Influence of Natural and Anthropogenic Linear Canopy Openings on Forest Structural Patterns Investigated Using LiDAR


Abstract
:1. Introduction
2. Methods
2.1. Study Area
2.2. LiDAR Data
2.3. Site Selection
2.4. Data Analysis
2.4.1. Response Variables and Buffer Segments
2.4.2. Individual Tree Identification and Validation
2.4.3. Gap Definition and Delineation
2.4.4. Comparison of Forests around Linear Openings and Reference Forest
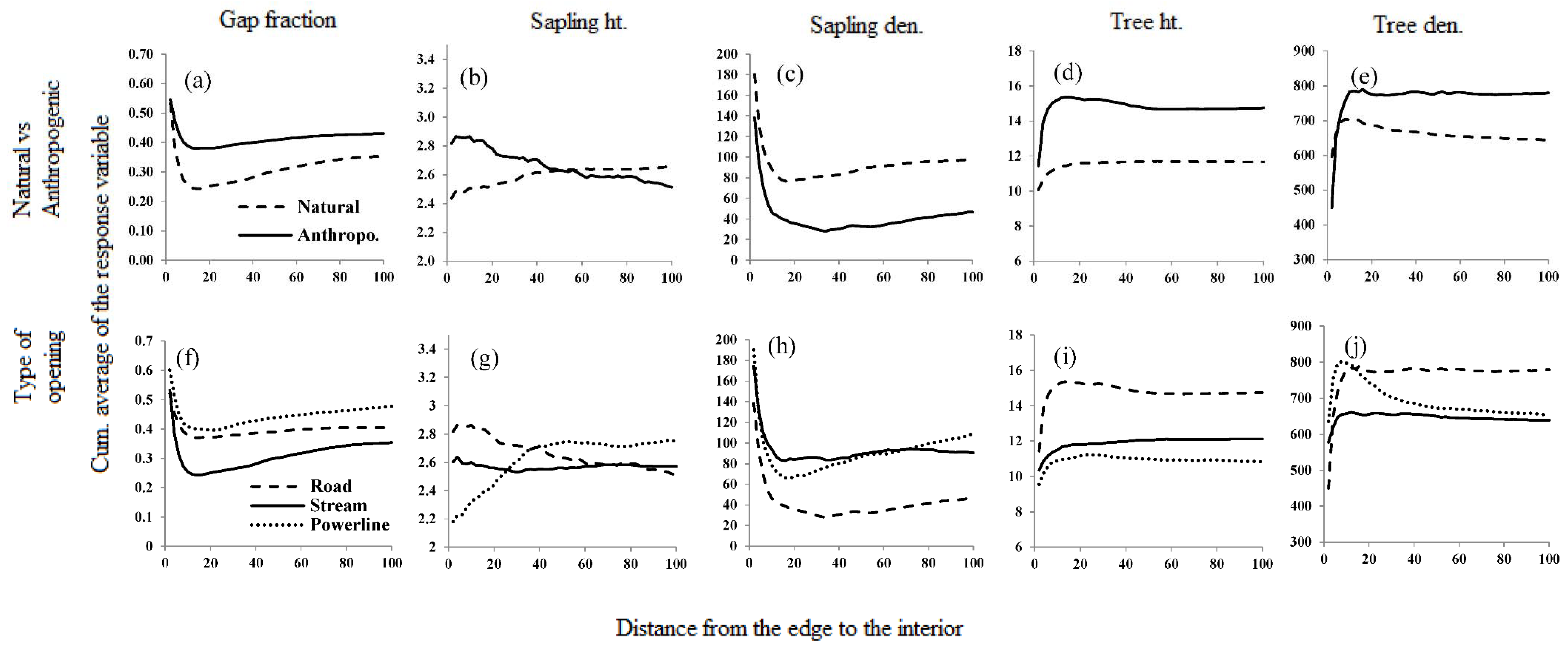
2.4.5. Depth of Influence—Average CUSUM
2.4.6. Magnitude of Edge Influence MEI
3. Results
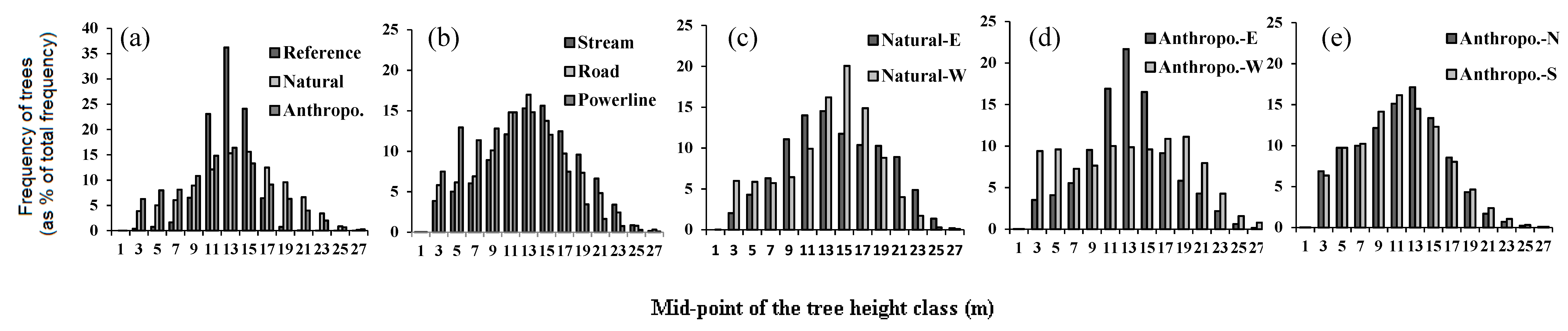
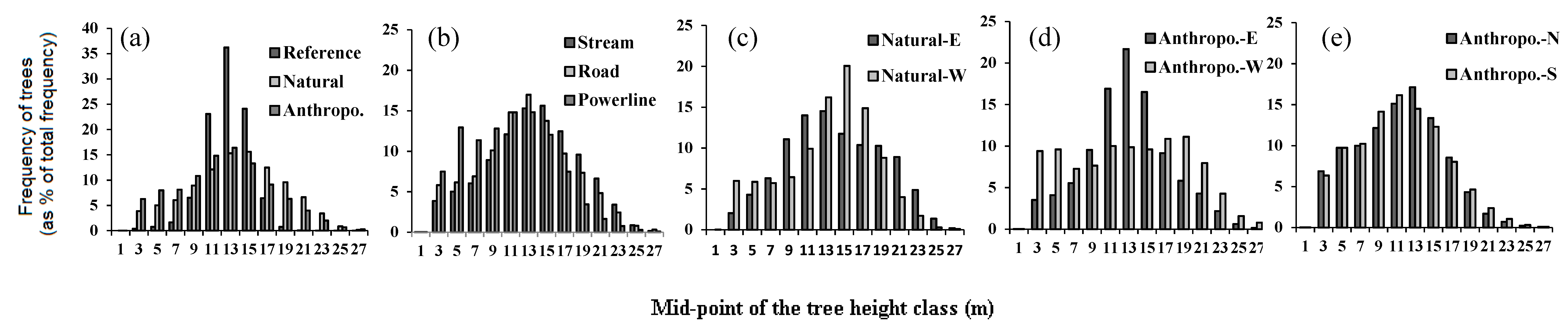
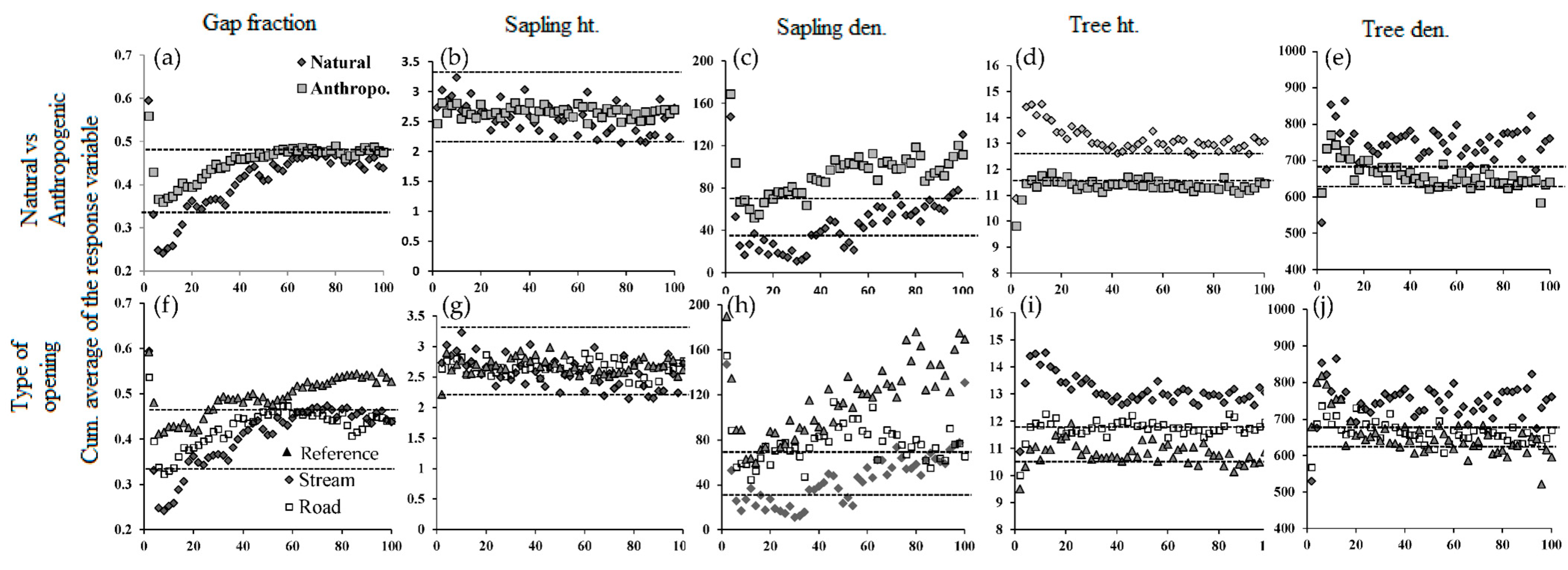
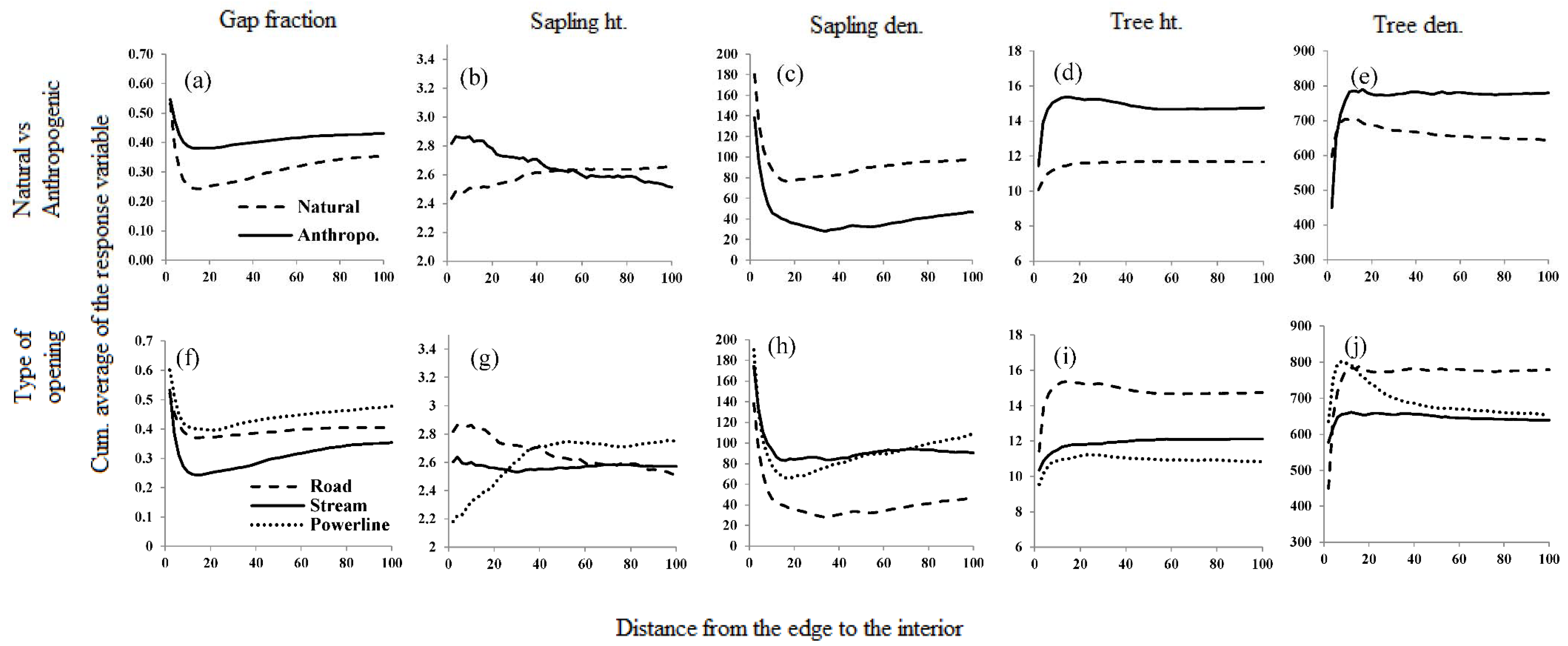
3.1. Reference Forest vs. Forests around Linear Openings
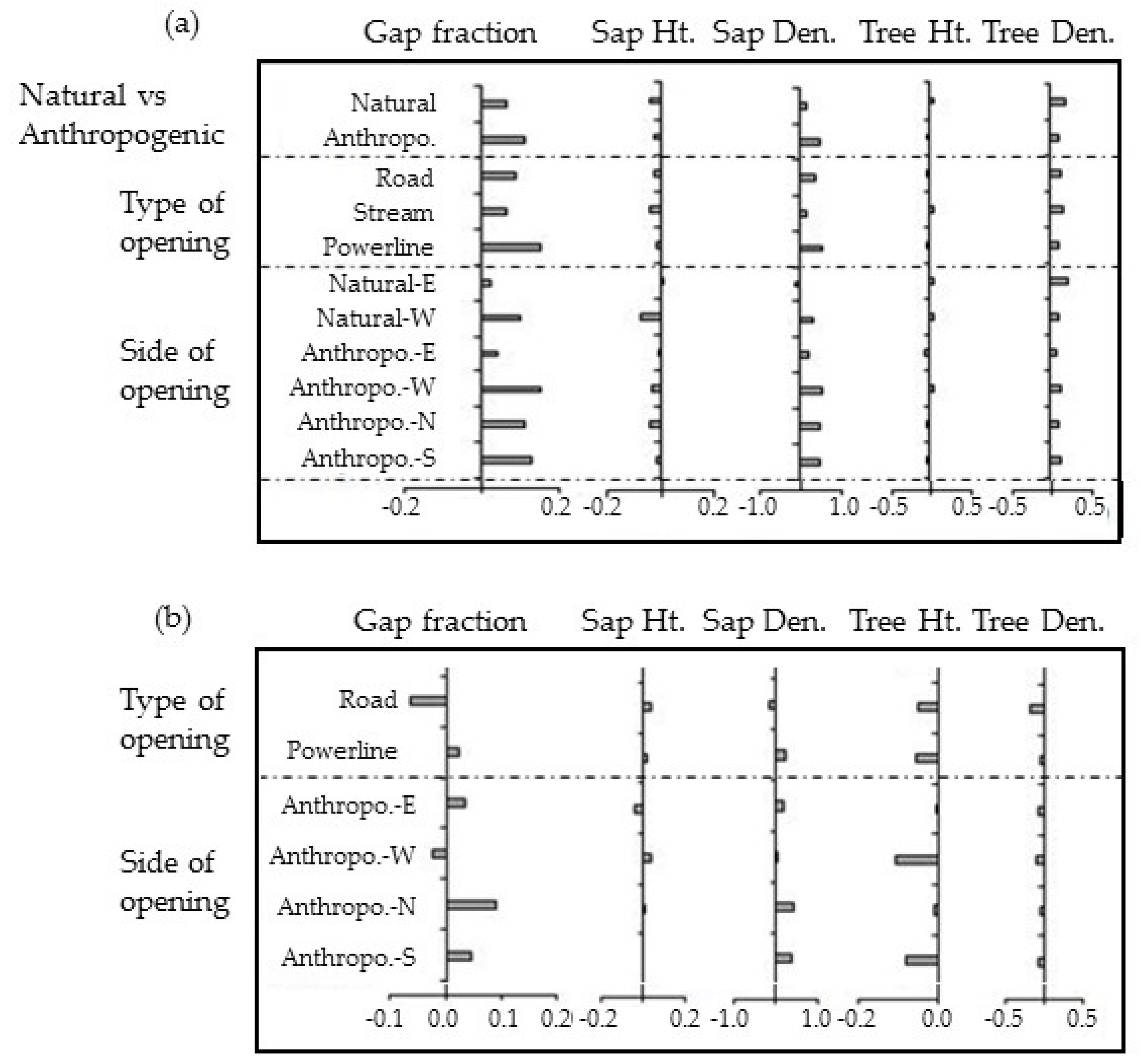
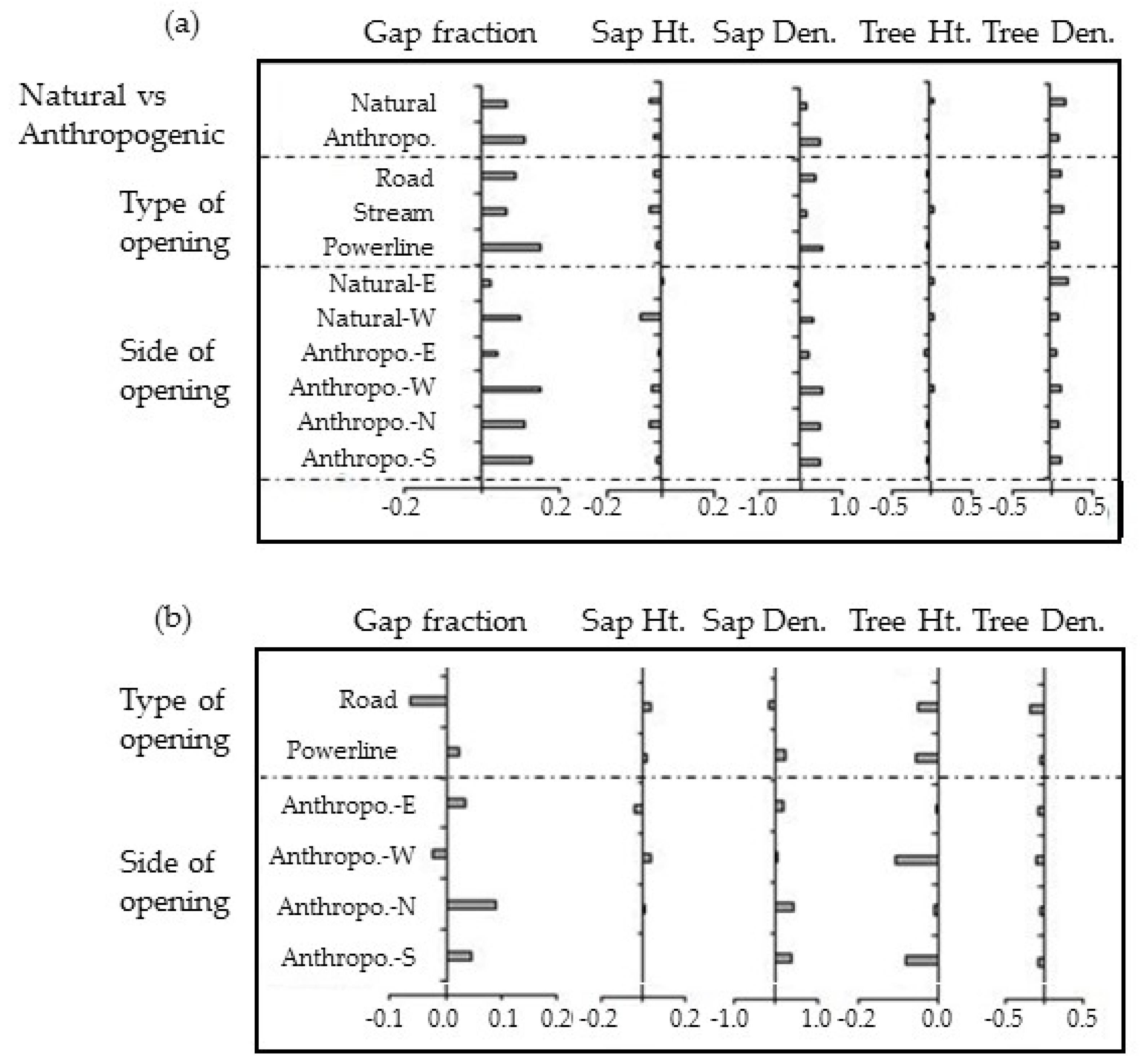
3.2. Natural vs. Anthropogenic Openings
4. Discussion
4.1. Do Natural and Anthropogenic Linear Openings Cause the Same Effects?
4.2. The Influence of Orientation
4.3. How Far Do Effects Extend?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bourgeois, L.; Kneeshaw, D.; Boisseau, G. Les routes forestières au québec: Les impacts environnementaux, sociaux et économiques. VertigO-La Rev. Électron. Sci. L’environ. 2005, 6, 2. [Google Scholar] [CrossRef]
