In Vitro Studies of Lipopolysaccharide-Mediated DNA Release of Podovirus HK620


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Deletion of ompA or ompC Genes from E. coli H TD2158
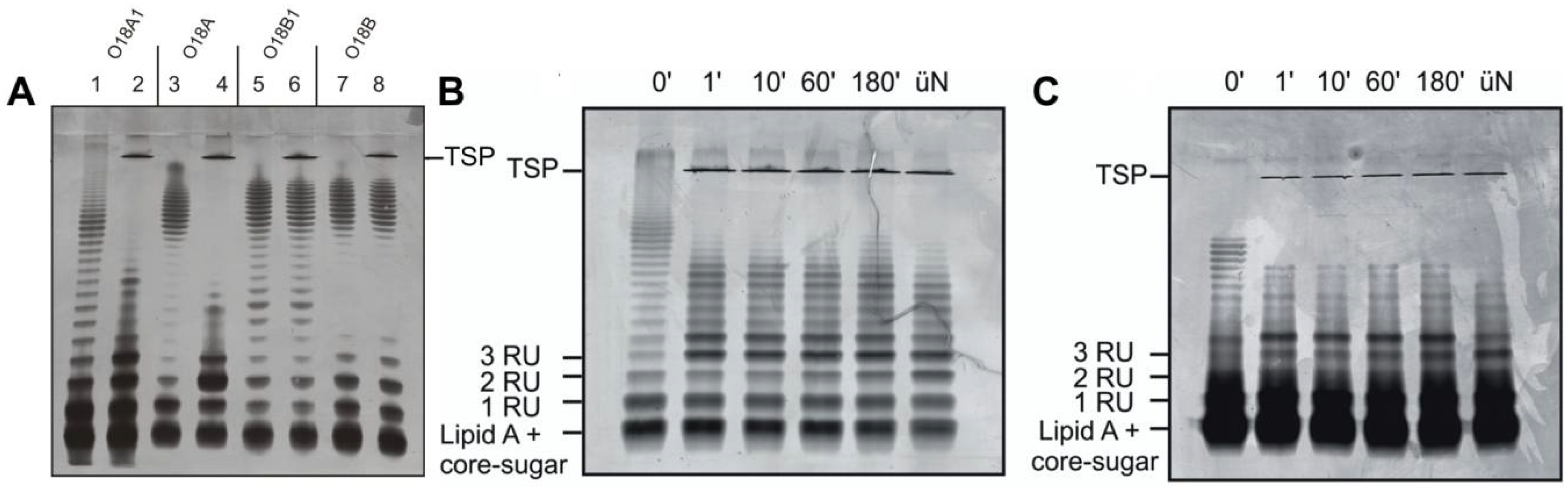
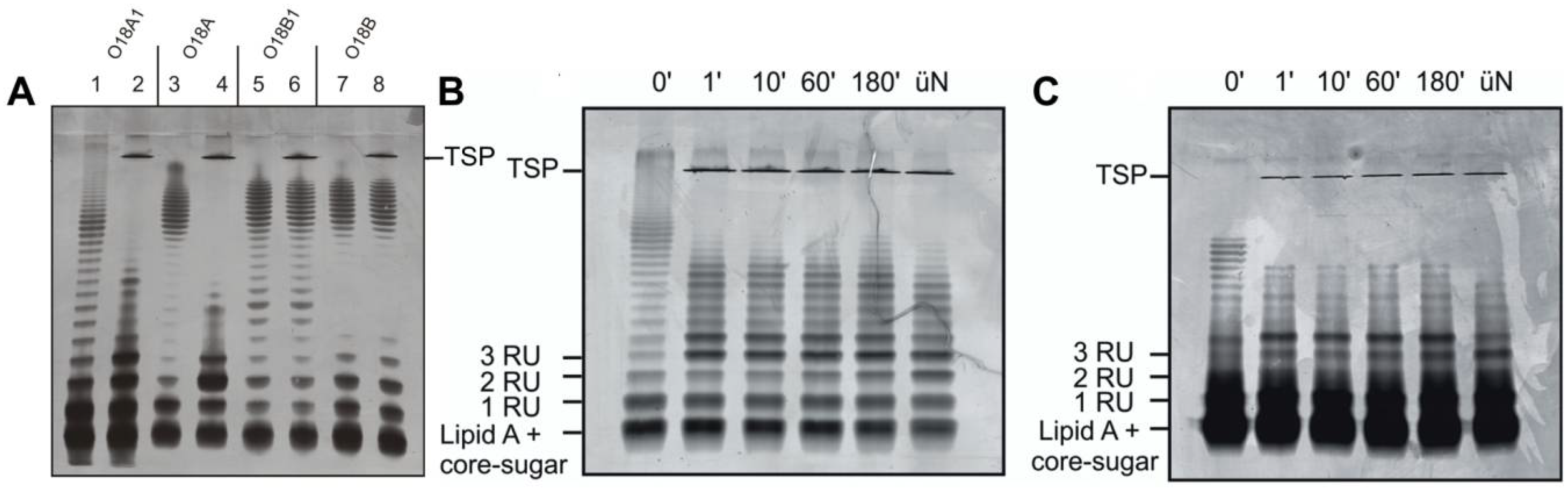
2.3. Preparation of LPS, Lipid A, PS and TSP-Digested LPS
2.4. DNA Ejection and Phage Inhibition Experiments
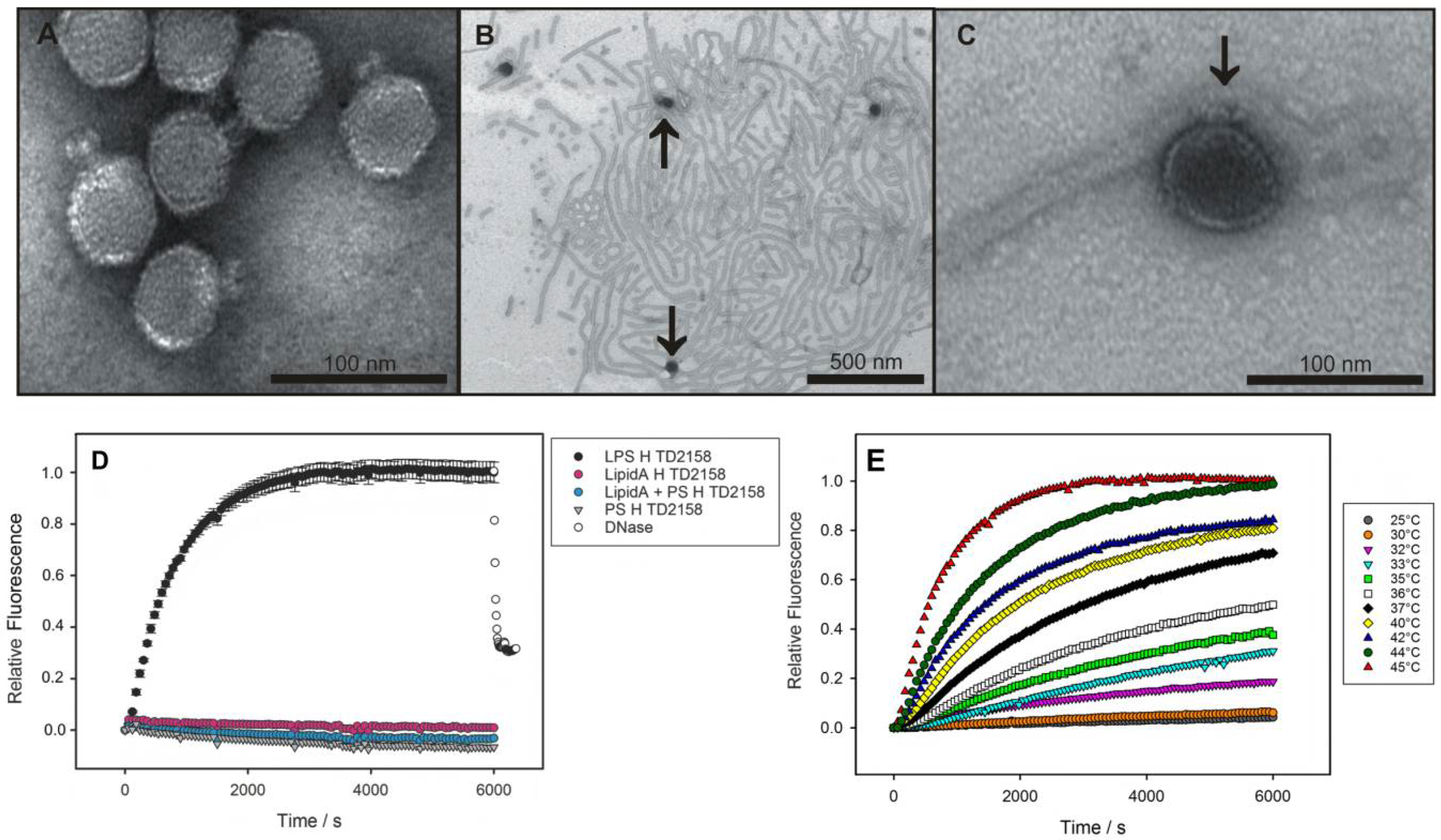
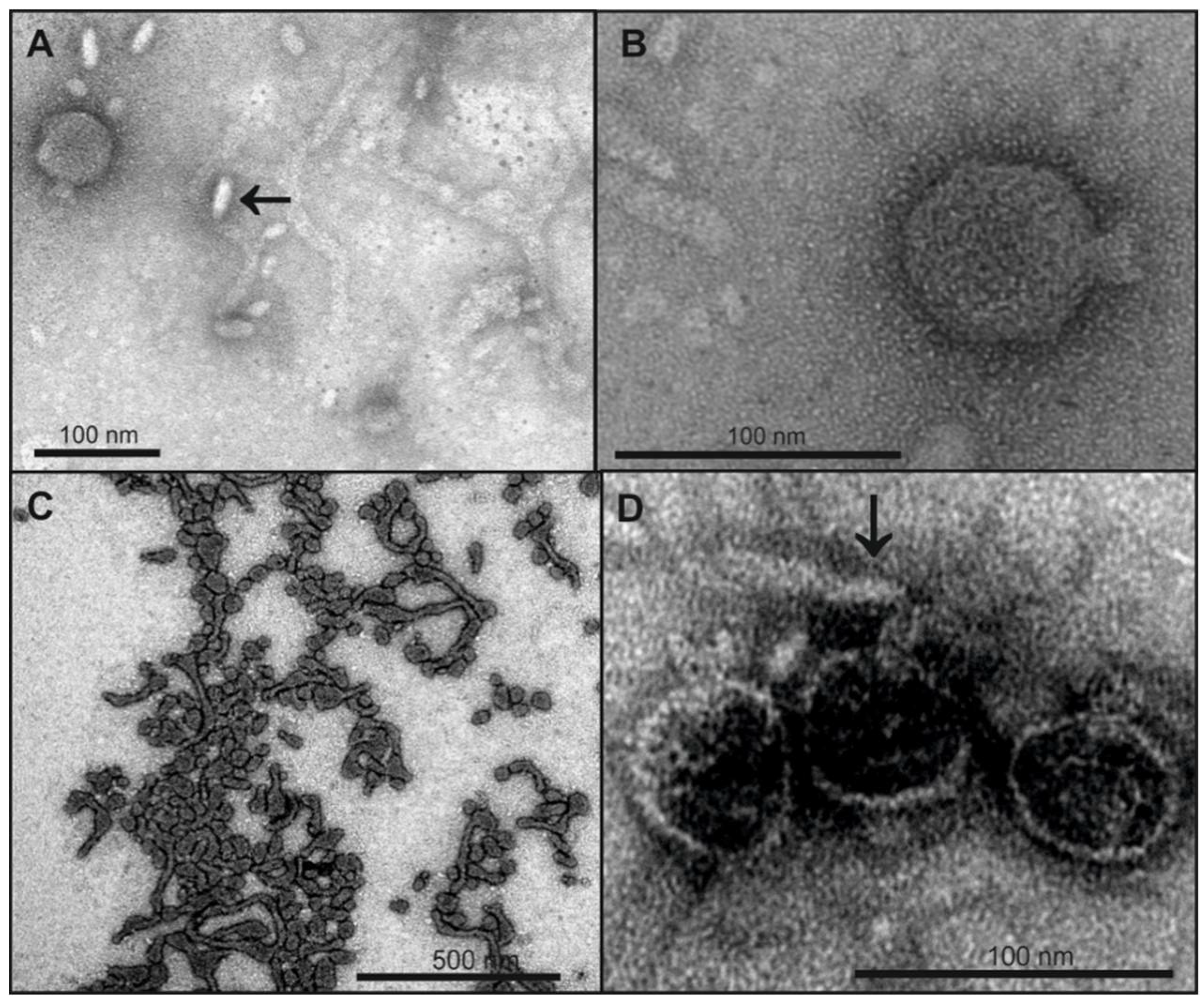
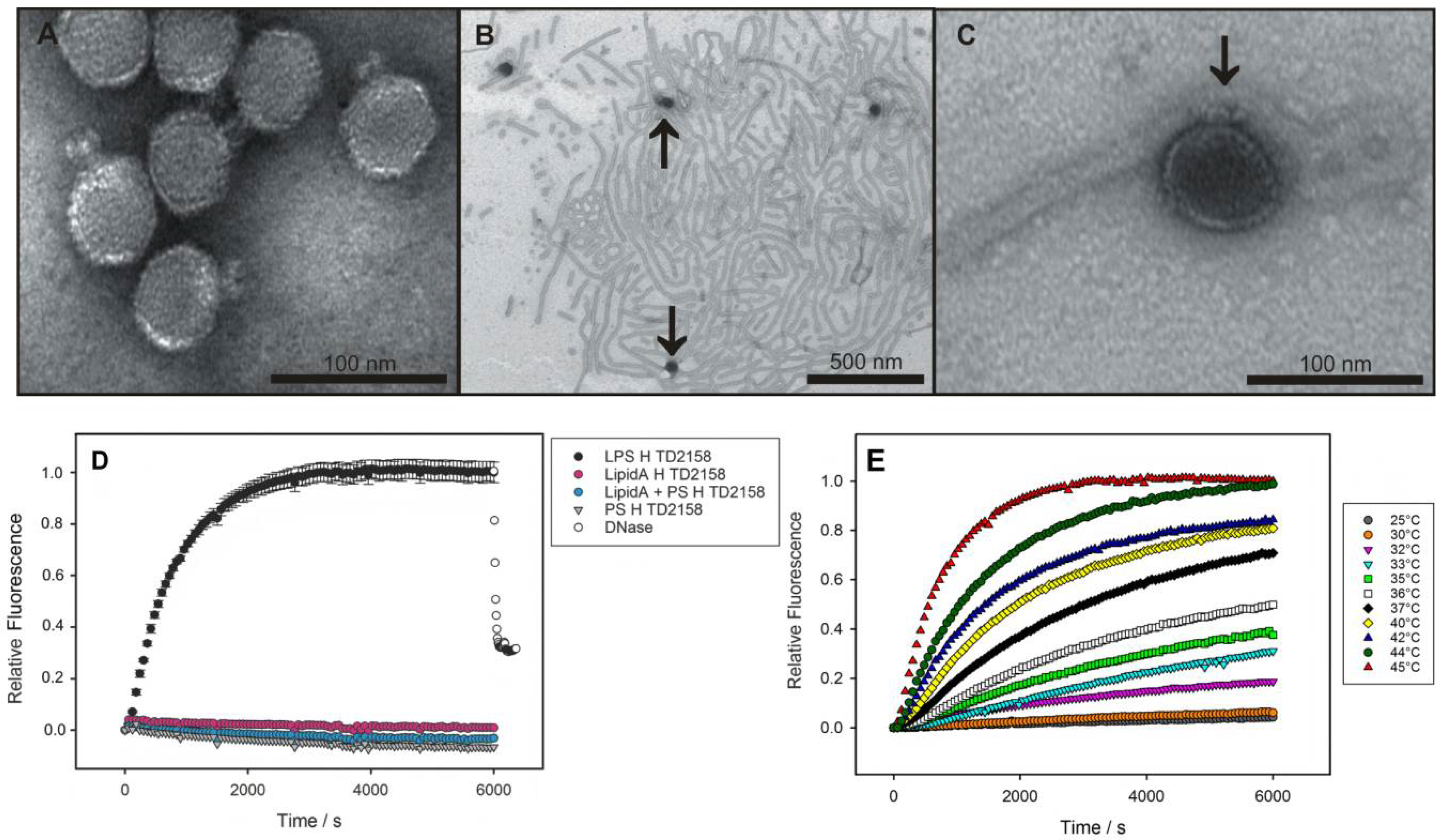
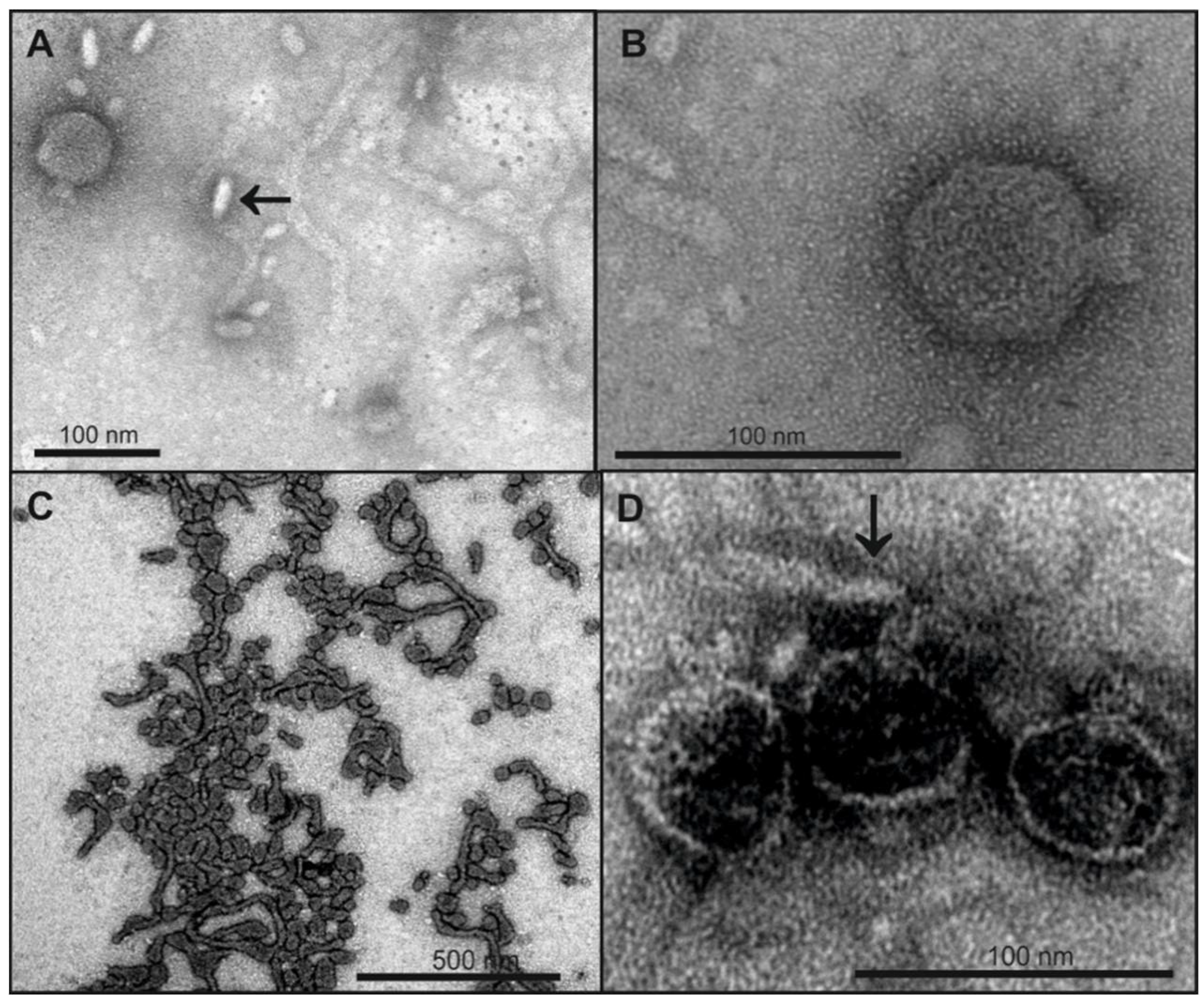
2.5. LPS Morphology Studies
3. Results
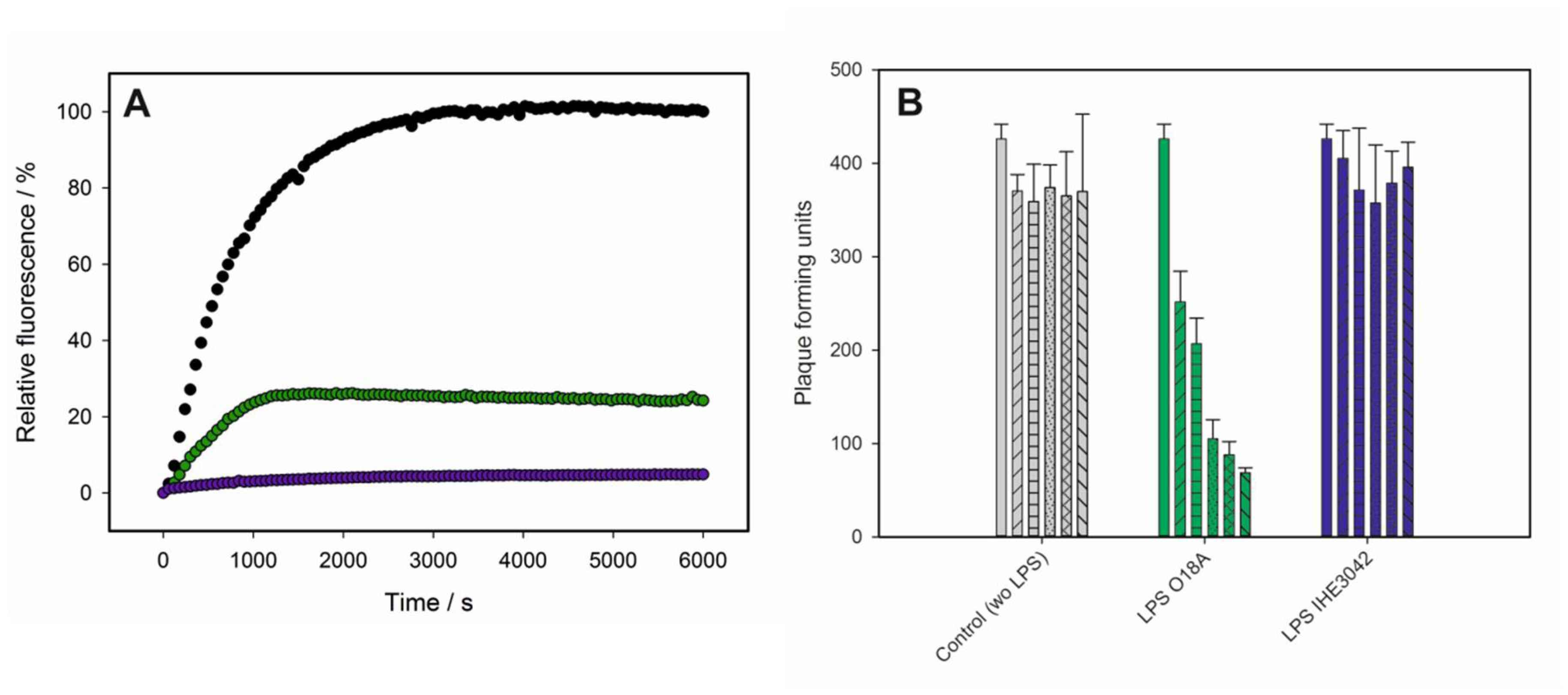
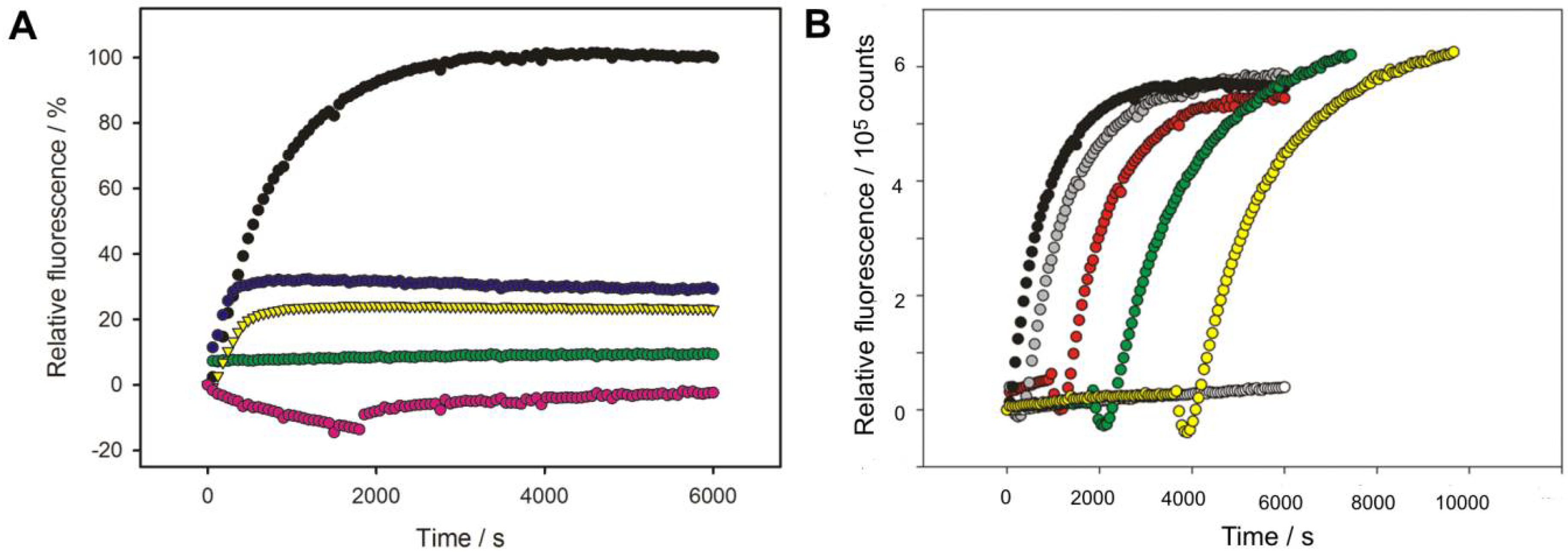
3.1. Bacteriophage HK620 Releases Its DNA Upon LPS Incubation In Vitro
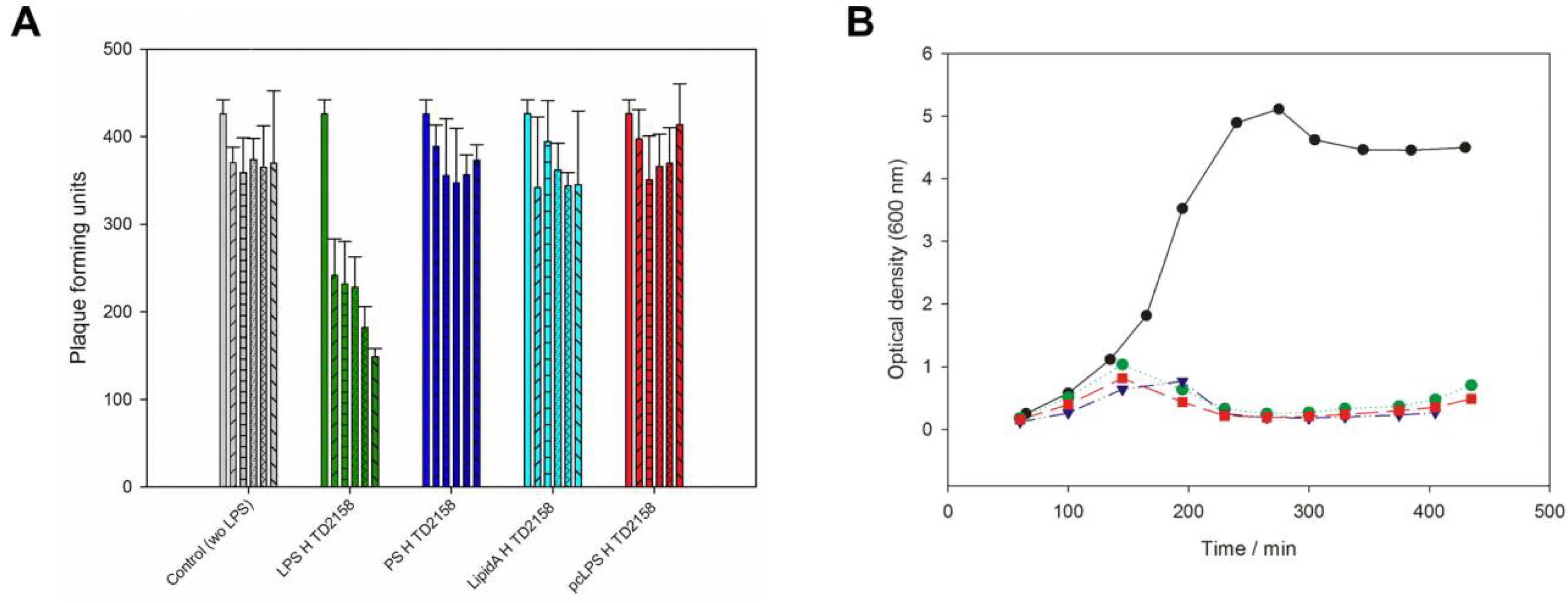
3.2. LPS Is the Receptor Sufficient for HK620 Host Cell Infection
3.3. O-Antigen Specificity of TSP Guides Phage HK620 Host Specificity
3.4. O-Antigen Chain Length Determines the Properties of the LPS Receptor
3.5. Phage Adsorption in Competition with TSP-Mediated O-Antigen Cleavage
4. Discussion
4.1. Role of O-Antigen Receptor Composition
4.2. In Vitro Particle Opening
4.3. Properties of the LPS Receptor Influencing In Vitro Particle Opening
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Citorik, R.J.; Mimee, M.; Lu, T.K. Bacteriophage-based synthetic biology for the study of infectious diseases. Curr. Opin. Microbiol. 2014, 19, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.R.; Molineux, I.J. Short noncontractile tail machines: Adsorption and DNA delivery by podoviruses. In Viral Molecular Machines; Rossmann, M.G., Rao, V.B., Eds.; Springer: Berlin, Germany, 2012; Volume 726, pp. 143–179. [Google Scholar]
- Broeker, N.K.; Barbirz, S. Not a barrier but a key: How bacteriophages exploit host’s O-antigen as an essential receptor to initiate infection. Mol. Microbiol. 2017, 105, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; van Valen, D.; Hu, Q.; Phillips, R. Ion-dependent dynamics of DNA ejections for bacteriophage lambda. Biophys. J. 2010, 99, 1101–1109. [Google Scholar] [CrossRef] [PubMed]
