Assessing Climate Change Impact on Forest Habitat Suitability and Diversity in the Korean Peninsula

 ,
, 
 ,
, 
Abstract
1. Introduction
2. Data and Methods
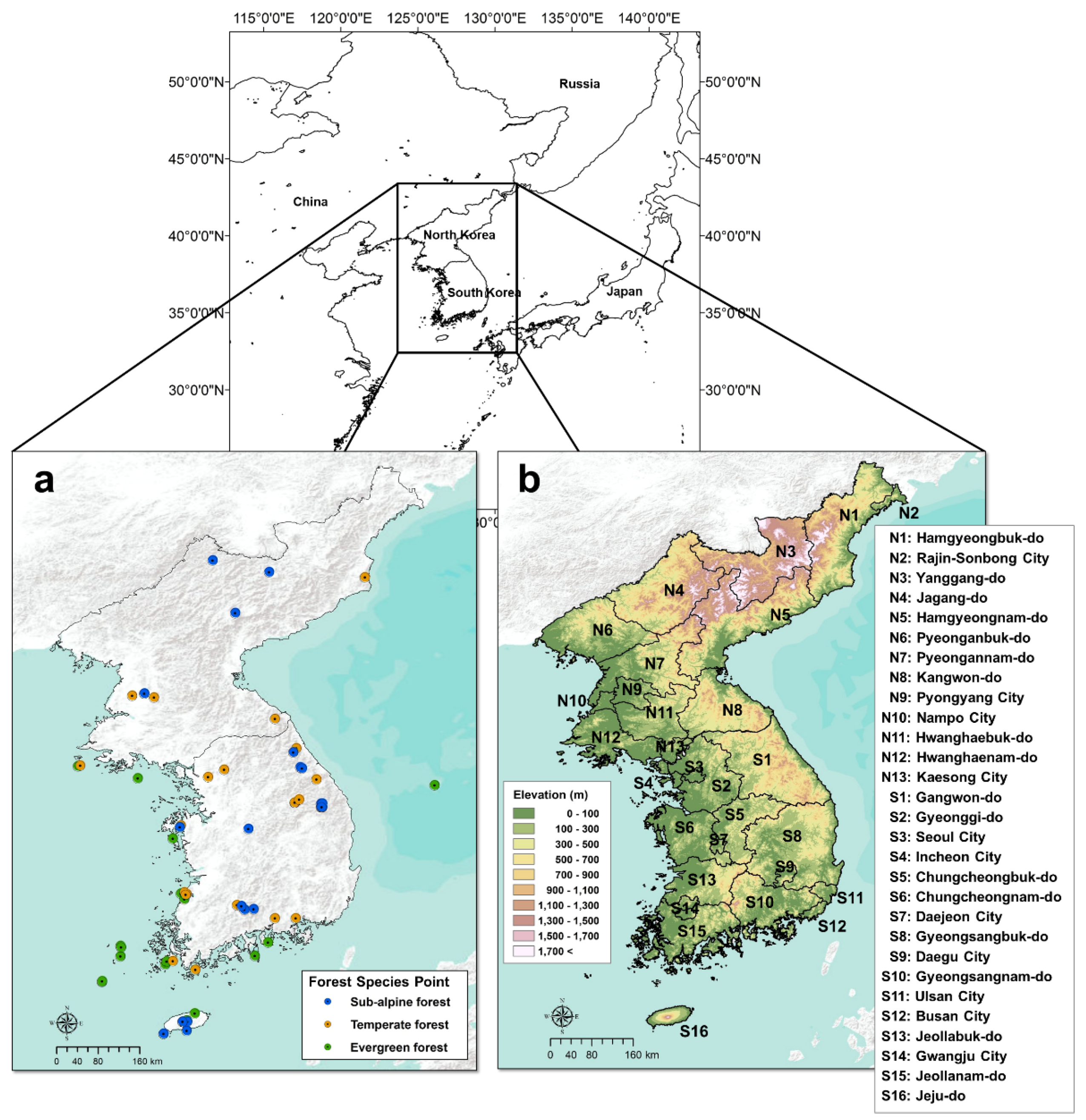
2.1. Study Area
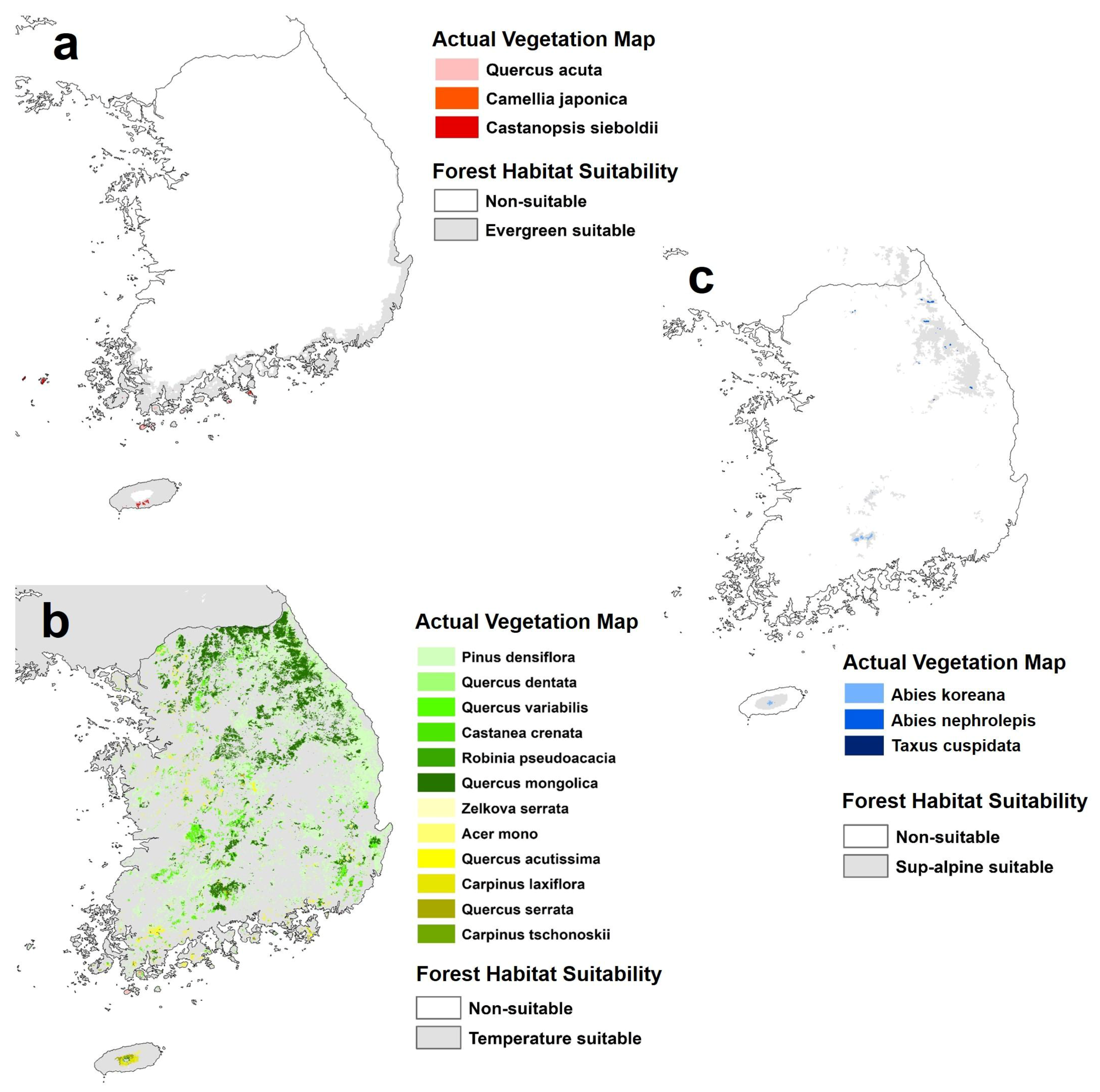
2.2. Forest Habitat Suitability, Diversity, and Forest Species Data
- Warm temperate evergreen forest: Camellia japonica L., Castanopsis sieboldii (Makino) Hatus, Quercus acuta Thunb., Machilus thunbergii Sieb. & Zucc., and Pinus thunbergii Parl.
- Temperate forest: Pinus densiflora Siebold & Zucc., Quercus dentate Thunb., Quercus variabilis Blume, Castanea crenata Siebold & Zucc., Robinia pseudoacacia L., Quercus mongolica Fisch. ex Ledeb., Zelkova serrate (Thunb.) Makino, Acer mono Maxim., Quercus acutissima Carruth., Carpinus laxiflora (Siebold & Zucc.) Blume, Quercus serrate Murray, and Carpinus tschonoskii Maxim.(Betulaceae)
- Subalpine forest: Abies koreana E.H.Wilson, Abies nephrolepis (Trautv. ex Maxim.) Maxim., and Taxus cuspidate Siebold & Zucc.
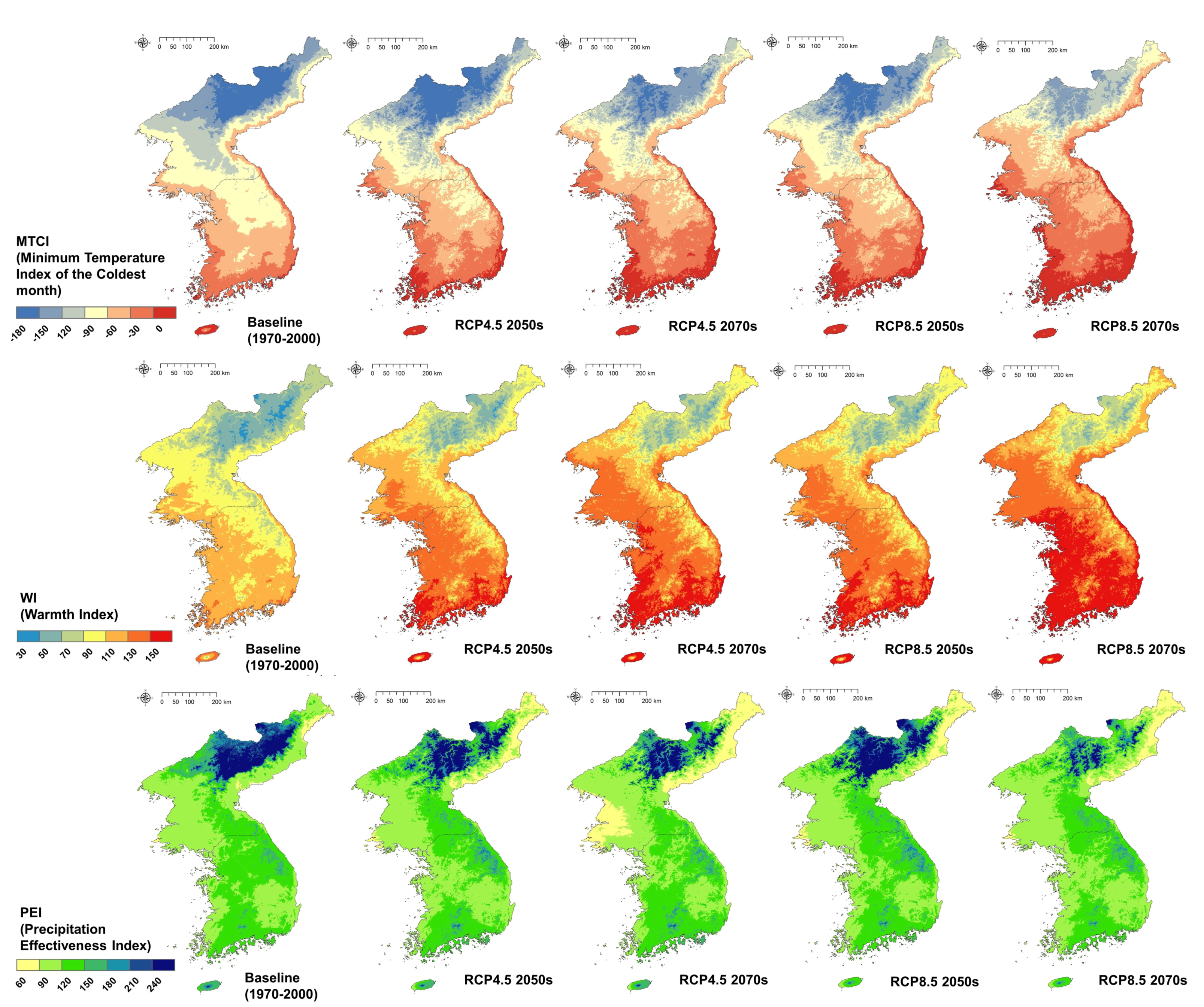
2.3. Climate Data
2.4. MaxEnt Modeling and Classification
2.5. Evaluation of Model Performance
3. Results and Discussion
3.1. Calculating Three Climate Indices
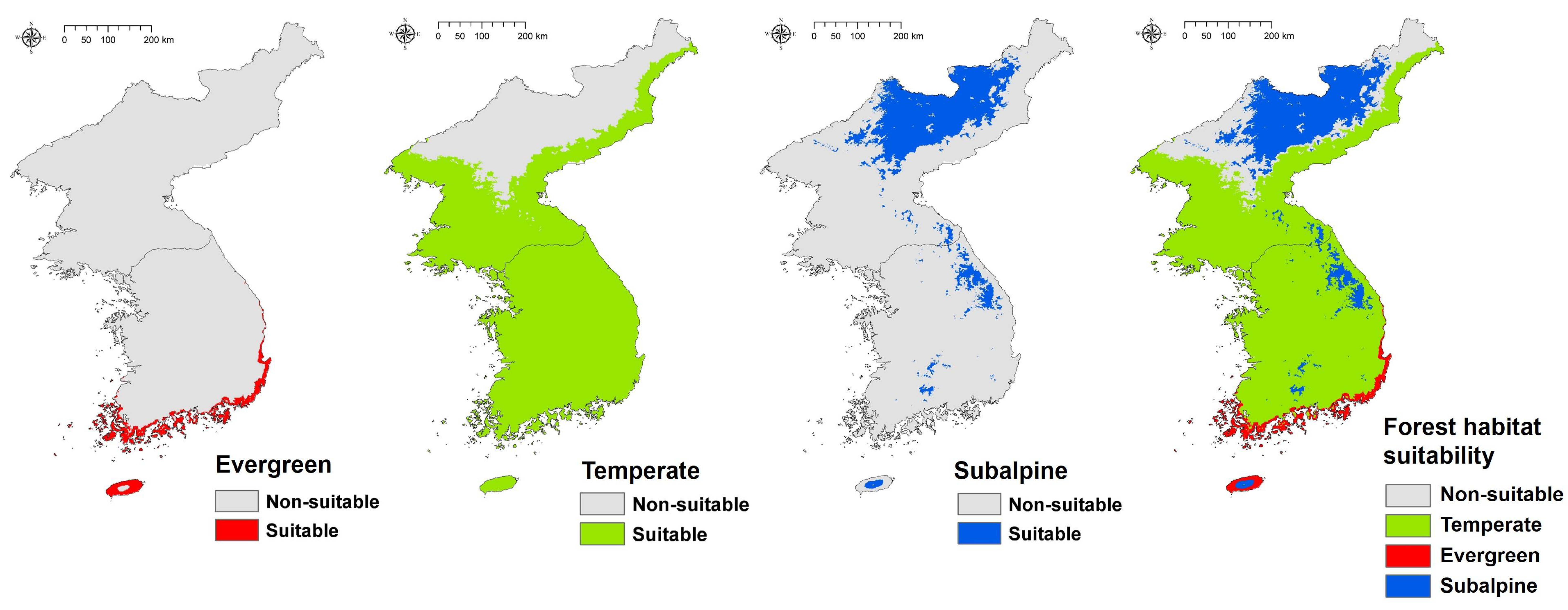
3.2. Estimating Baseline Forest Habitat Suitability
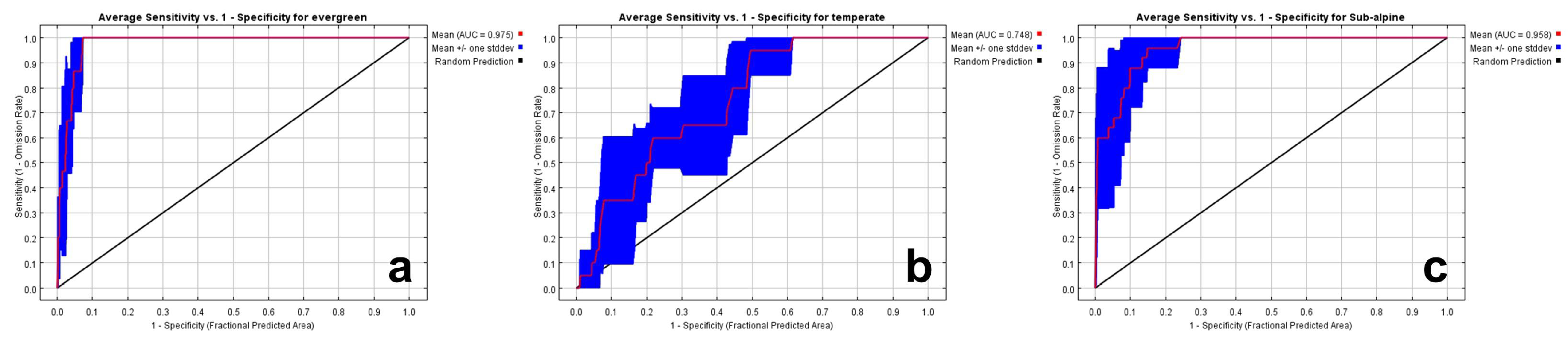
3.3. Evaluation of Model Performance
3.3.1. Evaluation Using AUC and ROC
3.3.2. Evaluation Using the Actual Vegetation Map of South Korea
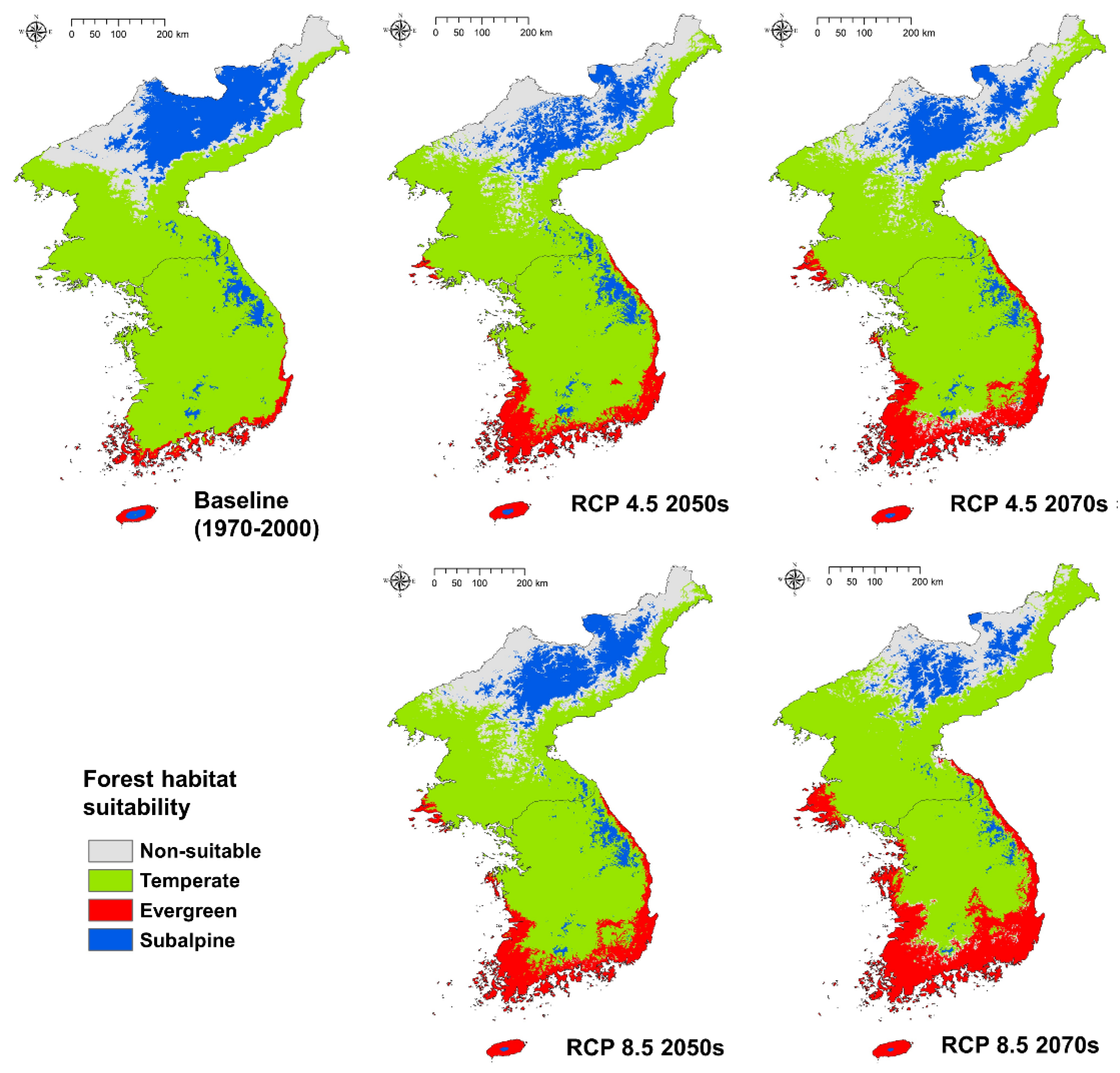
3.4. Climate Change Impact on Forest Habitat Suitability
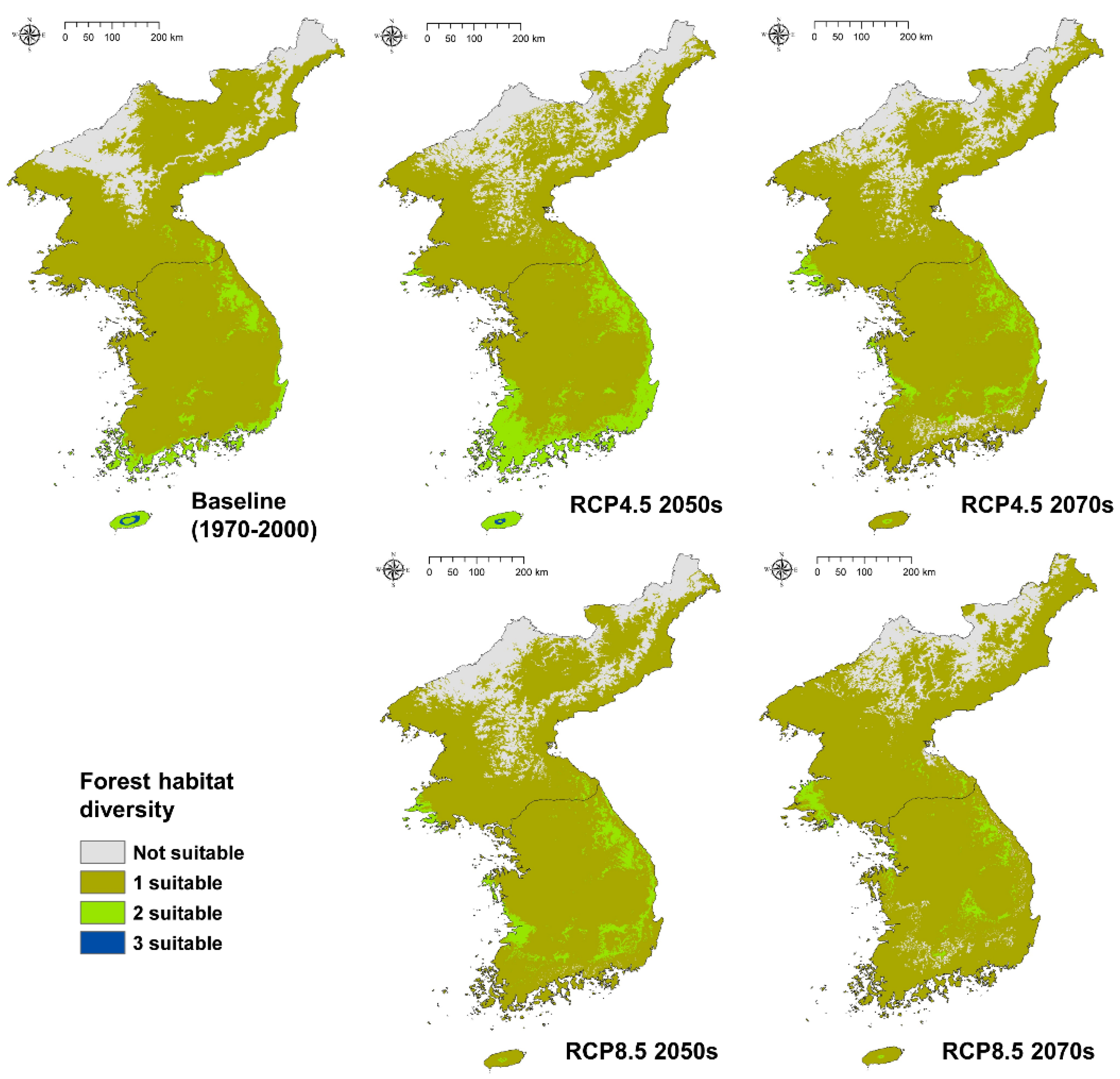
3.5. Converting to Forest Habitat Diversity Analysis under Climate Change
3.6. Implications of Forest Suitability and Diversity Assessment Attributable to Climate Change
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Intergovernmental Panel on Climate Change (IPCC). Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Anadon, J.D.; Sala, O.E.; Maestre, F.T. Climate change will increase savannas at the expense of forests and treeless vegetation in tropical and subtropical Americas. J. Ecol. 2014, 102, 1363–1373. [Google Scholar] [CrossRef]
