There Is Always Another Way! Cytomegalovirus’ Multifaceted Dissemination Schemes


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
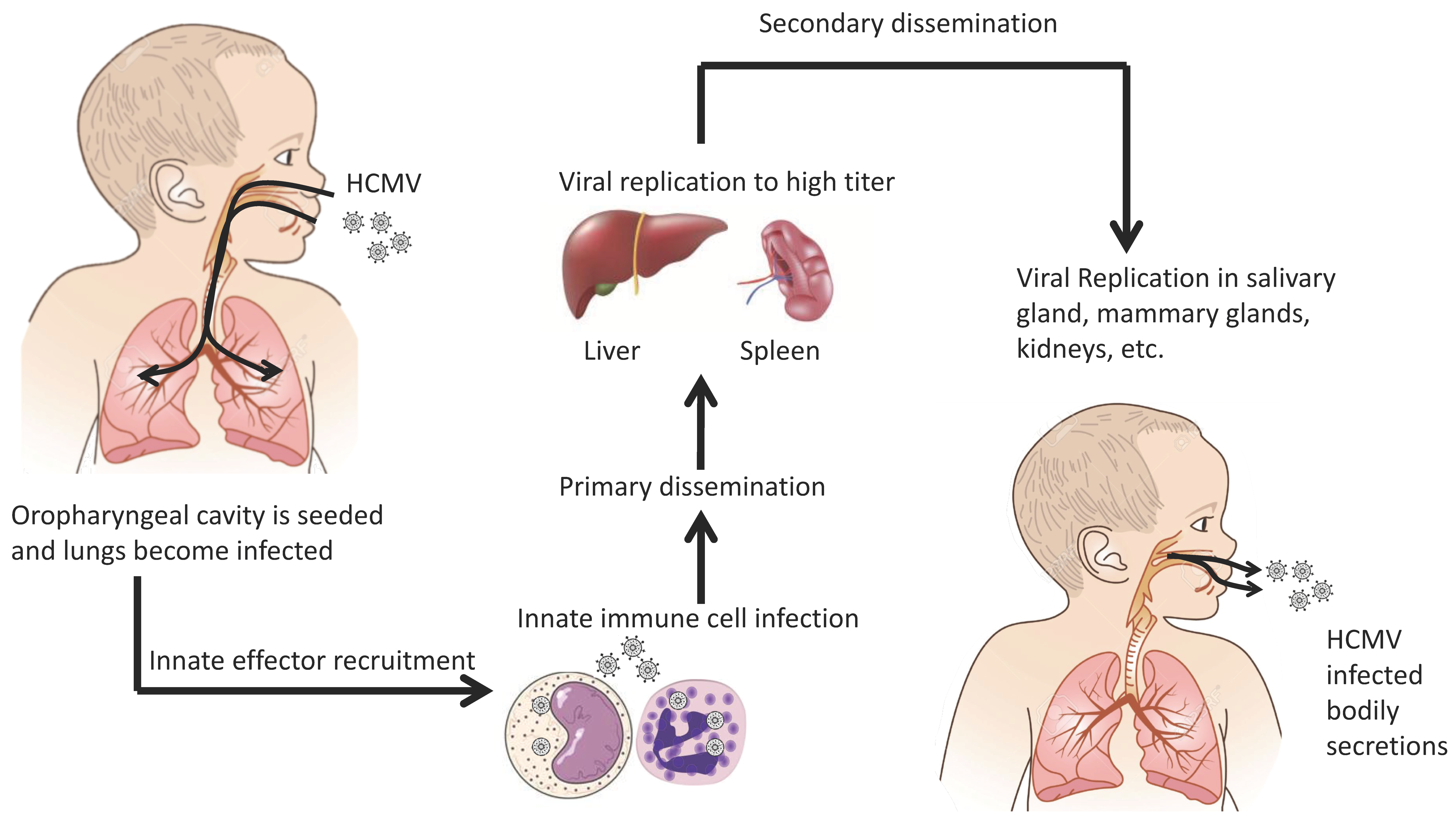
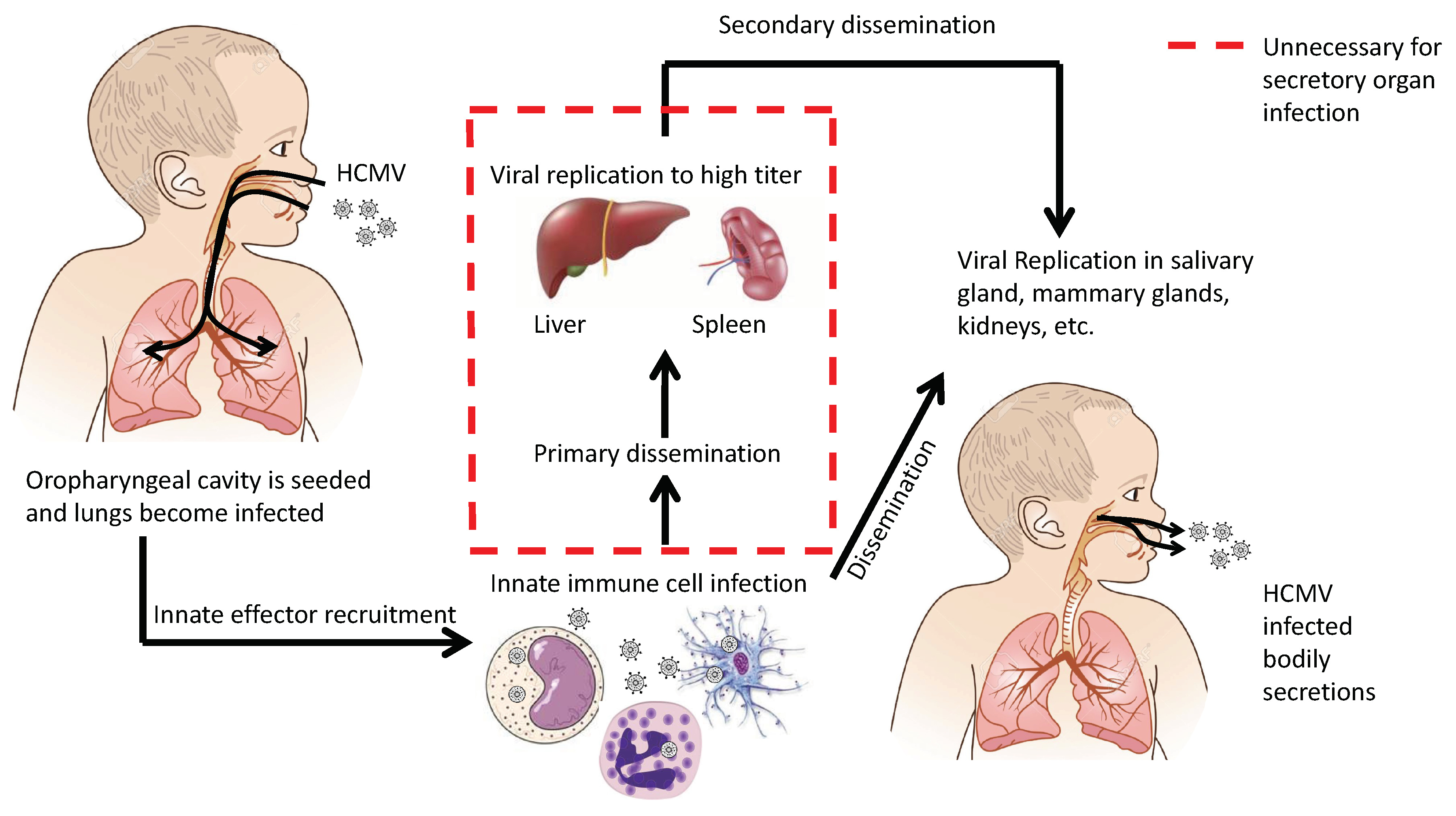
2. Transmission and Initial Infection
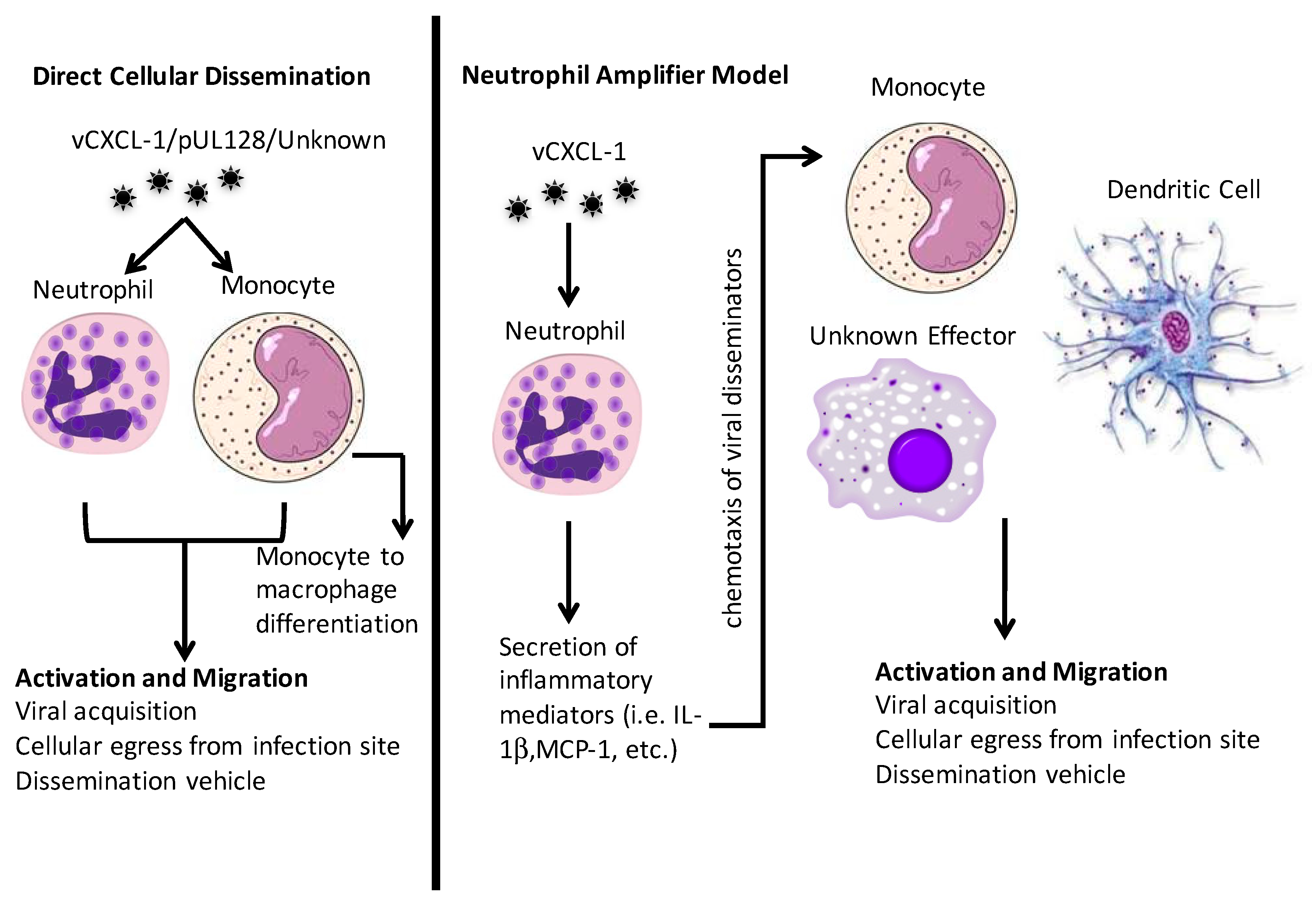
3. Cell-Mediated Dissemination
3.1. PMNs
3.2. Monocytes and Macrophages
4. Dissemination in Animal Models
4.1. The Mouse Model
4.2. Other Small Animal Models
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Staras, S.A.S.; Dollard, S.C.; Radford, K.W.; Flanders, W.D.; Pass, R.F.; Cannon, M.J. Seroprevalence of cytomegalovirus infection in the united states, 1988–1994. Clin. Infect. Dis. 2006, 43, 1143–1151. [Google Scholar] [CrossRef] [PubMed]
- Liu, L. Fields virology, 6th edition. Clin. Infect. Dis. 2014, 59, 613. [Google Scholar] [CrossRef]
- Britt, W. Virus entry into host, establishment of infection, spread in host, mechanisms of tissue damage. In Human Herpesviruses: Biology, Therapy, and Immunoprophylaxis; Arvin, A., Campadelli-Fiume, G., Mocarski, E., Moore, P.S., Roizman, B., Whitley, R., Yamanishi, K., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Li, F.; Kenyon, K.W.; Kirby, K.A.; Fishbein, D.P.; Boeckh, M.; Limaye, A.P. Incidence and clinical features of ganciclovir-resistant cytomegalovirus disease in heart transplant recipients. Clin. Infect. Dis. 2007, 45, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, J.; Sissons, P. Latency and reactivation of human cytomegalovirus. J. Gen. Virol. 2006, 87, 1763–1779. [Google Scholar] [CrossRef] [PubMed]
- Demmler, G.J. Congenital cytomegalovirus infection. Semin. Pediatr. Neurol. 1994, 1, 36–42. [Google Scholar] [PubMed]
- Manicklal, S.; Emery, V.C.; Lazzarotto, T.; Boppana, S.B.; Gupta, R.K. The “silent” global burden of congenital cytomegalovirus. Clin. Microbiol. Rev. 2013, 26, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Fowler, K.B.; Dahle, A.J.; Boppana, S.B.; Pass, R.F. Newborn hearing screening: Will children with hearing loss caused by congenital cytomegalovirus infection be missed? J. Pediatr. 1999, 135, 60–64. [Google Scholar] [CrossRef]
- Schleiss, M.R. Cytomegalovirus in the neonate: Immune correlates of infection and protection. Clin. Dev. Immunol. 2013, 2013, 501801. [Google Scholar] [CrossRef] [PubMed]
- Stagno, S.; Pass, R.F.; Dworsky, M.E.; Britt, W.J.; Alford, C.A. Congenital and perinatal cytomegalovirus infections: Clinical characteristics and pathogenic factors. Birth Defects Orig. Artic. Ser. 1984, 20, 65–85. [Google Scholar] [PubMed]
- Stagno, S.; Reynolds, D.W.; Huang, E.S.; Thames, S.D.; Smith, R.J.; Alford, C.A. Congenital cytomegalovirus-infection—Occurrence in an immune population. N. Engl. J. Med. 1977, 296, 1254–1258. [Google Scholar] [CrossRef] [PubMed]
- Schopfer, K.; Lauber, E.; Krech, U. Congenital cytomegalovirus-infection in newborn-infants of mothers infected before pregnancy. Arch. Dis. Child 1978, 53, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Diosi, P.; Babusceac, L.; Nevinglovschi, O.; Kun-Stoicu, G. Cytomegalovirus infection associated with pregnancy. Lancet 1967, 2, 1063–1066. [Google Scholar] [CrossRef]
- Dworsky, M.; Yow, M.; Stagno, S.; Pass, R.F.; Alford, C. Cytomegalovirus infection of breast milk and transmission in infancy. Pediatrics 1983, 72, 295–299. [Google Scholar] [PubMed]
- Hamprecht, K.; Maschmann, J.; Vochem, M.; Dietz, K.; Speer, C.P.; Jahn, G. Epidemiology of transmission of cytomegalovirus from mother to preterm infant by breastfeeding. Lancet 2001, 357, 513–518. [Google Scholar] [CrossRef]
- Stagno, S.; Reynolds, D.W.; Pass, R.F.; Alford, C.A. Breast milk and the risk of cytomegalovirus infection. N. Engl. J. Med. 1980, 302, 1073–1076. [Google Scholar] [CrossRef] [PubMed]
- Adler, S.P. Cytomegalovirus and child day care: Risk factors for maternal infection. Pediatr. Infect. Dis. J. 1991, 10, 590–594. [Google Scholar] [CrossRef] [PubMed]
