Influenza Virus Infection of Human Lymphocytes Occurs in the Immune Cell Cluster of the Developing Antiviral Response

Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Sources and Culture Conditions
2.2. Purification of Peripheral Blood Monocytes-Macrophages, Total Lymphocytes, and Subsets of Lymphocytes
2.3. Exposure to Infectious Influenza Virus
2.4. Incremental PBMC Monocyte/Macrophage Depletion and Collection of Resting And Proliferating Lymphocytes
2.5. Flow Cytometry Analyses of Cell Phenotype, Cell Cycle, and Virus Uptake
2.6. Microscopic Analyses of Cell Clusters and Viral Protein Expression
2.7. Analyses of Protein Synthesis
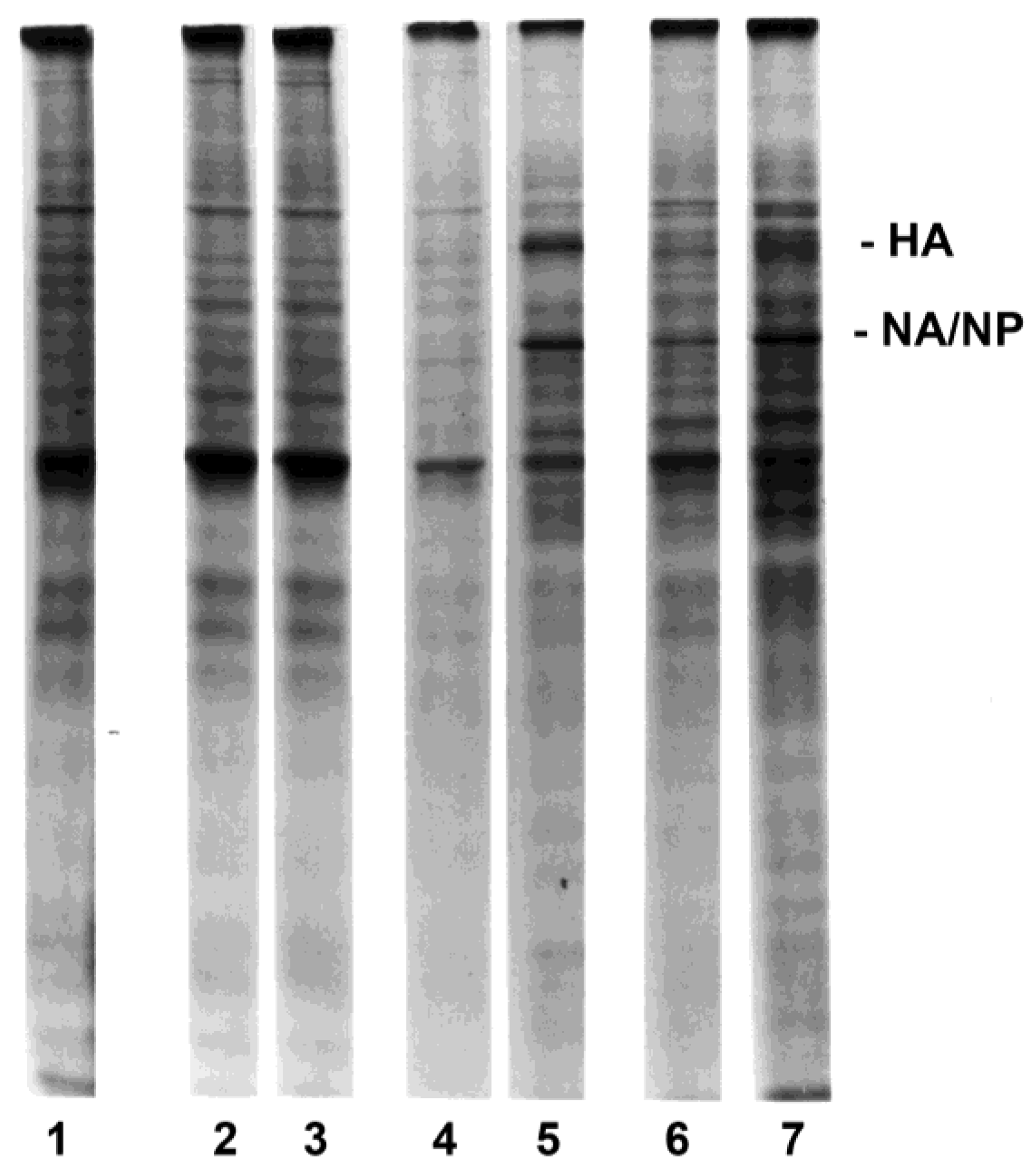
2.8. Northern Blot Analyses for Influenza Virus RNA
2.9. Infectious Focus Assay for Influenza Virus Infection
3. Results
3.1. Susceptibility of PBMC Monocyte/Macrophage and Lymphocyte Subpopulations to Influenza Virus Infection
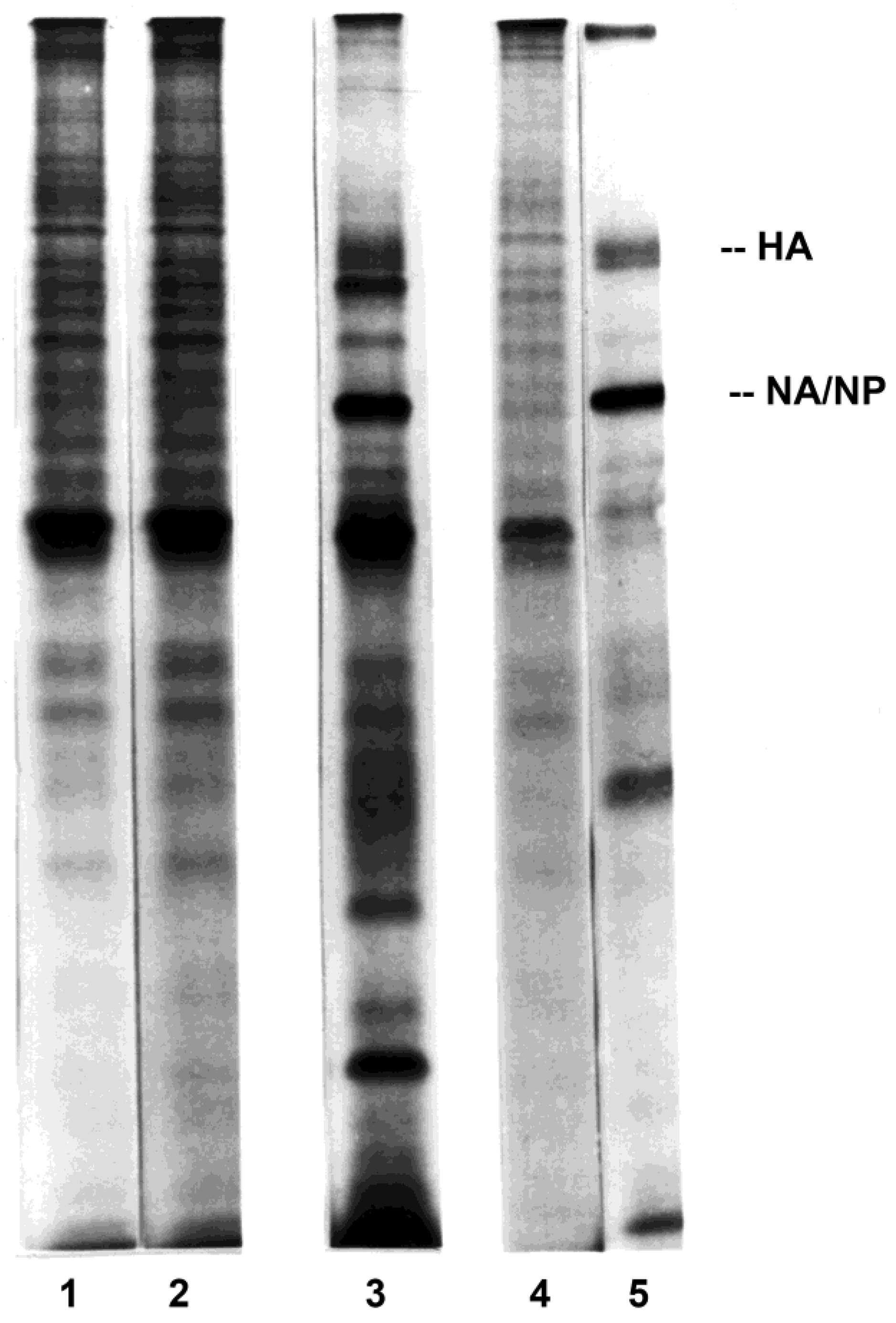
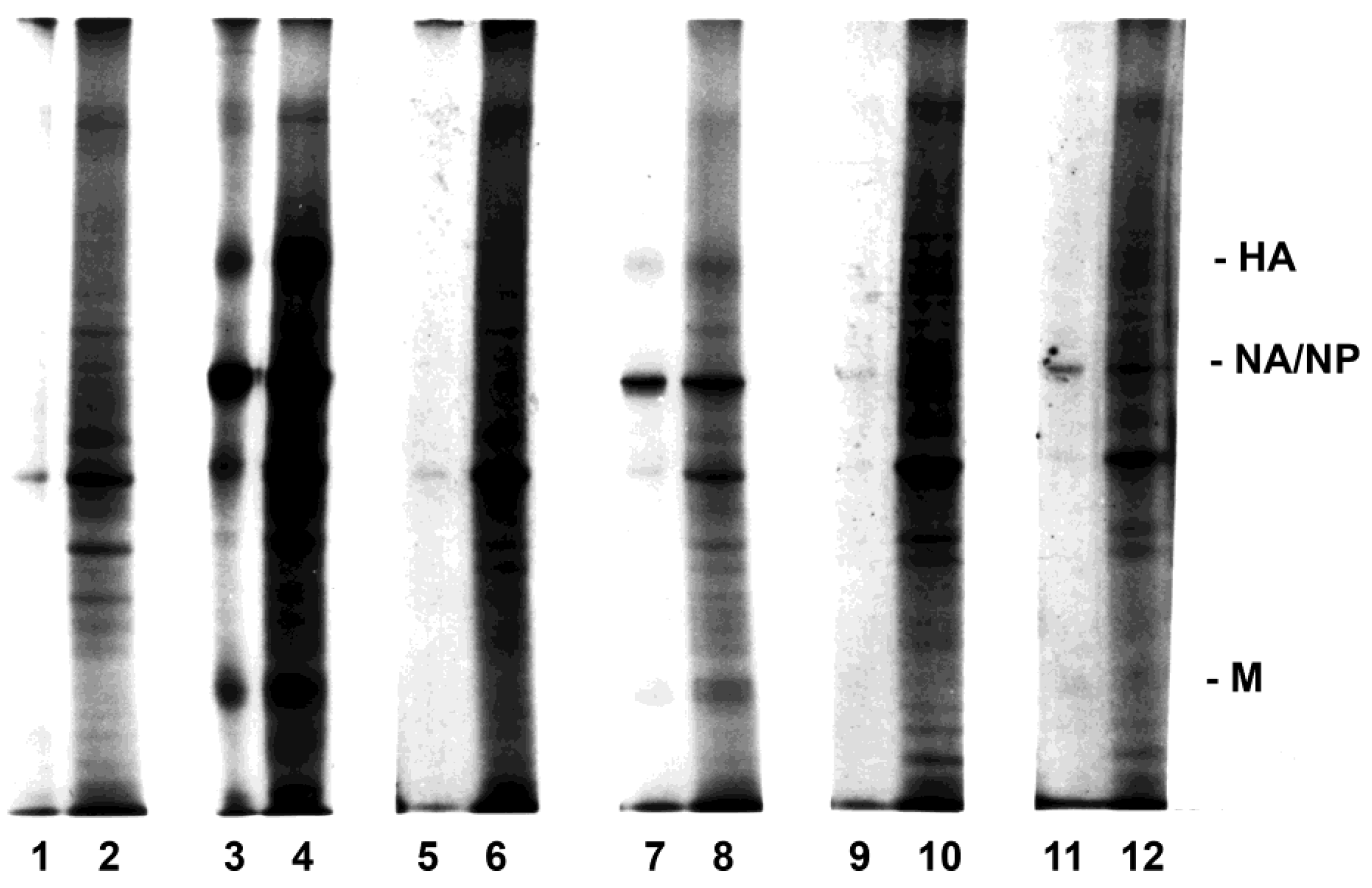
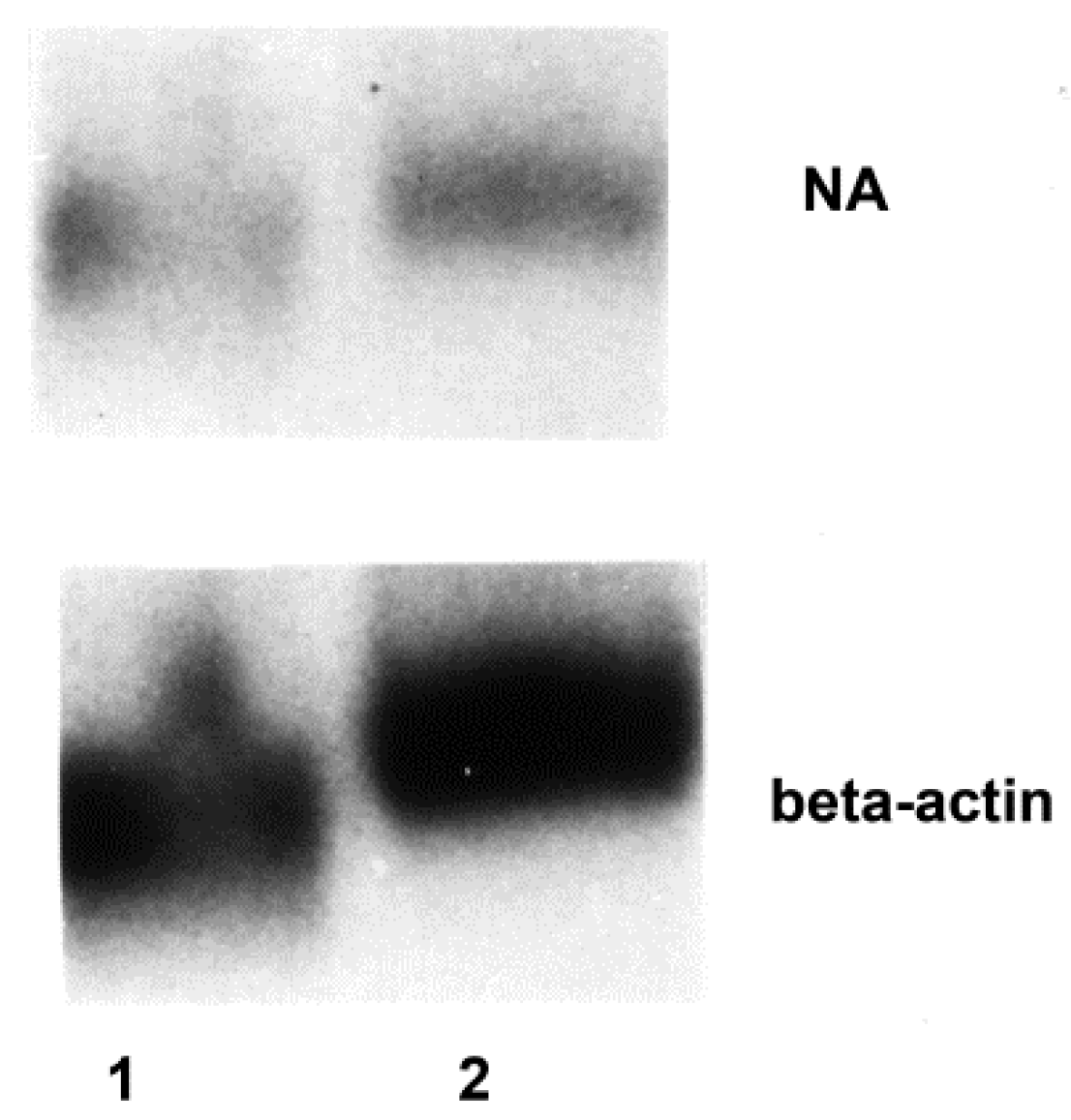
3.2. Influenza Virus Neuraminidase Transcription
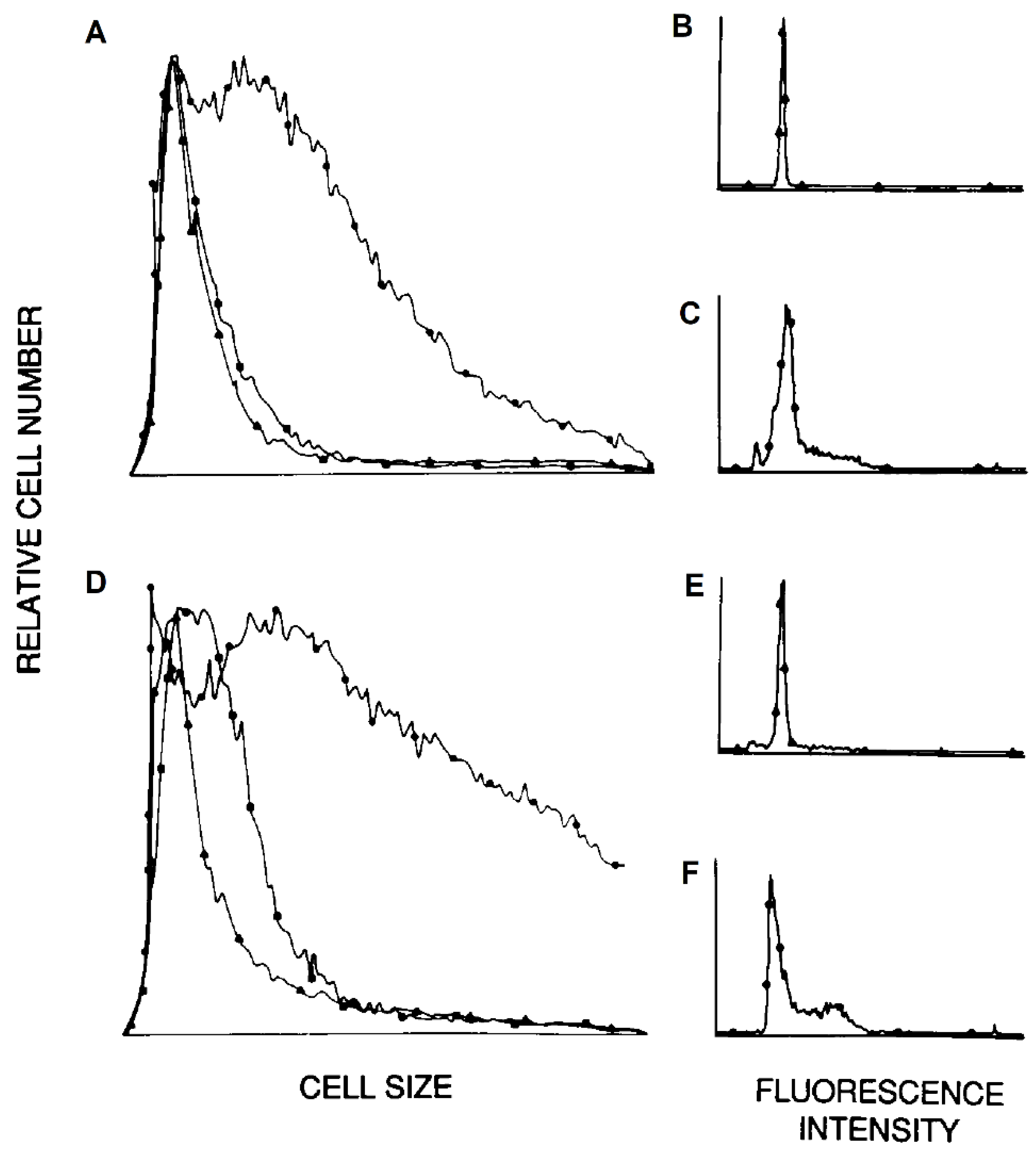
3.3. Lymphocyte Proliferation and Influenza Virus Infection
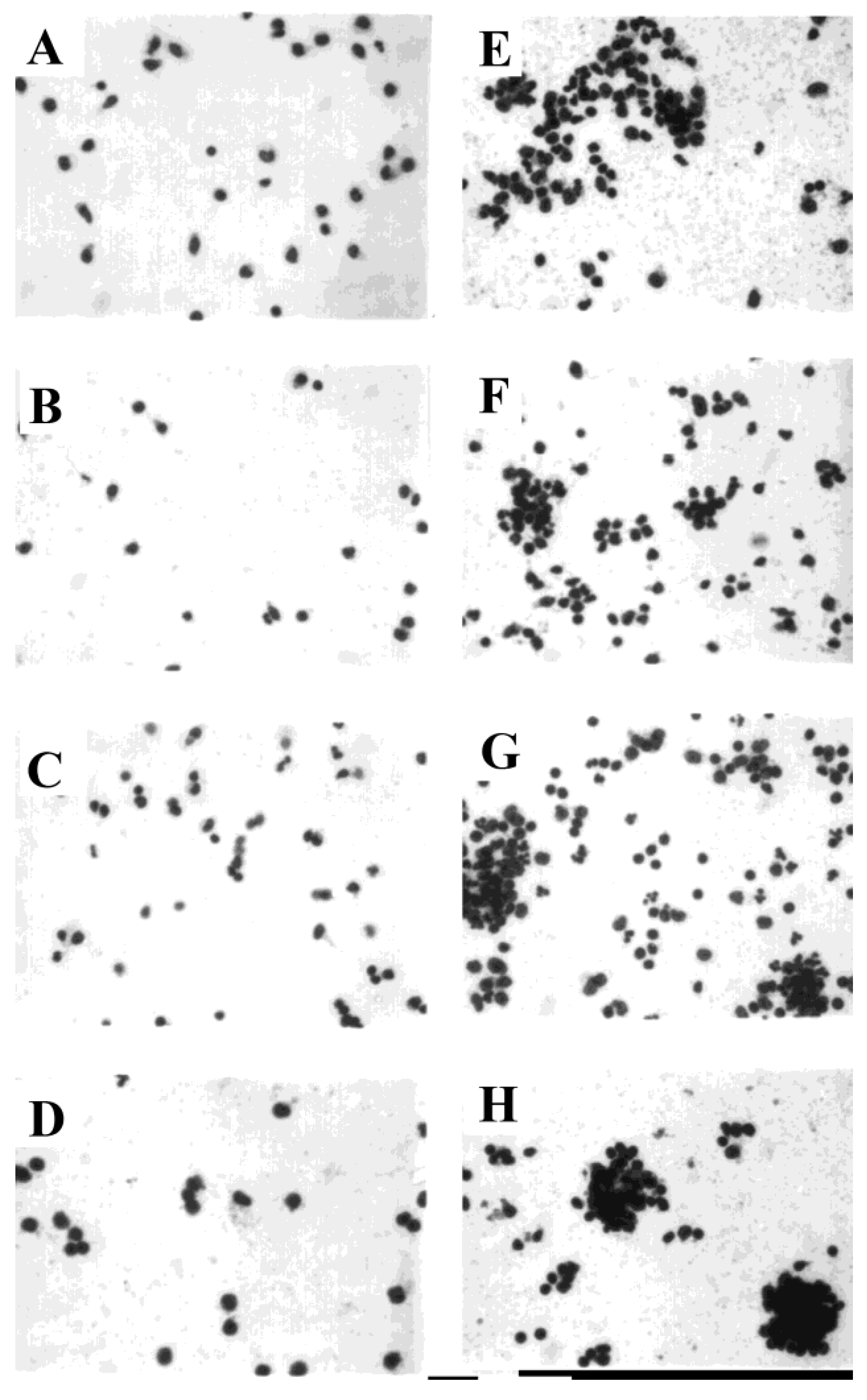
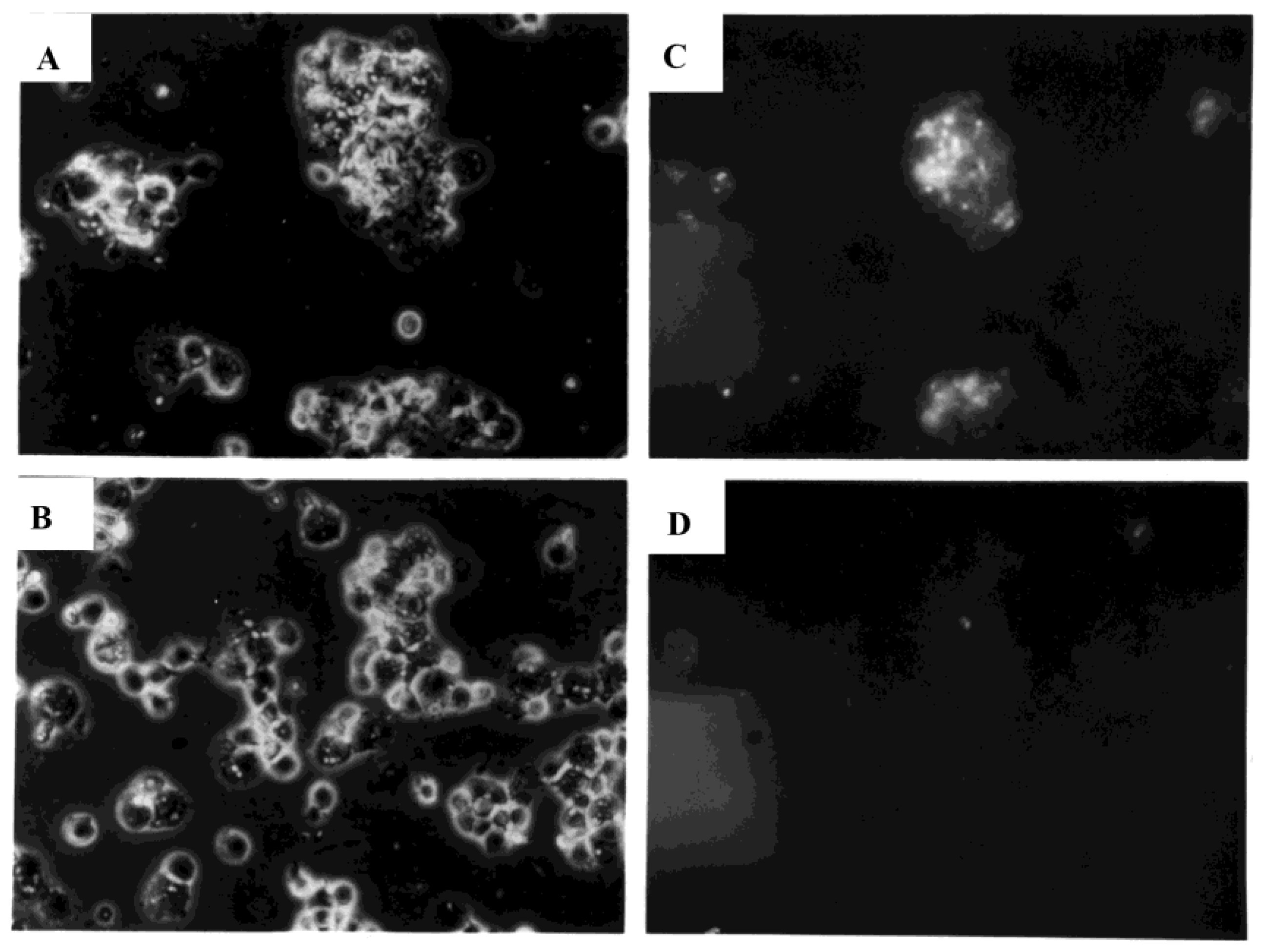
3.4. Development of Cell Clusters and Expression of Viral Proteins
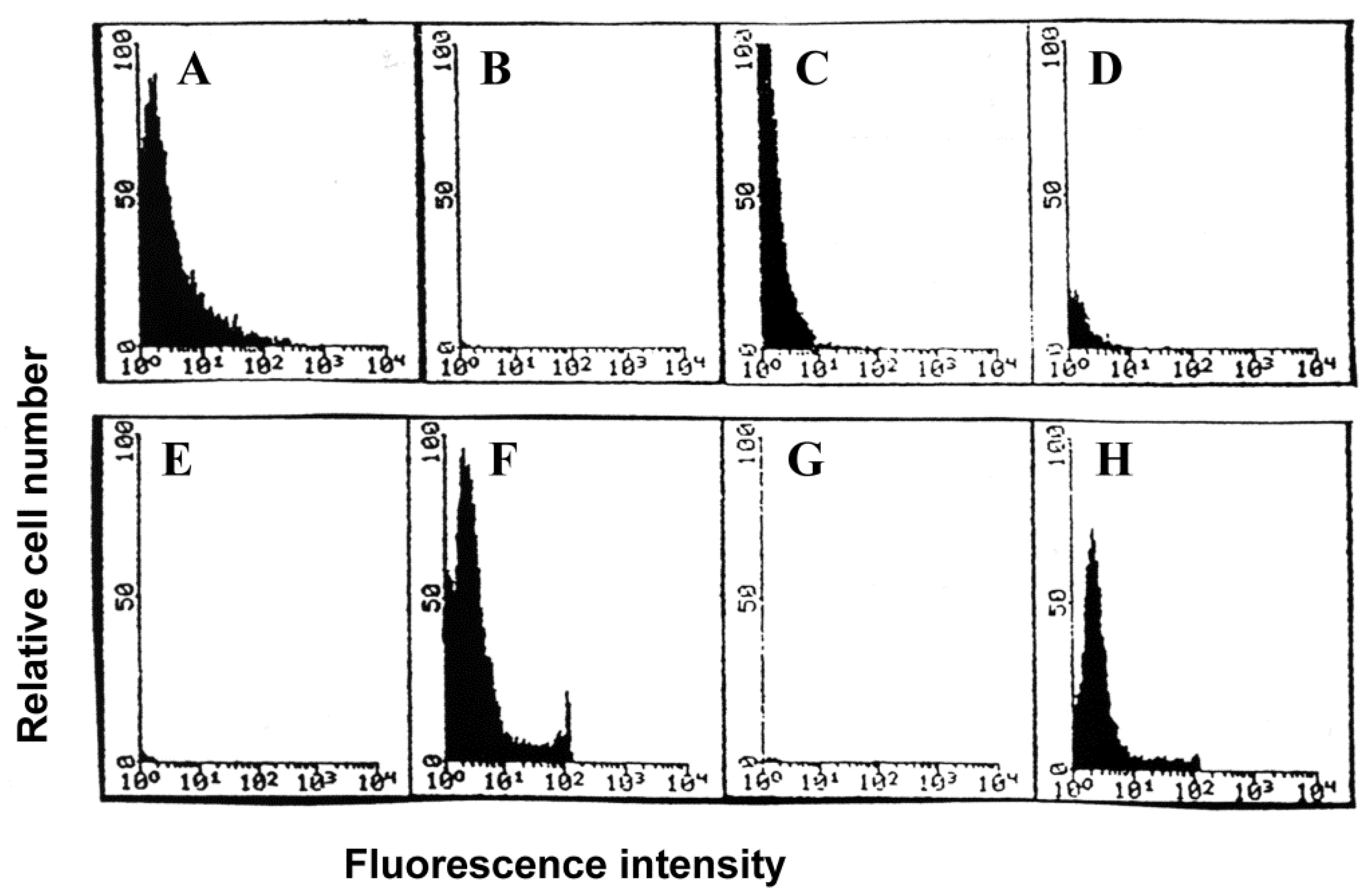
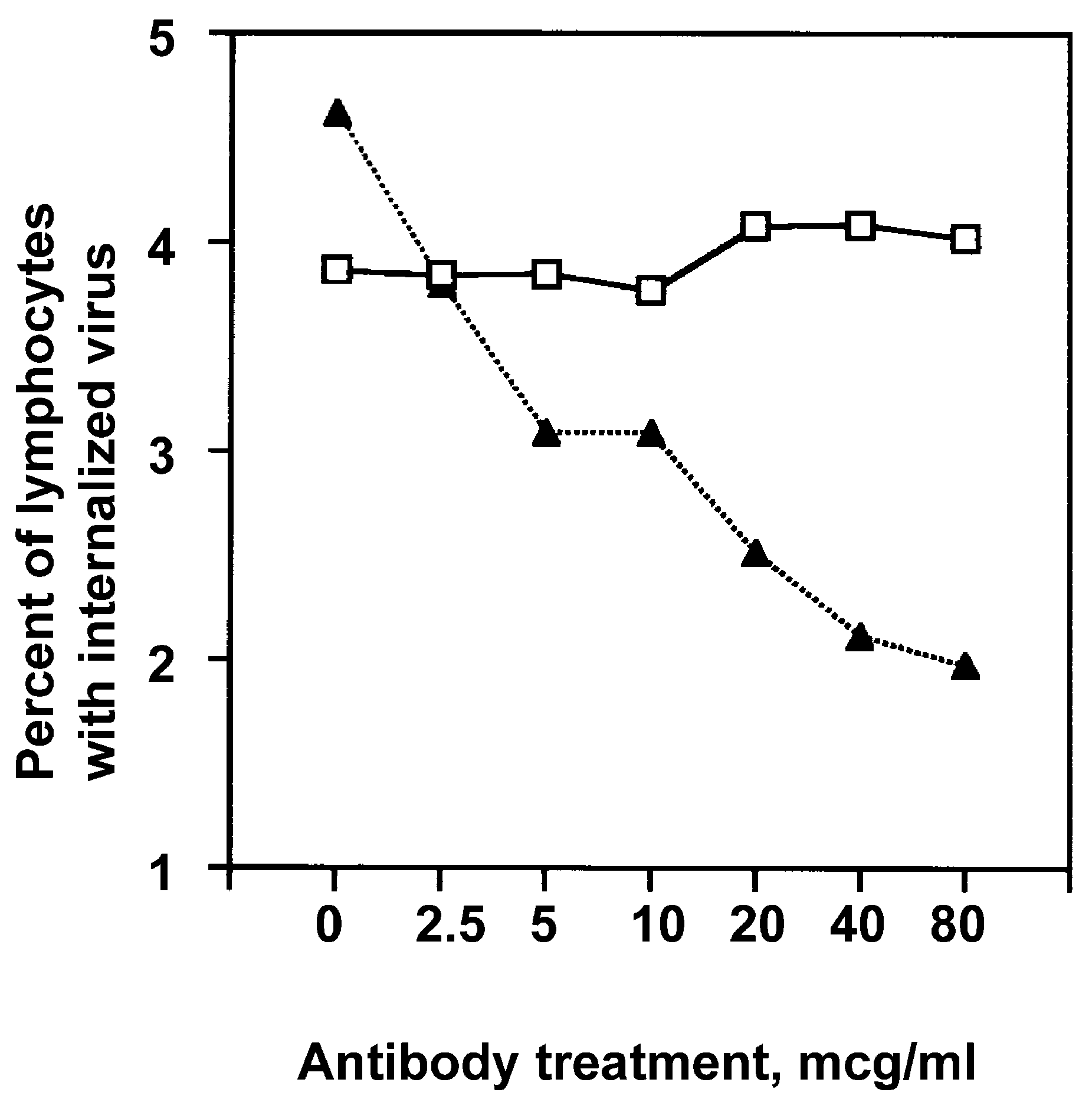
3.5. Uptake of FITC-Conjugated Influenza Virus by Lymphocytes
3.6. Uptake of FITC-Labeled Virus by Alveolar Lavage Versus Peripheral Blood Lymphocytes
3.7. Infectious Focus Potential of Influenza Virus-Exposed Lymphocytes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wyde, P.R.; Peavy, D.L.; Cate, T.R. Morphological and Cytochemical Characterization of Cells Infiltrating Mouse Lungs After Influenza Infection. Infect. Immun. 1978, 21, 140–146. [Google Scholar] [PubMed]
- Ennis, F.A.; Wells, M.A.; Butchko, G.M.; Albrecht, P. Evidence That Cytotoxic T Cells Are Part of the Host’s Response to Influenza Pneumonia. J. Exp. Med. 1978, 148, 1241–1250. [Google Scholar] [CrossRef] [PubMed]
- Cerwenka, A.; Morgan, T.M.; Dutton, R.W. Naive, Effector, and Memory CD8 T Cells in Protection Against Pulmonary Influenza Virus Infection: Homing Properties Rather Than Initial Frequencies Are Crucial. J. Immunol. 1999, 163, 5535–5543. [Google Scholar] [PubMed]
- Roman, E.; Miller, E.; Harmsen, A.; Wiley, J.; Von Andrian, U.H.; Huston, G.; Swain, S.L. CD4 Effector T Cell Subsets in the Response to Influenza: Heterogeneity, Migration, and Function. J. Exp. Med. 2002, 196, 957–968. [Google Scholar] [CrossRef] [PubMed]