- Wernberg, T.; Bennett, S.; Babcock, R.C.; de Bettignies, T.; Cure, K.; Depczynski, M.; Dufois, F.; Fromont, J.; Fulton, C.J.; Hovey, R.K.; et al. Climate-driven regime shift of a temperate marine ecosystem. Science 2016, 353, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Schlaepfer, D.R.; Bradford, J.B.; Lauenroth, W.K.; Munson, S.M.; Tietjen, B.; Hall, S.A.; Wilson, S.D.; Duniway, M.C.; Jia, G.; Pyke, D.A.; et al. Climate change reduces extent of temperate drylands and intensifies drought in deep soils. Nat. Commun. 2017, 8, 14196. [Google Scholar] [CrossRef] [PubMed]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; de Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Torres, D.A.; Navarro-Sigüenza, A.G.; Santiago-Alarcon, D.; Rojas-Soto, O.R. Response of the endangered tropical dry forests to climate change and the role of Mexican Protected Areas for their conservation. Glob. Chang. Biol. 2016, 22, 364–379. [Google Scholar] [CrossRef] [PubMed]
- Dyderski, M.K.; Paź, S.; Frelich, L.E.; Jagodziński, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2017, 24, 1150–1163. [Google Scholar] [CrossRef] [PubMed]
- Akhter, S.; McDonald, M.A.; van Breugel, P.; Sohel, S.; Kjær, E.D.; Mariott, R. Habitat distribution modelling to identify areas of high conservation value under climate change for Mangifera sylvatica Roxb. of Bangladesh. Land Use Policy 2017, 60, 223–232. [Google Scholar] [CrossRef]
- Mair, L.; Harrison, P.J.; Räty, M.; Bärring, L.; Strandberg, G.; Snäll, T. Forest management could counteract distribution retractions forced by climate change. Ecol. Appl. 2017, 27, 1485–1497. [Google Scholar] [CrossRef] [PubMed]