- Handsfield, H.H.; Chandler, S.H.; Caine, V.A.; Meyers, J.D.; Corey, L.; Medeiros, E.; McDougall, J.K. Cytomegalovirus infection in sex partners: Evidence for sexual transmission. J. Infect. Dis. 1985, 151, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Meyers, J.D. Cytomegalovirus infection following marrow transplantation: Risk, treatment, and prevention. Birth Defects Orig. Artic. Ser. 1984, 20, 101–117. [Google Scholar] [PubMed]
- Vandenberg, A.P.; Vanson, W.J.; Jiwa, N.M.; Vanderbij, W.; Schirm, J.; Vandergiessen, M.; The, T.H. Recent advances in the diagnosis of active cytomegalovirus-infection after organ-transplantation. Transplant. Proc. 1990, 22, 226–228. [Google Scholar]
- Sinclair, J.; Poole, E. Human cytomegalovirus latency and reactivation in and beyond the myeloid lineage. Future Virol. 2014, 9, 557–563. [Google Scholar] [CrossRef]
- Fenner, F. The clinical features and pathogenesis of mouse-pox (infectious ectromelia of mice). J. Pathol. Bacteriol. 1948, 60, 529–552. [Google Scholar] [CrossRef]
- Fenner, F. The pathogenesis of the acute exanthems—An interpretation based on experimental investigations with mousepox (infectious ectromelia of mice). Lancet 1948, 252, 915–920. [Google Scholar] [CrossRef]
- Collins, T.M.; Quirk, M.R.; Jordan, M.C. Biphasic viremia and viral gene-expression in leukocytes during acute cytomegalovirus-infection of mice. J. Virol. 1994, 68, 6305–6311. [Google Scholar] [PubMed]
- Sacher, T.; Andrassy, J.; Kalnins, A.; Dolken, L.; Jordan, S.; Podlech, J.; Ruzsics, Z.; Jauch, K.W.; Reddehase, M.J.; Koszinowski, U.H. Shedding light on the elusive role of endothelial cells in cytomegalovirus dissemination. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Sacher, T.; Podlech, J.; Mohr, C.A.; Jordan, S.; Ruzsics, Z.; Reddehase, M.J.; Koszinowski, U.H. The major virus-producing cell type during murine cytomegalovirus infection, the hepatocyte, is not the source of virus dissemination in the host. Cell Host Microbe 2008, 3, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.E.; Bruce, K.; Lawler, C.; Oliveira, M.; Cardin, R.; Davis-Poynter, N.; Stevenson, P.G. Murine cytomegalovirus spreads by dendritic cell recirculation. mBio 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Daley-Bauer, L.P.; Roback, L.J.; Wynn, G.M.; Mocarski, E.S. Cytomegalovirus hijacks cx3cr1hi patrolling monocytes as immune-privileged vehicles for dissemination in mice. Cell Host Microbe 2014, 15, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Mayer, B.T.; Krantz, E.M.; Swan, D.; Ferrenberg, J.; Simmons, K.; Selke, S.; Huang, M.L.; Casper, C.; Corey, L.; Wald, A.; et al. Transient oral human cytomegalovirus infections indicate inefficient viral spread from very few initially infected cells. J. Virol. 2017, 92. [Google Scholar] [CrossRef] [PubMed]
- Cannon, M.J.; Hyde, T.B.; Schmid, D.S. Review of cytomegalovirus shedding in bodily fluids and relevance to congenital cytomegalovirus infection. Rev. Med. Virol. 2011, 21, 240–255. [Google Scholar] [CrossRef] [PubMed]
- Dela Pena, M.G.; Strelow, L.; Barry, P.A.; Abel, K. Use of specific-pathogen-free (spf) rhesus macaques to better model oral pediatric cytomegalovirus infection. J. Med. Primatol. 2012, 41, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.E.; Lawler, C.; Tan, C.S.E.; MacDonald, K.; Bruce, K.; Mach, M.; Davis-Poynter, N.; Stevenson, P.G. Murine cytomegalovirus exploits olfaction to enter new hosts. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.E.; Lawler, C.; Oliveira, M.T.; Davis-Poynter, N.; Stevenson, P.G. Alveolar macrophages are a prominent but nonessential target for murine cytomegalovirus infecting the lungs. J. Virol. 2016, 90, 2756–2766. [Google Scholar] [CrossRef] [PubMed]
- Heldwein, E.E. Gh/gl supercomplexes at early stages of herpesvirus entry. Curr. Opin. Virol. 2016, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ryckman, B.J.; Jarvis, M.A.; Drummond, D.D.; Nelson, J.A.; Johnson, D.C. Human cytomegalovirus entry into epithelial and endothelial cells depends on genes ul128 to ul150 and occurs by endocytosis and low-ph fusion. J. Virol. 2006, 80, 710–722. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Shenk, T. Human cytomegalovirus virion protein complex required for epithelial and endothelial cell tropism. Proc. Natl. Acad. Sci. USA 2005, 102, 18153–18158. [Google Scholar] [CrossRef] [PubMed]