- Chiaruttini, N.; Letellier, L.; Viasnoff, V. A novel method to couple electrophysiological measurements and fluorescence imaging of suspended lipid membranes: The example of T5 bacteriophage DNA ejection. PLoS ONE 2013, 8, e84376. [Google Scholar] [CrossRef] [PubMed]
- Raspaud, E.; Forth, T.; Sao-Jose, C.; Tavares, P.; de Frutos, M. A kinetic analysis of DNA ejection from tailed phages revealing the prerequisite activation energy. Biophys. J. 2007, 93, 3999–4005. [Google Scholar] [CrossRef] [PubMed]
- Andres, D.; Roske, Y.; Doering, C.; Heinemann, U.; Seckler, R.; Barbirz, S. Tail morphology controls DNA release in two salmonella phages with one lipopolysaccharide receptor recognition system. Mol. Microbiol. 2012, 83, 1244–1253. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Garcia, V.A.; Pulido-Cid, M.; Garcia-Doval, C.; Bocanegra, R.; van Raaij, M.J.; Martin-Benito, J.; Cuervo, A.; Carrascosa, J.L. Conformational changes leading to T7 DNA delivery upon interaction with the bacterial receptor. J. Biol. Chem. 2015, 290, 10038–10044. [Google Scholar] [CrossRef] [PubMed]
- Jeembaeva, M.; Jonsson, B.; Castelnovo, M.; Evilevitch, A. DNA heats up: Energetics of genome ejection from phage revealed by isothermal titration calorimetry. J. Mol. Biol. 2010, 395, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Brussow, H. Phage therapy: The Escherichia coli experience. Microbiology 2005, 151, 2133–2140. [Google Scholar] [CrossRef] [PubMed]
- Bolocan, A.S.; Callanan, J.; Forde, A.; Ross, P.; Hill, C. Phage therapy targeting Escherichia coli—A story with no end? FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Mizushima, S. Roles of lipopolysaccharide and outer membrane protein OmpC of Escherichia coli K-12 in the receptor function for bacteriophage T4. J. Bacteriol. 1982, 151, 718–722. [Google Scholar] [PubMed]