- Clark, J.S.; Gelfand, A.E.; Woodall, C.W.; Zhu, K. More than the sum of the parts: Forest climate response from joint species distribution models. Ecol. Appl. 2014, 24, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Koo, K.A.; Patten, B.C.; Madden, M. Predicting effects of climate change on habitat suitability of red spruce (Picea rubens Sarg.) in the southern Appalachian Mountains of the USA: Understanding complex systems mechanisms through modeling. Forests 2015, 6, 1208–1226. [Google Scholar] [CrossRef]
- De Rigo, D.; Caudullo, G.; San-Miguel-Ayanz, J.; Barredo, J.I. Robust Modelling of the Impacts of Climate Change on the Habitat Suitability of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2017; 58p. [Google Scholar]
- Choi, S.; Lee, W.K.; Kwak, D.A.; Lee, S.; Son, Y.; Lim, J.H.; Saborowski, J. Predicting forest cover changes in future climate using hydrological and thermal indices in South Korea. Clim. Res. 2011, 49, 229–245. [Google Scholar] [CrossRef][Green Version]
- Nam, K.; Lee, W.K.; Kim, M.; Kwak, D.A.; Byun, W.H.; Yu, H.; Kwak, H.; Kwon, T.; Sung, J.; Chung, D.J.; Lee, S.H. Spatio-temporal change in forest cover and carbon storage considering actual and potential forest cover in South Korea. Sci. China Life Sci. 2015, 58, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.; Minor, E.S.; Lee, D.; Park, C.R. Predicting impacts of climate change on habitat connectivity of Kalopanax septemlobus in South Korea. Acta Oecol. 2016, 71, 31–38. [Google Scholar] [CrossRef]
- Park, S.U.; Koo, K.A.; Seo, C.; Hong, S. Climate-related range shifts of Ardisia japonica in the Korean Peninsula: A role of dispersal capacity. J. Ecol. Environ. 2017, 41, 38. [Google Scholar] [CrossRef]
- Koo, K.A.; Park, S.U.; Seo, C. Effects of Climate Change on the Climatic Niches of Warm-Adapted Evergreen Plants: Expansion or Contraction? Forests 2017, 8, 500. [Google Scholar] [CrossRef]
- Huang, J.; Li, G.; Li, J.; Zhang, X.; Yan, M.; Du, S. Projecting the Range Shifts in Climatically Suitable Habitat for Chinese Sea Buckthorn under Climate Change Scenarios. Forests 2018, 9, 9. [Google Scholar] [CrossRef]