- Wyde, P.R.; Wilson, M.R.; Cate, T.R. Interferon Production by Leukocytes Infiltrating the Lungs of Mice During Primary Influenza Virus Infection. Infect. Immun. 1982, 38, 1249–1255. [Google Scholar] [PubMed]
- MacKenzie, C.D.; Taylor, P.M.; Askonas, B.A. Rapid Recovery of Lung Histology Correlates With Clearance of Influenza Virus by Specific CD8+ Cytotoxic T Cells. Immunology 1989, 67, 375–381. [Google Scholar] [PubMed]
- Yap, K.L.; Ada, G.L.; McKenzie, I.F.C. Transfer of Specific Cytotoxic T Lymphocytes Protects Mice Inoculated With Influenza Virus. Nature 1978, 273, 238–239. [Google Scholar] [CrossRef] [PubMed]
- Wells, M.A.; Ennis, F.A.; Albrecht, P. Recovery From a Viral Respiratory Infection. II. Passive Transfer of Immune Spleen Cells to Mice With Influenza Pneumonia. J. Immunol. 1981, 126, 1042–1046. [Google Scholar] [PubMed]
- Singer, S.H.; Noguchi, P.; Kirschstein, R.L. Respiratory Diseases in Cyclophosphamide-Treated Mice. II. Decreased Virulence of PR8 Influenza Virus. Infect. Immun. 1972, 5, 957–960. [Google Scholar] [PubMed]
- Suzuki, F.; Ohya, J.; Ishida, N. Effect of Antilymphocyte Serum on Influenza Virus Infection. Proc. Soc. Exp. Biol. Med. 1974, 146, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Wells, M.A.; Albrecht, P.; Ennis, F.A. Recovery From a Viral Respiratory Infection. I. Influenza Pneumonia in Normal and T-Deficient Mice. J. Immunol. 1981, 126, 1036–1041. [Google Scholar] [PubMed]
- Roberts, N.J., Jr.; Domurat, F. Virus-Induced Immunosuppression: Influenza Virus. In Virus-Induced Immunosuppression; Specter, S., Bendinelli, M., Friedman, H., Eds.; Plenum Publishing, Inc.: New York, NY, USA, 1989. [Google Scholar]
- Grakoui, A.; Bromley, S.K.; Sumen, C.; Davis, M.M.; Shaw, A.S.; Allen, P.M.; Dustin, M.L. The Immunological Synapse: A Molecular Machine Controlling T Cell Activation (See Comments). Science 1999, 285, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Dustin, M.L.; Chakraborty, A.K.; Shaw, A.S. Understanding the Structure and Function of the Immunological Synapse. Cold Spring Harb. Perspect. Biol. 2010, 2, a002311. [Google Scholar] [CrossRef] [PubMed]
- Braendstrup, O.; Werdelin, O.; Shevach, E.M.; Rosenthal, A.S. Macrophage-Lymphocyte Clusters in the Immune Response to Soluble Protein Antigen in Vitro. VII. Genetically Restricted and Nonrestricted Physical Interactions. J. Immunol. 1979, 122, 1608–1613. [Google Scholar] [PubMed]
