Non-Structural Protein NSm of Tomato Spotted Wilt Virus Is an Avirulence Factor Recognized by Resistance Genes of Tobacco and Tomato via Different Elicitor Active Sites

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plasmid Construction
2.2. Plant Materials, Virus Inoculation, and Agroinfiltration
2.3. RNA Isolation, RT-PCR, and Quantitative Real-time PCR (qRT-PCR)
2.4. DAB Staining
2.5. Protein Extraction and Western Blot Analysis
2.6. Enzyme-linked Immunosorbent Assay (ELISA)
3. Results
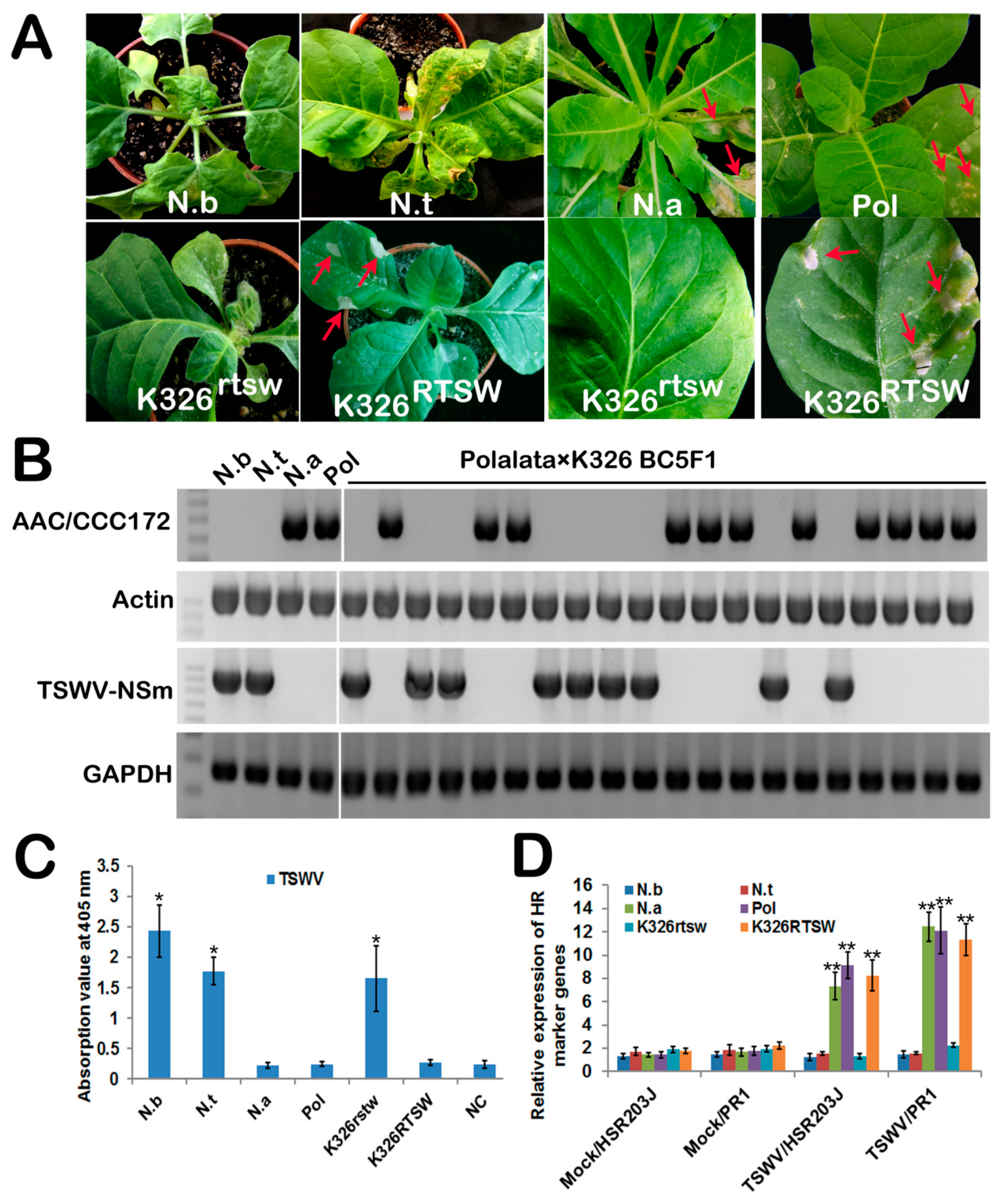
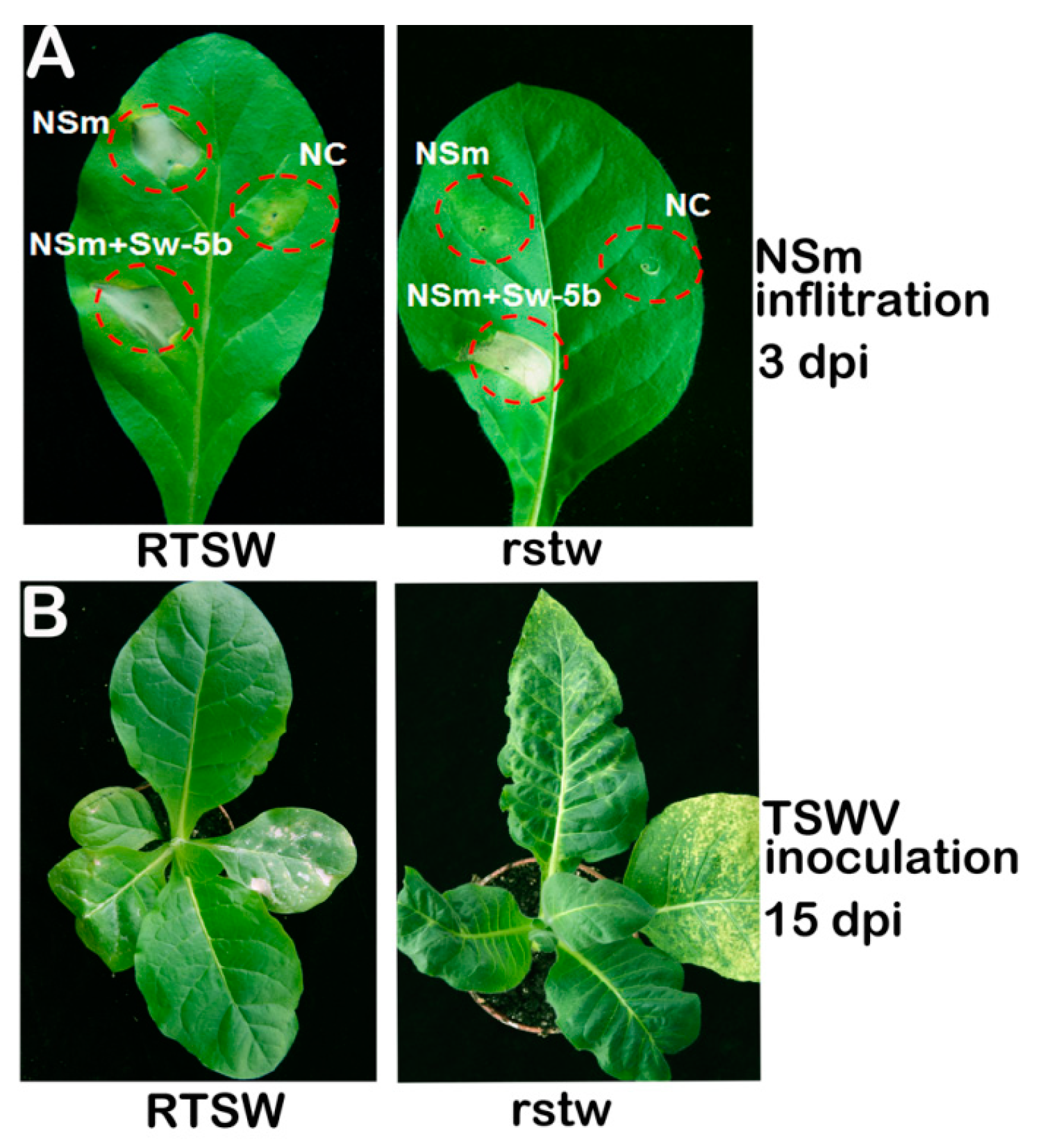
3.1. Tobacco RTSW Confers Resistance to TSWV by a Hypersensitive Response (HR)
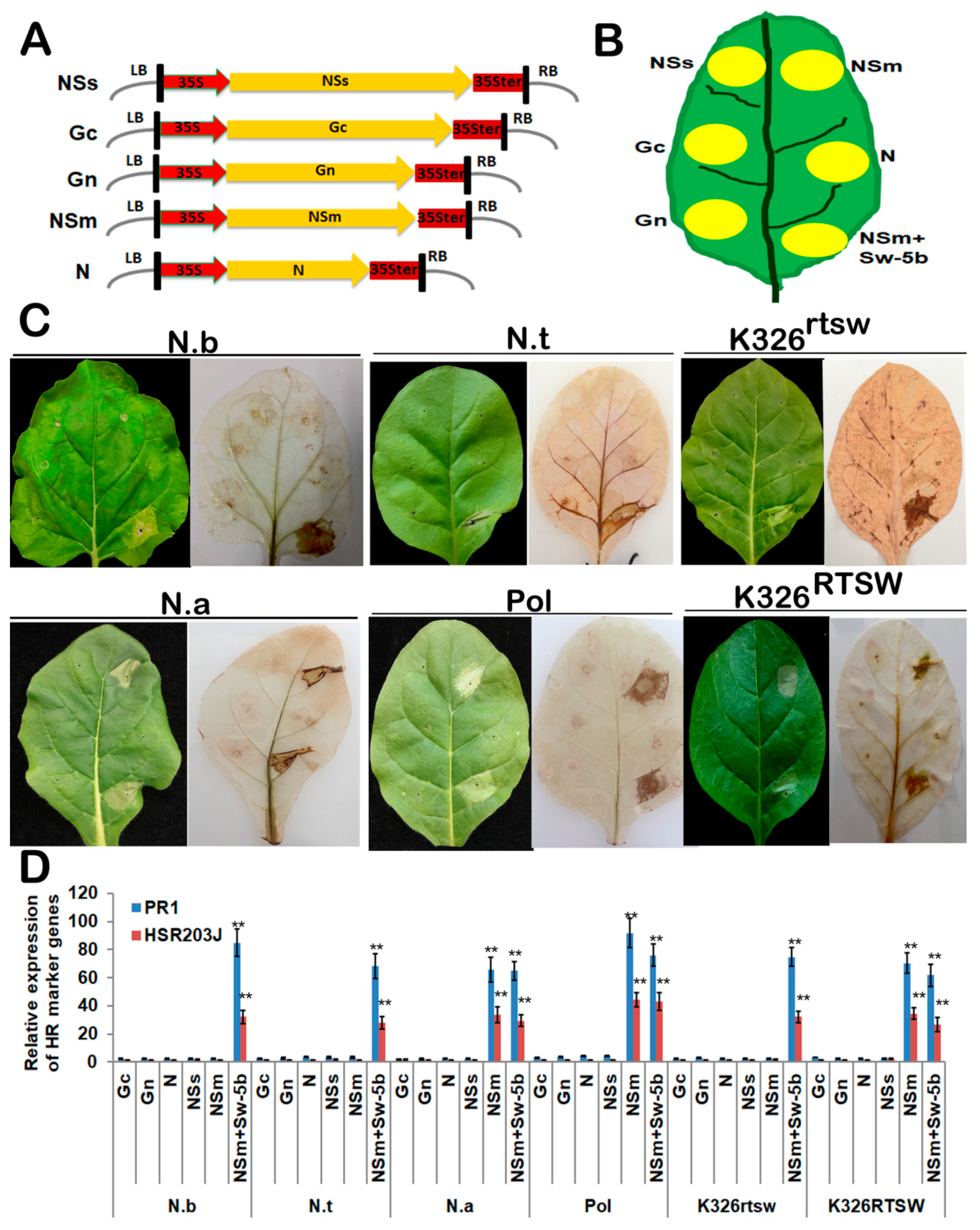
3.2. NSm Protein is an Elicitor of RTSW-based Resistance to TSWV
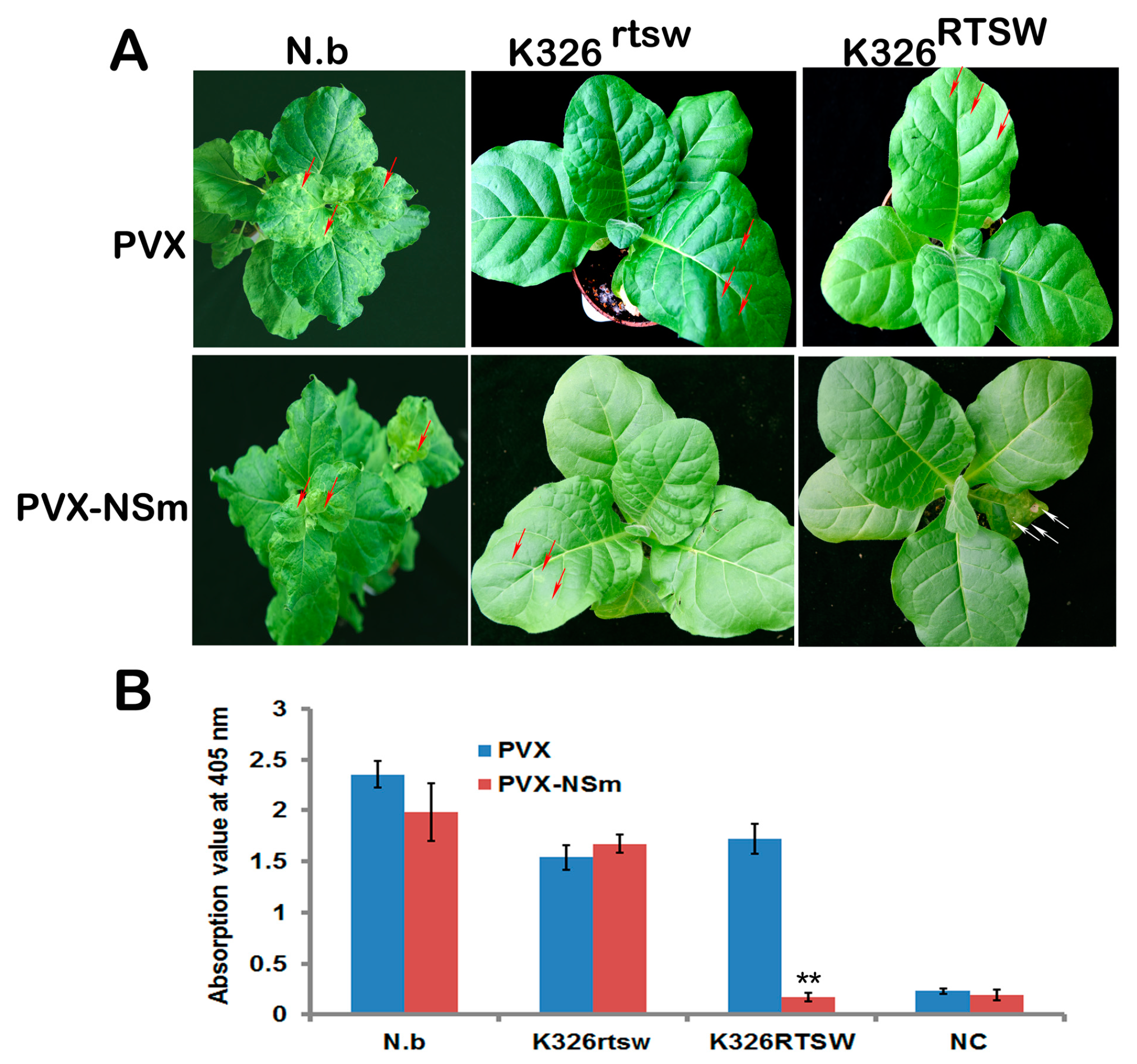
3.3. TSWV NSm Elicits HR-type Cell Death in a PVX Expression Assay Against RTSW Background
3.4. Role of NSm in HR Induction is Uncoupled from its Role in Plasmodesmata (PD) Targeting and Intercellular Movement
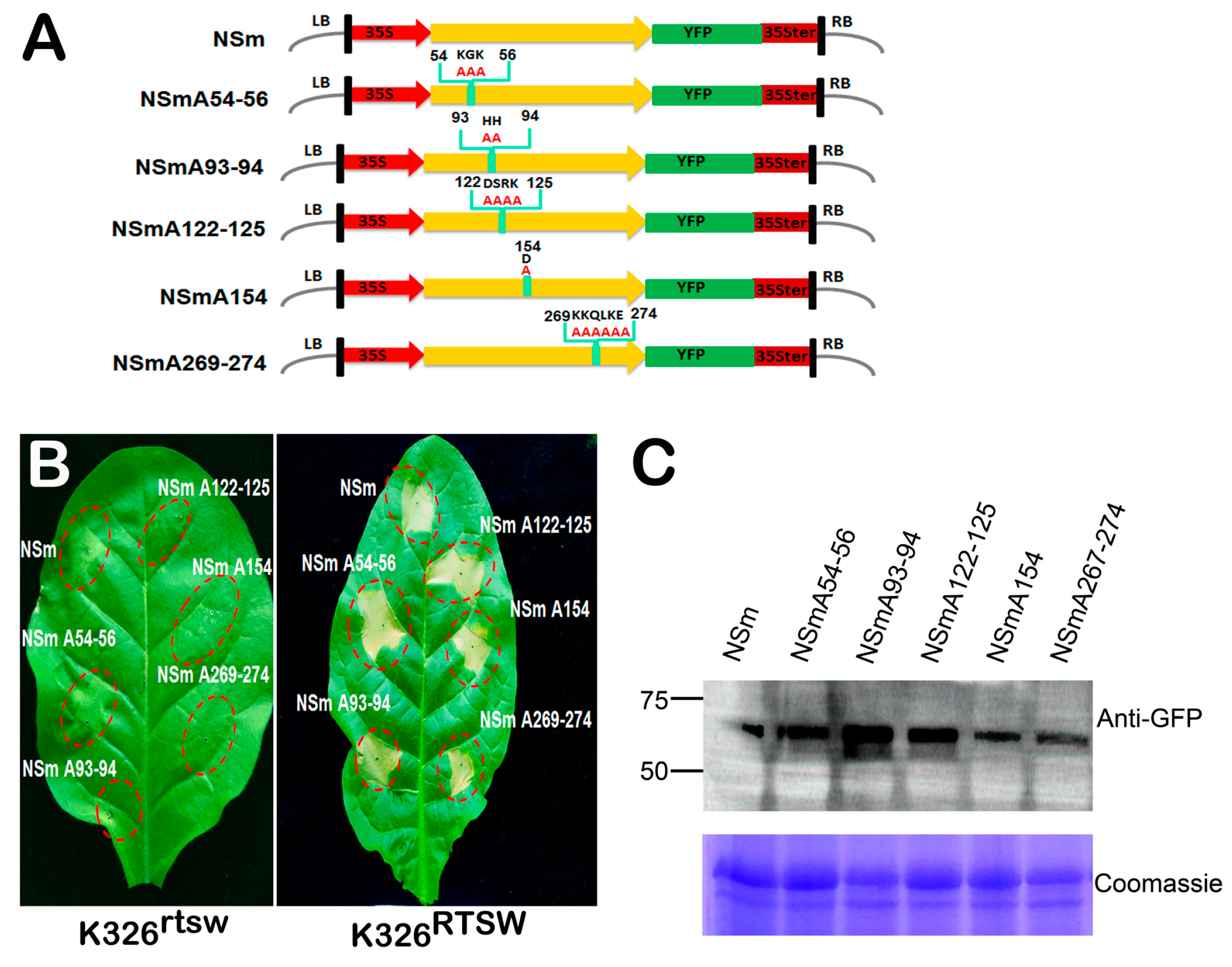
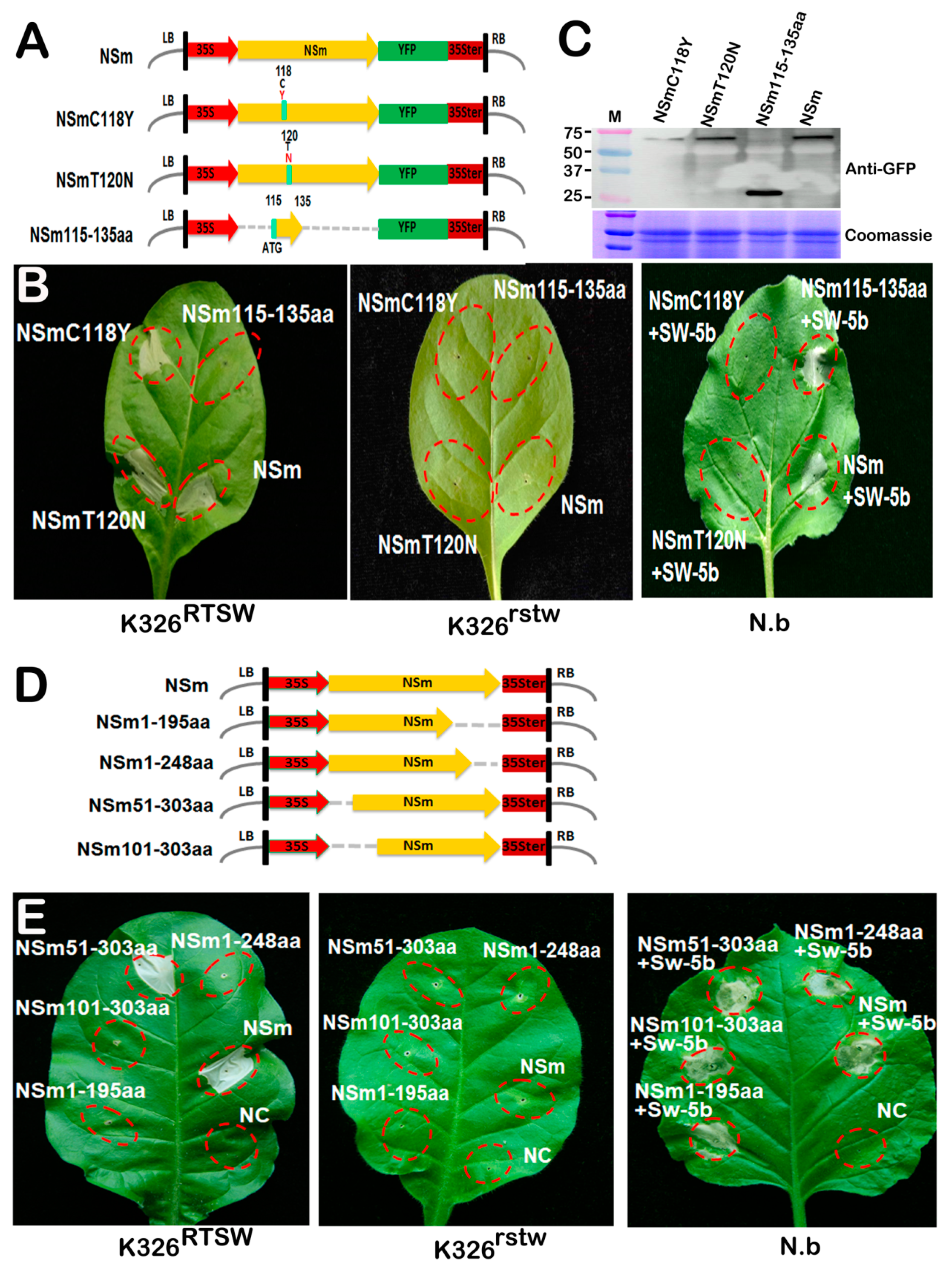
3.5. Key Amino Acids and a Conserved Motif Required for NSm-Sw-5b Interaction are Not Essential for RTSW Resistance
3.6. Application of the NSm Gene in Resistance Identification
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′→3′) |
|---|---|
| Primers used for the construction of expression vectors | |
| NSmBamHIF | CGGGATCCATGTTGACTTTTTTTGGTA |
| NSmBamHIR | CGGGATCCTATCTCATCAAAAGATAACT |
| GnBamHIF | CCCGGATCCATGAGAATTCTAAAACTA |
| GnBamHIR | CCCGGATCCTCATTCCATGCTAGTCCACT |
| GcBamHIF | CGGGATCCATGTGGTTCCATCTAATAGTGAAC |
| GcBamHIR | CGGGATCCTCAGACAAGGTGAGAGAAATC |
| NSsBamHIF | CGGGATCCATGTCTTCAAGTGTTTATGAG |
| NSsBamHIR | CGGGATCCTTATTTTGATCCTGAAGCATATG |
| NBamHIF | CGGGATCCATGTCTAAGGTTAAGCTCACT |
| NBamHIR | CGGGATCCTTAAGCAAGTTCTGCAAGT |
