Abstract
Seasonal timing of prescribed fire and alterations to the structure and composition of fuels in savannas and woodlands can release understory hardwoods, potentially resulting in a global increase of closed-canopy forest and a loss of biodiversity. We hypothesized that growing-season fire, high overstory density, and wiregrass presence in longleaf pine woodlands would reduce the number and stature of understory hardwoods, and that because evergreen hardwoods retain live leaves, dormant-season fire would reduce performance and survival of evergreen more than deciduous hardwoods. Understory hardwood survival and height were monitored over seven years in longleaf pine woodlands in southwest Georgia with a range of overstory density, groundcover composition, and season of application of prescribed fire. Hardwood stem survival decreased with increasing overstory density, and deciduous hardwoods were more abundant in the absence of wiregrass. Contrary to expectations, evergreen hardwood growth increased following dormant-season fire. Differences in hardwood stem survival and height suggest that low fire intensity in areas with low overstory density increase the risk that hardwoods will grow out of the understory. These results indicate a need for focused research into the effects of groundcover composition on hardwood stem dynamics and emphasize that adequate overstory density is important in longleaf ecosystem management.
1. Introduction
Loss of savannas and grassy woodlands to woody species encroachment is occurring globally [1], and could have profound effects ecologically and economically [2,3]. In large parts of their ranges, savannas and woodlands are maintained by frequent fires that regulate the size and occurrence of woody species that would otherwise convert these biomes into closed-canopy forests [4,5]. Changes in rangeland, forest or livestock management can affect savanna and woodland vegetation configuration and structure, which can reduce fire’s efficacy in controlling woody species and could shift these ecosystems towards closed-canopy forest.
Longleaf pine (Pinus palustris Mill.) woodlands once covered 24.3 million ha of the Southeastern Coastal Plain of the United States [6], and featured a persistent and sometimes problematic understory of hardwood trees. Prior to European settlement, lightning- and human-ignited fires that occurred every 1–10 years created an open, pine-dominated canopy by minimizing recruitment of understory hardwoods into the midstory and overstory [7,8,9]. Because of landscape fragmentation, naturally ignited fires today are typically limited in extent [10], and prescribed fire is presently the most common and effective means for maintaining longleaf pine woodland structure and ecosystem function [11,12].
Typically, prescribed fire every 1–3 years in longleaf pine woodlands kills the aboveground portions of understory hardwoods (top-kills), thus preventing hardwood recruitment into the mid- and overstory [9,13]. Understory hardwoods are nonetheless common because they persist as sprouting rootstocks which often survive fire. Woody sprouting species have been characterized as being trapped in the understory by frequent fire [14]. Under this paradigm, sprouting hardwoods that fail to reach a threshold height between fires are top-killed and must rebuild height by sprouting from ground level. With repeated fires, a depletion of carbohydrates may occur resulting in death [15,16]. Alternatively, hardwood stems that exceed a threshold height may escape the trap [17]. These individuals rapidly increase in fire resistance because aboveground meristems that survive fire inhibit sprouting from the stem and root crown [18], and focus resource allocation to growth of surviving stems. As applied to longleaf pine woodland, reductions in fire duration or temperature could result in a lower threshold height and more hardwood recruitment into the midstory.
The efficacy of fire for management of understory hardwoods in longleaf pine woodlands may depend on the time of year the burn takes place (fire season), as well as the type, amount and arrangement of fuels. It is widely accepted among prescribed fire practitioners that better hardwood control is obtained with growing-season fire, denser pine canopies and groundcover dominated by wiregrass (Aristida stricta Michx.) [19].
During much of the twentieth century, most prescribed burning on the Southeastern Coastal Plain was conducted during the dormant season, partly for fear that more intense growing-season fires would damage overstory trees and reduce growth [20]. Yet it has become recognized that seasonal timing of fire may influence hardwood control [21,22]. Growing-season fire has been linked to reduced density of understory and midstory hardwoods and increased plant biodiversity [9,21]. Many shrub species sprout less profusely after growing-season fire than dormant-season fire [13]. Effects of fire season may differ by leaf habit: deciduous oaks are less affected by leaf-off (dormant-season) fires than by leaf-on (growing-season) fires [9]. By reducing understory and midstory hardwood density, and enhancing flowering in the groundcover, growing-season fires favor grasses and other herbaceous species. Evidence for ecological benefits of growing-season burning notwithstanding, operational considerations (e.g., smoke production, hunting restrictions, heat stress for burn personnel) often restrict burning to cooler seasons [23,24].
Both wiregrass and longleaf pine litter increase flame length, time of burning, and time of smoldering relative to other fuels [25,26,27,28]. Among North American pine species, longleaf pine needles have been shown to burn with the longest flame length [26], and wiregrass features strong, horizontally tending blades that hold pine needle litter above the soil surface, creating a dry, well-aerated fuel-bed [25,29,30,31]. Because of this fuel-bed structure, fires in woodlands with wiregrass-dominated groundcover and abundant longleaf pine litter burn more intensely and may increase hardwood mortality compared to open forests with old-field groundcover [32].
Silvicultural treatments, agricultural legacies, and natural disturbance can affect fuel characteristics [33,34]. Silvicultural methods that create openings in the canopy that exceed the height of a single tree, such as group selection, shelterwood, and seed-tree methods [35], may decrease canopy fuel inputs in localized areas. Agricultural legacies generally include the absence of wiregrass, which does not readily recolonize on disturbed soils [36]. Natural disturbances include tornadic windstorms and tropical cyclones [37]. Any of these conditions may lead to interruptions in fuelbed continuity, increasing the likelihood that hardwood stems will survive fire and will grow into the midstory [38], leading to undesirable changes in forest condition.
When overstory density is reduced either through management or natural disturbance, hardwood control via prescribed fire often becomes less effective. Reduction in overstory density reduces the contribution of fine fuel from the canopy (i.e., pine needles) to the fuel bed. Reduction in the amount of pine litter in the fuel bed results in lower fire intensity [39], which decreases aboveground hardwood mortality and increases the likelihood that surviving stems will grow into the midstory [38]. Thus, hardwood control with fire may be compromised in large overstory openings where fire intensity is likely to be lower.
Despite considerable knowledge about their individual effects, it is largely unknown how interactions between overstory density, groundcover composition and fire season affect understory dynamics. This study seeks to improve our understanding of the roles that fire season, overstory pine density and groundcover composition play in hardwood control using prescribed fire in frequently burned longleaf pine woodlands. To clarify how season of fire affects understory hardwoods in a variety of groundcover types and overstory densities, we asked the following questions: does growing-season fire increase top-kill relative to dormant-season fire? Are deciduous hardwoods more affected by fire seasonality than evergreen hardwoods? Is there an overstory density below which hardwood sprouts can escape top-kill by fire? And does groundcover dominated by wiregrass provide better control over understory hardwoods than groundcover consisting of old-field vegetation? The overarching goal of this study is to improve natural resource managers’ capacity to use prescribed fire to manage hardwoods in longleaf pine woodlands in the Southeastern United States and other savanna and woodland ecosystems worldwide.
2. Materials and Methods
2.1. Study Area
This study was conducted at the Joseph W. Jones Ecological Research Center at Ichauway in southwest Georgia, USA. Ichauway is an 11,000-ha research property composed primarily of longleaf pine woodland dominated by trees which became established between 1900 and 1920 following cessation of commercial forestry activities (e.g., timber and turpentine production) or abandonment of agricultural fields [40,41]. Since 1990, longleaf pine woodlands at Ichauway have been managed primarily by application of dormant-season fires at 2-year intervals. Longleaf pine woodland at Ichauway occurs on landforms ranging from xeric sandhills to periodically wet woodland on poorly drained soils, but the majority occurs on well-drained, upland soils.
