H5N1 Virus Evolution in Europe—An Updated Overview

Abstract
:1. Introduction
2. The emergence of divergent genetic clades
3. The westward spread of clade 2.2

{kind=link}
| Country | Animal category | No. outbreaks in poultry** |
|---|---|---|
| Albania | Poultry | 3 |
| Austria | Wild birds | |
| Belgium | Wild birds (illegal trade) | |
| Bosnia Herzegovina | Wild birds | |
| Bulgaria | Wild birds | |
| Croatia | Wild birds | |
| Czech Republic | Poultry; Wild birds | 4 |
| Denmark | Poultry; Wild birds | 1 |
| France | Poultry ; Wild birds | 1 |
| Germany | Poultry; Wild birds | 8 |
| Greece | Wild birds | |
| Hungary | Poultry; Wild birds | 9 |
| Italy | Wild birds | |
| Poland | Poultry; Wild birds | 10 |
| Romania | Poultry; Wild birds | 163 |
| Russia | Poultry; Wild birds | 149 |
| Serbia & Montenegro | Poultry; Wild birds | 1 |
| Slovakia | Wild birds | |
| Slovenia | Wild birds | |
| Spain | Wild birds | |
| Sweden | Poultry; Wild birds | 1 |
| Switzerland | Wild birds | |
| Ukraine | Poultry; Wild birds | 42 |
| United Kingdom | Poultry; Wild birds | 3 |
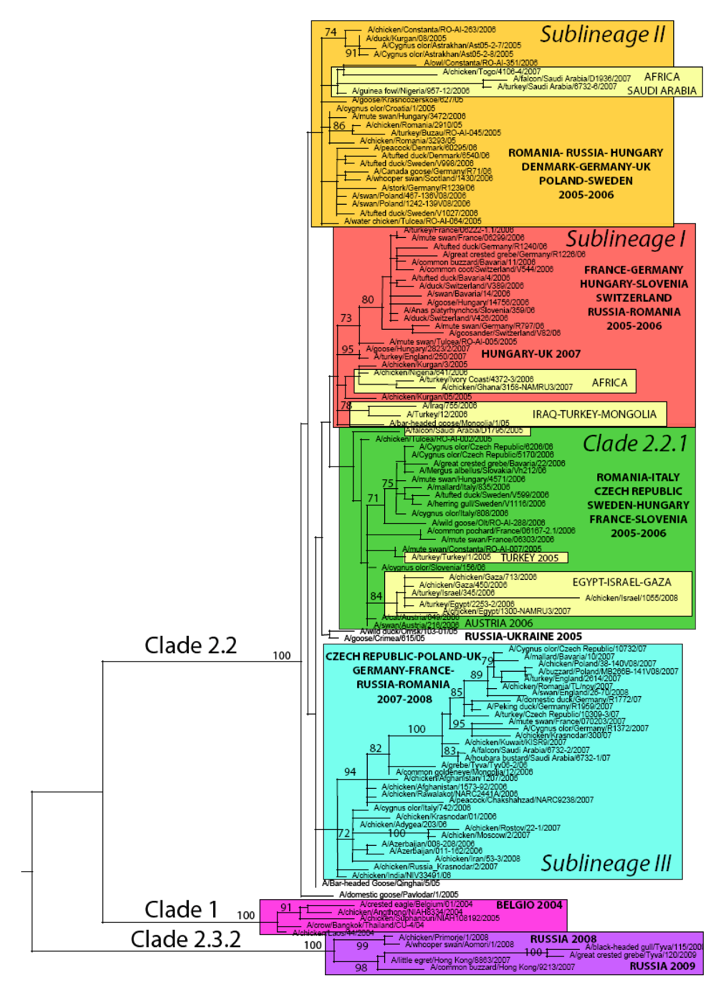
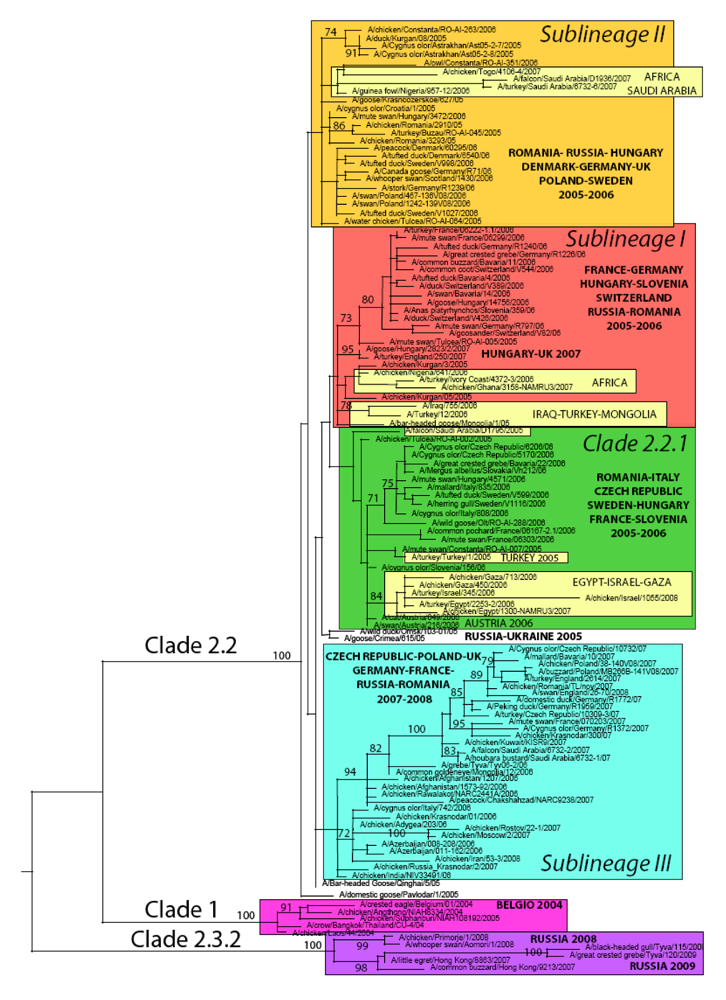
4. Genetic diversity of HPAI A/H5N1 viruses in Europe
5. Updating the molecular epidemiology of HPAI A/H5N1 in Europe
5.1. Phylogeny
| Year | ||||||
|---|---|---|---|---|---|---|
| Country | 2004 | 2005 | 2006 | 2007 | 2008 | 2009 |
| Austria | 2.2.1 | |||||