| NSmCalIF | GGCATCGATATGTTGACTTTTTTTGGTA |
| NSmSalIR | GGCGTCGACCTATATCTCATCAAAAGATAAC |
| Primers used for qRT-PCR | |
| PR1qRTF | AGCTCGTGCAGATGTAGGTGTAGA |
| PR1qRTR | TTCGCCGTATTGACCATGAGAATGT |
| HSR203JqRTF | CGCAATCATCGTCTCCGTCTTCC |
| HSR203JqRTR | CTCCGATAAGACCGCACGAACC |
| GAPDHF | GCAGTGAACGACCCATTTATCTC |
| GAPDHR | AACCTTCTTGGCACCACCCT |
| Primers used for genotyping | |
| S-AAC/CCC172F | AGCTTCTTTTCTCTTTCCATTTTT |
| S-AAC/CCC172R | CAGAAGAAAAACTGCTGGAGCTAT |
| Actin1000F | ATGGGCCAGAAGGATGCTTATG |
| Actin1000R | CTGCTGGAATGTGCTGAGAGAG |
References
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Hogenhout, S.A.; Van der Hoorn, R.A.L.; Terauchi, R.; Kamoun, S. Emerging concepts in effector biology of plant-associated organisms. Mol. Plant Microbe Interact. 2009, 22, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Kamoun, S. Groovy times: Filamentous pathogen effectors revealed. Curr. Opin. Plant Biol. 2007, 10, 358–365. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, B.C.; Calil, I.P.; Machado, J.P.B.; Santos, A.A.; Fontes, E.P.B. Immune receptors and co-receptors in antiviral innate immunity in plants. Front. Microbiol. 2017, 7, 2139. [Google Scholar] [CrossRef] [PubMed]
- Andolfo, G.; Jupe, F.; Witek, K.; Etherington, G.; Ercolano, M.; Jones, J. Defining the full tomato NB-LRR resistance gene repertoire using genomic and cDNA RenSeq. BMC Plant Biol. 2014, 14, 120. [Google Scholar] [CrossRef] [PubMed]
- Jupe, F.; Witek, K.; Verweij, W.; Sliwka, J.; Pritchard, L.; Etherington, G.J.; Maclean, D.; Cock, P.J.; Leggett, R.M.; Bryan, G.J.; et al. Resistance gene enrichment sequencing (RenSeq) enables reannotation of the NB-LRR gene family from sequenced plant genomes and rapid mapping of resistance loci in segregating populations. Plant J. 2013, 76, 530–544. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.; Kim, S.; Yeom, S.-I.; Choi, D. Genome-wide comparative analyses reveal the dynamic evolution of nucleotide-binding leucine-rich repeat gene family among Solanaceae plants. Front. Plant Sci. 2016, 7, 1205. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, G.; Mazzucotelli, E.; Marone, D.; Crosatti, C.; Michelotti, V.; Valè, G.; Mastrangelo, A.M. Regulation and evolution of NLR genes: A close interconnection for plant immunity. Int. J. Mol. Sci. 2018, 19, 1662. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.Q.; Harrach, B.; Harrison, R.L.; Knowles, N.J.; Kropinski, A.M.; Krupovic, M.; Kuhn, J.H.; Mushegian, A.R.; et al. Changes to taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses. Arch. Virol. 2017, 162, 2505–2538. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.E.; Whitfield, A.E. The genus Tospovirus: Emerging bunyaviruses that threaten food security. Annu. Rev. Virol. 2016, 3, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Turina, M.; Kormelink, R.; Resende, R.O. Resistance to Tospoviruses in vegetable crops: Epidemiological and molecular aspects. Annu. Rev. Phytopathol. 2016, 54, 347–371. [Google Scholar] [CrossRef] [PubMed]
- Scholthof, K.B.; Adkins, S.; Czosnek, H.; Palukaitis, P.; Jacquot, E.; Hohn, T.; Hohn, B.; Saunders, K.; Candresse, T.; Ahiquist, P.; et al. Top 10 plant viruses in molecular plant pathology. Mol. Plant Pathol. 2011, 12, 938–954. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, A.E.; Ullman, D.E.; German, T.L. Tospovirus-thrips interactions. Annu. Rev. Phytopathol. 2005, 43, 459–489. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zhu, Q.; Chen, Y.; Liu, Y.-G. CRISPR/Cas9 platforms for genome editing in plants: Developments and applications. Mol. Plant 2016, 9, 961–974. [Google Scholar] [CrossRef] [PubMed]