- Wille, P.T.; Knoche, A.J.; Nelson, J.A.; Jarvis, M.A.; Johnson, D.C. A human cytomegalovirus go-null mutant fails to incorporate gh/gl into the virion envelope and is unable to enter fibroblasts and epithelial and endothelial cells. J. Virol. 2010, 84, 2585–2596. [Google Scholar] [CrossRef] [PubMed]
- Wille, P.T.; Wisner, T.W.; Ryckman, B.; Johnson, D.C. Human cytomegalovirus (hcmv) glycoprotein gb promotes virus entry in trans acting as the viral fusion protein rather than as a receptor-binding protein. mBio 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Grefte, A.; Plachter, B.; Gouw, A.S.; The, T.H.; Jahn, G. Fibroblasts, epithelial cells, endothelial cells and smooth muscle cells are major targets of human cytomegalovirus infection in lung and gastrointestinal tissues. J. Gen. Virol. 1995, 76 Pt 4, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Poole, E.; Juss, J.K.; Krishna, B.; Herre, J.; Chilvers, E.R.; Sinclair, J. Alveolar macrophages isolated directly from human cytomegalovirus (hcmv)-seropositive individuals are sites of hcmv reactivation in vivo. J. Infect. Dis. 2015, 211, 1936–1942. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.C.; Schroer, J.; Shenk, T. Human cytomegalovirus cell-to-cell spread in the absence of an essential assembly protein. Proc. Natl. Acad. Sci. USA 2005, 102, 2081–2086. [Google Scholar] [CrossRef] [PubMed]
- Noriega, V.M.; Gardner, T.J.; Redmann, V.; Bongers, G.; Lira, S.A.; Tortorella, D. Human cytomegalovirus us28 facilitates cell-to-cell viral dissemination. Viruses 2014, 6, 1202–1218. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Schmidt, K.; Knapp, J.; Kahl, M.; Beck, R.; Waldman, J.; Hebart, H.; Einsele, H.; Jahn, G. Modification of human cytomegalovirus tropism through propagation in vitro is associated with changes in the viral genome. J. Gen. Virol. 1999, 80, 2867–2877. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Knapp, J.; Plachter, B.; Schmidt, K.; Jahn, G. Quantification of replication of clinical cytomegalovirus isolates in cultured endothelial cells and fibroblasts by a focus expansion assay. J. Virol. Methods 1997, 63, 103–112. [Google Scholar] [CrossRef]
- Grundy, J.E.; Lawson, K.M.; MacCormac, L.P.; Fletcher, J.M.; Yong, K.L. Cytomegalovirus-infected endothelial cells recruit neutrophils by the secretion of c-x-c chemokines and transmit virus by direct neutrophil-endothelial cell contact and during neutrophil transendothelial migration. J. Infect. Dis. 1998, 177, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Waldman, W.J.; Knight, D.A.; Huang, E.H.; Sedmak, D.D. Bidirectional transmission of infectious cytomegalovirus between monocytes and vascular endothelial-cells—An in-vitro model. J. Infect. Dis. 1995, 171, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Hahn, G.; Revello, M.G.; Patrone, M.; Percivalle, E.; Campanini, G.; Sarasini, A.; Wagner, M.; Gallina, A.; Milanesi, G.; Koszinowski, U.; et al. Human cytomegalovirus ul131-128 genes are indispensable for virus growth in endothelial cells and virus transfer to leukocytes. J. Virol. 2004, 78, 10023–10033. [Google Scholar] [CrossRef] [PubMed]
- Bentz, G.L.; Jarquin-Pardo, M.; Chan, G.; Smith, M.S.; Sinzger, C.; Yurochko, A.D. Human cytomegalovirus (hcmv) infection of endothelial cells promotes naive monocyte extravasation and transfer of productive virus to enhance hematogenous dissemination of hcmv. J. Virol. 2006, 80, 11539–11555. [Google Scholar] [CrossRef] [PubMed]