Ichauway is located on the Gulf Coastal Plain where the climate is humid subtropical. The site receives an average of 1360 mm of precipitation spread evenly throughout the year [42]. Mean monthly temperatures range from 20 to 33 °C during the summer and from 3 to 18 °C during the winter. Over the seven-year study period from 2010 to 2016, Ichauway received approximately 89% of long-term mean annual precipitation but precipitation varied considerably from year to year. Mean annual precipitation averaged 70% of the long-term normal over the first three years of the study, and 103% of normal over the final four years. The study took place in stands devoted to an experiment on selection harvest methods. In 2009, longleaf pine basal area in fourteen 4-ha treatment stands was reduced through single-tree and group-selection harvests. In addition, there were 4 uncut control stands. Single-tree harvest treatments were based on the Stoddard-Neel approach to ecological forestry [43] and reduced basal areas by approximately 15%. Group selection harvest treatments were targeted to have the same residual basal area as the single-tree selection harvest in the same treatment block. Residual basal areas ranged from 10.8 to 18.7 m2/ha. In 2010, eighty one 4 × 4 m plots were established across the treatments to monitor hardwood demography.
Hardwood plot locations were chosen based on overstory density and groundcover type. An index of overstory density was characterized over the treatment areas. Trees ≥ 10 cm dbh (diameter at breast height) were mapped with GPS (A101 Smart Antenna, Hemisphere GNSS, Scottsdale, AZ; Nomad 1050 handheld computer, Trimble, Sunnyvale, CA, USA); a few trees were mapped using a compass and tape. Overstory abundance index (OAI) was calculated at the center of each grid cell on a 4 m grid using the formula
where A is the cross-sectional area at breast height of each tree i ≥ 10 cm dbh within 15 m of the center of each grid cell, and d is the distance to each tree [44,45]. This overstory abundance index, which has units of cm2 m−1, attributes greater influence to larger trees and/or trees that are nearer to the cell center. The grid cells were grouped by overstory density (as indicated by OAI) into three classes with similar numbers of observations. The ranges for the classes are low (0–471), medium (472–1414) and high (1415–3359). Two cells per overstory abundance index class were randomly selected as sample plots in nine stands, and one cell per overstory abundance index class was randomly selected in the other nine stands. Groundcover in nine of the treatment stands was dominated by wiregrass, and groundcover in the other nine stands was absent wiregrass and dominated by ruderal, old-field species typical of disturbed soils and abandoned agricultural plots (e.g., Andropogon virginicus L., Pityopsis graminifolia (Michx.) Nutt., Dyschoriste oblongifolia (Michx.) Kuntze, Scleria ciliata (Michx), and Aclypha gracilens A. Gray).
All stands were burned biennially in February and early March (dormant season) or in late April and May (growing season). Dormant-season burns were conducted in 2014, and growing-season burns were conducted in 2010, 2012 and 2016. The goals of all prescribed burns were to consume accumulated surface fuels and kill/reduce aboveground woody understory species while minimizing scorch to overstory longleaf pine trees. All burns were conducted with relative humidity between 20% and 60%, and wind speeds <9 m/s (Table 1). Because wiregrass can cause high crown scorch when burned in warmer, drier conditions and can burn well when relative humidity is high, wiregrass stands were preferentially burned under cooler, more humid conditions than old-field stands (Scott Smith, personal communication). The downwind side of a burn unit containing sample plots was ignited, and fire allowed to back into the unit to create a buffer at the downwind perimeter. Ignition patterns upwind and in the interior of burn units varied based on weather and fire behavior at the times of ignition.

Table 1.
Average fire-weather during prescribed fires.
Fire intensity was not measured during prescribed fires. Increased overstory density and wiregrass groundcover were assumed to increase fire intensity: longleaf pine needles burns with the longest flame length of any North American pine species and are positively related to fire intensity [26,39]; wiregrass biomass is positively linked with increased flame length, and longleaf pine needles perched in wiregrass tussocks increase duration of burning [25]. Growing-season fire was hypothesized to decrease genet and stem survival because a larger proportion of a plant’s resources are aboveground and vulnerable to fire at that time, and because the ambient temperature is high in the growing season, the temperature of tissue death (60 °C) can be reached more quickly [9].
Plots were sampled once per year in late summer or early fall for six years from 2011 to 2016. Starting from the northwest corner of the 4 × 4 m plot, a 2 m2 area was examined, and a labeled stake was placed near each sprout or group of sprouts with a common rootstock. Sprouts were determined to have a common rootstock by carefully following each sprout to its base. In most cases, sprouts of the same species in a group were found to branch at or just below the soil surface from a common root-crown. Because groups of understory oaks are often from the same genet [46], these groups were assumed to be genetically related and were classified as belonging to a single genet. Hardwood trees studied were laurel oak, live oak and water oak, classified here as evergreen, and bluejack oak, post oak, southern red oak, common persimmon, sassafras and winged sumac, classified as deciduous (Table 2). If fewer than eight genets of a hardwood species were found in the first 2 m2 of the plot, the search area was increased by 2 m2. After that, the area sampled was increased by 4 m2 increments until at least eight genets of each species were found or 16 m2 had been sampled. In the first year of the study, the number of sprouts per genet were counted and the height of the tallest live sprout in each genet was estimated visually as belonging to one of 4 height classes: <0.5 m, 0.5 to <1 m, 1 to <1.5 m, or ≥1.5 m (we refer to “genet height” because only one sprout per genet was measured). Each sprout was painted near the base. In subsequent years, we counted the number of sprouts per genet that died, survived, or were new, and continued to estimate genet height. New and surviving sprouts were marked with a different color of paint than the previous year, and dead sprouts were clipped. If no new or surviving sprouts were observed for two consecutive years, genet death was recorded as having occurred on the first of those two years.
Table 2.
Leafing behavior and taxonomic affinity of common hardwood tree species that persist in shrub form in frequently burned longleaf pine woodland in southwest Georgia, USA.
2.2. Statistical Analyses
The hypotheses that fire season, overstory density and the composition of ground cover affected hardwood performance were tested using generalized linear models, generalized linear mixed models, and ordinal regression in R version 3.3.2 [48,49,50]. Our general strategy was to propose models incorporating all possible effects, e.g., overstory density, ground cover composition, and fire season, and then to drop effects by backward elimination applying a multi-step protocol [51]. For every successive model iteration, two-tailed z-test scores and p-values were calculated for each level of fixed effects. The least significant fixed effect was successively eliminated from the model if it was not part of an interaction, was not a level of a categorical fixed effect that had one or more other significant levels, was not significant at a 95% confidence level, and its removal resulted in a lower value of Akaike’s information criterion (AIC). To ensure that this protocol did not result in type-II error by retaining marginally significant effects in the model, analysis of variance (ANOVA) tests of fixed effects were run using type II or III sums of squares depending on whether the model contained only simple or interactive effects, respectively. Estimates of the mean number of genets at each level of significant fixed effects were generated using least-squares means.