- Spassova, M.I.; Prins, T.W.; Folkertsma, R.T.; Klein-Lankhorst, R.M.; Hille, J.; Goldbach, R.W. The tomato gene Sw5 is a member of the coiled coil, nucleotide binding, leucine-rich repeat class of plant resistance genes and confers resistance to TSWV in tobacco. Mol. Breeding 2001, 7, 151–161. [Google Scholar] [CrossRef]
- Zhu, M.; Jiang, L.; Bai, B.; Zhao, W.; Chen, X.; Li, J.; Liu, Y.; Chen, Z.; Wang, B.; Wang, C.; et al. The intracellular immune receptor Sw-5b confers broad-spectrum resistance to Tospoviruses through recognition of a conserved 21-amino acid viral effector epitope. Plant Cell 2017, 29, 2214–2232. [Google Scholar] [CrossRef] [PubMed]
- Hallwass, M.; de Oliveira, A.S.; Dianese, E.D.; Lohuis, D.; Boiteux, L.S.; Inoue-Nagata, A.K.; Resende, R.O.; Kormelink, R. The Tomato spotted wilt virus cell-to-cell movement protein (NSM) triggers a hypersensitive response in Sw-5-containing resistant tomato lines and in Nicotiana benthamiana transformed with the functional Sw-5b resistance gene copy. Mol. Plant Pathol. 2014, 15, 871–880. [Google Scholar] [PubMed]
- Peiró, A.; Canizares, M.C.; Rubio, L.; Lopez, C.; Moriones, E.; Aramburu, J.; Sánchez-Navarro, J. The movement protein (NSm) of Tomato spotted wilt virus is the avirulence determinant in the tomato Sw-5 gene-based resistance. Mol. Plant Pathol. 2014, 15, 802–813. [Google Scholar] [CrossRef] [PubMed]
- De Ronde, D.; Butterbach, P.; Lohuis, D.; Hedil, M.; van Lent, J.W.; Kormelink, R. Tsw gene-based resistance is triggered by a functional RNA silencing suppressor protein of the Tomato spotted wilt virus. Mol. Plant Pathol. 2013, 14, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Opoka, B. Resistance of wild Nicotiana species to Lycopersicon 3. Hodowia Rosl. Helimatyzaga Nasienn 1969, 13, 83–87. [Google Scholar]