- Nielsen, M.H.; Jenson, H.; Braendstrup, O.; Werdelin, O. Macrophage-Lymphocyte Clusters in the Immune Response to Soluble Protein Antigen in Vitro. II. Ultrastructure of Clusters Formed during the Early Response. J. Exp. Med. 1974, 140, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- Werdelin, O.; Braendstrup, O.; Pedersen, E. Macrophage-Lymphocyte Clusters in the Immune Response to Soluble Protein Antigen in Vitro. I. Roles of Lymphocytes and Macrophages in Cluster Formation. J. Exp. Med. 1974, 140, 1245–1259. [Google Scholar] [CrossRef] [PubMed]
- Kuijpers, K.C.; Kuijpers, T.W.; Zeijlemaker, W.P.; Lucas, C.J.; van Lier, R.A.W.; Miedema, F. Analysis of the Role of Leukocyte Function-Associated Antigen-1 in Activation of Human Influenza Virus-Specific T Cell Clones. J. Immunol. 1990, 144, 3281–3287. [Google Scholar] [PubMed]
- Salkind, A.R.; Nichols, J.E.; Roberts, N.J., Jr. Suppressed Expression of ICAM-1 and LFA-1 and Abrogation of Leukocyte Collaboration After Exposure of Human Mononuclear Leukocytes to Respiratory Syncytial Virus in Vitro: Comparison With Exposure to Influenza Virus. J. Clin. Investig. 1991, 88, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Mock, D.J.; Domurat, F.; Roberts, N.J., Jr.; Walsh, E.E.; Licht, M.R.; Keng, P. Macrophages Are Required for Influenza Virus Infection of Human Lymphocytes. J. Clin. Investig. 1987, 79, 620–624. [Google Scholar] [CrossRef] [PubMed]
- Hackeman, M.M.A.; Denman, A.M.; Tyrrell, D.A.J. Inactivation of Influenza Virus by Human Lymphocytes. Clin. Exp. Immunol. 1974, 16, 583–591. [Google Scholar]
- Roberts, N.J., Jr.; Horan, P.K. Expression of Viral Antigens After Infection of Human Lymphocytes, Monocytes, and Macrophages With Influenza Virus. J. Infect. Dis. 1985, 151, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Sweet, C.; Smith, H. Pathogenicity of Influenza Virus. Microbiol. Rev. 1980, 44, 303–330. [Google Scholar] [PubMed]
- Lipsky, P.; Rosenthal, A.S. Macrophage-Lymphocyte Interaction. II. Antigen-Mediated Physical Interactions Between Immune Guinea Pig Lymph Node Lymphocytes and Syngeneic Macrophages. J. Exp. Med. 1975, 141, 138–154. [Google Scholar] [CrossRef] [PubMed]
- Boyum, A. Isolation of Mononuclear Cells and Granulocytes From Human Blood. Scand. J. Clin. Lab. Investig. 1968, 21 (Suppl. S97), 77–89. [Google Scholar]
- Frampton, M.W.; Smeglin, A.M.; Roberts, N.J., Jr.; Morrow, P.E.; Utell, M.J. Nitrogen Dioxide Exposure in Vivo and Human Alveolar Macrophage Inactivation of Influenza Virus in Vitro. Environ. Res. 1989, 48, 179–192. [Google Scholar] [CrossRef]
- Roberts, N.J., Jr.; Steigbigel, R.T. Effect of in Vitro Virus Infection on Response of Human Monocytes and Lymphocytes to Mitogen Stimulation. J. Immunol. 1978, 121, 1052–1058. [Google Scholar] [PubMed]