- Riitters Kurt, H.; Wickham James, D. How far to the nearest road? Front. Ecol. Environ. 2003, 1, 125–129. [Google Scholar] [CrossRef]
- Daigle, P. A summary of the environmental impacts of roads, management responses, and research gaps: A literature review. J. Ecosyst. Manag. 2010, 10, 65–89. [Google Scholar]
- Coulombe, G.; Huot, J.; Arsenault, J.; Bauce, E.; Bernard, J.-T.; Bouchard, A.; Liboiron, M.A.; Szaraz, G. Commission D’étude sur la Gestion de la Forêt Publique Québécoise; Bibliothèque Nationale du Québec: Québec City, QC, Canada, 2004; p. 307. [Google Scholar]
- Alverson, W.S.; Waller, D.M.; Solheim, S.L. Forests too deer: Edge effects in northern Wisconsin. Conserv. Biol. 1988, 2, 348–358. [Google Scholar] [CrossRef]
- Gascon, C.; Williamson, G.B.; da Fonseca, G.A.B. Receding forest edges and vanishing reserves. Science 2000, 288, 1356–1358. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F.; Goosem, M.; Laurance, S.G.W. Impacts of roads and linear clearings on tropical forests. Trends Ecol. Evol. 2009, 24, 659–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forman, R.T.T.; Godron, M. Landscape Ecology; Jhon Wiley & Sons: New York, NY, USA, 1986; p. 619. [Google Scholar]
- Williams-Linera, G. Vegetation structure and environmental conditions of forest edges in Panama. J. Ecol. 1990, 78, 356–373. [Google Scholar] [CrossRef]