- Gajos, Z. Polalta-a tobacco variety resistant to Tomato spotted wilt virus (TSWV) and black root rot (Thielaviopsis basicola Ferr). Biul. CLPT 1988, 1, 7–25. [Google Scholar]
- Laskowska, D.; Doroszewska, T.; Depta, A.; Kursa, K.; Olszak-Przybyś, H.; Czubacka, A. A survey of Nicotiana germplasm for resistance to Tomato spotted wilt virus (TSWV). Euphytica 2013, 193, 207–219. [Google Scholar] [CrossRef]
- Moon, H.; Nicholson, J.S. AFLP and SCAR markers linked to Tomato spotted wilt virus resistance in tobacco. Crop Sci. 2007, 47, 1887–1894. [Google Scholar] [CrossRef]
- Thomas, C.L.; Jones, L.; Baulcombe, D.C.; Maule, A.J. Size constraints for targeting post-transcriptional gene silencing and for RNA-directed methylation in Nicotiana benthamiana using a potato virus X vector. Plant J. 2001, 25, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Jiang, L.; Feng, Z.; Chen, X.; Huang, Y.; Xue, F.; Huang, C.; Liu, Y.; Li, F.; Liu, Y.; et al. Plasmodesmata targeting and intercellular trafficking of Tomato spotted wilt tospovirus movement protein NSm is independent of its function in HR induction. J. Gen. Virol. 2016, 97, 1990–1997. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.J.; Liu, Y.; Yu, H.Q.; Liu, B.Z.; Qing, L. Bidens pilosa is a natural host of Tomato spotted wilt virus in Yunnan Province, China. Plant Dis. 2016, 100, 1957–1957. [Google Scholar] [CrossRef]
- Huang, C.; Cun, Y.; Yu, H.; Tong, Z.; Xiao, B.; Song, Z.; Wang, B.; Li, Y.; Liu, Y. Transcriptomic profile of tobacco in response to Tomato zonate spot orthotospovirus infection. Virol. J. 2017, 14, 153. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Hou, H.; Shen, Q.; Cai, X.; Sunter, G.; Zhou, X. RepA protein encoded by Oat dwarf virus elicits a temperature-sensitive hypersensitive response–type cell death that involves jasmonic acid–dependent signaling. Mol. Plant Microbe Interact. 