- Roberts, N.J., Jr.; Douglas, R.G., Jr.; Simons, R.L.; Diamond, M.E. Virus-Induced Interferon Production by Human Macrophages. J. Immunol. 1979, 123, 365–369. [Google Scholar] [PubMed]
- Keng, P.; Li, C.K.N.; Wheeler, K.T. Characterization of the Separation Properties of the Beckman Elutriator System. Cell Biochem. Biophys. 1981, 3, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Wahl, L.M.; Katona, I.M.; Wilder, R.L.; Winter, C.C.; Haraouri, B.; Scher, I.; Wahl, S.M. Isolation of Human Mononuclear Cell Subsets by Counterflow Centrifugal Elutriation (CCE). I. Characterization of B-Lymphocyte-, T-Lymphocyte-, and Monocyte-Enriched Fractions by Flow Cytometry Analysis. Cell. Immunol. 1984, 85, 373–383. [Google Scholar] [CrossRef]
- Domurat, F.M.; Keng, P.; Mock, D.J.; Prill, A.H.; Roberts, N.J., Jr. Early Identification and Retrieval or Deletion of Human Lymphocyte Subpopulations Responding to Influenza Virus or Respiratory Syncytial Virus Challenge. Cell Biophys. 1989, 15, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Wysocki, L.J.; Sato, V.L. “Panning” for Lymphocytes: A Method for Cell Selection. Proc. Natl. Acad. Sci. USA 1978, 75, 2844–2848. [Google Scholar] [CrossRef] [PubMed]
- Eisenlohr, L.C.; Gerhard, W.; Hackett, C.J. Role of Receptor-Binding Activity of the Viral Hemagglutinin Molecule in the Presentation of Influenza Virus Antigens to Helper T Cells. J. Virol. 1987, 61, 1375–1383. [Google Scholar] [PubMed]
- Matlin, K.S.; Reggio, H.; Helenius, A.; Simons, K. Infectious Entry Pathway of Influenza Virus in a Canine Kidney Cell Line. J. Cell Biol. 1981, 91, 601–613. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.E.; Mock, D.J.; Roberts, N.J., Jr. Use of FITC-Labeled Influenza Virus and Flow Cytometry to Assess Binding and Internalization of Virus by Monocytes-Macrophages and Lymphocytes. Arch. Virol. 1992, 130, 441–455. [Google Scholar] [CrossRef]
- Yoshimura, A.; Ohnishi, S.-I. Uncoating of Influenza Virus in Endosomes. J. Virol. 1984, 51, 497–504. [Google Scholar] [PubMed]
- Ettensohn, D.B.; Roberts, N.J., Jr. Human Alveolar Macrophage Support of Lymphocyte Responses to Mitogens and Antigens: Analysis and Comparison With Autologous Peripheral-Blood-Derived Monocytes and Macrophages. Am. Rev. Respir. Dis. 1983, 128, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Roberts, N.J., Jr.; Nichols, J.E. Regulation of Proliferation After Influenza Virus Infection of Human Mononuclear Leukocytes. J. Med. Virol. 1989, 27, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Domurat, F.; Nikaein, A.; Mock, D.J.; Keng, P.C.; Roberts, N.J., Jr. Demonstration and Selective Abrogation of Class II HLA-DR Restricted Activity in Human Mononuclear Leukocytes Exposed to Respiratory Syncytial Virus. Hum. Immunol. 1985, 14, 164–165. [Google Scholar]