- Alsterberg, C.; Roger, F.; Sundbäck, K.; Juhanson, J.; Hulth, S.; Hallin, S.; Gamfeldt, L. Habitat diversity and ecosystem multifunctionality—The importance of direct and indirect effects. Sci. Adv. 2017, 3, e1601475. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, M.L. Species diversity gradients: We know more and less than we thought. J. Mammal. 1992, 73, 715–730. [Google Scholar] [CrossRef]
- Paquette, A.; Messier, C. The effect of biodiversity on tree productivity: From temperate to boreal forests. Glob. Ecol. Biogeogr. 2011, 20, 170–180. [Google Scholar] [CrossRef]
- Müllerová, J.; Hédl, R.; Szabó, P. Coppice abandonment and its implications for species diversity in forest vegetation. For. Ecol. Manag. 2015, 343, 88–100. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Lee, W.K.; Choi, G.M.; Song, C.; Lim, C.H.; Moon, J.; Piao, D.; Kraxner, F.; Shividenko, A.; Forsell, N. Modeling stand-level mortality based on maximum stem number and seasonal temperature. For. Ecol. Manag. 2017, 386, 37–50. [Google Scholar] [CrossRef]
- Yoo, S.; Lee, W.K.; Kim, M.; Lim, C.H.; Song, C.; Kim, S.J. Predicting Endangered Coniferous Tree Species Distribution under Climate Change: Implication of HyTAG Model. J. Environ. Impact Assess. 2018. under review. [Google Scholar]
- Lim, C.H.; Kim, S.H.; Choi, Y.; Kafatos, M.C.; Lee, W.K. Estimation of the Virtual Water Content of Main Crops on the Korean Peninsula Using Multiple Regional Climate Models and Evapotranspiration Methods. Sustainability 2017, 9, 1172. [Google Scholar] [CrossRef]
- Lamchin, M.; Lee, W.K.; Jeon, S.W.; Wang, S.W.; Lim, C.H.; Song, C.; Sung, M. Long-term trend and correlation between vegetation greenness and climate variables in Asia based on satellite data. Sci. Total Environ. 2018, 618, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.H.; Choi, Y.; Kim, M.; Lee, S.J.; Folberth, C.; Lee, W.K. Spatially Explicit Assessment of Agricultural Water Equilibrium in the Korean Peninsula. Sustainability 2018, 10, 201. [Google Scholar] [CrossRef]
- Koo, K.A.; Park, S.U.; Hong, S.; Jang, I.; Seo, C. Future distributions of warm-adapted evergreen trees, Neolitsea sericea and Camellia japonica under climate change: Ensemble forecasts and predictive uncertainty. Ecol. Res. 2018, 33, 313–325. [Google Scholar] [CrossRef]