2015, 29, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yu, C.; Yang, C.; Zhou, X. Monoclonal antibodies against the recombinant nucleocapsid protein of Tomato spotted wilt virus and its application in virus detection. J. Phytopathol. 2009, 157, 344–349. [Google Scholar] [CrossRef]
- Takahashi, Y.; Uehara, Y.; Berberich, T.; Ito, A.; Saitoh, H.; Miyazaki, A.; Terauchi, R.; Kusano, T. A subset of hypersensitive response marker genes, including HSR203J, is the downstream target of a spermine signal transduction pathway in tobacco. Plant J. 2004, 40, 586–595. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.T.; Yao, N. The role and regulation of programmed cell death in plant–pathogen interactions. Cell Microbiol. 2004, 6, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Xue, F.; Xu, M.; Chen, X.; Zhao, W.; Garcia-Murria, M.J.; Mingarro, I.; Liu, Y.; Huang, Y.; Jiang, L.; et al. The ER-Membrane transport system is critical for intercellular trafficking of the NSm movement protein and Tomato spotted wilt tospovirus. PLoS Pathog. 2016, 12, e1005443. [Google Scholar] [CrossRef] [PubMed]
- Kormelink, R.; Storms, M.; Van Lent, J.; Peters, D.; Goldbach, R. Expression and subcellular location of the NSM protein of tomato spotted wilt virus (TSWV), a putative viral movement protein. Virology 1994, 200, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Lewandowski, D.J.; Adkins, S. The tubule-forming NSm protein from Tomato spotted wilt virus complements cell-to-cell and long-distance movement of Tobacco mosaic virus hybrids. Virology 2005, 342, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Lewandowski, D.J.; Hilf, M.E.; Adkins, S. Identification of domains of the Tomato spotted wilt virus NSm protein involved in tubule formation, movement and symptomatology. Virology 2009, 390, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Lopez, C.; Aramburu, J.; Galipienso, L.; Soler, S.; Nuez, F.; Rubio, L. Evolutionary analysis of tomato Sw-5 resistance-breaking isolates of Tomato spotted wilt virus. J. Gen. Virol. 