2.3. Genet Survival
We used linear models to determine how overstory density and groundcover composition affected abundance and composition of understory hardwoods. Genet counts at each plot were transformed to density (genets m−2) based on the area sampled for each species. These data were log-transformed to improve normality. Initial models included leaf habit, overstory density and groundcover composition and all two-way interactions as fixed effects. Final models were chosen using backwards elimination as described above. Least-squares means estimates of genet density at each level of significant fixed effects were back-transformed to the original scale.
To determine how leaf habit, overstory, groundcover, and fire season influenced genet survival, we used generalized linear mixed models based on a binomial distribution. Initial models for genet survival included groundcover, leaf habit, overstory density, fire season (dormant or growing) and all two-way interactions as fixed effects, and a random effect to account for variation between genets. Final models were chosen using backward elimination.
2.4. Sprouting
We sought to determine if the presence of an active aboveground meristem that had survived fire would suppress sprouting. Linear mixed models were used to compare the counts of new sprouts following fires in fire-years. Stem survival was classified by genet as either no or at least one stem surviving prescribed burning. The initial model included pre-fire genet height, stem survival and their interaction as fixed effects, and species and genet as random effects.
2.5. Stem Survival
We also asked whether deciduous and evergreen understory hardwood stem survival differed by fire season in varying overstory and groundcover conditions. Generalized linear mixed models were used to compare the probability that one or more stems would survive fire. For this analysis, stem survival was again classified by genet as either no or at least one stem that survived prescribed burning, but was used as the response variable, and modelled on a binomial distribution. The initial model included the fixed effects of leaf habit, pre-fire genet height class, overstory density class, groundcover composition, fire season and all two-way interactions; variation between genets was accounted for in a random effect.
To determine whether there is a size threshold at which hardwood stems begin to escape topkill by fire, we analyzed height class with respect to the main experimental factors. Genet height was analyzed with proportional-odds models, because it was recorded in four ordered classes. Proportional-odds models fit logistic regressions to the proportions of data below and above a series of thresholds that correspond to the ordinal levels of the data [51]. Probabilities of stem height >1 m (>height class 2) were generated using least-squares means to compare fixed effects. Initial models included the effects of leaf habit, overstory density, groundcover type and fire season and all possible interactions with genet included as a random effect.
3. Results
3.1. Genet Survival
Groundcover composition affected the number and composition of genets found per plot (Figure 1), but overstory density did not. In old-field groundcover, density of deciduous genets (0.44 m−2) was 132% higher than evergreen genets (0.19 m−2; p = 0.001). The density of deciduous genets in old-field groundcover was also 120% higher than deciduous genets in wiregrass (0.20 m−2; p < 0.001) and 214% higher than evergreen genets in wiregrass (0.14 m−2; p < 0.001).
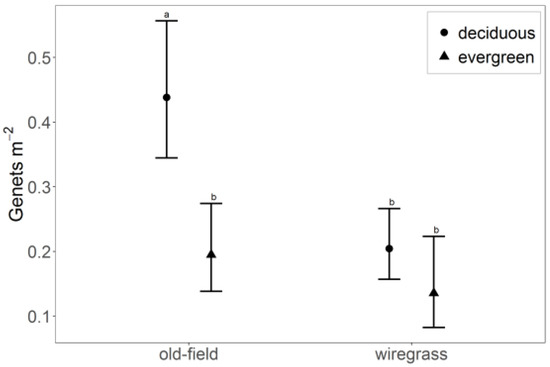
Figure 1.
Hardwood genet density (least-squares means with 95% confidence intervals) in longleaf pine woodland according to groundcover type and leaf habit. Old-field groundcover is indicative of areas that have a history of soil disturbance (e.g., agriculture), whereas wiregrass is indicative of relatively undisturbed soils. Effects that do not share superscripted letters are significantly different at a 95% confidence level.
Fire season interacted with groundcover composition, leaf habit and overstory density to significantly affect genet survival (Figure 2). The primary effect due to groundcover type was that hardwood genets in old-field groundcover had a lower probability of survival following growing-season fire (92.9%) than following dormant-season fire (96.7%; p = 0.001; Figure 2A). Given the documented effectiveness of wiregrass in hardwood control, it was surprising to find that genet survival in old-field groundcover following growing-season fire was significantly lower than genet survival in wiregrass following either growing-season fire (95.8%; p ≤ 0.043) or dormant-season fire (96.2%; p ≤ 0.034). This may be due a practice common at Ichauway of burning wiregrass in lower ambient temperature and higher fuel moisture relative to old-field groundcover (Scott Smith, personal communication). The main effect related to leaf habit was that evergreen hardwood genet survival was significantly higher following dormant-season fire (97.9%) than deciduous hardwood genet survival following both dormant-season fires (94.1%; p = 0.006; Figure 2B) and growing-season fires (94.2%; p = 0.009). The effects of overstory density on genet survival were complex (Figure 2C). Genet survival was higher following dormant-season fire (97.6%) than following growing-season fire (93.0%) in medium overstory density (p = 0.003), but fire season did not have a significant effect on genet survival in low or high overstory density. Genet survival was also higher following dormant-season fire in medium overstory density than following dormant-season fire in low overstory density (94.8%; p = 0.017) and following growing-season fire in high overstory density (94.2%; p = 0.039).
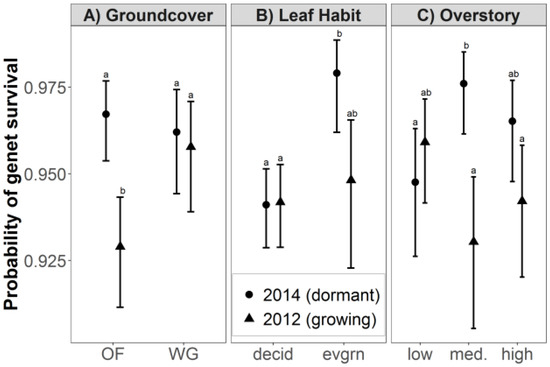
Figure 2.
Probabilities (least-square means with 95% confidence intervals) that understory hardwood genets will survive biennial prescribed fire in longleaf pine woodland in SW Georgia, USA (A) Interactive effects of fire-season and groundcover composition; (B) Fire-season and leaf habit; (C) Fire-season and overstory density. Within each panel, effects that do not share superscripted letters are significantly different at a 95% confidence level. Abbreviations: old-field, OF; wiregrass, WG; deciduous, decid.; evergreen, evgrn; medium, med.
3.2. New Sprout Counts
Counts of new sprouts following fire in burn years were significantly affected by an interaction between stem survival and pre-fire stem height. The presence of stems that survived fire resulted in significantly fewer new post-fire sprouts in all height classes (p < 0.001). There were no significant differences in new sprout counts among pre-fire height classes when stems survived previous fire (mean = 0.7 per genet). When no stems survived previous fires, the number of new sprouts per genet increased 28% from height class one to two (3.2 and 4.1 sprouts per genet, respectively), and 27% from height class two to three (5.2 sprouts per genet; p < 0.001). New sprout counts in height classes three and four (5.3 sprouts per genet) did not differ statistically.
3.3. Stem Survival
The probability that one or more stems would survive prescribed fire varied by the simple effect of fire year, and by interactive effects between overstory density and groundcover composition, and pre-fire stem height and leaf habit (Figure 3). Fire season did not have a significant effect on stem survival, but there was significant variation among dormant-season fires (Figure 3A). Probability of stem survival following the 2012 growing-season prescribed fires (0.082) was 116% higher than in the 2014 dormant-season prescribed fires (0.038; p < 0.001), which was more than 400% higher than survival following the 2016 dormant-season prescribed fires (0.007; p < 0.001).