- Gerna, G.; Baldanti, F.; Revello, M.G. Pathogenesis of human cytomegalovirus infection and cellular targets. Hum. Immunol. 2004, 65, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Gerna, G.; Percivalle, E.; Baldanti, F.; Sozzani, S.; Lanzarini, P.; Genini, E.; Lilleri, D.; Revello, M.G. Human cytomegalovirus replicates abortively in polymorphonuclear leukocytes after transfer from infected endothelial cells via transient microfusion events. J. Virol. 2000, 74, 5629–5638. [Google Scholar] [CrossRef] [PubMed]
- Mercer, F.; Ng, S.H.; Brown, T.M.; Boatman, G.; Johnson, P.J. Neutrophils kill the parasite trichomonas vaginalis using trogocytosis. PLoS Biol. 2018, 16, e2003885. [Google Scholar] [CrossRef] [PubMed]
- Adler, B.; Sinzger, C. Endothelial cells in human cytomegalovirus infection: One host cell out of many or a crucial target for virus spread? Thromb. Haemost. 2009, 102, 1057–1063. [Google Scholar] [PubMed]
- Grefte, A.; Vandergiessen, M.; Vanson, W.; The, T.H. Circulating cytomegalovirus (cmv)-infected endothelial-cells in patients with an active cmv infection. J. Infect. Dis. 1993, 167, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Percivalle, E.; Revello, M.G.; Vago, L.; Morini, F.; Gerna, G. Circulating endothelial giant-cells permissive for human cytomegalovirus (hcmv) are detected in disseminated hcmv infections with organ involvement. J. Clin. Investig. 1993, 92, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Boom, R.; Sol, C.J.A.; Schuurman, T.; van Breda, A.; Weel, J.F.L.; Beld, M.; ten Berge, I.J.M.; Wertheim-van Dillen, P.M.E.; de Jong, M.D. Human cytomegalovirus DNA in plasma and serum specimens of renal transplant recipients is highly fragmented. J. Clin. Microbiol. 2002, 40, 4105–4113. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y.; Hou, P.F.; Bi, J.; Ying, C.M. Detection of human cytomegalovirus DNA in various blood components after liver transplantation. Braz. J. Med. Biol. Res. 2014, 47, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Eisenfeld, L.; Silver, H.; Mclaughlin, J.; Klevjeranderson, P.; Mayo, D.; Anderson, J.; Herson, V.; Krause, P.; Savidakis, J.; Lazar, A.; et al. Prevention of transfusion-associated cytomegalovirus-infection in neonatal patients by the removal of white cells from blood. Transfusion 1992, 32, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, G.L.; Hayes, K.; Hudson, I.L.; James, J. Prevention of transfusion-acquired cytomegalovirus infection in infants by blood filtration to remove leucocytes. Neonatal cytomegalovirus infection study group. Lancet 1989, 1, 1228–1231. [Google Scholar] [CrossRef]
- Hassan-Walker, A.F.; Mattes, F.M.; Griffiths, P.D.; Emery, V.C. Quantity of cytomegalovirus DNA in different leukocyte populations during active infection in vivo and the presence of gb and ul18 transcripts. J. Med. Virol. 2001, 64, 283–289. [Google Scholar] [CrossRef] [PubMed]
- Schafer, P.; Tenschert, W.; Cremaschi, L.; Schroter, M.; Gutensohn, K.; Laufs, R. Cytomegalovirus cultured from different major leukocyte subpopulations: Association with clinical features in cmv immunoglobulin g-positive renal allograft recipients. J. Med. Virol. 2000, 61, 488–496. [Google Scholar] [CrossRef]
- Grefte, A.; Harmsen, M.C.; van der Giessen, M.; Knollema, S.; van Son, W.J.; The, T.H. Presence of human cytomegalovirus (hcmv) immediate early mrna but not ppul83 (lower matrix protein pp65) mrna in polymorphonuclear and mononuclear leukocytes during active hcmv infection. J. Gen. Virol. 1994, 75 Pt 8, 1989–1998. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, C.R., Jr.; Black, P.H.; Hirsch, M.S. Interaction of cytomegalovirus with leukocytes from patients with mononucleosis due to cytomegalovirus. J. Infect. Dis. 1977, 136, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Fiala, M.; Payne, J.E.; Berne, T.V.; Moore, T.C.; Henle, W.; Montgomerie, J.Z.; Chatterjee, S.N.; Guze, L.B. Epidemiology of cytomegalovirus infection after transplantation and immunosuppression. J. Infect. Dis. 1975, 132, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Miller-Kittrell, M.; Sparer, T.E. Feeling manipulated: Cytomegalovirus immune manipulation. Virol. J. 2009, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Mocarski, E.S., Jr. Immunomodulation by cytomegaloviruses: Manipulative strategies beyond evasion. Trends Microbiol. 2002, 10, 332–339. [Google Scholar] [CrossRef]