- Crissman, H.A.; Tobey, R.A. Cell-Cycle Analysis in 20 Minutes. Science 1974, 184, 1297–1298. [Google Scholar] [CrossRef] [PubMed]
- Darzynkiewicz, A.; Traganos, R.; Melamed, M.R. New Cell Cycle Compartments Identified by Multi-Parameter Flow Cytometry. Cytometry 1981, 1, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Fattorossi, A.; Nisini, R.; Pizzolo, J.G.; D’Amelio, R. New, Simple Flow Cytometry Technique to Discriminate Between Internalized and Membrane-Bound Particles in Phagocytosis. Cytometry 1989, 10, 320–325. [Google Scholar] [CrossRef] [PubMed]
- Salkind, A.R.; McCarthy, D.O.; Nichols, J.E.; Domurat, F.M.; Walsh, E.E.; Roberts, N.J., Jr. Interleukin-1 Inhibitor Activity Induced by Respiratory Syncytial Virus: Abrogation of Virus-Specific and Alternate Human Lymphocyte Proliferative Responses. J. Infect. Dis. 1991, 163, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins During the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Van Wyck, K.L.; Bean, W.J., Jr.; Webster, R.G. Monoclonal Antibodies to the Influenza A Virus Nucleoprotein Affecting RNA Transcription. J. Virol. 1981, 39, 313–317. [Google Scholar]
- Chomczynski, P.; Sacchi, N. Single Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef]
- Wahl, G.M.; Meinkoth, J.L.; Kimmel, A.R. Northern and Southern Blots. In Guide to Molecular Cloning Techniques. Methods in Enzymology; Berger, S.L., Kimmel, A.R., Eds.; Academic Press, Inc.: San Diego, CA, USA, 1987; Volume 152. [Google Scholar]
- Tobita, K.; Suguia, A.; Enomote, C.; Furuyama, M. Plaque Assay and Primary Isolation of Influenza A Viruses in an Established Line of Canine Kidney Cells (MDCK) in the Presence of Trypsin. Med. Microbiol. Immunol. 1975, 162, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Brownson, J.M.; Mahy, B.W.J.; Hazleman, B.L. Interaction of Influenza A Virus With Human Peripheral Blood Lymphocytes. Infect. Immun. 1979, 25, 749–756. [Google Scholar] [PubMed]
- Roberts, N.J., Jr.; NYU School of Medicine, New York, NY, USA. Unpublished work. 2018.
- Fooksman, D.R.; Vardhana, S.; Vasiliver-Shamis, G.; Liese, J.; Blair, D.A.; Waite, J.; Sacristan, C.; Victora, G.D.; Zanin-Zhorov, A.; Dustin, M.L. Functional Anatomy of T Cell Activation and Synapse Formation. Annu. Rev. Immunol. 2010, 28, 79–105. [Google Scholar] [CrossRef] [PubMed]