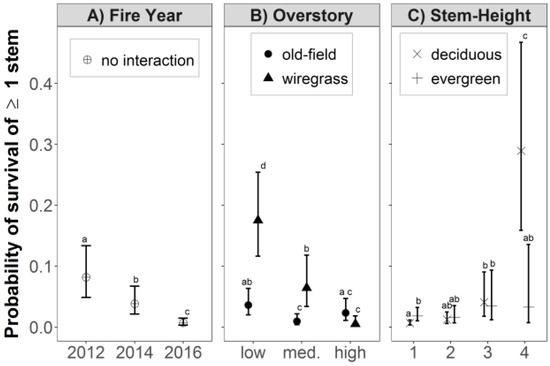
Figure 3.
Probability (least-squares means with 95% confidence intervals) that ≥1 stem per shrub-form hardwood genet will survive biennial prescribed fire in longleaf pine woodland in SW Georgia, USA. (A) Effect of fire year; (B) Interactive effect of overstory density and groundcover composition, and (C) Interactive effect of pre-fire stem height class 1 (<0.5 m), 2 (0.5 to <1 m), 3 (1 to <1.5 m), and 4 (≥1.5 m); and leaf habit. Within each panel, effects that do not share superscripted letters are significantly different at a 95% confidence level. Abbreviations: old-field, OF; wiregrass, WG; deciduous, decid.; evergreen, evgrn; medium, med.
The effects of overstory density on hardwood stem survival varied by groundcover composition (Figure 3B). Probability of stem survival in wiregrass groundcover increased 1180% from high to medium overstory density (0.005 and 0.064, respectively), and increased 173% from medium to low overstory density (0.175; p ≤ 0.003). In old-field groundcover, stem survival increased 800% from medium to low overstory density (0.004 and 0.036, respectively; p = 0.003). Comparisons in stem survival between groundcover classes shows that stem survival increased 386% from old-field to wiregrass groundcover in low overstory density (0.036 and 0.175, respectively; p < 0.001), and 611% from old-field to wiregrass groundcover in medium overstory density (0.064 and 0.009, respectively; p < 0.001). There was no difference in stem survival in old-field and wiregrass dominated groundcover where overstory density was high.
Probability of stem survival was >0.05 for both deciduous and evergreen species in height classes 1–3 (0–1.5 m), and varied significantly by leaf habit but only in pre-fire height class four (≥1.5 m; Figure 3C). Stem survival in height class four was 776% higher for deciduous species (0.289) than for evergreen species (0.033%, p = 0.039). There were no other significant differences in stem survival between deciduous and evergreen species in any other pre-fire height class.
3.4. Genet Height
Genet height was significantly affected by fire year, groundcover composition, leaf habit and overstory density (Figure 4). Fire year had no effect on genet height of deciduous species, but the probability of height >1 m (height class 2) in evergreen species was 167% higher following the 2014 dormant-season fires (0.32) than following the 2010 growing-season fire (0.12) and 113% higher than following the 2012 growing-season fire (0.15; p < 0.001; Figure 4A). In addition, the probability of height >1 m in evergreen species was 300% higher than in deciduous genets following the 2014 growing-season fires (0.08, p < 0.001).
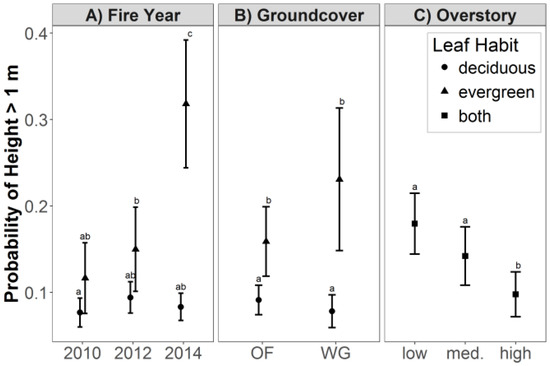
Figure 4.
Probability of stem height >1 m (least-squares means with 95% confidence intervals) of shrub-form deciduous and evergreen hardwood trees in longleaf pine woodland in SW Georgia. (A) Effects of leaf habit and fire year; (B) Effects of leaf habit and groundcover; and (C) Effect of overstory density. Within each panel, effects that do not share superscripted letters are significantly different at a 95% confidence level. Abbreviations: old-field; OF; wiregrass; WG; medium, med.
An interaction between groundcover composition and leaf habit also significantly affected genet height (Figure 4B). Probability of height >1 m was 78% higher in evergreen species than in deciduous species in old-field groundcover (0.16 and 0.09, respectively; p = 0.008) and 188% higher in wiregrass groundcover (0.23 and 0.08, respectively; p = 0.002). Probabilities of height >1 m for deciduous genets were higher in old-field than in wiregrass and lower for evergreen genets in old-field than in wiregrass, but these differences were not statistically significant.
Genet height was affected by the simple effect of overstory density (Figure 4C). Probability of height >1 m was 80% higher in low overstory density (0.18) and 40% higher in medium overstory density (0.14) than in high overstory density (0.10; p ≤ 0.011).
4. Discussion
Canopy closure and the associated reduction in understory light due to site capture by hardwoods are serious threats to groundcover species, the locus of longleaf pine woodland biodiversity [52]. In canopy openings where pine fuels are limited and/or where herbaceous fuels are sparse, hardwoods often grow large enough to survive low-intensity fire [53]. High overstory density, ground cover dominated by wiregrass and growing-season fires give longleaf pine managers flexibility in choosing ignition patterns and weather conditions that will burn intensely enough to control both deciduous and evergreen understory hardwoods; however, variability inherent in longleaf pine ecosystems can lead to unintended results [34,54]. Growing-season burning is advocated from a biological diversity standpoint, and for optimal hardwood control [13,38]. Nevertheless, the barriers to application are higher in the growing season due to smoke impacts on air quality, higher potential of damaging valuable trees, and risk of heat stress for fire personnel. Improved understanding of seasonal fire effects may assist managers in determining whether benefits of growing-season burns outweigh the disadvantages. While the results of this study highlight interactions between frequent fire and hardwood sprouts, they also illustrate high variability in these ecosystems and the need to consider additional factors to gain a more complete understanding of these complicated relationships.