| Belgium | 1 | |||||
| Croatia | 2.2 Sub II | |||||
| Czech Republic | 2.2.1 | 2.2 Sub III | ||||
| Danimark | 2.2 Sub II | |||||
| France | 2.2 Sub I | 2.2 Sub III | ||||
| Germany | 2.2 Sub I 2.2 Sub II 2.2.1 | 2.2 Sub III | ||||
| Hungary | 2.2 Sub I 2.2 Sub II 2.2.1 | 2.2 Sub I | ||||
| Italy | 2.2.1 2.2 Sub III | |||||
| Romania | 2.2.1 2.2 Sub I 2.2 Sub II | 2.2.1 2.2 Sub I 2.2 Sub II | 2.2 Sub III | |||
| Russia | 2.2 Sub I 2.2 Sub II 2.2.1 | 2.2 Sub III | 2.2 Sub III | 2.3.2 | 2.3.2 | |
| Slovakia | 2.2.1 | |||||
| Slovenia | 2.2.1 2.2 Sub I | |||||
| Sweden | 2.2 Sub II 2.2.1 | |||||
| Switzerland | 2.2 Sub I | |||||
| Ukraine | 2.2 (undefined sublineage) | 2.2 Sub III | ||||
| UK | 2.2 Sub II | 2.2 Sub I 2.2 Sub III | 2.2 Sub III | |||
| Polonia | 2.2 Sub II | 2.2 Sub III | ||||
| Spain | 2.2.1 | |||||
5.2. Spatial migration of H5N1 virus in Europe
6. Conclusions
References
- Xu, X.; Subbarao, K.; Cox, N.J.; Guo, Y. Genetic characterization of the pathogenic influenza A/Goose/Guangdong/1/96 (H5N1) virus: similarity of its hemagglutinin gene to those of H5N1 viruses from the 1997 outbreaks in Hong Kong. Virology 1999, 261, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Capua, I.; Alexander, D.J. Avian influenza and human health. Acta Trop. 2002, 83, 1–6. [Google Scholar] [CrossRef]
- Sims, L.D.; Brown, I.H. Swayne, D.E., Ed.; Blackwell Publishing Professional: Ames, Iowa, USA, 2008; pp. 251–286.