- Comerford, I.; McColl, S.R. Mini-review series: Focus on chemokines. Immunol. Cell Biol. 2011, 89, 183–184. [Google Scholar] [CrossRef] [PubMed]
- Rot, A.; von Andrian, U.H. Chemokines in innate and adaptive host defense: Basic chemokinese grammar for immune cells. Annu. Rev. Immunol. 2004, 22, 891–928. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.; Dogra, P.; Masi, T.J.; Pitt, E.A.; de Kruijf, P.; Smit, M.J.; Sparer, T.E. Novel human cytomegalovirus viral chemokines, vcxcl-1s, display functional selectivity for neutrophil signaling and function. J. Immunol. 2015, 195, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Penfold, M.E.; Dairaghi, D.J.; Duke, G.M.; Saederup, N.; Mocarski, E.S.; Kemble, G.W.; Schall, T.J. Cytomegalovirus encodes a potent alpha chemokine. Proc. Natl. Acad. Sci. USA 1999, 96, 9839–9844. [Google Scholar] [CrossRef] [PubMed]
- Yamin, R.; Lecker, L.S.M.; Weisblum, Y.; Vitenshtein, A.; Le-Trilling, V.T.K.; Wolf, D.G.; Mandelboim, O. Hcmv vcxcl1 binds several chemokine receptors and preferentially attracts neutrophils over nk cells by interacting with cxcr2. Cell Rep. 2016, 15, 1542–1553. [Google Scholar] [CrossRef] [PubMed]
- Luttichau, H.R. The cytomegalovirus ul146 gene product vcxcl1 targets both cxcr1 and cxcr2 as an agonist. J. Biol. Chem. 2010, 285, 9137–9146. [Google Scholar] [CrossRef] [PubMed]
- Lurain, N.S.; Fox, A.M.; Lichy, H.M.; Bhorade, S.M.; Ware, C.F.; Huang, D.D.; Kwan, S.P.; Garrity, E.R.; Chou, S.W. Analysis of the human cytomegalovirus genomic region from ul146 through ul147a reveals sequence hypervariability, genotypic stability, and overlapping transcripts. Virol. J. 2006, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Hassan-Walker, A.F.; Okwuadi, S.; Lee, L.; Griffiths, P.D.; Emery, V.C. Sequence variability of the alpha-chemokine ul146 from clinical strains of human cytomegalovirus. J. Med. Virol. 2004, 74, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Schraufstatter, I.U.; Chung, J.; Burger, M. Il-8 activates endothelial cell cxcr1 and cxcr2 through rho and rac signaling pathways. Am. J. Physiol.-Lung C 2001, 280, L1094–L1103. [Google Scholar] [CrossRef] [PubMed]
- Bonecchi, R.; Facchetti, F.; Dusi, S.; Luini, W.; Lissandrini, D.; Simmelink, M.; Locati, M.; Bernasconi, S.; Allavena, P.; Brandt, E.; et al. Induction of functional il-8 receptors by il-4 and il-13 in human monocytes. J. Immunol. 2000, 164, 3862–3869. [Google Scholar] [CrossRef] [PubMed]
- Polec, A.; Raki, M.; Abyholm, T.; Tanbo, T.G.; Fedorcsak, P. Interaction between granulosa-lutein cells and monocytes regulates secretion of angiogenic factors in vitro. Hum. Reprod. 2011, 26, 2819–2829. [Google Scholar] [CrossRef] [PubMed]
- Gerszten, R.E.; Garcia-Zepeda, E.A.; Lim, Y.C.; Yoshida, M.; Ding, H.A.; Gimbrone, M.A., Jr.; Luster, A.D.; Luscinskas, F.W.; Rosenzweig, A. Mcp-1 and il-8 trigger firm adhesion of monocytes to vascular endothelium under flow conditions. Nature 1999, 398, 718–723. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, C.R., Jr.; Stossel, T.P.; Black, P.H.; Hirsch, M.S. Polymorphonuclear leukocyte function during cytomegalovirus mononucleosis. Clin. Immunol. Immunopathol. 1979, 12, 331–334. [Google Scholar] [CrossRef]
- Gerna, G.; Zipeto, D.; Percivalle, E.; Parea, M.; Revello, M.G.; Maccario, R.; Peri, G.; Milanesi, G. Human cytomegalovirus-infection of the major leukocyte subpopulations and evidence for initial viral replication in polymorphonuclear leukocytes from viremic patients. J. Infect. Dis. 1992, 166, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Plachter, B.; Grefte, A.; The, T.H.; Jahn, G. Tissue macrophages are infected by human cytomegalovirus in vivo. J. Infect. Dis. 1996, 173, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Van der Strate, B.W.; Hillebrands, J.L.; Lycklama a Nijeholt, S.S.; Beljaars, L.; Bruggeman, C.A.; Van Luyn, M.J.; Rozing, J.; The, T.H.; Meijer, D.K.; Molema, G.; et al. Dissemination of rat cytomegalovirus through infected granulocytes and monocytes in vitro and in vivo. J. Virol. 2003, 77, 11274–11278. [Google Scholar] [CrossRef] [PubMed]