- Israel, J.V.; Anderson, T.F.; Levine, M. In vitro morphogenesis of phage P22 from heads and base-plate parts. Proc. Natl. Acad. Sci. USA 1967, 57, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Prokhorov, N.S.; Riccio, C.; Zdorovenko, E.L.; Shneider, M.M.; Browning, C.; Knirel, Y.A.; Leiman, P.G.; Letarov, A.V. Function of bacteriophage G7C esterase tailspike in host cell adsorption. Mol. Microbiol. 2017, 105, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Richter, W.; Vogel, V.; Howe, J.; Steiniger, F.; Brauser, A.; Koch, M.H.; Roessle, M.; Gutsmann, T.; Garidel, P.; Mäntele, W.; et al. Morphology, size distribution, and aggregate structure of lipopolysaccharide and lipid A dispersions from enterobacterial origin. Innate Immun. 2011, 17, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Frirdich, E.; Whitfield, C. Lipopolysaccharide inner core oligosaccharide structure and outer membrane stability in human pathogens belonging to the Enterobacteriaceae. J. Endotoxin Res. 2005, 11, 133–144. [Google Scholar] [PubMed]
- Lindberg, A.A. Bacteriophage receptors. Annu. Rev. Microbiol. 1973, 27, 205–241. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.; Thuman-Commike, P. Evolution of mosaically related tailed bacteriophage genomes seen through the lens of phage P22 virion assembly. Virology 2011, 411, 393–415. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Weigele, P.; King, J.; Chiu, W.; Jiang, W. Cryo-EM asymmetric reconstruction of bacteriophage P22 reveals organization of its DNA packaging and infecting machinery. Structure 2006, 14, 1073–1082. [Google Scholar] [CrossRef] [PubMed]
- Steinbacher, S.; Baxa, U.; Miller, S.; Weintraub, A.; Seckler, R.; Huber, R. Crystal structure of phage P22 tailspike protein complexed with Salmonella sp. O-antigen receptors. Proc. Natl. Acad. Sci. USA 1996, 93, 10584–10588. [Google Scholar] [CrossRef] [PubMed]
- Andres, D.; Hanke, C.; Baxa, U.; Seul, A.T.; Barbirz, S.; Seckler, R. Tailspike interactions with lipopolysaccharide effect DNA ejection from phage P22 particles in vitro. J. Biol. Chem. 2010, 285, 36768–36775. