Maternal Immunity and the Natural History of Congenital Human Cytomegalovirus Infection


Abstract
1. Introduction
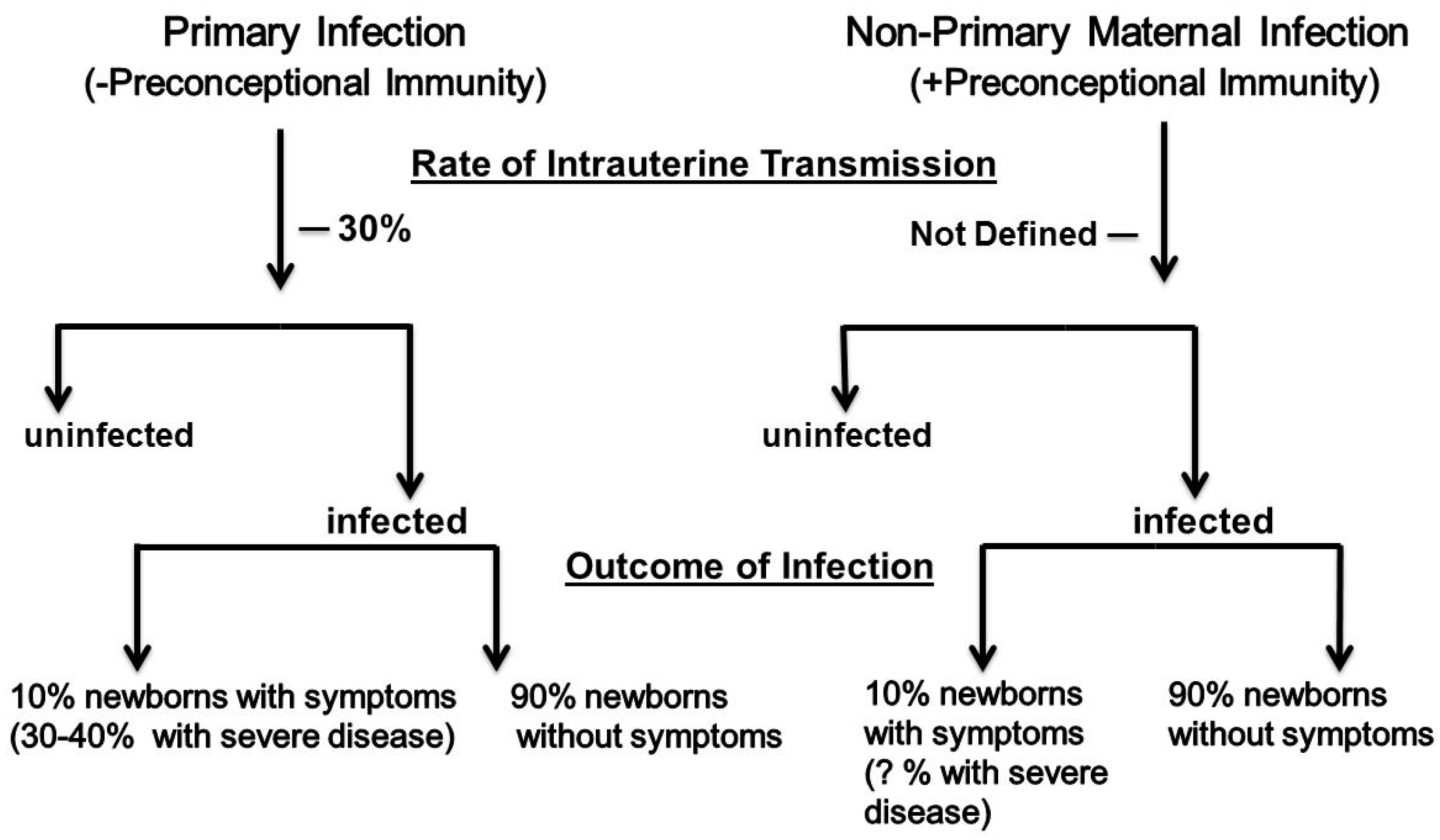
2. Epidemiology of cCMV Infections
3. Evidence That Adaptive Immunity Can Modify but not Prevent HCMV Infections
4. Adaptive Antiviral Responses and Intrauterine Transmission of HCMV
5. Adaptive Antiviral Responses and the Severity and Long Term Outcome of cCMV Infection
6. The Impact of Adaptive Immunity on cCMV Infections: Lessons from Vaccine Trials
7. Conclusions
Funding
Conflicts of Interest
References
- Boppana, S.B.; Ross, S.A.; Shimamura, M.; Palmer, A.L.; Ahmed, A.; Michaels, M.G.; Sanchez, P.J.; Bernstein, D.I.; Tolan, R.W., Jr.; Novak, Z.; et al. Saliva polymerase-chain-reaction assay for cytomegalovirus screening in newborns. N. Engl. J. Med. 2011, 364, 2111–2118. [Google Scholar] [CrossRef] [PubMed]
- Mussi-Pinhata, M.M.; Yamamoto, A.Y.; Moura Brito, R.M.; de Lima Isaac, M.; de Carvalho e Oliveira, P.F.; Boppana, S.; Britt, W.J. Birth prevalence and natural history of congenital cytomegalovirus infection in a highly seroimmune population. Clin. Infect. Dis. 2009, 49, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Manicklal, S.; Emery, V.C.; Lazzarotto, T.; Boppana, S.B.; Gupta, R.K. The “silent” global burden of congenital cytomegalovirus. Clin. Microbiol. Rev. 2013, 26, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Kenneson, A.; Cannon, M.J. Review and meta-analysis of the epidemiology of congenital cytomegalovirus (CMV) infection. Rev. Med. Virol. 2007, 17, 253–276. [Google Scholar] [CrossRef] [PubMed]
- Olusanya, B.O.; Slusher, T.M.; Boppana, S.B. Prevalence of congenital cytomegalovirus infection in Nigeria: A pilot study. Pediatr. Infect. Dis. J. 2015, 34, 322–324. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, N.G.; Yamamoto, A.Y.; Duarte, G.; Aragon, D.C.; Fowler, K.B.; Boppana, S.; Britt, W.J.; Mussi-Pinhata, M.M. Cytomegalovirus (CMV) Shedding in Seropositive Pregnant Women from a High Seroprevalence Population: “The Brazilian Cytomegalovirus Hearing and Maternal Secondary Infection Study” (BraCHS). Clin. Infect. Dis. 2018. [Google Scholar] [CrossRef] [PubMed]
- Fowler, K.B.; Ross, S.A.; Shimamura, M.; Ahmed, A.; Palmer, A.L.; Michaels, M.G.; Bernstein, D.I.; Sanchez, P.J.; Feja, K.N.; Stewart, A.; et al. Racial and Ethnic Differences in the Prevalence of Congenital Cytomegalovirus Infection. J. Pediatr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Alford, C.A.; Pass, R.F.; Stagno, S. Chronic congenital infections: Common environmental causes for severe and subtle birth defects. Birth Defects Orig. Artic. Ser. 1983, 19, 87–96. [Google Scholar] [PubMed]
- Britt, W. Controversies in the natural history of congenital human cytomegalovirus infection: The paradox of infection and disease in offspring of women with immunity prior to pregnancy. Med. Microbiol. Immunol. 2015, 204, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Cutts, F.T.; Vynnycky, E. Modelling the incidence of congenital rubella syndrome in developing countries. Int. J. Epidemiol. 1999, 28, 1176–1184. [Google Scholar] [CrossRef] [PubMed]
- Cooper, L.Z.; Preblud, S.R.; Alford, C.A. Infectious Diseases of the Fetus and Newborn Infant, 4th ed.; Remington, J.S., Klein, J.O., Eds.; WB Saunders: Philadelphia, PA, USA, 1995; pp. 268–311. [Google Scholar]
- Netto, E.M.; Moreira-Soto, A.; Pedroso, C.; Höser, C.; Funk, S.; Kucharski, A.J.; Rockstroh, A.; Kümmerer, B.M.; Sampaio, G.S.; Luz, E.; et al. High Zika Virus Seroprevalence in Salvador, Northeastern Brazil Limits the Potential for Further Outbreaks. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Stagno, S.; Cloud, G.; Pass, R.F.; Britt, W.J.; Alford, C.A. Factors associated with primary cytomegalovirus infection during pregnancy. J. Med. Virol. 1984, 13, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Fowler, K.B.; Stagno, S.; Pass, R.F. Maternal age and congenital cytomegalovirus infection: Screening of two diverse newborn populations, 1980–1990. J. Infect. Dis. 1993, 168, 552–556. [Google Scholar] [CrossRef] [PubMed]