- Murcia, C. Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 1995, 10, 58–62. [Google Scholar] [CrossRef]
- Johnson, H.B.; Vasek, F.C.; Yonkers, T. Productivity, diversity and stability relationships in mojave desert roadside vegetation. Bull. Torrey Bot. Club 1975, 102, 106–115. [Google Scholar] [CrossRef]
- Lamont, B.B.; Rees, R.G.; Witkowski, E.T.F.; Whitten, V.A. Comparative size, fecundity and ecophysiology of roadside plants of banksia hookeriana. J. Appl. Ecol. 1994, 31, 137–144. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E. Structure and composition of riparian boreal forest: New methods for analyzing edge influence. Ecology 2001, 82, 649–659. [Google Scholar] [CrossRef]
- Kardynal, K.J.; Morissette, J.L.; Van Wilgenburg, S.L.; Bayne, E.M.; Hobson, K.A. Avian responses to experimental harvest in southern boreal mixedwood shoreline forests: Implications for riparian buffer management. Can. J. For. Res. 2011, 41, 2375–2388. [Google Scholar] [CrossRef]
- Arienti, M.C.; Cumming, S.G.; Krawchuk, M.A.; Boutin, S. Road network density correlated with increased lightning fire incidence in the Canadian western boreal forest. Int. J. Wildland Fire 2009, 18, 970–982. [Google Scholar] [CrossRef]
- Burton, P.J. Effects of clearcut edges on trees in the sub-boreal spruce zone of northwest-central British Columbia. Silva Fenn. 2002, 36, 329–352. [Google Scholar] [CrossRef]