- World Health Organization. H5N1 avian influenza: timeline of major events. Available online: http://www.who.int/csr/disease/avian_influenza/ai_timeline/en/index.html (accessed 27 August 2009).
- Guan, Y.; Smith, G.J.; Webby, R.; Webster, R.G. Molecular epidemiology of H5N1 avian influenza. Rev. Sci. Tech. 2009, 28, 39–47. [Google Scholar] [PubMed]
- Li, K.S.; Guan, Y.; Wang, J.; Smith, G.J.; Xu, K.M.; Duan, L.; Rahardjo, A.P.; Puthavathana, P.; Buranathai, C.; Nguyen, T.D.; Estoepangestie, A.T.; Chaisingh, A.; Auewarakul, P.; Long, H.T.; Hanh, N.T.; Webby, R.J.; Poon, L.L.; Chen, H.; Shortridge, K.F.; Yuen, K.Y.; Webster, R.G.; Peiris, J.S. Genesis of a highly pathogenic and potentially pandemic H5N1 influenza virus in eastern Asia. Nature. 2004, 430, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Vijaykrishna, D.; Bahl, J.; Riley, S.; Duan, L.; Zhang, J.X.; Chen, H.; Peiris, J.S.; Smith, G.J.; Guan, Y. Evolutionary dynamics and emergence of panzootic H5N1 influenza viruses. PLoS Pathog 2008, 4, e1000161. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization/World Organisation for Animal Health/Food and Agriculture Organization H5N1 Evolution Working Group. Toward a unified nomenclature system for highly pathogenic avian influenza virus (H5N1) [conference summary]. Emerg. Infect. Dis.[serial on the Internet]. 2008. Available online: Available online: http://www.cdc.gov/EID/content/14/7/e1.htm. [CrossRef]
- Chen, H.; Li, Y.; Li, Z.; Shi, J.; Shinya, K.; Deng, G.; Qi, Q.; Tian, G.; Fan, S.; Zhao, H.; Sun, Y.; Kawaoka, Y. Properties and dissemination of H5N1 viruses isolated during an influenza outbreak in migratory waterfowl in western China. J. Virol. 2006, 80, 5976–5983. [Google Scholar] [CrossRef] [PubMed]
- Editorial team. Highly pathogenic avian influenza reported in Russian bird populations. Euro Surveill. 2005, 10. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=2771.
- Salzberg, S.L.; Kingsford, C.; Cattoli, G.; Spiro, D.J.; Janies, D.A.; Aly, M.M.; Brown, I.H.; Couacy-Hymann, E.; De Mia, G.M.; Dung Do, H.; Guercio, A.; Joannis, T.; Maken Ali, A.S.; Osmani, A.; Padalino, I.; Saad, M.D.; Savić, V.; Sengamalay, N.A.; Yingst, S.; Zaborsky, J.; Zorman-Rojs, O.; Ghedin, E.; Capua, I. Genome analysis linking recent European and African influenza (H5N1) viruses. Emerg. Infect. Dis. 2007, 13, 713–718. [Google Scholar] [PubMed]
- World Organisation for Animal Health (OIE). Avian Influenza. Available online: Available online: http://www.oie.int/eng/info_ev/en_AI_avianinfluenza.htm (accessed 27 August 2009).