- Staras, S.A.; Flanders, W.D.; Dollard, S.C.; Pass, R.F.; McGowan, J.E., Jr.; Cannon, M.J. Cytomegalovirus seroprevalence and childhood sources of infection: A population-based study among pre-adolescents in the United States. J. Clin. Virol. 2008, 43, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Drew, W.L.; Mintz, L.; Miner, R.C.; Sands, M.; Ketterer, B. Prevalence of cytomegalovirus infection in homosexual men. J. Infect. Dis. 1981, 143, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Collier, A.C.; Handsfield, H.H.; Ashley, R.; Roberts, P.L.; DeRouen, T.; Meyers, J.D.; Corey, L. Cervical but not urinary excretion of cytomegalovirus is related to sexual activity and contraceptive practices in sexually active women. J. Infect. Dis. 1995, 171, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Chandler, S.H.; Holmes, K.K.; Wentworth, B.B.; Gutman, L.T.; Wiesner, P.J.; Alexander, E.R.; Handsfield, H.H. The epidemiology of cytomegaloviral infection in women attending a sexually transmitted disease clinic. J. Infect. Dis. 1985, 152, 597–605. [Google Scholar] [CrossRef] [PubMed]
- Remis, R.S.; Liu, J.; Loutfy, M.R.; Tharao, W.; Rebbapragada, A.; Huibner, S.; Kesler, M.; Halpenny, R.; Grennan, T.; Brunetta, J.; et al. Prevalence of Sexually Transmitted Viral and Bacterial Infections in HIV-Positive and HIV-Negative Men Who Have Sex with Men in Toronto. PLoS ONE 2016, 11, e0158090. [Google Scholar] [CrossRef] [PubMed]
- Dreher, A.M.; Arora, N.; Fowler, K.B.; Novak, Z.; Britt, W.J.; Boppana, S.B.; Ross, S.A. Spectrum of Disease and Outcome in Children with Symptomatic Congenital Cytomegalovirus Infection. J. Pediatr. 2014, 164, 855–859. [Google Scholar] [CrossRef] [PubMed]
- Boppana, S.; Britt, W.J. Synopsis of clinical aspects of human cytomegalovirus disease. In Cytomegaloviruses: From Molecular Pathogenesis to Intervention; Reddehase, M., Ed.; Casister Academic Press: Norfolk, UK, 2013; Volume 2, pp. 1–25. [Google Scholar]
- Britt, W.J. Congenital Human Cytomegalovirus Infection and the Enigma of Maternal Immunity. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Fowler, K.B. Congenital cytomegalovirus infection: Audiologic outcome. Clin. Infect. Dis. 2013, 57 (Suppl. 4), S182–S184. [Google Scholar] [CrossRef]
- Yamamoto, A.Y.; Mussi-Pinhata, M.M.; Isaac Mde, L.; Amaral, F.R.; Carvalheiro, C.G.; Aragon, D.C.; Manfredi, A.K.; Boppana, S.B.; Britt, W.J. Congenital cytomegalovirus infection as a cause of sensorineural hearing loss in a highly immune population. Pediatr. Infect. Dis. J. 2011, 30, 1043–1046. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, S.; Yamashita, M.; Maeda, M.; Misawa, K.; Mineta, H. Audiological outcome of infants with congenital cytomegalovirus infection in a prospective study. Audiol. Neurootol. 2007, 12, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Lanzieri, T.M.; Chung, W.; Flores, M.; Blum, P.; Caviness, A.C.; Bialek, S.R.; Grosse, S.D.; Miller, J.A.; Demmler-Harrison, G. Hearing Loss in Children With Asymptomatic Congenital Cytomegalovirus Infection. Pediatrics 2017, 139, e20162610. [Google Scholar] [CrossRef] [PubMed]
- Morton, C.C.; Nance, W.E. Newborn hearing screening—A silent revolution. N. Engl. J. Med. 2006, 354, 2151–2164. [Google Scholar] [CrossRef] [PubMed]
- Stagno, S.; Pass, R.F.; Cloud, G.; Britt, W.J.; Henderson, R.E.; Walton, P.D.; Veren, D.A.; Page, F.; Alford, C.A. Primary cytomegalovirus infection in pregnancy. Incidence, transmission to fetus, and clinical outcome. JAMA 1986, 256, 1904–1908. [Google Scholar] [CrossRef] [PubMed]
- Manicklal, S.; van Niekerk, A.M.; Kroon, S.M.; Hutto, C.; Novak, Z.; Pati, S.K.; Chowdhury, N.; Hsiao, N.Y.; Boppana, S.B. Birth prevalence of congenital cytomegalovirus among infants of HIV-infected women on prenatal antiretroviral prophylaxis in South Africa. Clin. Infect. Dis. 2014, 58, 1467–1472. [Google Scholar] [CrossRef] [PubMed]
- Mwaanza, N.; Chilukutu, L.; Tembo, J.; Kabwe, M.; Musonda, K.; Kapasa, M.; Chabala, C.; Sinyangwe, S.; Mwaba, P.; Zumla, A.; et al. High rates of congenital cytomegalovirus infection linked with maternal HIV infection among neonatal admissions at a large referral center in sub-Saharan Africa. Clin. Infect. Dis. 2014, 58, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Adachi, K.; Xu, J.; Ank, B.; Watts, D.H.; Mofenson, L.M.; Pilotto, J.H.; Joao, E.; Santos, B.; Fonseca, R.; Kreitchmann, R.; et al. Cytomegalovirus Urinary Shedding in HIV-infected Pregnant Women and Congenital Cytomegalovirus Infection. Clin. Infect. Dis. 2017, 65, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Enders, G.; Daiminger, A.; Bader, U.; Exler, S.; Enders, M. Intrauterine transmission and clinical outcome of 248 pregnancies with primary cytomegalovirus infection in relation to gestational age. J. Clin. Virol. 2011, 52, 244–246. [Google Scholar] [CrossRef] [PubMed]