- Gauthier, S.; Vaillancourt, M.-A.; Leduc, A.; De Grandpré, L.; Kneeshaw, D.; Morin, H.; Drapeau, P.; Bergeron, Y. Ecosystem Management in the Boreal Forest; Presses de l’Université du Québec: Québec, QC, Canada, 2009; p. 539. [Google Scholar]
- Angelstam, P.K. Maintaining and restoring biodiversity in european boreal forests by developing natural disturbance regimes. J. Veg. Sci. 1998, 9, 593–602. [Google Scholar] [CrossRef]
- Didham, R.K.; Lawton, J.H. Edge structure determines the magnitude of changes in microclimate and vegetation structure in tropical forest fragments. Biotropica 1999, 31, 17–30. [Google Scholar] [CrossRef]
- Kane, V.R.; Gersonde, R.F.; Lutz, J.A.; McGaughey, R.J.; Bakker, J.D.; Franklin, J.F. Patch dynamics and the development of structural and spatial heterogeneity in pacific northwest forests. Can. J. For. Res. 2011, 41, 2276–2291. [Google Scholar] [CrossRef]
- Vepakomma, U.; St-Onge, B.; Kneeshaw, D. Spatially explicit characterization of boreal forest gap dynamics using multi-temporal lidar data. Remote Sens. Environ. 2008, 112, 2326–2340. [Google Scholar] [CrossRef]
- Vierling, K.T.; Vierling, L.A.; Gould, W.A.; Martinuzzi, S.; Clawges, R.M. Lidar: Shedding new light on habitat characterization and modeling. Front. Ecol. Environ. 2008, 6, 90–98. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J.P. Paysages Régionaux du Québec Méridional. Direction de la Gestion des Stocks Forestiers et Direction des Relations Publiques, Ministère des Ressources Naturelles du Québec; Les Publications du Québec: Québec, QC, Canada, 1998. [Google Scholar]
- Thibault, M.; Hotte, D. Les Régions Écologiques du Québec Méridional: Deuxième Approximation; Ministère de L’énergie et des Ressources, Service de la Cartographie: Québec, QC, Canada, 1985. [Google Scholar]
- Rowe, J.S. Forest Regions of Canada; Service, C.F., Ed.; Fisheries and Environment Canada: Ottawa, ON, Canada, 1972; p. 172. [Google Scholar]
- Bouchard, M.; Pothier, D.; Gauthier, S. Fire return intervals and tree species succession in the north shore region of Eastern Quebec. Can. J. For. Res. 2008, 38, 1621–1633. [Google Scholar] [CrossRef]
- Blais, J.R. Les forêts de la côte nord au québec sont-elles sujettes aux déprédations par la tordeuse? For. Chron. 1983, 59, 17–20. [Google Scholar] [CrossRef]
- Bouchard, M.; Pothier, D. Spatiotemporal variability in tree and stand mortality caused by spruce budworm outbreaks in eastern Quebec. Can. J. For. Res. 2010, 40, 86–94. [Google Scholar] [CrossRef]
- Pham, A.T.; Grandpré, L.D.; Gauthier, S.; Bergeron, Y. Gap dynamics and replacement patterns in gaps of the northeastern boreal forest of Quebec. Can. J. For. Res. 2004, 34, 353–364. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Bergeron, Y. Spatial and temporal patterns of seedling and sapling recruitment within canopy gaps caused by spruce budworm. Ecoscience 1999, 6, 214–222. [Google Scholar] [CrossRef]