- World Health Organization. Cumulative Number of Confirmed Human Cases of Avian Influenza A/(H5N1) Reported to WHO . 11 8 2009. Available online: http://www.who.int/csr/disease/avian_influenza/country/cases_table_2009_08_11/en/index.html (accessed 27 August 2009).
- WHO/OIE/FAO H5N1 Evolution Working Group. Continuing progress towards a unified nomenclature for the highly pathogenic H5N1 avian influenza viruses: divergence of clade 2.2 viruses. Influenza Other Respi. Viruses 2009, 3, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Cattoli, G.; Monne, I.; Fusaro, A.; Joannis, T.M.; Lombin, L.H.; Aly, M.M.; Arafa, A.S.; Sturm-Ramirez, K.M.; Couacy-Hymann, E.; Awuni, J.A.; Batawui, K.B.; Awoume, K.A.; Aplogan, G.L.; Sow, A.; Ngangnou, A.C.; El Nasri Hamza, I.M.; Gamatié, D.; Dauphin, G.; Domenech, J.M.; Capua, I. Highly pathogenic avian influenza virus subtype H5N1 in Africa: a comprehensive phylogenetic analysis and molecular characterization of isolates (Epub 17 March 2009.). PLoS One 2009, 4, e4842. [Google Scholar] [CrossRef] [PubMed]
- Lipatov, A.S.; Evseenko, V.A.; Yen, H.L.; Zaykovskaya, A.V.; Durimanov, A.G.; Zolotykh, S.I.; Netesov, S.V.; Drozdov, I.G.; Onishchenko, G.G.; Webster, R.G.; Shestopalov, A.M. Influenza (H5N1) viruses in poultry, Russian Federation, 2005-2006. Emerg. Infect. Dis. 2007, 13, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Lvov, D.K.; Kaverin, N.V. Avian Influenza. Klenk, H.-D., Matrosovich, M.N., Stech, J., Eds.; 2008; Karger: Basel, Switzerland. [Google Scholar]
- Coulombier, D.; Paget, W.J.; Meijer, A.; Ganter, B. Highly pathogenic avian influenza reported to be spreading into western Russia. Euro Surveill. 2005, 10. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=2776.
- Anonym. Avian influenza suspected in Turkey: EC restricts imports. Vet. Rec. 2005, 157, 458. [Google Scholar]
- Anonym. H5N1 avian influenza virus reaches Europe. Vet. Rec. 2005, 157, 494. [Google Scholar] [PubMed]
- Van Borm, S.; Thomas, I.; Hanquet, G.; Lambrecht, B.; Boschmans, M.; Dupont, G.; Decaestecker, M.; Snacken, R.; van den Berg, T. Highly pathogenic H5N1 influenza virus in smuggled Thai eagles, Belgium. Emerg. Infect. Dis. 2005, 11, 702–705. [Google Scholar] [PubMed]
- Brown, I. Overview of animal outbreaks of AI H5N1 globally since 1997. FAO/OIE/WHO joint technical consultation on avian influenza at the human-animal interface; Verona, Italy, 7-9 10 2008. Available online: http://www.fao.org/docs/eims/upload//250653/aj140e00.pdf (accessed 27 August 2009).