- Delforge, M.L.; Costa, E.; Brancart, F.; Goldman, D.; Montesinos, I.; Zaytouni, S.; Marchant, A.; Donner, C. Presence of Cytomegalovirus in urine and blood of pregnant women with primary infection might be associated with fetal infection. J. Clin. Virol. 2017, 90, 14–17. [Google Scholar] [CrossRef] [PubMed]
- Simonazzi, G.; Curti, A.; Cervi, F.; Gabrielli, L.; Contoli, M.; Capretti, M.G.; Rizzo, N.; Guerra, B.; Farina, A.; Lazzarotto, T. Perinatal Outcomes of Non-Primary Maternal Cytomegalovirus Infection: A 15-Year Experience. Fetal Diagn. Ther. 2018, 43, 138–142. [Google Scholar] [CrossRef] [PubMed]
- Mele, F.; Fornara, C.; Jarrossay, D.; Furione, M.; Arossa, A.; Spinillo, A.; Lanzavecchia, A.; Gerna, G.; Sallusto, F.; Lilleri, D. Phenotype and specificity of T cells in primary human cytomegalovirus infection during pregnancy: IL-7Rpos long-term memory phenotype is associated with protection from vertical transmission. PLoS ONE 2017, 12, e0187731. [Google Scholar] [CrossRef] [PubMed]
- Lilleri, D.; Gerna, G.; Furione, M.; Zavattoni, M.; Spinillo, A. Neutralizing and ELISA IgG antibodies to human cytomegalovirus glycoprotein complexes may help date the onset of primary infection in pregnancy. J. Clin. Virol. 2016, 81, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Leruez-Ville, M.; Magny, J.F.; Couderc, S.; Pichon, C.; Parodi, M.; Bussieres, L.; Guilleminot, T.; Ghout, I.; Ville, Y. Risk Factors for Congenital Cytomegalovirus Infection Following Primary and Nonprimary Maternal Infection: A Prospective Neonatal Screening Study Using Polymerase Chain Reaction in Saliva. Clin. Infect. Dis. 2017, 65, 398–404. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, K.; Tairaku, S.; Morioka, I.; Ozaki, K.; Nagamata, S.; Morizane, M.; Deguchi, M.; Ebina, Y.; Minematsu, T.; Yamada, H. Universal Screening With Use of Immunoglobulin G Avidity for Congenital Cytomegalovirus Infection. Clin. Infect. Dis. 2017, 65, 1652–1658. [Google Scholar] [CrossRef] [PubMed]
- Ahlfors, K.; Ivarsson, S.A.; Harris, S.; Svanberg, L.; Holmqvist, R.; Lernmark, B.; Theander, G. Congenital cytomegalovirus infection and disease in Sweden and the relative importance of primary and secondary maternal infections. Scand. J. Infect. Dis. 1984, 16, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, X.; Bialek, S.; Cannon, M.J. Attribution of congenital cytomegalovirus infection to primary versus non-primary maternal infection. Clin. Infect. Dis. 2011, 52, e11–e13. [Google Scholar] [CrossRef] [PubMed]
- Ahlfors, K.; Ivarsson, S.A.; Harris, S. Report on a long-term study of maternal and congenital cytomegalovirus infection in Sweden. Review of prospective studies available in the literature. Scand. J. Infect. Dis. 1999, 31, 443–457. [Google Scholar] [PubMed]
- Townsend, C.L.; Forsgren, M.; Ahlfors, K.; Ivarsson, S.A.; Tookey, P.A.; Peckham, C.S. Long-term outcomes of congenital cytomegalovirus infection in Sweden and the United Kingdom. Clin. Infect. Dis. 2013, 56, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Dar, L.; Pati, S.K.; Patro, A.R.; Deorari, A.K.; Rai, S.; Kant, S.; Broor, S.; Fowler, K.B.; Britt, W.J.; Boppana, S.B. Congenital cytomegalovirus infection in a highly seropositive semi-urban population in India. Pediatr. Infect. Dis. J. 2008, 27, 841–843. [Google Scholar] [CrossRef] [PubMed]
- Boppana, S.B.; Fowler, K.B.; Britt, W.J.; Stagno, S.; Pass, R.F. Symptomatic congenital cytomegalovirus infection in infants born to mothers with preexisting immunity to cytomegalovirus. Pediatrics 1999, 104, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Boppana, S.B.; Rivera, L.B.; Fowler, K.B.; Mach, M.; Britt, W.J. Intrauterine transmission of cytomegalovirus to infants of women with preconceptional immunity. N. Engl. J. Med. 2001, 344, 1366–1371. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.A.; Fowler, K.B.; Ashrith, G.; Stagno, S.; Britt, W.J.; Pass, R.F.; Boppana, S.B. Hearing loss in children with congenital cytomegalovirus infection born to mothers with preexisting immunity. J. Pediatr. 2006, 148, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, A.Y.; Mussi-Pinhata, M.M.; Boppana, S.B.; Novak, Z.; Wagatsuma, V.M.; Oliveira Pde, F.; Duarte, G.; Britt, W.J. Human cytomegalovirus reinfection is associated with intrauterine transmission in a highly cytomegalovirus-immune maternal population. Am. J. Obstet. Gynecol. 2010, 202, 297.e1–297.e8. [Google Scholar] [CrossRef] [PubMed]