- Bale, J.F.; Kern, E.R.; Overall, J.C.; Baringer, J.R. Impaired migratory and chemotactic activity of neutrophils during murine cytomegalovirus-infection. J. Infect. Dis. 1983, 148, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Pocock, J.M.; Storisteanu, D.M.L.; Reeves, M.B.; Juss, J.K.; Wills, M.R.; Cowburn, A.S.; Chilvers, E.R. Human cytomegalovirus delays neutrophil apoptosis and stimulates the release of a prosurvival secretome. Front. Immunol. 2017, 8, 1185. [Google Scholar] [CrossRef] [PubMed]
- Slobedman, B.; Mocarski, E.S. Quantitative analysis of latent human cytomegalovirus. J. Virol. 1999, 73, 4806–4812. [Google Scholar] [PubMed]
- Eash, K.J.; Greenbaum, A.M.; Gopalan, P.K.; Link, D.C. Cxcr2 and cxcr4 antagonistically regulate neutrophil trafficking from murine bone marrow. J. Clin. Investig. 2010, 120, 2423–2431. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.S.; Bentz, G.L.; Alexander, J.S.; Yurochko, A.D. Human cytomegalovirus induces monocyte differentiation and migration as a strategy for dissemination and persistence. J. Virol. 2004, 78, 4444–4453. [Google Scholar] [CrossRef] [PubMed]
- Jakubzick, C.V.; Randolph, G.J.; Henson, P.M. Monocyte differentiation and antigen-presenting functions. Nat. Rev. Immunol. 2017, 17, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Sinzger, C.; Jahn, G. Human cytomegalovirus cell tropism and pathogenesis. Intervirology 1996, 39, 302–319. [Google Scholar] [CrossRef] [PubMed]
- Taylorwiedeman, J.; Sissons, P.; Sinclair, J. Induction of endogenous human cytomegalovirus gene-expression after differentiation of monocytes from healthy carriers. J. Virol. 1994, 68, 1597–1604. [Google Scholar]
- Soderberg, C.; Larsson, S.; Bergstedtlindqvist, S.; Moller, E. Identification of blood mononuclear-cells permissive of cytomegalovirus-infection invitro. Transplant. Proc. 1993, 25, 1416–1418. [Google Scholar] [PubMed]
- Maciejewski, J.P.; Bruening, E.E.; Donahue, R.E.; Sellers, S.E.; Carter, C.; Young, N.S.; Stjeor, S. Infection of mononucleated phagocytes with human cytomegalovirus. Virology 1993, 195, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.; Nogalski, M.T.; Yurochko, A.D. Human cytomegalovirus stimulates monocyte-to-macrophage differentiation via the temporal regulation of caspase 3. J. Virol. 2012, 86, 10714–10723. [Google Scholar] [CrossRef] [PubMed]
- Ibanez, C.E.; Schrier, R.; Ghazal, P.; Wiley, C.; Nelson, J.A. Human cytomegalovirus productively infects primary differentiated macrophages. J. Virol. 1991, 65, 6581–6588. [Google Scholar] [PubMed]
- Sinzger, C.; Eberhardt, K.; Cavignac, Y.; Weinstock, C.; Kessler, T.; Jahn, G.; Davignon, J.L. Macrophage cultures are susceptible to lytic productive infection by endothelial-cell-propagated human cytomegalovirus strains and present viral ie1 protein to cd4(+) t cells despite late downregulation of mhc class ii molecules. J. Gen. Virol. 2006, 87, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Frascaroli, G.; Varani, S.; MoeppS, B.; Sinzger, C.; Landini, M.P.; Mertens, T. Human cytomegalovirus subverts the functions of monocytes, impairing chemokine-mediated migration and leukocyte recruitment. J. Virol. 2006, 80, 7578–7589. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Tao, R.; Gao, H.H.; Xu, J.; Shang, S.Q.; Zhao, N. Hcmv-encoded ul128 enhances tnf-alpha and il-6 expression and promotes pbmc proliferation through the mapk/erk pathway in vitro. Viral Immunol. 2012, 25, 98–105. [Google Scholar] [CrossRef] [PubMed]
- McSharry, B.P.; Avdic, S.; Slobedman, B. Human cytomegalovirus encoded homologs of cytokines, chemokines and their receptors: Roles in immunomodulation. Viruses 2012, 4, 2448–2470. [Google Scholar] [CrossRef] [PubMed]