- Bogner, P.; Capua, I.; Cox, N.J.; Lipman, D.J.; and others. A global initiative on sharing avian flu data. Nature 2006, 442, 981. [Google Scholar] [CrossRef]
- Bragstad, K.; Jørgensen, P.H.; Handberg, K.; Hammer, A.S.; Kabell, S.; Fomsgaard, A. First introduction of highly pathogenic H5N1 avian influenza A viruses in wild and domestic birds in Denmark, Northern Europe. Virol. J. 2007, 4, 43. [Google Scholar] [CrossRef] [PubMed]
- Weber, S.; Harder, T.; Starick, E.; Beer, M.; Werner, O.; Hoffmann, B.; Mettenleiter, T.C.; Mundt, E. Molecular analysis of highly pathogenic avian influenza virus of subtype H5N1 isolated from wild birds and mammals in northern Germany. J. Gen. Virol. 2007, 88, 554–558. [Google Scholar] [CrossRef] [PubMed]
- Rinder, M.; Lang, V.; Fuchs, C.; Hafner-Marx, A.; Bogner, K.H.; Neubauer, A.; Büttner, M.; Rinder, H. Genetic evidence for multi-event imports of avian influenza virus A(H5N1) into Bavaria, Germany. J. Vet. Diagn. Invest. 2007, 19, 279–282. [Google Scholar] [PubMed]
- Starick, E.; Beer, M.; Hoffmann, B.; Staubach, C.; Werner, O.; Globig, A.; Strebelow, G.; Grund, C.; Durban, M.; Conraths, F.J.; Mettenleiter, T.; Harder, T. Phylogenetic analyses of highly pathogenic avian influenza virus isolates from Germany in 2006 and 2007 suggest at least three separate introductions of H5N1 virus. Vet. Microbiol. 2008, 28, 243–252. [Google Scholar] [CrossRef]
- Harder, T.C.; Teuffert, J.; Starick, E.; Gethmann, J.; Grund, C.; Fereidouni, S.; Durban, M.; Bogner, K.H.; Neubauer-Juric, A.; Repper, R.; Hlinak, A.; Engelhardt, A.; Nöckler, A.; Smietanka, K.; Minta, Z.; Kramer, M.; Globig, A.; Mettenleiter, T.C.; Conraths, F.J.; Beer, M. Highly pathogenic avian influenza virus (H5N1) in frozen duck carcasses, Germany, 2007. Emerg. Infect. Dis. 2009, 15, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Globig, A.; Staubach, C.; Beer, M.; Köppen, U.; Fiedler, W.; Nieburg, M.; Wilking, H.; Starick, E.; Teifke, J.P.; Werner, O.; Unger, F.; Grund, C.; Wolf, C.; Roost, H.; Feldhusen, F.; Conraths, F.J.; Mettenleiter, T.C.; Harder, T.C. Epidemiological and ornithological aspects of outbreaks of highly pathogenic avian influenza virus H5N1 of Asian lineage in wild birds in Germany, 2006 and 2007. Transbound. Emerg. Dis. 2009, 56, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Gall-Reculé, G.L.; Briand, F.X.; Schmitz, A.; Guionie, O.; Massin, P.; Jestin, V. Double introduction of highly pathogenic H5N1 avian influenza virus into France in early 2006. Avian Pathol. 2008, 37, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Machova, J.; Hornickova, J.; Tomci, M.; Nagl, I.; Horyna, B.; Holko, I. Highly pathogenic avian influenza virus subtype H5N1 in Mute swans in the Czech Republic. Vet. Microbiol. 2007, 120, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Vostinakova, V.; Pindova, Z.; Hornickova, J.; Cernikova, L.; Sedlak, K.; Dirbakova, Z.; Machova, J. Molecular and phylogenetic analysis of the H5N1 avian influenza virus caused the first highly pathogenic avian influenza outbreak in poultry in the Czech Republic in 2007. Vet. Microbiol. 2009, 133, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Irvine, R.M.; Banks, J.; Londt, B.Z.; Lister, S.A.; Manvell, R.J.; Outtrim, L.; Russell, C.; Cox, W.J.; Ceeraz, V.; Shell, W.; Landeg, F.J.; Wilesmith, J.W.; Alexander, D.J.; Brown, I.H. Outbreak of highly pathogenic avian influenza caused by Asian lineage H5N1 virus in turkeys in Great Britain in January 2007. Vet. Rec. 2007, 161, 100–101. [Google Scholar] [PubMed]