- Becker, S.; Harris, D.T.; Koren, H.S. Characterization of Normal Human Lung Lymphocytes and Interleukin-2-Induced Lung T Cell Lines. Am. J. Respir. Cell Mol. Biol. 1990, 3, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Frampton, M.W.; Voter, K.Z.; Morrow, P.E.; Roberts, N.J., Jr.; Culp, D.J.; Cox, C.; Utell, M.J. Sulfuric Acid Aerosol Exposure in Humans Assessed by Bronchoalveolar Lavage. Am. Rev. Respir. Dis. 1992, 146, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Ettensohn, D.B.; Frampton, M.W.; Nichols, J.E.; Roberts, N.J., Jr. Human Alveolar Macrophages May Not Be Susceptible to Direct Infection by a Human Influenza Virus. J. Infect. Dis. 2016, 214, 1658–1665. [Google Scholar] [CrossRef] [PubMed]
- He, X.S.; Mahmood, K.; Maecker, H.T.; Holmes, T.H.; Kemble, G.W.; Arvin, A.M.; Greenberg, H.B. Analysis of the Frequencies and of the Memory T Cell Phenotypes of Human CD8+ T Cells Specific for Influenza A Viruses. J. Infect. Dis. 2003, 187, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.E.; Walsh, E.E.; Falsey, A.R.; Liu, N.; Liu, D.; Divekar, A.; Snyder-Cappione, J.E.; Mosmann, T.R. The Balance Between Influenza- and RSV-Specific CD4 T Cells Secreting IL-10 or IFNγ in Young and Healthy-Elderly Subjects. Mech. Ageing Dev. 2005, 126, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Fleming, E.H.; Ochoa, E.E.; Nichols, J.E.; O’Banion, M.K.; Salkind, A.R.; Roberts, N.J., Jr. Reduced Activation and Proliferation of Human Lymphocytes Exposed to Respiratory Syncytial Virus Compared to Cells Exposed to Influenza Virus. J. Med. Virol. 2018, 90, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.L.; Manicassamy, B.; Lamb, R.A. Influenza A Virus Uses Intercellular Connections to Spread to Neighboring Cells. J. Virol. 2015, 89, 1537–1549. [Google Scholar] [CrossRef] [PubMed]
- Mock, D.J.; Roberts, N.J., Jr. Proposed Immunopathogenic Factors Associated With Progression From Human Immunodeficiency Virus Seropositivity to Clinical Disease. J. Clin. Microbiol. 1987, 25, 1817–1821. [Google Scholar] [PubMed]
- Zhong, P.; Agosto, L.M.; Munro, J.B.; Mothes, W. Cell-to-Cell Transmission of Viruses. Curr. Opin. Virol. 2013, 3, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Bracq, L.; Xie, M.; Benichou, S.; Bouchet, J. Mechanisms for Cell-to-Cell Transmission of HIV-1. Front. Immunol. 2018, 9, 260. [Google Scholar] [CrossRef] [PubMed]