Over multiple fires in frequent fire ecosystems, sprouting hardwood trees may either lose carbohydrates and eventually die [16], reach an equilibrium state in which they survive in the understory by replenishing carbohydrates lost to previous fire [55], or gain carbohydrates and eventually grow to a fire-resistant size and out of the understory. Prescribed burning may also cause understory hardwood genet mortality more directly through heating of belowground buds or meristems to lethal temperatures during fires [16]. We found overall low stem survival which agrees with high rates of topkill in previous research in a frequently burned hardwood forest [56], but we also expected that genet and stem survival would be lower under conditions predisposing to high fire intensity such as wiregrass ground cover, high overstory density, and growing-season fire. Nevertheless, multiple contingencies precluded simple generalizations. Decreased genet survival following growing-season fire in old-field groundcover, in medium overstory density and for evergreen species all suggest that fire intensity was higher during growing-season fires than during dormant-season fires. The lack of responses at the other levels of groundcover, leaf habit and overstory density are difficult to interpret. Differences in fire weather at the times of prescribed fires may have dampened the effect of groundcover on understory hardwoods. Land managers at Ichauway typically burn wiregrass-dominated groundcover at higher fuel moisture and relative humidity than old-field groundcover. Areas dominated by wiregrass burned at lower fuel moisture and relative humidity often result in undesirable levels of overstory crown scorch (Scott Smith, personal communication). The lack of leaf habit and overstory density responses may, more than anything, reveal a shortcoming of the data. Because rootstocks of genets in frequently burned ecosystems can survive for decades despite repeatedly losing aboveground stems to fire [14,57], seven years of data are likely not sufficient to fully investigate hardwood genet survival. Genet mortality in this study may reflect the effects of decades of repeated prescribed fires. The patterns of lower genet survival in fire years versus non-fire years, and decreasing stem survival following each successive fire suggest that genets may have lost carbohydrates over the study period. Carbohydrate depletion likely resulted in fewer stems reaching a fire-resistant size prior to the next prescribed fire. Fewer genets in wiregrass groundcover indicate that groundcover composition affects understory hardwood abundance over multiple fire cycles. In this study, differences in groundcover composition originated primarily from agricultural soil disturbance ending approximately 100 years prior [41]. Because wiregrass burns intensely under a wide range of fuel moisture and fire weather conditions [25], it seems likely that fires in wiregrass are more damaging to understory hardwoods, and hasten genet mortality related to repeated fires.
We also expected that growing-season fires would result in lower stem survival, and because evergreen hardwoods have resources in leaves during the dormant season, that dormant-season fires would result in higher evergreen hardwood mortality. Contrary to previous research, we found no difference in stem survival by fire season in deciduous or evergreen hardwoods. However, our study differs from Glitzenstein et al. [9], because we focused on smaller hardwoods largely confined to the understory rather than sapling-sized, more fire-resistant midstory trees. It appears that increased fire intensity may not have a strong effect on smaller stems because even low-intensity fires cause high rates of stem mortality. Because many hardwood species become less likely to sprout as they grow larger [58], sapling-sized hardwood genet survival may be more strongly affected by changes in fire intensity related to burn season. In addition, the assumption of greater carbohydrate and nutrient loss in evergreen hardwoods relative to deciduous hardwoods following dormant-season fires may have been incorrect, especially considering the February-to-early-March timing of our dormant-season fires. Southeastern evergreen hardwoods shed the previous season’s leaves and concurrently begin growing new leaves in late February and early March [59], so by the time of our dormant-season prescribed fires, evergreen leaves were already approaching senescence. Thus, the effect of losing leaves to dormant-season fire in evergreen species was reduced to the loss of several weeks of photosynthesis during a time of year when production was relatively low.
When hardwood sprouts survive fire, genet heights should increase, sprouting should decrease and, over time, there should be an overall decline in the number of sprouts present. Apical meristems inhibit the development of new shoots from the root crown [18,60]. Genets with stems that survive fire with intact apical meristems should therefore experience decreased post-fire sprouting and increased average genet height by suppressing new sprouts and continuing to grow from the apical meristem. Indeed, new sprouts decreased in genets that had one or more stems survive prescribed fire. Overall stem survival following fires was approximately 1%, so average genet height over time was dominated by new stems in lower height classes.
Average genet height did not increase over time in deciduous hardwoods and the increase in genet height in evergreen hardwoods in 2014 (Figure 4A) may have been related to the dormant-season timing of fires that year. This could have provided additional time for growth compared to deciduous hardwoods which may initiate height growth later in the spring. Furthermore, evergreen hardwoods are more physiologically active than deciduous hardwoods during the dormant season, and may be better able to take advantage of increased post-fire soil nutrient availability [61].
The striking relationship of stem survival to height class, in which deciduous hardwoods’ stem survival increased dramatically once they were in the ≥1.5 m height class, attests to profound differences in life history strategies between deciduous and evergreen oaks. The southern deciduous oaks—turkey oak, sand post oak, southern red oak, bluejack oak—accrue thick bark rapidly as juveniles [62]. This contrasts with the relatively thinner bark of southern evergreen hardwoods, which tend to occur in habitats in which fire is less frequent [63]. Thin bark may allow increased allocation of internal resources to height growth [64]. The ubiquity of evergreen oaks in upland habitats, despite their affinity for more mesic habitats [47] indicates the wide ecological amplitude of these species. The greater density of deciduous oaks, at least in old-field groundcover, suggests that the strategies associated with deciduous oak may prevail in the long term.
Longleaf canopy density affects hardwood understories by contributing long-burning needle litter to the fuelbed, casting shade, and competing for water and nutrients [26,65,66]. Based on these principles and on previous research, we expected understory hardwoods to perform poorly (decreased height growth, stem and genet survival) at higher overstory density. Consistent with our expectations, hardwood stem survival and height growth both decreased as overstory density increased. Nevertheless, it was surprising to find little effect of overstory density on genet mortality other than decreased genet survival at medium overstory density during growing-season fires. A dense canopy may provide a facilitative effect in the hot summers of the Southeastern Coastal Plain, which may forestall genet mortality [67], but this must remain conjectural. Certainly, the strong effects of canopy density on hardwood height growth and stem mortality imply that woodland managers should guard against excessive local reduction in basal area when applying selection treatments, lest understory hardwoods initiate rapid growth.
Fire-understory interactions can determine successional trajectories, and careful groundcover management is key to perpetuation of open-canopy woodlands [68]. Because wiregrass burns intensely [25,27] and appears to reduce the density of understory hardwoods [36,69], wiregrass was expected to adversely affect hardwood performance (i.e., decrease height growth, and survival of sprouts and genets), and to be associated with lower hardwood density. Consistent with this expectation, we found fewer hardwood stems on average in stands with wiregrass groundcover, but all other indicators of hardwood performance in wiregrass groundcover were surprising: height growth was unrelated to understory composition, stem survival was higher when wiregrass was present (low- and medium- density stands), and genet survival after growing-season fire was higher in wiregrass compared to old-field understories. These counterintuitive findings may be related to land use legacies, and may also be indicative of the mechanisms of wiregrass impacts on fire behavior.
Old agricultural plots are not distributed randomly on the landscape, and old fields, even long after succession to forest, may maintain cryptic fertility gradients that affect hardwood growth. For example, soils under abandoned agricultural plots maintained highly elevated phosphorus availability 14 years after longleaf pine plantation establishment compared to never-tilled soils under longleaf pine woodland [70]. Hardwood performance may be influenced by such land-use legacies, leading to improved performance in old fields which lack wiregrass. Additionally, in the absence of wiregrass, more ruderal species populate the groundcover. Broomsedge (Andropogon virginicus), an old-field colonizer which was common in our study plots, can inhibit nitrogen fixation in the soil [71] and is a strong competitor for soil resources [72]. The effect of wiregrass on fire behavior is to increase flame length and maximum temperature rather than to increase duration of heating [25]. Such increased temperatures may have little effect on shrub mortality, and duration of heating is likely to be more important [13,73].
The absence of a finding of a positive relationship between wiregrass presence and hardwood control, however, should not be construed as an argument against groundcover restoration with wiregrass. Wiregrass provides managers with more flexibility to meet burn objectives. Wiregrass burns well in a wide range of fire weather and fuel moisture conditions [25], and reduces needle and leaf litter fuel moisture by keeping it elevated above the soil surface [30]. In addition, some of the highest rates of species richness and endemism in the temperate world are found in fire-maintained longleaf pine/wiregrass ecosystems [74].