- Lim, C.H.; Choi, Y.; Kim, M.; Jeon, S.W.; Lee, W.K. Impact of deforestation on agro-environmental variables in cropland, North Korea. Sustainability 2017, 9, 1354. [Google Scholar] [CrossRef]
- Lim, C.H. A Cross-Sectoral Approach in the Water-Food-Ecosystem Nexus to Climate Change in the Korean Peninsula. Ph.D. Thesis, Korea University, Seoul, Korea, 2017. [Google Scholar]
- Chao, A.; Chiu, C.H.; Jost, L. Statistical challenges of evaluating diversity patterns across environmental gradients in mega-diverse communities. J. Veg. Sci. 2016, 27, 437–438. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Bachelet, D.; Lenihan, J.M.; Daly, C.; Neilson, R.P.; Ojima, D.S.; Parton, W.J. MC1: A Dynamic Vegetation Model for Estimating the Distribution of Vegetation and Associated Ecosystem Fluxes of Carbon, Nutrients, and Water; Pacific Northwest Station General Technical Report PNW-GTR-508; USDA: Washington, DC, USA, 2001.
- Neilson, R.P. A model for predicting continental-scale vegetation distribution and water balance. Ecol. Appl. 1995, 5, 362–385. [Google Scholar] [CrossRef]
- Kira, T. A New Classification of Climate in Eastern Asia as the Basis for Agricultural Geography; Horticultural Institute Kyoto University: Kyoto, Japan, 1945. [Google Scholar]
- Yim, Y.J. Distribution of forest vegetation and climate in the Korean Peninsula: III. Distribution of tree species along the thermal gradient. Jpn. J. Ecol. 1977, 27, 177–189. [Google Scholar]
- Thornthwaite, C.W. The climates of North America: According to a new classification. Geogr. Rev. 1931, 21, 633–655. [Google Scholar] [CrossRef]
- Vörösmarty, C.J.; Douglas, E.M.; Green, P.A.; Revenga, C. Geospatial indicators of emerging water stress: An application to Africa. Ambio 2005, 34, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Lim, C.H.; Ryu, J.; Jeon, S.W. Bioclimatic Classification of the Northeast Asia Reflecting Social Factors: Development and Characterization. Sustainability 2017, 9, 1137. [Google Scholar] [CrossRef]
