton, C.; Lee, J.H.; Crotty, S. Tfh cells and HIV bnAbs, an immunodominance model of the HIV neutralizing antibody generation problem. Immunol. Rev. 2017, 275, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Lynch, R.M.; Gautam, R.; Matus-Nicodemos, R.; Schmidt, S.D.; Boswell, K.L.; Darko, S.; Wong, P.; Sheng, Z.; Petrovas, C.; et al. Quality and quantity of Tfh cells are critical for broad antibody development in shivad8 infection. Sci. Transl. Med. 2015, 7, 298ra120. [Google Scholar] [CrossRef] [PubMed]
- Hancock, G.; Yang, H.; Yorke, E.; Wainwright, E.; Bourne, V.; Frisbee, A.; Payne, T.L.; Berrong, M.; Ferrari, G.; Chopera, D.; et al. Identification of effective subdominant anti-HIV-1 CD8+ T cells within entire post-infection and post-vaccination immune responses. PLoS Pathog. 2015, 11, e1004658. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, V.; Valentin, A.; Rosati, M.; Alicea, C.; Singh, A.K.; Jalah, R.; Broderick, K.E.; Sardesai, N.Y.; Le Gall, S.; Mothe, B.; et al. Altered response hierarchy and increased T-cell breadth upon HIV-1 conserved element DNA vaccination in macaques. PLoS ONE 2014, 9, e86254. [Google Scholar] [CrossRef] [PubMed]
- Ondondo, B.; Abdul-Jawad, S.; Bridgeman, A.; Hanke, T. Characterization of T-cell responses to conserved regions of the HIV-1 proteome in BALB/C mice. Clin. Vaccine Immunol. 2014, 21, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Valentin, A.; Cai, Y.; Dayton, F.; Rosati, M.; Ramirez-Salazar, E.G.; Kulkarni, V.; Broderick, K.E.; Sardesai, N.Y.; Wyatt, L.S.; et al. DNA-vaccine induced long-lasting cytotoxic T cells targeting conserved elements of HIV Gag are boosted upon DNA or rMVA vaccination. Hum. Gene Ther. 2018. [Google Scholar] [CrossRef] [PubMed]
- Streeck, H.; Lu, R.; Beckwith, N.; Milazzo, M.; Liu, M.; Routy, J.P.; Little, S.; Jessen, H.; Kelleher, A.D.; Hecht, F.; et al. Emergence of individual HIV-specific CD8 T cell responses during primary HIV-1 infection can determine long-term disease outcome. J. Virol. 2014, 88, 12793–12801. [Google Scholar] [CrossRef] [PubMed]
- Schultz, B.T.; Teigler, J.E.; Pissani, F.; Oster, A.F.; Kranias, G.; Alter, G.; Marovich, M.; Eller, M.A.; Dittmer, U.; Robb, M.L.; et al. Circulating HIV-specific interleukin-21+ CD4+ T cells represent peripheral Tfh cells with antigen-dependent helper functions. Immunity 2016, 44, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Locci, M.; Havenar-Daughton, C.; Landais, E.; Wu, J.; Kroenke, M.A.; Arlehamn, C.L.; Su, L.F.; Cubas, R.; Davis, M.M.; Sette, A.; et al. Human circulating PD-1+CXCR3-CXCR5+ memory Tfh cells are highly functional and correlate with broadly neutralizing HIV antibody responses. Immunity 2013, 39, 758–769. [Google Scholar] [CrossRef] [PubMed]
- Claireaux, M.; Galperin, M.; Benati, D.; Nouel, A.; Mukhopadhyay, M.; Klingler, J.; de Truchis, P.; Zucman, D.; Hendou, S.; Boufassa, F.; et al. A high frequency of HIV-specific circulating follicular helper t cells is associated with preserved memory B cell responses in HIV controllers. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Ciabattini, A.; Pettini, E.; Medaglini, D. CD4+ T cell priming as biomarker to study immune response to preventive vaccines. Front. Immunol. 2013, 4, 421. [Google Scholar] [CrossRef] [PubMed]