Management Implications
The results of this study highlight the importance of prescribed fire in longleaf pine woodland management, but also show that high variability in fuel structure, fuel composition and burn conditions can introduce considerable uncertainty. Taller hardwood stems in areas with low overstory density coupled with increased stem survival for deciduous stems ≥1.5 m suggest that hardwood control via prescribed fire is more challenging in areas with low pine needle litter density. Gap-based regeneration strategies may be attractive in longleaf pine woodlands because of a possible benefit to longleaf pine seedling growth [75,76]; however, understory hardwoods and other understory species [66] threaten to outcompete longleaf pine seedlings in gaps and may create conditions that jeopardize successful longleaf pine regeneration [53]. Therefore, special attention should be paid to gap size in any gap-based management strategy.
While understory hardwoods in this study performed better in old-field vegetation, it may still be important to consider groundcover restoration to wiregrass in management. Wiregrass may allow managers to meet burn objectives when fire conditions are not ideal [25,73]. Dense wiregrass groundcover appears to limit the presence of understory hardwoods [36]. Although understory hardwood performance is lower in old-field vegetation, longleaf pine seedling growth and survival may also be lower in old-field groundcover, and restoration of groundcover to wiregrass may ultimately aid in successful longleaf pine regeneration.
5. Conclusions
Oaks are an important component of longleaf pine woodlands, but managers prefer deciduous oaks over evergreen because they are less detrimental to fire regime and thus tend not to foster further colonization by trees and shrubs. Our results show that deciduous oaks >1.5 m have enhanced stem survival in comparison to evergreens, and suggest that some local reduction in overstory canopy density may be necessary, even in relatively open stands, to achieve good height growth and recruitment into the midstory. Evergreen hardwoods can grow quickly following dormant-season fire but recruitment to the midstory is unlikely because stem survival does not increase with stem height. Given the complex historic and environmental composition of the ecosystem, it is perhaps not surprising that some of our results were more complicated. Groundcover had significant effects on hardwood sprouts, but many of those effects contradicted hypotheses and some results seemed to conflict with others. Managers intend to limit adverse effects of burning to overstory trees by burning wiregrass in cooler, more humid conditions than old-field groundcover, but this practice likely dampened the hypothesized effect of greater hardwood control in wiregrass. In addition, because understory hardwood rootstocks can be persistent in frequently burned woodlands, our genet survival results may reflect the effects of land use legacy and decades of prescribed fire. Although management of understory hardwoods with fire is a complicated and multi-faceted problem, fire researchers and practitioners alike can benefit from a deeper understanding of how fire season and the overstory and understory woodland environment affect fire.
Author Contributions
Conceptualization, S.B.J.; Formal analysis, A.W.; Investigation, M.F.N.; Methodology, A.W., S.W.B. and S.B.J.; Supervision, S.W.B. and S.B.J.; Visualization, A.W.; Writing—original draft, A.W. and S.W.B.
Funding
This research was funded by the Robert W. Woodruff Foundation.
Acknowledgments
Bob Mitchell helped design the study and Noah A. Jansen developed the hardwood sampling scheme.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Mitchard, E.T.A.; Flintrop, C.M. Woody encroachment and forest degradation in sub-Saharan Africa’s woodlands and savannas 1982–2006. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368. [Google Scholar] [CrossRef] [PubMed]
- Scholes, R.J.; Archer, S.R. Tree-Grass Interactions in Savannas 1. Annu. Rev. Ecol. Syst. 1997, 28, 517–544. [Google Scholar] [CrossRef]
- Liu, Y.Y.; Van Dijk, A.I.J.M.; De Jeu, R.A.M.; Canadell, J.G.; McCabe, M.F.; Evans, J.P.; Wang, G. Recent reversal in loss of global terrestrial biomass. Nat. Clim. Chang. 2015, 5, 470–474. [Google Scholar] [CrossRef]
- Bond, W.J.; Woodward, F.I.; Midgley, G.F. The global distribution of ecosystems in a world without fire. New Phytol. 2005, 165, 525–538. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, M.; Hanan, N.P.; Scholes, R.J.; Ratnam, J.; Augustine, D.J.; Cade, B.S.; Gignoux, J.; Higgins, S.I.; Le Roux, X.; Ludwig, F.; et al. Determinants of woody cover in African savannas. Nature 2005, 438, 846–849. [Google Scholar] [CrossRef] [PubMed]
- Outcalt, K.W. Lightning, fire and longleaf pine: Using natural disturbance to guide management. For. Ecol. Manag. 2008, 255, 3351–3359. [Google Scholar] [CrossRef]
- Heyward, F. The relation of fire to stand composition of longleaf pine forests. Ecology 1939, 20, 287–304. [Google Scholar] [CrossRef]
- Christensen, N.L. Fire regimes in southeastern ecosystems. In Proceedings of the Conference Fire Regimes and Ecosystem Properties, Honolulu, HI, USA, 11–15 December 1978; Mooney, H.A., Bonnicksen, T.M., Christensen, N.L., Lotspeich, F.B., Reiners, W.A., Eds.; USDA Forest Service: Washington, DC, USA, 1981; pp. 112–136. [Google Scholar]
- Glitzenstein, J.S.; Platt, W.J.; Streng, D.R. Effects of fire regime and habitat on tree dynamics in north florida longleaf pine savannas. Ecol. Monogr. 1995, 65, 441–476. [Google Scholar] [CrossRef]
- Duncan, B.W.; Schmalzer, P.A. Anthropogenic influences on potential fire spread in a pyrogenic ecosystem of Florida, USA. Landsc. Ecol. 2004, 19, 153–165. [Google Scholar] [CrossRef]
- Kirkman, L.; Goebel, P.; Palik, B.; West, L. Predicting plant species diversity in a longleaf pine landscape. Ecoscience 2004, 11, 80–93. [Google Scholar] [CrossRef]
- Van Lear, D.H.; Carroll, W.D.; Kapeluck, P.R.; Johnson, R. History and restoration of the longleaf pine-grassland ecosystem: Implications for species at risk. For. Ecol. Manag. 2005, 211, 150–165. [Google Scholar] [CrossRef]
- Drewa, P.B.; Platt, W.J.; Moser, A.E.B. Fire effects on resprouting of shrubs in headwaters of southeastern longleaf pine savannas. Ecology 2002, 83, 755–767. [Google Scholar] [CrossRef]
- Bond, W.J.; Van Wilgen, B.W. Fire and Plants, 1st ed.; Chapman & Hall: London, UK, 1996; ISBN 0412475405. [Google Scholar]
- Waldrop, T.A.; White, D.L.; Jones, S.M. Fire regimes for pine-grassland communities in the southeastern United States. For. Ecol. Manag. 1992, 47, 195–210. [Google Scholar] [CrossRef]