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, T.S.; Poon, A.P.; Chan, D.; Clark, A.J. General transducing phages like salmonella phage P22 isolated using a smooth strain of Escherichia coli as host. FEMS Microbiol. Lett. 1998, 161, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Zaccheus, M.V.; Broeker, N.K.; Lundborg, M.; Uetrecht, C.; Barbirz, S.; Widmalm, G. Structural studies of the o-antigen polysaccharide from Escherichia coli TD2158 having O18 serogroup specificity and aspects of its interaction with the tailspike endoglycosidase of the infecting bacteriophage HK620. Carbohydr. Res. 2012, 357, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Barbirz, S.; Muller, J.J.; Uetrecht, C.; Clark, A.J.; Heinemann, U.; Seckler, R. Crystal structure of Escherichia coli phage HK620 tailspike: Podoviral tailspike endoglycosidase modules are evolutionarily related. Mol. Microbiol. 2008, 69, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Menouni, R.; Champ, S.; Espinosa, L.; Boudvillain, M.; Ansaldi, M. Transcription termination controls prophage maintenance in Escherichia coli genomes. Proc. Natl. Acad. Sci. USA 2013, 110, 14414–14419. [Google Scholar] [CrossRef] [PubMed]
- Clark, A.J.; Inwood, W.; Cloutier, T.; Dhillon, T.S. Nucleotide sequence of coliphage HK620 and the evolution of lambdoid phages. J. Mol. Biol. 2001, 311, 657–679. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.; Walker-Kopp, N.; Casjens, S.R.; Cingolani, G. An evolutionarily conserved family of virion tail needles related to bacteriophage P22 GP26: Correlation between structural stability and length of the alpha-helical trimeric coiled coil. J. Mol. Biol. 2009, 391, 227–245. [Google Scholar] [CrossRef] [PubMed]
- Broeker, N.K.; Gohlke, U.; Müller, J.J.; Uetrecht, C.; Heinemann, U.; Seckler, R.; Barbirz, S. Single amino acid exchange in bacteriophage HK620 tailspike protein results in thousand-fold increase of its oligosaccharide affinity. Glycobiology 2013, 23, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Parent, K.N.; Gilcrease, E.B.; Casjens, S.R.; Baker, T.S. Structural evolution of the P22-like phages: Comparison of Sf6 and P22 procapsid and virion architectures. Virology 2012, 427, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Leavitt, J.C.; Gogokhia, L.; Gilcrease, E.B.; Bhardwaj, A.; Cingolani, G.; Casjens, S.R. The tip of the tail needle affects the rate of DNA delivery by bacteriophage P22. PLoS ONE 2013, 8, e70936. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Ara, T.; Hasegawa, M.; Takai, Y.; Okumura, Y.; Baba, M.; Datsenko, K.A.; Tomita, M.; Wanner, B.L.; Mori, H. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: The keio collection. Mol. Syst. Biol. 2006, 2, 2006.0008. [Google Scholar] [CrossRef] [PubMed]
- Freeman, G.G.; Philpot, J.S. The preparation and properties of a specific polysaccharide from bact. Typhosum Ty(2). Biochem. J. 1942, 36, 340–356. [Google Scholar] [CrossRef] [PubMed]
- Kemmer, G.; Keller, S. Nonlinear least-squares data fitting in excel spreadsheets. Nat. Protoc. 2010, 5, 267–281. [Google Scholar] [CrossRef] [PubMed]
- Provencher, S.W. Contin—A general purpose constrained regularization program for inverting noisy linear equations. Comput. Phys. Commun. 1982, 27, 229–242. [Google Scholar] [CrossRef]
- Parent, K.N.; Erb, M.L.; Cardone, G.; Nguyen, K.; Gilcrease, E.B.; Porcek, N.B.; Pogliano, J.; Baker, T.S.; Casjens, S.R. OmpA and OmpC are critical host factors for bacteriophage Sf6 entry in Shigella. Mol. Microbiol. 2014, 92, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Andres, D.; Baxa, U.; Hanke, C.; Seckler, R.; Barbirz, S. Carbohydrate binding of salmonella phage P22 tailspike protein and its role during host cell infection. Biochem. Soc. Trans. 2010, 38, 1386–1389. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, A.; Rabsch, W.; Broeker, N.K.; Barbirz, S. Bacteriophage tailspike protein based assay to monitor phase variable glucosylations in salmonella o-antigens. BMC Microbiol. 2016, 16, 207. [Google Scholar] [CrossRef] [PubMed]
- Jann, B.; Shashkov, A.S.; Gupta, D.S.; Jann, K. The O18 antigens (lipopolysaccharides) of Escherichia coli. Structural characterization of the O18A, O18A1, O18B and O18B1-specific polysaccharides. Eur. J. Biochem. 1992, 210, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Patel, D.S.; Park, S.; Slusky, J.; Klauda, J.B.; Widmalm, G.; Im, W. Bilayer properties of lipid A from various gram-negative bacteria. Biophys. J. 2016, 111, 1750–1760. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed]