- Buchbinder, S.P.; Grunenberg, N.A.; Sanchez, B.J.; Seaton, K.E.; Ferrari, G.; Moody, M.A.; Frahm, N.; Montefiori, D.C.; Hay, C.M.; Goepfert, P.A.; et al. Immunogenicity of a novel clade B HIV-1 vaccine combination: Results of phase 1 randomized placebo controlled trial of an HIV-1 GM-CSF-expressing DNA prime with a modified vaccinia Ankara vaccine boost in healthy HIV-1 uninfected adults. PLoS ONE 2017, 12, e0179597. [Google Scholar] [CrossRef] [PubMed]
- Joachim, A.; Munseri, P.J.; Nilsson, C.; Bakari, M.; Aboud, S.; Lyamuya, E.F.; Tecleab, T.; Liakina, V.; Scarlatti, G.; Robb, M.L.; et al. Three-year durability of immune responses induced by HIV-DNA and HIV-modified vaccinia virus Ankara and effect of a late HIV-modified vaccinia virus Ankara boost in tanzanian volunteers. AIDS Res. Hum. Retroviruses 2017, 33, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Thompson, M.; Heath, S.L.; Sweeton, B.; Williams, K.; Cunningham, P.; Keele, B.F.; Sen, S.; Palmer, B.E.; Chomont, N.; Xu, Y.; et al. DNA/MVA vaccination of HIV-1 infected participants with viral suppression on antiretroviral therapy, followed by treatment interruption: Elicitation of immune responses without control of re-emergent virus. PLoS ONE 2016, 11, e0163164. [Google Scholar] [CrossRef] [PubMed]
- Pitman, M.C.; Lau, J.S.Y.; McMahon, J.H.; Lewin, S.R. Barriers and strategies to achieve a cure for HIV. Lancet HIV 2018, 5, e317–e328. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Segments | Length (aa) | Position on HXB2 | HIV-1 Protein | Coverage by HIV-1 Clade B Consensus Peptide Pools |
|---|---|---|---|---|
| S1 | 85 | 17–101 | Gag-p17 | Gag-1 |
| S2 | 72 | 138–209 | Gag-p24 | Gag-1 |
| S3 | 104 | 259–362 | Gag-p24 | Gag-2 |
| S4 | 107 | 101–207 | Pol-Prt | GPN-1 |
| S5 | 34 | 81–114 | Nef | GPN-2 |
| S6 | 13 | 56–68 | Nef | GPN-2 |
| S7 | 99 | 399–497 | Pol-RT | GPN-3 |
| S8 | 55 | 328–382 | Pol-RT | GPN-4 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perdiguero, B.; Raman, S.C.; Sánchez-Corzo, C.; S. Sorzano, C.O.; Valverde, J.R.; Esteban, M.; Gómez, C.E. Potent HIV-1-Specific CD8 T Cell Responses Induced in Mice after Priming with a Multiepitopic DNA-TMEP and Boosting with the HIV Vaccine MVA-B. Viruses 2018, 10, 424. https://doi.org/10.3390/v10080424
Perdiguero B, Raman SC, Sánchez-Corzo C, S. Sorzano CO, Valverde JR, Esteban M, Gómez CE. Potent HIV-1-Specific CD8 T Cell Responses Induced in Mice after Priming with a Multiepitopic DNA-TMEP and Boosting with the HIV Vaccine MVA-B. Viruses. 2018; 10(8):424. https://doi.org/10.3390/v10080424
Chicago/Turabian StylePerdiguero, Beatriz, Suresh C. Raman, Cristina Sánchez-Corzo, Carlos Oscar S. Sorzano, José Ramón Valverde, Mariano Esteban, and Carmen Elena Gómez. 2018. "Potent HIV-1-Specific CD8 T Cell Responses Induced in Mice after Priming with a Multiepitopic DNA-TMEP and Boosting with the HIV Vaccine MVA-B" Viruses 10, no. 8: 424. https://doi.org/10.3390/v10080424
APA StylePerdiguero, B., Raman, S. C., Sánchez-Corzo, C., S. Sorzano, C. O., Valverde, J. R., Esteban, M., & Gómez, C. E. (2018). Potent HIV-1-Specific CD8 T Cell Responses Induced in Mice after Priming with a Multiepitopic DNA-TMEP and Boosting with the HIV Vaccine MVA-B. Viruses, 10(8), 424. https://doi.org/10.3390/v10080424