- Lim, C.H.; Kim, G.S.; Lee, E.J.; Heo, S.; Kim, T.; Kim, Y.S.; Lee, W.K. Development on Crop Yield Forecasting Model for Major Vegetable Crops using Meteorological Information of Main Production Area. J. Clim. Chang. Res. 2016, 7, 193–203. [Google Scholar] [CrossRef]
- Deblauwe, V.; Barbier, N.; Couteron, P.; Lejeune, O.; Bogaert, J. The global biogeography of semi-arid periodic vegetation patterns. Glob. Ecol. Biogeogr. 2008, 17, 715–723. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with MaxEnt: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Chen, F.; Du, Y.; Niu, S.; Zhao, J. Modeling forest lightning fire occurrence in the Daxinganling Mountains of Northeastern China with MAXENT. Forests 2015, 6, 1422–1438. [Google Scholar] [CrossRef]
- Ikegami, M.; Jenkins, T.A. Estimate global risks of a forest disease under current and future climates using species distribution model and simple thermal model–Pine Wilt disease as a model case. For. Ecol. Manag. 2018, 409, 343–352. [Google Scholar] [CrossRef]
- Xiaofeng, L.; Yi, Q.; Diqiang, L.; Shirong, L.; Xiulei, W.; Bo, W.; Chunquan, Z. Habitat evaluation of wild Amur tiger (Panthera tigris altaica) and conservation priority setting in north-eastern China. J. Environ. Manag. 2011, 92, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Jenks, G.F. The Data Model Concept in Statistical Mapping. In International Yearbook of Cartography; George Philip: London, UK, 1967; Volume 7, pp. 186–190. [Google Scholar]
- Vilar, L.; Gómez, I.; Martínez-Vega, J.; Echavarría, P.; Riaño, D.; Martín, M.P. Multitemporal modelling of socio-economic wildfire drivers in central Spain between the 1980s and the 2000s: Comparing generalized linear models to machine learning algorithms. PLoS ONE 2016, 11, e0161344. [Google Scholar] [CrossRef] [PubMed]
- Kafatos, M.C.; Kim, S.H.; Lim, C.-H.; Kim, J.; Lee, W.-K. Responses of Agroecosystems to Climate Change: Specifics of Resilience in the Mid-Latitude Region. Sustainability 2017, 9, 1361. [Google Scholar] [CrossRef]