- Kintz, E.; Davies, M.R.; Hammarlof, D.L.; Canals, R.; Hinton, J.C.D.; van der Woude, M.W. A BTP1 prophage gene present in invasive non-typhoidal salmonella determines composition and length of the O-antigen of the lipopolysaccharide. Mol. Microbiol. 2015, 96, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.R.; Broadbent, S.E.; Harris, S.R.; Thomson, N.R.; van der Woude, M.W. Horizontally acquired glycosyltransferase operons drive salmonellae lipopolysaccharide diversity. PLoS Genet. 2013, 9, e1003568. [Google Scholar] [CrossRef] [PubMed]
- Bohm, K.; Porwollik, S.; Chu, W.; Dover, J.A.; Gilcrease, E.B.; Casjens, S.R.; McClelland, M.; Parent, K.N. Genes affecting progression of bacteriophage P22 infection in salmonella identified by transposon and single gene deletion screens. Mol. Microbiol. 2018, 108. [Google Scholar] [CrossRef] [PubMed]
- Cota, I.; Sanchez-Romero, M.A.; Hernandez, S.B.; Pucciarelli, M.G.; Garcia-del Portillo, F.; Casadesus, J. Epigenetic control of salmonella enterica O-antigen chain length: A tradeoff between virulence and bacteriophage resistance. PLoS Genet. 2015, 11, e1005667. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Margolin, W.; Molineux, I.J.; Liu, J. The bacteriophage T7 virion undergoes extensive structural remodeling during infection. Science 2013, 339, 576–579. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Margolin, W.; Molineux, I.J.; Liu, J. Structural remodeling of bacteriophage T4 and host membranes during infection initiation. Proc. Natl. Acad. Sci. USA 2015, 112, E4919–E4928. [Google Scholar] [CrossRef] [PubMed]
- McNulty, R.; Cardone, G.; Gilcrease, E.B.; Baker, T.S.; Casjens, S.R.; Johnson, J.E. Cryo-EM elucidation of the structure of bacteriophage P22 virions after genome release. Biophys. J. 2018, 114, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Chiaruttini, N.; de Frutos, M.; Augarde, E.; Boulanger, P.; Letellier, L.; Viasnoff, V. Is the in vitro ejection of bacteriophage DNA quasistatic? A bulk to single virus study. Biophys. J. 2010, 99, 447–455. [Google Scholar] [CrossRef] [PubMed]