- Ahlfors, K.; Ivarsson, S.A.; Harris, S. Secondary maternal cytomegalovirus infection—A significant cause of congenital disease. Pediatrics 2001, 107, 1227–1228. [Google Scholar] [CrossRef] [PubMed]
- Fowler, K.B.; Stagno, S.; Pass, R.F.; Britt, W.J.; Boll, T.J.; Alford, C.A. The outcome of congenital cytomegalovirus infection in relation to maternal antibody status. N. Engl. J. Med. 1992, 326, 663–667. [Google Scholar] [CrossRef] [PubMed]
- Rubin, R.H.; Russell, P.S.; Levin, M.; et al. Summary of a workshop on cytomegalovirus infections during organ transplantation. J. Infect. Dis. 1979, 139, 728–734. [Google Scholar] [CrossRef] [PubMed]
- Autran, B.; Carcelain, G.; Li, T.S.; Blanc, C.; Mathez, D.; Tubiana, R.; Katlama, C.; Debre, P.; Leibowitch, J. Positive effects of combined antiretroviral therapy on CD4+ T cell homeostasis and function in advanced HIV disease. Science 1997, 277, 112–116. [Google Scholar] [CrossRef] [PubMed]
- Komanduri, K.V.; Viswanathan, M.N.; Wieder, E.D.; Schmidt, D.K.; Bredt, B.M.; Jacobson, M.; McCune, J.M. Restoration of cytomegalovirus-specific CD4+ T-lymphocyte responses after ganciclovir and highly active antiretroviral therapy in individuals infected with HIV-1. Nat. Med. 1998, 4, 953–956. [Google Scholar] [CrossRef] [PubMed]
- Bronke, C.; Palmer, N.M.; Jansen, C.A.; Westerlaken, G.H.; Polstra, A.M.; Reiss, P.; Bakker, M.; Miedema, F.; Tesselaar, K.; van Baarle, D. Dynamics of cytomegalovirus (CMV)-specific T cells in HIV-1-infected individuals progressing to AIDS with CMV end-organ disease. J. Infect. Dis. 2005, 191, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Gerna, G.; Lilleri, D.; Chiesa, A.; Zelini, P.; Furione, M.; Comolli, G.; Pellegrini, C.; Sarchi, E.; Migotto, C.; Bonora, M.R.; et al. Virologic and immunologic monitoring of cytomegalovirus to guide preemptive therapy in solid-organ transplantation. Am. J. Transplant. 2011, 11, 2463–2471. [Google Scholar] [CrossRef] [PubMed]
- Gratama, J.W.; Boeckh, M.; Nakamura, R.; Cornelissen, J.J.; Brooimans, R.A.; Zaia, J.A.; Forman, S.J.; Gaal, K.; Bray, K.R.; Gasior, G.H.; et al. Immune monitoring with iT Ag MHC tetramers for prediction of recurrent or persistent cytomegalovirus infection or disease in allogeneic hematopoietic stem cell transplant recipients: A prospective multicenter study. Blood 2010, 116, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Lucia, M.; Crespo, E.; Melilli, E.; Cruzado, J.M.; Luque, S.; Llaudo, I.; Niubo, J.; Torras, J.; Fernandez, N.; Grinyo, J.M.; et al. Preformed frequencies of cytomegalovirus (CMV)-specific memory T and B cells identify protected CMV-sensitized individuals among seronegative kidney transplant recipients. Clin. Infect. Dis. 2014, 59, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Ganepola, S.; Gentilini, C.; Hilbers, U.; Lange, T.; Rieger, K.; Hofmann, J.; Maier, M.; Liebert, U.G.; Niederwieser, D.; Engelmann, E.; et al. Patients at high risk for CMV infection and disease show delayed CD8+ T-cell immune recovery after allogeneic stem cell transplantation. Bone Marrow Transpl. 2007, 39, 293–299. [Google Scholar] [CrossRef] [PubMed]
- Reusser, P.; Riddell, S.R.; Meyers, J.D.; Greenberg, P.D. Cytotoxic T-lymphocyte response to cytomegalovirus after human allogeneic bone marrow transplantation: Pattern of recovery and correlation with cytomegalovirus infection and disease. Blood 1991, 78, 1373–1380. [Google Scholar] [PubMed]
- Li, C.R.; Greenberg, P.D.; Gilbert, M.J.; Goodrich, J.M.; Riddell, S.R. Recovery of HLA-restricted cytomegalovirus (CMV)-specific T-cell responses after allogeneic bone marrow transplant: Correlation with CMV disease and effect of ganciclovir prophylaxis. Blood 1994, 83, 1971–1979. [Google Scholar] [PubMed]
- Egli, A.; Humar, A.; Kumar, D. State-of-the-art monitoring of cytomegalovirus-specific cell-mediated immunity after organ transplant: A primer for the clinician. Clin. Infect. Dis. 2012, 55, 1678–1689. [Google Scholar] [CrossRef] [PubMed]
- Gabanti, E.; Lilleri, D.; Ripamonti, F.; Bruno, F.; Zelini, P.; Furione, M.; Colombo, A.A.; Alessandrino, E.P.; Gerna, G. Reconstitution of Human Cytomegalovirus-Specific CD4+ T Cells is Critical for Control of Virus Reactivation in Hematopoietic Stem Cell Transplant Recipients but Does Not Prevent Organ Infection. Biol. Blood Marrow Transpl. 2015, 21, 2192–2202. [Google Scholar] [CrossRef] [PubMed]