- Goldwich, A.; Prechtel, A.T.; Muhl-Zurbes, P.; Pangratz, N.M.; Stossel, H.; Romani, N.; Steinkasserer, A.; Kummer, M. Herpes Simplex Virus Type I (HSV-1) Replicates in Mature Dendritic Cells but Can Only Be Transferred in a Cell-Cell Contact-Dependent Manner. J. Leukoc. Biol. 2011, 89, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Denman, A.M.; Rager-Zisman, B.; Merigan, T.C.; Tyrrell, D.A.J. Replication or Inactivation of Different Viruses by Human Lymphocyte Preparations. Infect. Immun. 1974, 9, 373–376. [Google Scholar] [PubMed]
- Duc-Nguyen, H.; Henle, W. Replication of Mumps Virus in Human Leukocyte Cultures. J. Bacteriol. 1966, 92, 258–265. [Google Scholar] [PubMed]
- Folks, T.; Kelly, J.; Benn, S.; Kinter, A.; Justement, J.; Gold, J.; Redfield, R.; Sell, K.W.; Fauci, A.S. Susceptibility of Normal Human Lymphocytes to Infection With HTLV III/LAV. J. Immunol. 1986, 136, 4049–4053. [Google Scholar] [PubMed]
- McChesney, M.B.; Oldstone, M.B.A. Viruses Perturb Lymphocyte Functions: Selected Principles Characterizing Virus-Induced Immunosuppression. Ann. Rev. Immunol. 1987, 5, 279–304. [Google Scholar] [CrossRef] [PubMed]
- McDougal, J.S.; Mawle, A.; Cort, S.P.; Nicholson, J.K.A.; Cross, G.D.; Scheppler-Campbell, J.A.; Hicks, D.; Sligh, J. Cellular Tropism of the Human Retrovirus HTLV-III/LAV. I. Role of T Cell Activation and Expression of the T4 Antigen. J. Immunol. 1985, 135, 3151–3162. [Google Scholar] [PubMed]
- Miller, G.; Enders, J.F. Vaccinia Virus Replication and Cytopathic Effect in Cultures of Phytohemagglutinin-Treated Human Peripheral Blood Leukocytes. J. Virol. 1968, 2, 787–792. [Google Scholar] [PubMed]
- Woodruff, J.F.; Woodruff, J.J. T Lymphocyte Interaction With Viruses and Virus-Infected Tissues. Prog. Med. Virol. 1975, 19, 120–160. [Google Scholar] [PubMed]
- Nikolaidis, N.M.; Noel, J.G.; Pitstick, L.B.; Gardner, J.C.; Uehara, Y.; Wu, H.; Saito, A.; Lewnard, K.E.; Liu, H.; White, M.R.; et al. Mitogenic Stimulation Accelerates Influenza-Induced Mortality by Increasing Susceptibility of Alveolar Type II Cells to Infection. Proc. Natl. Acad. Sci. USA 2017, 114, E6613–E6622. [Google Scholar] [CrossRef] [PubMed]
- Pizzolla, A.; Nguyen, T.H.; Sant, S.; Jaffar, J.; Loudovaris, T.; Mannering, S.I.; Thomas, P.G.; Westall, G.P.; Kedzierska, K.; Wakim, L.M. Influenza-Specific Lung-Resident Memory T Cells Are Proliferative and Polyfunctional and Maintain Diverse TCR Profiles. J. Clin. Investig. 2018, 128, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Lipscomb, M.F.; Yeakel-Houlihan, D.; Lyons, C.R.; Gleason, R.R.; Stein-Streilein, J. Persistence of Influenza As an Immunogen in Pulmonary Antigen-Presenting Cells. Infect. Immun. 1983, 42, 965–972. [Google Scholar] [PubMed]