- Kim, T.; Song, C.; Lee, W.K.; Kim, M.; Lim, C.H.; Jeon, S.W.; Kim, J. Habitat Quality Valuation Using InVEST Model in Jeju Island. J. Korea Soc. Environ. Restor. Technol. 2015, 18, 1–11. [Google Scholar] [CrossRef]
- Cui, G.; Kwak, H.; Choi, S.; Kim, M.; Lim, C.H.; Lee, W.K.; Kim, J.; Chae, Y. Assessing vulnerability of forests to climate change in South Korea. J. For. Res. 2016, 27, 489–503. [Google Scholar] [CrossRef]
- Koo, K.A.; Kong, W.S.; Nibbelink, N.P.; Hopkinson, C.S.; Lee, J.H. Potential effects of climate change on the distribution of cold-tolerant evergreen broadleaved woody plants in the Korean Peninsula. PLoS ONE 2015, 10, e0134043. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.H; Lee, W.K. Climate-Environmental Impacts on Agriculture and Water Balance; LAP: Beau Bassin, Mauritius, 2018. [Google Scholar]
- Kim, M.; Lee, W.K.; Kim, Y.S.; Lim, C.H.; Song, C.; Park, T.; Son, Y.; Son, Y.M. Impact of thinning intensity on the diameter and height growth of Larix kaempferi stands in central Korea. For. Sci. Technol. 2016, 12, 77–87. [Google Scholar]
- Song, C.H.; Lee, W.K.; Choi, H.A.; Kim, J.; Jeon, S.W.; Kim, J.S. Spatial assessment of ecosystem functions and services for air purification of forests in South Korea. Environ. Sci. Policy 2016, 63, 27–34. [Google Scholar] [CrossRef]
- Kim, G.S.; Lim, C.H.; Kim, S.J.; Lee, J.; Son, Y.; Lee, W.K. Effect of national-scale afforestation on forest water supply and soil loss in South Korea, 1971–2010. Sustainability 2017, 9, 1017. [Google Scholar] [CrossRef]
- Lee, J.; Lim, C.H.; Kim, G.S.; Markandya, A.; Chowdhury, S.; Kim, S.J.; Lee, W.K.; Son, Y. Economic viability of the national-scale forestation program: The case of success in the Republic of Korea. Ecosyst. Serv. 2018, 29, 40–46. [Google Scholar] [CrossRef]
- Correia, R.A.; Bugalho, M.N.; Franco, A.M.; Palmeirim, J.M. Contribution of spatially explicit models to climate change adaptation and mitigation plans for a priority forest habitat. Mitig. Adapt. Strateg. Glob. Chang. 2018, 23, 371–386. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Type | Baseline | RCP 4.5 2050s | RCP 4.5 2070s | RCP 8.5 2050s | RCP 8.5 2070s |