- Bond, W.J.; Midgley, J.J. Ecology of sprouting in woody plants: The persistence niche. Trends Ecol. Evol. 2001, 16, 45–51. [Google Scholar] [CrossRef]
- Bond, W.J. What Limits Trees in C4 Grasslands and Savannas? Annu. Rev. Ecol. Evol. Syst. 2008, 39, 641–659. [Google Scholar] [CrossRef]
- Chao, W.S.; Foley, M.E.; Horvath, D.P.; Anderson, J. V Signals regulating dormancy in vegetative buds. Int. J. Plant Dev. Biol. 2007, 1, 49–56. [Google Scholar]
- Wade, D. Fuel moisture and prescribed burning. South. Fire Exch. Fact Sheet 2013, 5, 5–8. [Google Scholar]
- Boyer, W.D. Volume Growth Loss: A Hidden Cost of Periodic Prescribed Burning in Longleaf Pine? South. J. Appl. For. 1987, 11, 154–157. [Google Scholar]
- Streng, D.R.; Glitzenstein, J.S.; Platt, W.J. Evaluating effects of season of burn in longleaf pine forests: A critical literature review and some results from ongoing long-term study. In Proceedings of 18th Tall Timbers Fire Ecology Conference; Hermann, S.M., Ed.; Tall Timbers Research Station: Tallahassee, FL, USA, 1993; pp. 227–263. [Google Scholar]
- Robertson, K.M.; Hmielowski, T.L. Effects of fire frequency and season on resprouting of woody plants in southeastern US pine-grassland communities. Oecologia 2014, 174, 765–776. [Google Scholar] [CrossRef] [PubMed]
- Melvin, M.; McIntyre, R.K. Air quality and human health challenges to prescribed fire. In Ecological Restoration and Management of Longleaf Pine Forests; Kirkman, L.K., Jack, S.B., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 255–266. ISBN 9781498748186. [Google Scholar]
- Chiodi, A.M.; Larkin, N.S.; Varner, J.M. An analysis of Southeastern US prescribed burn weather windows: Seasonal variability and El Niño associations. Int. J. Wildl. Fire 2018, 27, 176–189. [Google Scholar] [CrossRef]
- Fill, J.M.; Moule, B.M.; Varner, J.M.; Mousseau, T.A. Flammability of the keystone savanna bunchgrass Aristida stricta. Plant Ecol. 2016, 217, 331–342. [Google Scholar] [CrossRef]
- Fonda, R.W. Burning Characteristics of Needles from Eight Pine Species. For. Sci. 2001, 47, 390–396. [Google Scholar]
- Wenk, E.S.; Wang, G.G.; Walker, J.L. Understory fuel variation at the Carolina Sandhills National Wildlife Refuge: A description of chemical and physical properties. In Proceedings of the 15th Biennial Southern Silvicultural Research Conference, Asheville, USA, 2013; Guldin, J.M., Ed.; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2013; Volume 175, pp. 351–356. [Google Scholar]
- Loudermilk, E.L.; O’Brien, J.J.; Mitchell, R.J.; Cropper, W.P.; Hiers, J.K.; Grunwald, S.; Grego, J.; Fernandez-Diaz, J.C. Linking complex forest fuel structure and fire behaviour at fine scales. Int. J. Wildl. Fire 2012, 21, 882–893. [Google Scholar] [CrossRef]
- Parrott, R.T. A Study of Wiregrass (Aristida Stricta Michx.) with Particular Reference to Fire. Master’s Thesis, Duke University, Durham, NC, USA, 1967. [Google Scholar]
- Hendricks, J.J.; Wilson, C.A.; Boring, L.R. Foliar litter position and decomposition in a fire-maintained longleaf pine-wiregrass ecosystem. Can. J. For. Res. 2002, 32, 928–941. [Google Scholar] [CrossRef]
- Williamson, G.B.; Black, E.M. High temperature of forest fires under pines as a selective advantage over oaks. Nature 1981, 293, 643–644. [Google Scholar] [CrossRef]
- Hiers, J.K.; O’Brien, J.J.; Mitchell, R.J.; Grego, J.M.; Loudermilk, E.L. The wildland fuel cell concept: An approach to characterize fine-scale variation in fuels and fire in frequently burned longleaf pine forests. Int. J. Wildl. Fire 2009, 18, 315–325. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.J.; Jack, S.B.; Engstrom, R.T. Silviculture that sustains: The nexus between silviculture, frequent prescribed fire, and conservation of biodiversity in longleaf pine forests of the southeastern United States. Can. J. For. Res. 2006, 36, 2724–2736. [Google Scholar] [CrossRef]
- Mitchell, R.J.; Hiers, J.K.; O’Brien, J.; Starr, G. Ecological forestry in the Southeast: Understanding the ecology of fuels. J. For. 107391-397 2009, 107, 391–397. [Google Scholar]
- Nyland, R.D. Silviculture: Concepts and Applications, 2nd ed.; McGraw-Hill: Boston, MA, USA, 2002; ISBN 9780073661902. [Google Scholar]
- Kirkman, L.K.; Giencke, L.M. Restoring and managing a diverse ground cover. In Ecological Restoration and Management of Longleaf Pine Forests; Kirkman, L.K., Jack, S.B., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 207–232. [Google Scholar]
- Miller, D.L.; Mora, C.I.; Grissino-Mayer, H.D.; Mock, C.J.; Uhle, M.E.; Sharp, Z. Tree-ring isotope records of tropical cyclone activity. Proc. Natl. Acad. Sci. USA 2006, 103, 14294–14297. [Google Scholar] [CrossRef] [PubMed]
- Hmielowski, T.L.; Robertson, K.M.; Platt, W.J. Influence of season and method of topkill on resprouting characteristics and biomass of Quercus nigra saplings from a southeastern U.S. pine-grassland ecosystem. Plant Ecol. 2014, 215, 1221–1231. [Google Scholar] [CrossRef]
- Platt, W.J.; Ellair, D.P.; Huffman, J.M.; Potts, S.E.; Beckage, B. Pyrogenic fuels produced by savanna trees can engineer humid savannas. Ecol. Monogr. 2016, 86, 352–372. [Google Scholar] [CrossRef]
- Palik, B.J.B.J.; Pederson, N. Overstory mortality and canopy disturbances in longleaf pine ecosystems. Can. J. For. Res. 1996, 26, 2035–2047. [Google Scholar] [CrossRef]
- Pederson, N.; Varner, J.M.; Palik, B.J. Canopy disturbance and tree recruitment over two centuries in a managed longleaf pine landscape. For. Ecol. Manag. 2008, 254, 85–95. [Google Scholar] [CrossRef]
- NCEI Monthly Normals, Albany SW Georgia Regional Airport, GA. Available online: https://www.ncdc.noaa.gov/cdo-web/datasets 9 (accessed on 30 November 2016).
- McIntyre, R.K.; Jack, S.B.; Mitchell, R.J.; Kevin Hiers, J.; Neel, W.L. Multiple Value Management: The Stoddard-Neel Approach to Ecological Forestry in Longleaf Pine Grasslands, 1st ed.; Joseph, W., Ed.; Jones Ecological Research Center: Newton, MA, USA, 2008. [Google Scholar]
- Palik, B.; Mitchell, R.J.; Pecot, S.; Battaglia, M.; Pu, M. Spatial Distribution of Overstory Retention Influences Resources and Growth of Longleaf Pine Seedlings. Ecol. Appl. 2003, 13, 674–686. [Google Scholar] [CrossRef]
- Stoll, P.; Weiner, J.; Schmid, B. Growth variation in a naturally established population of Pinus sylvestris. Ecology 1994, 75, 660–670. [Google Scholar] [CrossRef]
- Berg, E.E.; Hamrick, J.L. Spatial and Genetic Structure of Two Sandhills Oaks: Quercus laevis and Quercus margaretta (Fagaceae). Am. J. Bot. 1994, 81, 7–14. [Google Scholar] [CrossRef]
- Cavender-bares, A.J.; Kitajima, K.; Bazzaz, F.A. Multiple Trait Associations in Relation to Habitat Differentiation among 17 Floridian Oak Species. Ecol. Monogr. 2004, 74, 635–662. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 1. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 15 May 2017).