- Kellermayer, M.S.Z.; Voros, Z.; Csik, G.; Herenyi, L. Forced phage uncorking: Viral DNA ejection triggered by a mechanically sensitive switch. Nanoscale 2018, 10, 1898–1904. [Google Scholar] [CrossRef] [PubMed]
- Baxa, U.; Steinbacher, S.; Miller, S.; Weintraub, A.; Huber, R.; Seckler, R. Interactions of phage P22 tails with their cellular receptor, salmonella O-antigen polysaccharide. Biophys. J. 1996, 71, 2040–2048. [Google Scholar] [CrossRef]
- Freiberg, A.; Morona, R.; Van den Bosch, L.; Jung, C.; Behlke, J.; Carlin, N.; Seckler, R.; Baxa, U. The tailspike protein of shigella phage Sf6—A structural homolog of salmonella phage P22 tailspike protein without sequence similarity in the beta-helix domain. J. Biol. Chem. 2003, 278, 1542–1548. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Garcia, V.; Bocanegra, R.; Pulido-Cid, M.; Martin-Benito, J.; Cuervo, A.; Carrascosa, J.L. Characterization of the initial steps in the T7 DNA ejection process. Bacteriophage 2015, 5, e1056904. [Google Scholar] [CrossRef] [PubMed]
- Olia, A.S.; Casjens, S.; Cingolani, G. Structure of phage P22 cell envelope–penetrating needle. Nat. Struct. Mol. Biol. 2007, 14, 1221–1226. [Google Scholar] [CrossRef] [PubMed]
- Bhardwaj, A.; Molineux, I.J.; Casjens, S.R.; Cingolani, G. Atomic structure of bacteriophage Sf6 tail needle knob. J. Biol. Chem. 2011, 286, 30867–30877. [Google Scholar] [CrossRef] [PubMed]
- Freeman, K.G.; Behrens, M.A.; Streletzky, K.A.; Olsson, U.; Evilevitch, A. Portal stability controls dynamics of DNA ejection from phage. J. Phys. Chem. B 2016, 120, 6421–6429. [Google Scholar] [CrossRef] [PubMed]
- De Frutos, M.; Letellier, L.; Raspaud, E. DNA ejection from bacteriophage T5: Analysis of the kinetics and energetics. Biophys. J. 2005, 88, 1364–1370. [Google Scholar] [CrossRef] [PubMed]