- Walter, E.A.; Greenberg, P.D.; Gilbert, M.J.; Finch, R.J.; Watanabe, K.S.; Thomas, E.D.; Riddell, S.R. Reconstitution of cellular immunity against cytomegalovirus in recipients of allogeneic bone marrow by transfer of T-cell clones from the donor. N. Engl. J. Med. 1995, 333, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Snydman, D.R.; Werner, B.G.; Heinze-Lacey, B.; Berardi, V.P.; Tilney, N.L.; Kirkman, R.L.; Milford, E.L.; Cho, S.I.; Bush, H.L., Jr.; Levey, A.S. Use of cytomegalovirus immune globulin to prevent cytomegalovirus disease in renal-transplant recipients. N. Engl. J. Med. 1987, 317, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Snydman, D.R.; Ruthazer, R.; Griffith, J.; Werner, B.G.; Freeman, R.; Rohrer, R. Cytomegalovirus immune globulin (CMVIG) prophylaxis is associated with increased survival after orthotopic liver transplantation. The Boston Center for Liver Transplantation CMVIG Study Group. Clin. Transplant. 1997, 11, 432–437. [Google Scholar] [PubMed]
- Hibberd, P.L.; Tolkoff-Rubin, N.E.; Conti, D.; Stuart, F.; Thistlethwaite, J.R.; Neylan, J.F.; Snydman, D.R.; Freeman, R.; Lorber, M.I.; Rubin, R.H. Preemptive ganciclovir therapy to prevent cytomegalovirus disease in cytomegalovirus antibody-positive renal transplant recipients. A randomized controlled trial. Ann. Intern. Med. 1995, 123, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Ishida, J.H.; Patel, A.; Mehta, A.K.; Gatault, P.; McBride, J.M.; Burgess, T.; Derby, M.A.; Snydman, D.R.; Emu, B.; Feierbach, B.; et al. Phase 2 Randomized, Double-Blind, Placebo-Controlled Trial of RG7667, a Combination Monoclonal Antibody, for Prevention of Cytomegalovirus Infection in High-Risk Kidney Transplant Recipients. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Biron, C.A.; Byron, K.S.; Sullivan, J.L. Severe herpesvirus infections in an adolescent without natural killer cells. N. Engl. J. Med. 1989, 320, 1731–1735. [Google Scholar] [CrossRef] [PubMed]
- Barry, P.A. Exploiting viral natural history for vaccine development. Med. Microbiol. Immunol. 2015, 204, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Deere, J.D.; Barry, P.A. Using the nonhuman primate model of HCMV to guide vaccine development. Viruses 2014, 6, 1483–1501. [Google Scholar] [CrossRef] [PubMed]
- Wussow, F.; Yue, Y.; Martinez, J.; Deere, J.D.; Longmate, J.; Herrmann, A.; Barry, P.A.; Diamond, D.J. A vaccine based on the rhesus cytomegalovirus UL128 complex induces broadly neutralizing antibodies in rhesus macaques. J. Virol. 2013, 87, 1322–1332. [Google Scholar] [CrossRef] [PubMed]
- Abel, K.; Martinez, J.; Yue, Y.; Lacey, S.F.; Wang, Z.; Strelow, L.; Dasgupta, A.; Li, Z.; Schmidt, K.A.; Oxford, K.L.; et al. Vaccine-induced control of viral shedding following rhesus cytomegalovirus challenge in rhesus macaques. J. Virol. 2011, 85, 2878–2890. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.R.; Bialek, S.R.; Boppana, S.B.; Griffiths, P.D.; Laughlin, C.A.; Ljungman, P.; Mocarski, E.S.; Pass, R.F.; Read, J.S.; Schleiss, M.R.; et al. Priorities for CMV vaccine development. Vaccine 2013, 32, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Streblow, D.N.; Hwee, Y.K.; Kreklywich, C.N.; Andoh, T.; Denton, M.; Smith, P.; Hart, E.; Broekel, R.; Pallett, C.; Rogers, K.; et al. Rat Cytomegalovirus Vaccine Prevents Accelerated Chronic Rejection in CMV-Naive Recipients of Infected Donor Allograft Hearts. Am. J. Transpl. 2015, 15, 1805–1816. [Google Scholar] [CrossRef] [PubMed]
- Brizic, I.; Hirsl, L.; Britt, W.J.; Krmpotic, A.; Jonjic, S. Immune responses to congenital cytomegalovirus infection. Microbes Infect. 2017. [Google Scholar] [CrossRef] [PubMed]
- Slavuljica, I.; Busche, A.; Babic, M.; Mitrovic, M.; Gasparovic, I.; Cekinovic, D.; Markova Car, E.; Pernjak Pugel, E.; Cikovic, A.; Lisnic, V.J.; et al. Recombinant mouse cytomegalovirus expressing a ligand for the NKG2D receptor is attenuated and has improved vaccine properties. J. Clin. Investig. 2010, 120, 4532–4545. [Google Scholar] [CrossRef] [PubMed]
- Holtappels, R.; Podlech, J.; Grzimek, N.K.; Thomas, D.; Pahl-Seibert, M.F.; Reddehase, M.J. Experimental preemptive immunotherapy of murine cytomegalovirus disease with CD8 T-cell lines specific for ppM83 and pM84, the two homologs of human cytomegalovirus tegument protein ppUL83 (pp65). J. Virol. 2001, 75, 6584–6600. [Google Scholar] [CrossRef] [PubMed]