- Bernhagen, J.; Krohn, R.; Lue, H.; Gregory, J.L.; Zernecke, A.; Koenen, R.R.; Dewor, M.; Georgiev, I.; Schober, A.; Leng, L.; et al. Mif is a noncognate ligand of cxc chemokine receptors in inflammatory and atherogenic cell recruitment. Nat. Med. 2007, 13, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Lafemina, R.L.; Hayward, G.S. Differences in cell type-specific blocks to immediate early gene-expression and DNA-replication of human, simian and murine cytomegalo-virus. J. Gen. Virol. 1988, 69, 355–374. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Maul, G.G. Mouse cytomegalovirus crosses the species barrier with help from a few human cytomegalovirus proteins. J. Virol. 2006, 80, 7510–7521. [Google Scholar] [CrossRef] [PubMed]
- Lilja, A.E.; Shenk, T. Efficient replication of rhesus cytomegalovirus variants in multiple rhesus and human cell types. Proc. Natl. Acad. Sci. USA 2008, 105, 19950–19955. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.B. The murine cytomegalovirus as a model for the study of viral pathogenesis and persistent infections. Arch. Virol. 1979, 62, 1–29. [Google Scholar] [CrossRef] [PubMed]
- Dogra, P.; Sparer, T.E. What we have learned from animal models of hcmv. Methods Mol. Biol. 2014, 1119, 267–288. [Google Scholar] [PubMed]
- Wagner, F.M.; Brizic, I.; Prager, A.; Trsan, T.; Arapovic, M.; Lemmermann, N.A.W.; Podlech, J.; Reddehase, M.J.; Lemnitzer, F.; Bosse, J.B.; et al. The viral chemokine mck-2 of murine cytomegalovirus promotes infection as part of a gh/gl/mck-2 complex. PLoS Pathog. 2013, 9, e1003493. [Google Scholar] [CrossRef] [PubMed]
- Saederup, N.; Lin, Y.C.; Dairaghi, D.J.; Schall, T.J.; Mocarski, E.S. Cytomegalovirus-encoded beta chemokine promotes monocyte-associated viremia in the host. Proc. Natl. Acad. Sci. USA 1999, 96, 10881–10886. [Google Scholar] [CrossRef] [PubMed]
- Saederup, N.; Aguirre, S.A.; Sparer, T.E.; Bouley, D.M.; Mocarski, E.S. Murine cytomegalovirus cc chemokine homolog mck-2 (m131-129) is a determinant of dissemination that increases inflammation at initial sites of infection. J. Virol. 2001, 75, 9966–9976. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Aliberti, J.; Graemmel, P.; Sunshine, M.J.; Kreutzberg, G.W.; Sher, A.; Littman, D.R. Analysis of fractalkine receptor cx(3)cr1 function by targeted deletion and green fluorescent protein reporter gene insertion. Mol. Cell. Biol. 2000, 20, 4106–4114. [Google Scholar] [CrossRef] [PubMed]
- Auffray, C.; Sieweke, M.H.; Geissmann, F. Blood monocytes: Development, heterogeneity, and relationship with dendritic cells. Annu. Rev. Immunol. 2009, 27, 669–692. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, J.; Reeves, M. The intimate relationship between human cytomegalovirus and the dendritic cell lineage. Front. Microbiol. 2014, 5, 389. [Google Scholar] [CrossRef] [PubMed]
- Bittencourt, F.M.; Wu, S.E.; Bridges, J.P.; Miller, W.E. The m33 g protein-coupled receptor encoded by murine cytomegalovirus is dispensable for hematogenous dissemination but is required for growth within the salivary gland. J. Virol. 2014, 88, 11811–11824. [Google Scholar] [CrossRef] [PubMed]
- Dogra, P.; Miller-Kittrell, M.; Pitt, E.; Jackson, J.W.; Masi, T.; Copeland, C.; Wu, S.; Miller, W.; Sparer, T. A little cooperation helps murine cytomegalovirus (mcmv) go a long way: Mcmv co-infection rescues a chemokine salivary gland defect. J. Gen. Virol. 2016, 97, 2957–2972. [Google Scholar] [CrossRef] [PubMed]
- Selgrade, M.K.; Osborn, J.E. Role of macrophages in resistance to murine cytomegalovirus. Infect. Immun. 1974, 10, 1383–1390. [Google Scholar] [PubMed]
- Farrell, H.E.; Davis-Poynter, N.; Bruce, K.; Lawler, C.; Dolken, L.; Mach, M.; Stevenson, P.G. Lymph node macrophages restrict murine cytomegalovirus dissemination. J. Virol. 2015, 89, 7147–7158. [Google Scholar] [CrossRef] [PubMed]
- Farrell, H.E.; Bruce, K.; Lawler, C.; Cardin, R.D.; Davis-Poynter, N.J.; Stevenson, P.G. Type 1 interferons and nk cells limit murine cytomegalovirus escape from the lymph node subcapsular sinus. PLoS Pathog. 2016, 12. [Google Scholar] [CrossRef] [PubMed]