- Kiss, I.; Gyarmati, P.; Zohari, S.; Ramsay, K.W.; Metreveli, G.; Weiss, E.; Brytting, M.; Stivers, M.; Lindström, S.; Lundkvist, A.; Nemirov, K.; Thorén, P.; Berg, M.; Czifra, G.; Belák, S. Molecular characterization of highly pathogenic H5N1 avian influenza viruses isolated in Sweden in 2006. Virol. J. 2008, 5, 113. [Google Scholar] [CrossRef] [PubMed]
- Barral, M.; Alvarez, V.; Juste, R.A.; Agirre, I.; Inchausti, I. First case of highly pathogenic H5N1 avian influenza virus in Spain. BMC Vet. Res. 2008, 4, 50. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, M.A.; Renzullo, S.; Baumer, A. Phylogenetic characterization of H5N1 highly pathogenic avian influenza viruses isolated in Switzerland in 2006. Virus Genes. 2008, 37, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Szeleczky, Z.; Dán, A.; Ursu, K.; Ivanics, E.; Kiss, I.; Erdélyi, K.; Belák, S.; Muller, C.P.; Brown, I.H.; Bálint, A. Four different sublineages of highly pathogenic avian influenza H5N1 introduced in Hungary in 2006-2007. Vet. Microbiol. 2009, 139, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Ducatez, M.F.; Olinger, C.M.; Owoade, A.A.; De Landtsheer, S.; Ammerlaan, W.; Niesters, H.G.; Osterhaus, A.D.; Fouchier, R.A.; Muller, C.P. Avian flu: multiple introductions of H5N1 in Nigeria. Nature. 2006, 442, 37. [Google Scholar] [CrossRef] [PubMed]
- Wilgenbusch, J.C.; Swofford, D. Inferring evolutionary trees with PAUP*. Curr. Protoc. Bioinformatics 2003, Chapter 6, Unit 6.4. [Google Scholar] [PubMed]
- Anonym. Viruses responsible for avian influenza in Suffolk and Hungary "essentially identical". Vet. Rec. 2007, 17, 201. [Google Scholar] [PubMed]
- Fusaro, A.M; Joannis, T.; Monne, I.; Salviato, A.; Yakubu, B.; Meseko, C.; Oladokun, T.; Fassina, S.; Capua, I.; Cattoli, G. Introduction into Nigeria of a distinct genotype of avian influenza virus (H5N1). Emerg. Infect. Dis. 2009, 15, 445–447. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Lu, L.; Liu, P.; Pybus, O.G. J. Infect 2004, 190 Erratum in: J. Infect. Dis. 2005, 191, 487., 1098–1108. [CrossRef]
- Carrington, C.V.; Foster, J.E.; Pybus, O.G.; Bennett, S.N.; Holmes, E.C. Invasion and maintenance of dengue virus type 2 and type 4 in the Americas. J. Virol. 2005, 79, 14680–14687. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Chmura, A.A.; Gibbons, D.W.; Fleischer, R.C.; Marra, P.P.; Daszak, P. Predicting the global spread of H5N1 avian influenza. Proc. Natl. Acad. Sci. U S A. 2006, 103, 19368–19373. [Google Scholar] [CrossRef] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Cattoli, G.; Fusaro, A.; Monne, I.; Capua, I. H5N1 Virus Evolution in Europe—An Updated Overview. Viruses 2009, 1, 1351-1363. https://doi.org/10.3390/v1031351
Cattoli G, Fusaro A, Monne I, Capua I. H5N1 Virus Evolution in Europe—An Updated Overview. Viruses. 2009; 1(3):1351-1363. https://doi.org/10.3390/v1031351
Chicago/Turabian StyleCattoli, Giovanni, Alice Fusaro, Isabella Monne, and Ilaria Capua. 2009. "H5N1 Virus Evolution in Europe—An Updated Overview" Viruses 1, no. 3: 1351-1363. https://doi.org/10.3390/v1031351
APA StyleCattoli, G., Fusaro, A., Monne, I., & Capua, I. (2009). H5N1 Virus Evolution in Europe—An Updated Overview. Viruses, 1(3), 1351-1363. https://doi.org/10.3390/v1031351