- Holtappels, R.; Bohm, V.; Podlech, J.; Reddehase, M.J. CD8 T-cell-based immunotherapy of cytomegalovirus infection: “proof of concept” provided by the murine model. Med. Microbiol. Immunol. 2008, 197, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Podlech, J.; Holtappels, R.; Pahl-Seibert, M.F.; Steffens, H.P.; Reddehase, M.J. Murine model of interstitial cytomegalovirus pneumona in syngenic bone marrow transplantation: Persistence of protective pulmonary CD8-T-cell infiltrates after clearance of acute infection. J. Virol. 2000, 74, 7496–7507. [Google Scholar] [CrossRef] [PubMed]
- Sequar, G.; Britt, W.J.; Lakeman, F.D.; Lockridge, K.M.; Tarara, R.P.; Canfield, D.R.; Zhou, S.S.; Gardner, M.B.; Barry, P.A. Experimental coinfection of rhesus macaques with rhesus cytomegalovirus and simian immunodeficiency virus: Pathogenesis. J. Virol. 2002, 76, 7661–7671. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.G.; Piatak, M., Jr.; Ventura, A.B.; Hughes, C.M.; Gilbride, R.M.; Ford, J.C.; Oswald, K.; Shoemaker, R.; Li, Y.; Lewis, M.S.; et al. Immune clearance of highly pathogenic SIV infection. Nature 2013, 502, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Hansen, S.G.; Zak, D.E.; Xu, G.; Ford, J.C.; Marshall, E.E.; Malouli, D.; Gilbride, R.M.; Hughes, C.M.; Ventura, A.B.; Ainslie, E.; et al. Prevention of tuberculosis in rhesus macaques by a cytomegalovirus-based vaccine. Nat. Med. 2018, 24, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Woolf, N.K.; Jaquish, D.V.; Koehrn, F.J. Transplacental murine cytomegalovirus infection in the brain of SCID mice. Virol. J. 2007, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Bialas, K.M.; Tanaka, T.; Tran, D.; Varner, V.; Cisneros de la Rosa, E.; Chiuppesi, F.; Wussow, F.; Kattenhorn, L.; Macri, S.; Kunz, E.L.; et al. Maternal CD4+ T cells protect against severe congenital cytomegalovirus disease in a novel nonhuman primate model of placental cytomegalovirus transmission. Proc. Natl. Acad. Sci. USA 2015, 112, 13645–13650. [Google Scholar] [CrossRef] [PubMed]
- Griffith, B.P.; McCormick, S.R.; Booss, J.; Hsiung, G.D. Inbred guinea pig model of intrauterine infection with cytomegalovirus. Am. J. Pathol. 1986, 122, 112–119. [Google Scholar] [PubMed]
- Bia, F.J.; Miller, S.A.; Davidson, K.H. The guinea pig cytomegalovirus model of congenital human cytomegalovirus infection. Birth Defects 1984, 20, 233–241. [Google Scholar] [PubMed]
- Harrison, C.J.; Britt, W.J.; Chapan, N.M.; Mullican, J.; Tracy, S. Reduced congenital cytomegalovirus (CMV) infection after maternal immunization with a guinea pig CMV glycoprotein before gestational primary CMV infection in the guinea pig model. J. Infect. Dis. 1995, 172, 1212–1220. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.Y.; Root, M.; McGregor, A. A Novel Non-Replication-Competent Cytomegalovirus Capsid Mutant Vaccine Strategy Is Effective in Reducing Congenital Infection. J. Virol. 2016, 90, 7902–7919. [Google Scholar] [CrossRef] [PubMed]
- Schleiss, M.R.; Berka, U.; Watson, E.; Aistleithner, M.; Kiefmann, B.; Mangeat, B.; Swanson, E.C.; Gillis, P.A.; Hernandez-Alvarado, N.; Fernandez-Alarcon, C.; et al. Additive Protection against Congenital Cytomegalovirus Conferred by Combined Glycoprotein B/pp65 Vaccination Using a Lymphocytic Choriomeningitis Virus Vector. Clin. Vaccine Immunol. 2017, 24, e00300-16. [Google Scholar] [CrossRef] [PubMed]
- Swanson, E.C.; Gillis, P.; Hernandez-Alvarado, N.; Fernandez-Alarcon, C.; Schmit, M.; Zabeli, J.C.; Wussow, F.; Diamond, D.J.; Schleiss, M.R. Comparison of monovalent glycoprotein B with bivalent gB/pp65 (GP83) vaccine for congenital cytomegalovirus infection in a guinea pig model: Inclusion of GP83 reduces gB antibody response but both vaccine approaches provide equivalent protection against pup mortality. Vaccine 2015, 33, 4013–4018. [Google Scholar] [PubMed]
- Borune, N.; Schleiss, M.; Bravo, F.; Bernstein, D. Preconception immunization with a cytomegalovirus (CMV) glycoprotein vaccine improves pregnancy outcome in a guniea pig model of congenital CMV infection. J. Infect. Dis. 2001, 183, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Schleiss, M.R.; Bourne, N.; Stroup, G.; Bravo, F.J.; Jensen, N.J.; Bernstein, D.I. Protection against congenital cytomegalovirus infection and disease in guinea pigs, conferred by a purified recombinant glycoprotein B vaccine. J. Infect. Dis. 2004, 189, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, M.R.; Yan, D.; Vij, R.; Hongo, J.A.; Nakamura, G.; Vernes, J.M.; Meng, Y.G.; Lein, S.; Chan, P.; Ross, J.; et al. A neutralizing anti-gH/gL monoclonal antibody is protective in the guinea pig model of congenital CMV infection. PLoS Pathog. 2014, 10, e1004060. [Google Scholar] [CrossRef] [PubMed]