- Canham, C.D.; Denslow, J.S.; Platt, W.J.; Runkle, J.R.; Spies, T.A.; White, P.S. Light regimes beneath closed canopies and tree-fall gaps in temperate and tropical forests. Can. J. For. Res. 1990, 20, 620–631. [Google Scholar] [CrossRef]
- Vepakomma, U.; Schneider, R.; Berninger, F.; Mailly, D.; St-Onge, B.; Hopkinson, C. Estimating Height of the Green Canopy of Black Spruce Trees in a Heterogeneous Forest Using Lidar. In Proceedings of the 32nd Symposium on Remote Sensing on Monitoring a Changing World, Sherbrooke, QC, Canada, 13–16 June 2011. [Google Scholar]
- Vepakomma, U.; St-Onge, B.; Kneeshaw, D. Response of a boreal forest to canopy opening: Assessing vertical and lateral tree growth with multi-temporal lidar data. Ecol. Appl. 2011, 21, 99–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Baldocchi, D.; Gong, P.; Kelly, M. Isolating individual trees in a savanna woodland using small footprint lidar data. Photogramm. Eng. Remote. Sens. 2006, 72, 923–932. [Google Scholar] [CrossRef]
- Runkle, J.R. Guidelines and Sample Protocol for Sampling Forest Gaps; Gen. Tech. Rep. PNW-GTR-283; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1992; p. 283.