- Jakab, G.J.; Astry, C.L.; Warr, G.A. Alveolitis Induced by Influenza Virus. Am. Rev. Respir. Dis. 1983, 128, 730–739. [Google Scholar] [PubMed]
- Pichyangkul, S.; Yongvanitchit, K.; Limsalakpetch, A.; Kum-Arb, U.; Im-Erbsin, R.; Boonnak, K.; Thitithayanont, A.; Jongkaewwattana, A.; Wiboon-Ut, S.; Mongkolsirichaikul, D.; et al. Tissue Distribution of Memory T and B Cells in Rhesus Monkeys Following Influenza A Infection. J. Immunol. 2015, 195, 4378–4386. [Google Scholar] [CrossRef] [PubMed]
- McMaster, S.R.; Wilson, J.J.; Wang, H.; Kohlmeier, J.E. Airway-Resident Memory CD8 T Cells Provide Antigen-Specific Protection Against Respiratory Virus Challenge Through Rapid IFN-γ Production. J. Immunol. 2015, 195, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.E.; Niles, J.A.; Roberts, N.J. Human Lymphocyte Apoptosis After Exposure to Influenza A Virus. J. Virol. 2001, 75, 5921–5929. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Cells | Percent of Cells Infected (Mean ± SE) | ||
|---|---|---|---|
| CD3+ | CD4+ | CD8+ | |
| Peripheral blood | 2.78 ± 0.60 | 1.88 ± 0.33 | 1.78 ± 0.48 |
| Bronchoalveolar lavage | 5.96 ± 3.37 | 4.46 ± 2.00 | 4.18 ± 0.97 |
| Cells | Experiment | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | Mean ± SE | |
| T lymphocytes | 2 | 3 | 1 | ||||
| CD4+ lymphocytes | 4 | 1 | 5 | 0.4 | 2 | 2 | 2.4 ± 0.7 |
| CD8+ lymphocytes | 1 | 1 | 5 | 0.1 | 0.5 | 1 | 1.4 ± 0.7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mock, D.J.; Frampton, M.W.; Nichols, J.E.; Domurat, F.M.; Signs, D.J.; Roberts, N.J., Jr. Influenza Virus Infection of Human Lymphocytes Occurs in the Immune Cell Cluster of the Developing Antiviral Response. Viruses 2018, 10, 420. https://doi.org/10.3390/v10080420
Mock DJ, Frampton MW, Nichols JE, Domurat FM, Signs DJ, Roberts NJ Jr. Influenza Virus Infection of Human Lymphocytes Occurs in the Immune Cell Cluster of the Developing Antiviral Response. Viruses. 2018; 10(8):420. https://doi.org/10.3390/v10080420
Chicago/Turabian StyleMock, David J., Mark W. Frampton, Joan E. Nichols, Frank M. Domurat, Denise J. Signs, and Norbert J. Roberts, Jr. 2018. "Influenza Virus Infection of Human Lymphocytes Occurs in the Immune Cell Cluster of the Developing Antiviral Response" Viruses 10, no. 8: 420. https://doi.org/10.3390/v10080420
APA StyleMock, D. J., Frampton, M. W., Nichols, J. E., Domurat, F. M., Signs, D. J., & Roberts, N. J., Jr. (2018). Influenza Virus Infection of Human Lymphocytes Occurs in the Immune Cell Cluster of the Developing Antiviral Response. Viruses, 10(8), 420. https://doi.org/10.3390/v10080420