- Bratcher, D.F.; Bourne, N.; Bravo, F.J.; Schleiss, M.R.; Slaoui, M.; Myers, M.G.; Bernstein, D.I. Effect of passive antibody on congenital cytomegalovirus infection in guinea pigs. J. Infect. Dis. 1995, 172, 944–950. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, A.; Harrison, C.J.; Britt, W.J.; Bewtra, C. Modification of maternal and congenital cytomegalovirus infection by anti-glycoprotein b antibody transfer in guinea pigs. J. Infect. Dis. 2001, 183, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, K.; Yamada, S.; Katano, H.; Fukuchi, S.; Sato, Y.; Kato, M.; Yamaguchi, T.; Moriishi, K.; Inoue, N. Effects of immunization of pregnant guinea pigs with guinea pig cytomegalovirus glycoprotein B on viral spread in the placenta. Vaccine 2013, 31, 3199–3205. [Google Scholar] [CrossRef] [PubMed]
- Revello, M.G.; Lazzarotto, T.; Guerra, B.; Spinillo, A.; Ferrazzi, E.; Kustermann, A.; Guaschino, S.; Vergani, P.; Todros, T.; Frusca, T.; et al. A randomized trial of hyperimmune globulin to prevent congenital cytomegalovirus. N. Engl. J. Med. 2014, 370, 1316–1326. [Google Scholar] [CrossRef] [PubMed]
- Blazquez-Gamero, D.; Galindo Izquierdo, A.; Del Rosal, T.; Baquero-Artigao, F.; Izquierdo Mendez, N.; Soriano-Ramos, M.; Rojo Conejo, P.; Gonzalez-Tome, M.I.; Garcia-Burguillo, A.; Perez Perez, N.; et al. Prevention and treatment of fetal cytomegalovirus infection with cytomegalovirus hyperimmune globulin: A multicenter study in Madrid. J. Matern. Fetal Neonatal Med. 2017, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kagan, K.O.; Enders, M.; Schampera, M.S.; Baeumel, E.; Hoopmann, M.; Geipel, A.; Berg, C.; Goelz, R.; de Catte, L.; Wallwiener, D.; et al. Prevention of maternal-fetal transmission of CMV by hyperimmunoglobulin (HIG) administered after a primary maternal CMV infectionin early gestation. Ultrasound Obstet. Gynecol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Alford, C.A.; Hayes, K.; Britt, W. Primary cytomegalovirus infection in pregnancy: Comparison of antibody responses to virus-encoded proteins between women with and without intrauterine infection. J. Infect. Dis. 1988, 158, 917–924. [Google Scholar] [CrossRef] [PubMed]
- Lilleri, D.; Kabanova, A.; Revello, M.G.; Percivalle, E.; Sarasini, A.; Genini, E.; Sallusto, F.; Lanzavecchia, A.; Corti, D.; Gerna, G. Fetal human cytomegalovirus transmission correlates with delayed maternal antibodies to gH/gL/pUL128–130–131 complex during primary infection. PLoS ONE 2013, 8, e59863. [Google Scholar] [CrossRef] [PubMed]
- Boppana, S.B.; Britt, W.J. Antiviral antibody responses and intrauterine transmission after primary maternal cytomegalovirus infection. J. Infect. Dis. 1995, 171, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Lilleri, D.; Gerna, G. Maternal immune correlates of protection from human cytomegalovirus transmission to the fetus after primary infection in pregnancy. Rev. Med. Virol. 2017, 27. [Google Scholar] [CrossRef] [PubMed]
- Furione, M.; Rognoni, V.; Sarasini, A.; Zavattoni, M.; Lilleri, D.; Gerna, G.; Revello, M.G. Slow increase in IgG avidity correlates with prevention of human cytomegalovirus transmission to the fetus. J. Med. Virol. 2013, 85, 1960–1967. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.; Vanarsdall, A.; Chin, A.; Jardetzky, T.; Liu, J. (Eds.) Similar Levels of HCMV Pentamer- and Trimer-Specific Neutralizing Antibodies in Human Sera from Transplant Patients and Mothers and Neither Predict Outcome. In Proceedings of the International Herpesvirus Workshop, Vancouver, BC, Canada, 21–25 July 2018. [Google Scholar]
- Lilleri, D.; Fornara, C.; Furione, M.; Zavattoni, M.; Revello, M.G.; Gerna, G. Development of human cytomegalovirus-specific T cell immunity during primary infection of pregnant women and its correlation with virus transmission to the fetus. J. Infect. Dis. 2007, 195, 1062–1070. [Google Scholar] [CrossRef] [PubMed]