2011, 92, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Flor, H.H. Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 1971, 9, 275–296. [Google Scholar] [CrossRef]
- Feng, Z.; Chen, X.; Bao, Y.; Dong, J.; Zhang, Z.; Tao, X. Nucleocapsid of Tomato spotted wilt tospovirus forms mobile particles that traffic on an actin/endoplasmic reticulum network driven by myosin XI-K. New Phytol. 2013, 200, 1212–1224. [Google Scholar] [CrossRef] [PubMed]
- Paape, M.; Solovyev, A.G.; Erokhina, T.N.; Minina, E.A.; Schepetilnikov, M.V.; Lesemann, D.E.; Schiemann, J.; Morozov, S.Y.; Kellmann, J.-W. At-4/1, an interactor of the Tomato spotted wilt virus movement protein, belongs to a new family of plant proteins capable of directed intra- and intercellular trafficking. Mol. Plant Microbe Interact. 2006, 19, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Wang, G.; Ma, S.; Xie, X.; Wu, X.; Zhang, X.; Wu, Y.; Zhao, P.; Xia, Q. CRISPR/Cas9-mediated targeted mutagenesis in Nicotiana tabacum. Plant Mol. Biol. 2015, 87, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Zhang, T.; Yao, M.; Feng, Z.; Miriam, K.; Wu, J.; Zhou, X.; Tao, X. The 2a protein of Cucumber mosaic virus induces a hypersensitive response in cowpea independently of its replicase activity. Virus Res. 2012, 170, 169–173. [Google Scholar] [CrossRef] [PubMed]
- De Ronde, D.; Pasquier, A.; Ying, S.; Butterbach, P.; Lohuis, D.; Kormelink, R. Analysis of Tomato spotted wilt virus NSs protein indicates the importance of the N-terminal domain for avirulence and RNA silencing suppression. Mol. Plant Pathol. 