- Malojcic, G.; Andres, D.; Grabowicz, M.; George, A.H.; Ruiz, N.; Silhavy, T.J.; Kahne, D. LptE binds to and alters the physical state of LPS to catalyze its assembly at the cell surface. Proc. Natl. Acad. Sci. USA 2014, 111, 9467–9472. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.L.; Fleming, P.J.; Yeom, M.S.; Widmalm, G.; Klauda, J.B.; Fleming, K.G.; Im, W. E. coli outer membrane and interactions with OmpLA. Biophys. J. 2014, 106, 2493–2502. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Rokney, A.; Feeney, M.; Semsey, S.; Kessel, M.; Goldberg, M.B.; Adhya, S.; Oppenheim, A.B. Bacteriophage infection is targeted to cellular poles. Mol. Microbiol. 2008, 68, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Wu, E.L.; Engstrom, O.; Jo, S.; Stuhlsatz, D.; Yeom, M.S.; Klauda, J.B.; Widmalm, G.; Im, W. Molecular dynamics and NMR spectroscopy studies of E. coli lipopolysaccharide structure and dynamics. Biophys. J. 2013, 105, 1444–1455. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin of LPS Preparation a,b | Stokes Radius/nm |
|---|---|
| E. coli H TD2158 (O18A1) * | 96 |
| TSP digested, E coli H TD2158 (O18A1) | 54 |
| E. coli IHE3042 (O18A1) | 66 |
| E. coli DSM10809 (O18A) * | 68 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Broeker, N.K.; Kiele, F.; Casjens, S.R.; Gilcrease, E.B.; Thalhammer, A.; Koetz, J.; Barbirz, S. In Vitro Studies of Lipopolysaccharide-Mediated DNA Release of Podovirus HK620. Viruses 2018, 10, 289. https://doi.org/10.3390/v10060289
Broeker NK, Kiele F, Casjens SR, Gilcrease EB, Thalhammer A, Koetz J, Barbirz S. In Vitro Studies of Lipopolysaccharide-Mediated DNA Release of Podovirus HK620. Viruses. 2018; 10(6):289. https://doi.org/10.3390/v10060289
Chicago/Turabian StyleBroeker, Nina K., Franziska Kiele, Sherwood R. Casjens, Eddie B. Gilcrease, Anja Thalhammer, Joachim Koetz, and Stefanie Barbirz. 2018. "In Vitro Studies of Lipopolysaccharide-Mediated DNA Release of Podovirus HK620" Viruses 10, no. 6: 289. https://doi.org/10.3390/v10060289
APA StyleBroeker, N. K., Kiele, F., Casjens, S. R., Gilcrease, E. B., Thalhammer, A., Koetz, J., & Barbirz, S. (2018). In Vitro Studies of Lipopolysaccharide-Mediated DNA Release of Podovirus HK620. Viruses, 10(6), 289. https://doi.org/10.3390/v10060289