- Fornara, C.; Cassaniti, I.; Zavattoni, M.; Furione, M.; Adzasehoun, K.M.G.; de Silvestri, A.; Comolli, G.; Baldanti, F. Human Cytomegalovirus-Specific Memory CD4+ T-Cell Response and Its Correlation With Virus Transmission to the Fetus in Pregnant Women With Primary Infection. Clin. Infect. Dis. 2017, 65, 1659–1665. [Google Scholar] [CrossRef] [PubMed]
- Forner, G.; Saldan, A.; Mengoli, C.; Gussetti, N.; Palu, G.; Abate, D. Cytomegalovirus (CMV) Enzyme-Linked Immunosorbent Spot Assay but Not CMV QuantiFERON Assay Is a Novel Biomarker To Determine Risk of Congenital CMV Infection in Pregnant Women. J. Clin. Microbiol. 2016, 54, 2149–2154. [Google Scholar] [CrossRef] [PubMed]
- Stagno, S.; Pass, R.F.; Dworsky, M.E.; Henderson, R.E.; Moore, E.G.; Walton, P.D.; Alford, C.A. Congenital cytomegalovirus infection: The relative importance of primary and recurrent maternal infection. N. Engl. J. Med. 1982, 306, 945–949. [Google Scholar] [CrossRef] [PubMed]
- Yeager, A.S.; Arvin, A.M. Reasons for the absence of a history of recurrent genital infections in mothers of neonates infected with herpes simplex virus. Pediatrics 1984, 73, 188–193. [Google Scholar] [PubMed]
- Bootz, A.; Karbach, A.; Spindler, J.; Kropff, B.; Reuter, N.; Sticht, H.; Winkler, T.H.; Britt, W.J.; Mach, M. Protective capacity of neutralizing and non-neutralizing antibodies against glycoprotein B of cytomegalovirus. PLoS Pathog. 2017, 13, e1006601. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.S.; Cruz, D.V.; Tran, D.; Bialas, K.M.; Stamper, L.; Wu, H.; Gilbert, M.; Blair, R.; Alvarez, X.; Itell, H.; et al. Preexisting antibodies can protect against congenital cytomegalovirus infection in monkeys. JCI Insight 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Bravo, F.J.; Bourne, N.; Schleiss, M.R.; Bernstein, D.I. An animal model of neonatal cytomegalovirus infection. Antiviral Res. 2003, 60, 41–49. [Google Scholar] [CrossRef]
- Adler, S.P.; Starr, S.E.; Plotkin, S.A.; Hempfling, S.H.; Buis, J.; Manning, M.L.; Best, A.M. Immunity induced by primary human cytomegalovirus infection protects against secondary infection among women of childbearing age. J. Infect. Dis. 1995, 171, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Pass, R.F.; Zhang, C.; Evans, A.; Simpson, T.; Andrews, W.; Huang, M.L.; Corey, L.; Hill, J.; Davis, E.; Flanigan, C.; et al. Vaccine prevention of maternal cytomegalovirus infection. N. Engl. J. Med. 2009, 360, 1191–1199. [Google Scholar] [CrossRef] [PubMed]
- Sabbaj, S.; Pass, R.F.; Goepfert, P.A.; Pichon, S. Glycoprotein B vaccine is capable of boosting both antibody and CD4 T-cell responses to cytomegalovirus in chronically infected women. J. Infect. Dis. 2011, 203, 1534–1541. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, D.I.; Munoz, F.M.; Callahan, S.T.; Rupp, R.; Wootton, S.H.; Edwards, K.M.; Turley, C.B.; Stanberry, L.R.; Patel, S.M.; McNeal, M.M.; et al. Safety and efficacy of a cytomegalovirus glycoprotein B (gB) vaccine in adolescent girls: A randomized clinical trial. Vaccine 2016, 34, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.S.; Huffman, T.; Jenks, J.A.; Cisneros de la Rosa, E.; Xie, G.; Vandergrift, N.; Pass, R.F.; Pollara, J.; Permar, S.R. HCMV glycoprotein B subunit vaccine efficacy mediated by nonneutralizing antibody effector functions. Proc. Natl. Acad. Sci. USA 2018, 115, 6267–6272. [Google Scholar] [CrossRef] [PubMed]
- Kneiss, N.; Mach, M.; Fay, J.; Britt, W.J. Distribution of linear antigenic sites on glycoprotein gp55 of human cytomegalovirus. J. Virol. 1991, 65, 138–146. [Google Scholar]
- Urban, M.; Winkler, T.; Landini, M.P.; Britt, W.; Mach, M. Epitope-specific distribution of IgG subclasses against antigenic domains on glycoproteins of human cytomegalovirus. J. Infect. Dis. 1994, 169, 83–90. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Britt, W.J. Maternal Immunity and the Natural History of Congenital Human Cytomegalovirus Infection. Viruses 2018, 10, 405. https://doi.org/10.3390/v10080405
Britt WJ. Maternal Immunity and the Natural History of Congenital Human Cytomegalovirus Infection. Viruses. 2018; 10(8):405. https://doi.org/10.3390/v10080405
Chicago/Turabian StyleBritt, William J. 2018. "Maternal Immunity and the Natural History of Congenital Human Cytomegalovirus Infection" Viruses 10, no. 8: 405. https://doi.org/10.3390/v10080405
APA StyleBritt, W. J. (2018). Maternal Immunity and the Natural History of Congenital Human Cytomegalovirus Infection. Viruses, 10(8), 405. https://doi.org/10.3390/v10080405