|---|---|---|---|---|---|
| Evergreen | 8372 (3.68) | 22,302 (9.79) | 28,100 (12.34) | 27,254 (11.97) | 43,731 (19.21) |
| Temperate | 149,811 (65.79) | 139,074 (61.08) | 138,652 (60.09) | 132,375 (58.14) | 137,245 (60.28) |
| Subalpine | 39,128 (17.18) | 26,938 (11.83) | 24,907 (10.94) | 29,906 (13.13) | 16,779 (7.37) |
| Non-suitable | 30,383 (13.34) | 39,380 (17.29) | 36,035 (15.83) | 38,159 (16.76) | 29,939 (13.15) |
| Diversity | Baseline | RCP 4.5–2050s | RCP 4.5–2070s | RCP 8.5–2050s | RCP 4.5–2070s |
|---|---|---|---|---|---|
| Not suitable | 30,383 (13.34) | 39,926 (17.53) | 36,035 (15.83) | 38,159 (16.76) | 29,939 (13.15) |
| 1 suitable | 183,099 (80.41) | 159,309 (69.97) | 181,911 (79.89) | 177,624 (78.01) | 191,466 (84.09) |
| 2 suitable | 13,925 (6.12) | 28,304 (12.43) | 9748 (4.28) | 11,911 (5.23) | 6289 (2.76) |
| 3 suitable | 287 (0.13) | 155 (0.07) | - | - | - |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, C.-H.; Yoo, S.; Choi, Y.; Jeon, S.W.; Son, Y.; Lee, W.-K. Assessing Climate Change Impact on Forest Habitat Suitability and Diversity in the Korean Peninsula. Forests 2018, 9, 259. https://doi.org/10.3390/f9050259
Lim C-H, Yoo S, Choi Y, Jeon SW, Son Y, Lee W-K. Assessing Climate Change Impact on Forest Habitat Suitability and Diversity in the Korean Peninsula. Forests. 2018; 9(5):259. https://doi.org/10.3390/f9050259
Chicago/Turabian StyleLim, Chul-Hee, Somin Yoo, Yuyoung Choi, Seong Woo Jeon, Yowhan Son, and Woo-Kyun Lee. 2018. "Assessing Climate Change Impact on Forest Habitat Suitability and Diversity in the Korean Peninsula" Forests 9, no. 5: 259. https://doi.org/10.3390/f9050259
APA StyleLim, C.-H., Yoo, S., Choi, Y., Jeon, S. W., Son, Y., & Lee, W.-K. (2018). Assessing Climate Change Impact on Forest Habitat Suitability and Diversity in the Korean Peninsula. Forests, 9(5), 259. https://doi.org/10.3390/f9050259