- Christensen, R.H.B. Ordinal—Regression Models for Ordinal Data. Available online: https://cran.r-project.org/package=ordinal (accessed on 15 May 2017).
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2000; ISBN 0470582472. [Google Scholar]
- Gilliam, F.S.; Platt, W.J. Effects of Long-Term Fire Exclusion on Tree Species Composition and Stand Structure in an Reviewed work (s): Effects of long-term fire exclusion on tree species composition and stand structure in an old-growth Pinus palustris (Longleaf pine) forest. Plant Ecol. 1999, 140, 15–26. [Google Scholar] [CrossRef]
- Jack, S.B.; Mitchell, R.J.; Pecot, S.D. Silvicultural Alternatives in a Longleaf Pine/Wiregrass Woodland in Southwest Georgia: Understory Hardwood Response To Harvest-Created Gaps. In Proceedings of the 13th Southern Silvicultural Research Conference, Asheville, NC, USA, 2006; Connor, K.F., Ed.; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2006; pp. 85–89. [Google Scholar]
- Stambaugh, M.C.; Varner, J.M.; Noss, R.F.; Dey, D.C.; Christensen, N.L.; Baldwin, R.F.; Guyette, R.P.; Hanberry, B.B.; Harper, C.A.; Lindblom, S.G.; et al. Clarifying the role of fire in the deciduous forests of eastern North America: Reply to Matlack. Conserv. Biol. 2015, 29, 942–946. [Google Scholar] [CrossRef] [PubMed]
- Grady, J.M.; Hoffmann, W.A. Caught in a fire trap: Recurring fire creates stable size equilibria in woody resprouters. Ecology 2012, 93, 2052–2060. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.D.; Arthur, M.A.; Loftis, D.L.; Green, S.R. Survival and growth of upland oak and co-occurring competitor seedlings following single and repeated prescribed fires. For. Ecol. Manag. 2008, 256, 1021–1030. [Google Scholar] [CrossRef]
- Menaut, J.C.; Gignoux, J.; Prado, C.; Clobert, J. Tree Community Dynamics in a Humid Savanna of the Cote-d’Ivoire: Modelling the Effects of Fire and Competition with Grass and Neighbours. J. Biogeogr. 1990, 17, 471–481. [Google Scholar] [CrossRef]
- Keyser, T.L.; Loftis, D.L. Stump sprouting of 19 upland hardwood species 1 year following initiation of a shelterwood with reserves silvicultural system in the southern Appalachian Mountains, USA. New For. 2015, 46, 449–464. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. [Technical Coordinators] Silvics of North America: Volume 2. Hardwoods; Agricultural Handbook; United States Department of Agriculture (USDA), Forest Service: Washington, DC, USA, 1990; Volume 654.
- Meier, A.R.; Saunders, M.R.; Michler, C.H. Epicormic buds in trees: A review of bud establishment, development and dormancy release. Tree Physiol. 2012, 32, 565–584. [Google Scholar] [CrossRef] [PubMed]
- Lavoie, M.; Starr, G.; Mack, M.C.; Martin, T.A.; Gholz, H.L. Effects of a prescribed fire on understory vegetation, carbon pools, and soil nutrients in a longleaf pine-slash pine forest in Florida. Nat. Areas J. 2010, 30, 82–94. [Google Scholar] [CrossRef]
- Varner, J.M.; Kane, J.M.; Hiers, J.K.; Kreye, J.K.; Veldman, J.W. Suites of fire-adapted traits of oaks in the Southeastern USA: Multiple strategies for persistence. Fire Ecol. 2016, 12, 48–64. [Google Scholar] [CrossRef]
- Graves, S.J.; Rifai, S.W.; Putz, F.E. Outer bark thickness decreases more with height on stems of fire-resistant than fire-sensitive Floridian oaks (Quercus spp.; Fagaceae). Am. J. Bot. 2014, 101, 2183–2188. [Google Scholar] [CrossRef] [PubMed]
- Lawes, M.J.; Adie, H.; Russell-Smith, J.; Murphy, B.; Midgley, J.J. How do small savanna trees avoid stem mortality by fire? The roles of stem diameter, height and bark thickness. Ecosphere 2011, 2, 1–13. [Google Scholar] [CrossRef]
- Battaglia, M.A.; Mitchell, R.J.; Mou, P.P.; Pecot, S.D. Light Transmittance Estimates in a Longleaf Pine Woodland. For. Sci. 2003, 49, 752–762. [Google Scholar]
- Pecot, S.D.; Mitchell, R.J.; Palik, B.J.; Moser, E.B.; Hiers, J.K. Competitive responses of seedlings and understory plants in longleaf pine woodlands: Separating canopy influences above and below ground. Can. J. For. Res. 2007, 37, 634–648. [Google Scholar] [CrossRef]
- Wright, A.; Schnitzer, S.A.; Reich, P.B. Living close to your neighbors-the importance of both competition and facilitation in plant communities. Ecology 2014, 95, 2213–2223. [Google Scholar] [CrossRef] [PubMed]
- Werner, P.A. Growth of juvenile and sapling trees differs with both fire season and understorey type: Trade-offs and transitions out of the fire trap in an Australian savanna. Austral Ecol. 2012, 37, 644–657. [Google Scholar] [CrossRef]
- Fill, J.M.; Glitzenstein, J.S.; Streng, D.R.; Stowe, J.; Mousseau, T.A. Wiregrass ( Aristida beyrichiana ) May Limit Woody Plant Encroachment in Longleaf Pine ( Pinus palustris ) Ecosystems. Am. Midl. Nat. 2017. [Google Scholar] [CrossRef]
- Markewitz, D.; Sartori, F.; Craft, C. Soil change and carbon storage in longleaf pine stands planted on marginal agricultural lands. Ecol. Appl. 2002, 12, 1276–1285. [Google Scholar] [CrossRef]
- Rice, E.L. Allelopathic effects of Andropogon virginicus and its persistence in old fields. Am. J. Bot. 1972, 59, 752–755. [Google Scholar] [CrossRef]
- Daneshgar, P.; Jose, S. Role of species identity in plant invasions: Experimental test using Imperata cylindrica. Biol. Invasions 2009, 11, 1431–1440. [Google Scholar] [CrossRef]
- Gagnon, P.R.; Passmore, H.A.; Platt, W.J.; Myers, J.A.; Paine, C.E.T.; Harms, K.E. Does pyrogenicity protect burning plants? Ecology 2010, 91, 3481–3486. [Google Scholar] [CrossRef] [PubMed]
- Peet, R.; Allard, D. Longleaf pine vegetation of the southern Atlantic and eastern Gulf Coast regions: A preliminary classification. In Proceedings of 18th Tall Timbers Fire Ecology Conference, Tallahassee, USA, 1993; Hermann, S.M., Ed.; Tall Timbers Research Station: Tallahassee, FL, USA, 1993; pp. 45–81. [Google Scholar]
- Brockway, D.G.; Outcalt, K.W. Gap-phase regeneration in longleaf pine wiregrass ecosystems. For. Ecol. Manag. 1998, 106, 125–139. [Google Scholar] [CrossRef]
- McGuire, J.P.; Mitchell, R.J.; Moser, E.B.; Pecot, S.D.; Gjerstad, D.H.; Hedman, C.W. Gaps in a gappy forest: Plant resources, longleaf pine regeneration, and understory response to tree removal in longleaf pine savannas. Can. J. For. Res. 2001, 31, 765–778. [Google Scholar] [CrossRef]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).