- Hawkins, D.M.; Olwell, D.H. Cumulative Sum Charts and Charting for Quality Improvement; Springer Science & Business Media: Berlin, Germany, 2012; p. 246. [Google Scholar]
- Forman, R.T.T.; Alexander, L.E. Roads and their major ecological effects. Annu. Rev. Ecol. Syst. 1998, 29, 207–231. [Google Scholar] [CrossRef]
- Laurance, W.F. Ecosystem decay of Amazonian forest fragments: Implications for conservation. In Stability of Tropical Rainforest Margins; Springer: Berlin, Germany, 2007; pp. 9–35. [Google Scholar]
- Harper, K.A.; Bergeron, Y.; Gauthier, S.; Drapeau, P. Post-fire development of canopy structure and composition in black spruce forests of Abitibi, Québec: A landscape scale study. Silva Fenn. 2002, 36, 249–263. [Google Scholar] [CrossRef]
- Laurance, W.F.; Ferreira, L.V.; Rankin-de Merona, J.M.; Laurance, S.G. Rain forest fragmentation and the dynamics of amazonian tree communities. Ecology 1998, 79, 2032–2040. [Google Scholar] [CrossRef]
- Mesquita, R.C.G.; Delamônica, P.; Laurance, W.F. Effect of surrounding vegetation on edge-related tree mortality in Amazonian forest fragments. Biol. Conserv. 1999, 91, 129–134. [Google Scholar] [CrossRef]
- Fleming, R.L.; Crossfield, R.M. Strip Cutting in Shallow-Soil Upland Black Spruce Near Nipigon, Ontario, III: Windfall and Mortality in the Leave Strips: Preliminary Results; Information Report O-X-354; Great Lake Forestry Centre, Canadian Forest Service, Department of the Environment: Sault Ste. Marie, ON, Canada, 1983; p. 29. [Google Scholar]
- Ruel, J.C. Understanding windthrow: Silvicultural implications. For. Chron. 1995, 71, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Laurance, W.F.; Lovejoy, T.E.; Vasconcelos, H.L.; Bruna, E.M.; Didham, R.K.; Stouffer, P.C.; Gascon, C.; Bierregaard, R.O.; Laurance, S.G.; Sampaio, E. Ecosystem decay of amazonian forest fragments: A 22-year investigation. Conserv. Biol. 2002, 16, 605–618. [Google Scholar] [CrossRef]
- Strayer, D.L.; Power, M.E.; Fagan, W.F.; Pickett, S.T.A.; Belnap, J. A classification of ecological boundaries. BioScience 2003, 53, 723–729. [Google Scholar] [CrossRef]
- Harper, K.A.; Macdonald, S.E.; Burton, P.J.; Chen, J.; Brosofske, K.D.; Saunders, S.C.; Euskirchen, E.S.; Roberts, D.A.R.; Jaiteh, M.S.; Esseen, P.A. Edge influence on forest structure and composition in fragmented landscapes. Conserv. Biol. 2005, 19, 768–782. [Google Scholar] [CrossRef]
- Matlack, G.R. Microenvironment variation within and among forest edge sites in the eastern United States. Biol. Conserv. 1993, 66, 185–194. [Google Scholar] [CrossRef]
- Trombulak, S.C.; Frissell, C.A. Review of ecological effects of roads on terrestrial and aquatic communities. Conserv. Biol. 2000, 14, 18–30. [Google Scholar] [CrossRef]
- Kneeshaw, D.D.; Larouche, M.; Asselin, H.; Adam, M.C.; Saint-Arnaud, M.; Reyes, G. Road rash: Ecological and social impacts of road networks on first nations. In Planning Co-Existence: Aboriginal Considerations and Approaches in Land Use Planning; Stevenson, M.G., Natcher, D.C., Eds.; Canadian Circumpolar Institute Press: Edmonton, AB, Canada, 2010; pp. 171–184. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type | Sampled Area | Width (m) | Orientation (°) | Length (m) | Total Area Sampled (ha) | Mean Elevation (m) | |