2014, 15, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Leastro, M.O.; de Oliveira, A.S.; Pallás, V.; Sánchez-Navarro, J.A.; Kormelink, R.; Resende, R.O. The NSm proteins of phylogenetically related tospoviruses trigger Sw-5b–mediated resistance dissociated of their cell-to-cell movement function. Virus Res. 2017, 240, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Mandal, B.; Pappu, H.R.; Culbreath, A.K. Factors affecting mechanical transmission of Tomato spotted wilt virus to peanut (Arachis hypogaea). Plant Dis. 2001, 85, 1259–1263. [Google Scholar] [CrossRef]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, C.; Liu, Y.; Yu, H.; Yuan, C.; Zeng, J.; Zhao, L.; Tong, Z.; Tao, X. Non-Structural Protein NSm of Tomato Spotted Wilt Virus Is an Avirulence Factor Recognized by Resistance Genes of Tobacco and Tomato via Different Elicitor Active Sites. Viruses 2018, 10, 660. https://doi.org/10.3390/v10110660
Huang C, Liu Y, Yu H, Yuan C, Zeng J, Zhao L, Tong Z, Tao X. Non-Structural Protein NSm of Tomato Spotted Wilt Virus Is an Avirulence Factor Recognized by Resistance Genes of Tobacco and Tomato via Different Elicitor Active Sites. Viruses. 2018; 10(11):660. https://doi.org/10.3390/v10110660
Chicago/Turabian StyleHuang, Changjun, Yong Liu, Haiqin Yu, Cheng Yuan, Jianmin Zeng, Lu Zhao, Zhijun Tong, and Xiaorong Tao. 2018. "Non-Structural Protein NSm of Tomato Spotted Wilt Virus Is an Avirulence Factor Recognized by Resistance Genes of Tobacco and Tomato via Different Elicitor Active Sites" Viruses 10, no. 11: 660. https://doi.org/10.3390/v10110660
APA StyleHuang, C., Liu, Y., Yu, H., Yuan, C., Zeng, J., Zhao, L., Tong, Z., & Tao, X. (2018). Non-Structural Protein NSm of Tomato Spotted Wilt Virus Is an Avirulence Factor Recognized by Resistance Genes of Tobacco and Tomato via Different Elicitor Active Sites. Viruses, 10(11), 660. https://doi.org/10.3390/v10110660