- Vomaske, J.; Denton, M.; Kreklywich, C.; Andoh, T.; Osborn, J.M.; Chen, D.; Messaoudi, I.; Orloff, S.L.; Streblow, D.N. Cytomegalovirus cc chemokine promotes immune cell migration. J. Virol. 2012, 86, 11833–11844. [Google Scholar] [CrossRef] [PubMed]
- Kaptein, S.J.F.; van Cleef, K.W.R.; Gruijthuijsen, Y.K.; Beuken, E.V.H.; van Buggenhout, L.; Beisser, P.S.; Stassen, F.R.M.; Bruggeman, C.A.; Vink, C. The r131 gene of rat cytomegalovirus encodes a proinflammatory cc chemokine homolog which is essential for the production of infectious virus in the salivary glands. Virus Genes 2004, 29, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Streblow, D.N.; Kreklywich, C.; Yin, Q.; De La Melena, V.T.; Corless, C.L.; Smith, P.A.; Brakebill, C.; Cook, J.W.; Vink, C.; Bruggeman, C.A.; et al. Cytomegalovirus-mediated upregulation of chemokine expression correlates with the acceleration of chronic rejection in rat heart transplants. J. Virol. 2003, 77, 2182–2194. [Google Scholar] [CrossRef] [PubMed]
- Streblow, D.N.; Kreklywich, C.N.; Smith, P.; Soule, J.L.; Meyer, C.; Yin, M.; Beisser, P.; Vink, C.; Nelson, J.A.; Orloff, S.L. Rat cytomegalovirus-accelerated transplant vascular sclerosis is reduced with mutation of the chemokine-receptor r33. Am. J. Transplant. 2005, 5, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Haggerty, S.M.; Schleiss, M.R. A novel cc-chemokine homolog encoded by guinea pig cytomegalovirus. Virus Genes 2002, 25, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Penfold, M.; Miao, Z.; Wang, Y.; Haggerty, S.; Schleiss, M.R. A macrophage inflammatory protein homolog encoded by guinea pig cytomegalovirus signals via cc chemokine receptor 1. Virology 2003, 316, 202–212. [Google Scholar] [CrossRef]
- Schraff, S.A.; Brown, D.K.; Schleiss, M.R.; Meinzen-Derr, J.; Greinwald, J.H.; Choo, D.I. The role of cmv inflammatory genes in hearing loss. Otol. Neurotol. 2007, 28, 964–969. [Google Scholar] [CrossRef] [PubMed]
- Schraff, S.A.; Schleiss, M.R.; Brown, D.K.; Meinzen-Derr, J.; Choi, Y.; Greinwald, J.H.; Choo, D.I. Macrophage inflammatory proteins in cytomegalovirus-related inner ear injury. Otolaryngol. Head Neck 2007, 137, 612–618. [Google Scholar] [CrossRef] [PubMed]
- Leviton, M.P.; Lacayo, J.C.; Choi, K.Y.; Hernandez-Alvarado, N.; Wey, A.; Schleiss, M.R. An attenuated cytomegalovirus vaccine with a deletion of a viral chemokine gene is protective against congenital cmv transmission in a guinea pig model. Clin. Dev. Immunol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Auerbach, M.; Yan, D.; Fouts, A.; Xu, M.; Estevez, A.; Austin, C.D.; Bazan, F.; Feierbach, B. Characterization of the guinea pig cmv gh/gl/gp129/gp131/gp133 complex in infection and spread. Virology 2013, 441, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Schleiss, M.R.; McVoy, M.A. Guinea pig cytomegalovirus (gpcmv): A model for the study of the prevention and treatment of maternal-fetal transmission. Future Virol. 2010, 5, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Cekinovic, D.; Lisnic, V.J.; Jonjic, S. Rodent models of congenital cytomegalovirus infection. Methods Mol. Biol. 2014, 1119, 289–310. [Google Scholar] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson, J.W.; Sparer, T. There Is Always Another Way! Cytomegalovirus’ Multifaceted Dissemination Schemes. Viruses 2018, 10, 383. https://doi.org/10.3390/v10070383
Jackson JW, Sparer T. There Is Always Another Way! Cytomegalovirus’ Multifaceted Dissemination Schemes. Viruses. 2018; 10(7):383. https://doi.org/10.3390/v10070383
Chicago/Turabian StyleJackson, Joseph W., and Tim Sparer. 2018. "There Is Always Another Way! Cytomegalovirus’ Multifaceted Dissemination Schemes" Viruses 10, no. 7: 383. https://doi.org/10.3390/v10070383
APA StyleJackson, J. W., & Sparer, T. (2018). There Is Always Another Way! Cytomegalovirus’ Multifaceted Dissemination Schemes. Viruses, 10(7), 383. https://doi.org/10.3390/v10070383