|---|---|---|---|---|---|---|---|
| Side 1 * | Side 2 * | ||||||
| Stream | 1 | 50 | 0 | 550 | 11 | 107 (E) | 105 (W) |
| 2 | 70 | 0 | 500 | 10 | 107 (E) | 106 (W) | |
| Road | 1 | 20 | 0 | 530 | 10.6 | 293 (E) | 288 (W) |
| 2 | 20 | 25 | 1200 | 24 | 242 (E) | 238 (W) | |
| 3 | 20 | 35 | 1000 | 20 | 380 (N) | 376 (S) | |
| Powerline | 1 | 90 | 270 | 530 | 10.6 | 330 (N) | 340 (S) |
| 2 | 150 | 325 | 1000 | 20 | 376 (N) | 380 (S) | |
| Comparison | Type | Gap Fraction | Sapling Height (m) | Sap. Density (stems/ha) | Tree Height (m) | Tree Density (stems/ha) | ||
|---|---|---|---|---|---|---|---|---|
| Mean | Max | Mean | Mean | Max | Mean | Mean | ||
| Natural vs. Anthropogenic | Natural | 0.41 ± 0.07 | 4.0 | 2.6 ± 0.5 | 46 ± 27 | 31.3 | 13.1 ± 0.4 | 746.6 ± 51.4 |
| Anthropo. | 0.45 ± 0.04 | 4.0 | 2.7 ± 0.1 | 93 ± 20 | 32.5 | 11.4 ± 0.3 | 659.9 ± 35.4 | |
| Nat. vs. Anth. | p < 0.01 | p < 0.05 | p < 0.05 | p < 0.05 | p < 0.05 | |||
| All features | Reference | 0.36 | 3.9 | 2.8 ± 0.8 | 32 | 25.6 | 11.9 ± 2.2 | 520 |
| Road | 0.43 ± 0.42 | 4.0 | 2.6 ± 0.1 | 77 ± 19 | 32.5 | 11.8 ± 0.4 | 663.6 ± 31.8 | |
| Stream | 0.41 ± 0.07 | 4.0 | 2.6 ± 0.5 | 46 ± 27 | 31.3 | 13.1 ± 0.4 | 746.6 ± 51.4 | |
| Powerline | 0.49 ± 0.43 | 4.0 | 2.7 ± 0.1 | 116 ± 33 | 31.3 | 10.8 ± 0.4 | 654.3 ± 56.0 | |
| Reference vs. linear openings | Ref. vs. all types | p < 0.01 | p < 0.01 | ns | ns | p < 0.001 | ||
| Ref. vs. type | p < 0.01 | p < 0.05 | p < 0.01 | ns | p < 0.001 | |||
| Natural vs. Anthropogenic vs. orientation | Natural-E | 0.38 ± 0.06 | 4.0 | 2.8 ± 0.3 | 25 ± 15 | 31.3 | 13.2 ± 0.6 | 830.2 ± 78.7 |
| Natural-W | 0.44 ± 0.09 | 4.0 | 2.4 ± 0.4 | 66 ± 40 | 29.8 | 13.1 ± 0.9 | 663.2 ± 75.2 | |
| Nat. E vs. W | p < 0.001 | p < 0.001 | p < 0.001 | ns | p < 0.001 | |||
| Anthropo.-E | 0.39 ± 0.05 | 4.0 | 2.7 ± 0.2 | 49 ± 27 | 26.8 | 10.6 ± 0.5 | 604.2 ± 45.5 | |
| Anthropo.-W | 0.49 ± 0.1 | 4.0 | 2.6 ± 0.2 | 118 ± 45 | 32.5 | 12.9 ± 0.7 | 679.8 ± 70.7 | |
| Nat. vs. Anth.-E | p < 0.05 | p < 0.001 | p < 0.01 | ns | p < 0.001 | |||
| Nat. vs. Anth.-W | p < 0.05 | ns | p < 0.01 | p < 0.001 | p < 0.001 | |||
| Anthropo.-N | 0.45 ± 0.03 | 4.0 | 2.6 ± 0.2 | 98 ± 28 | 31.1 | 11.2 ± 0.4 | 649.5 ± 50.3 | |
| Anthropo.-S | 0.47 ± 0.03 | 4.0 | 2.7 ± 0.2 | 100 ± 25 | 31.3 | 11.0 ± 0.4 | 694.2 ± 42.7 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vepakomma, U.; Kneeshaw, D.D.; De Grandpré, L. Influence of Natural and Anthropogenic Linear Canopy Openings on Forest Structural Patterns Investigated Using LiDAR. Forests 2018, 9, 540. https://doi.org/10.3390/f9090540
Vepakomma U, Kneeshaw DD, De Grandpré L. Influence of Natural and Anthropogenic Linear Canopy Openings on Forest Structural Patterns Investigated Using LiDAR. Forests. 2018; 9(9):540. https://doi.org/10.3390/f9090540
Chicago/Turabian StyleVepakomma, Udayalakshmi, Daniel D. Kneeshaw, and Louis De Grandpré. 2018. "Influence of Natural and Anthropogenic Linear Canopy Openings on Forest Structural Patterns Investigated Using LiDAR" Forests 9, no. 9: 540. https://doi.org/10.3390/f9090540
APA StyleVepakomma, U., Kneeshaw, D. D., & De Grandpré, L. (2018). Influence of Natural and Anthropogenic Linear Canopy Openings on Forest Structural Patterns Investigated Using LiDAR. Forests, 9(9), 540. https://doi.org/10.3390/f9090540