Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers?



Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description
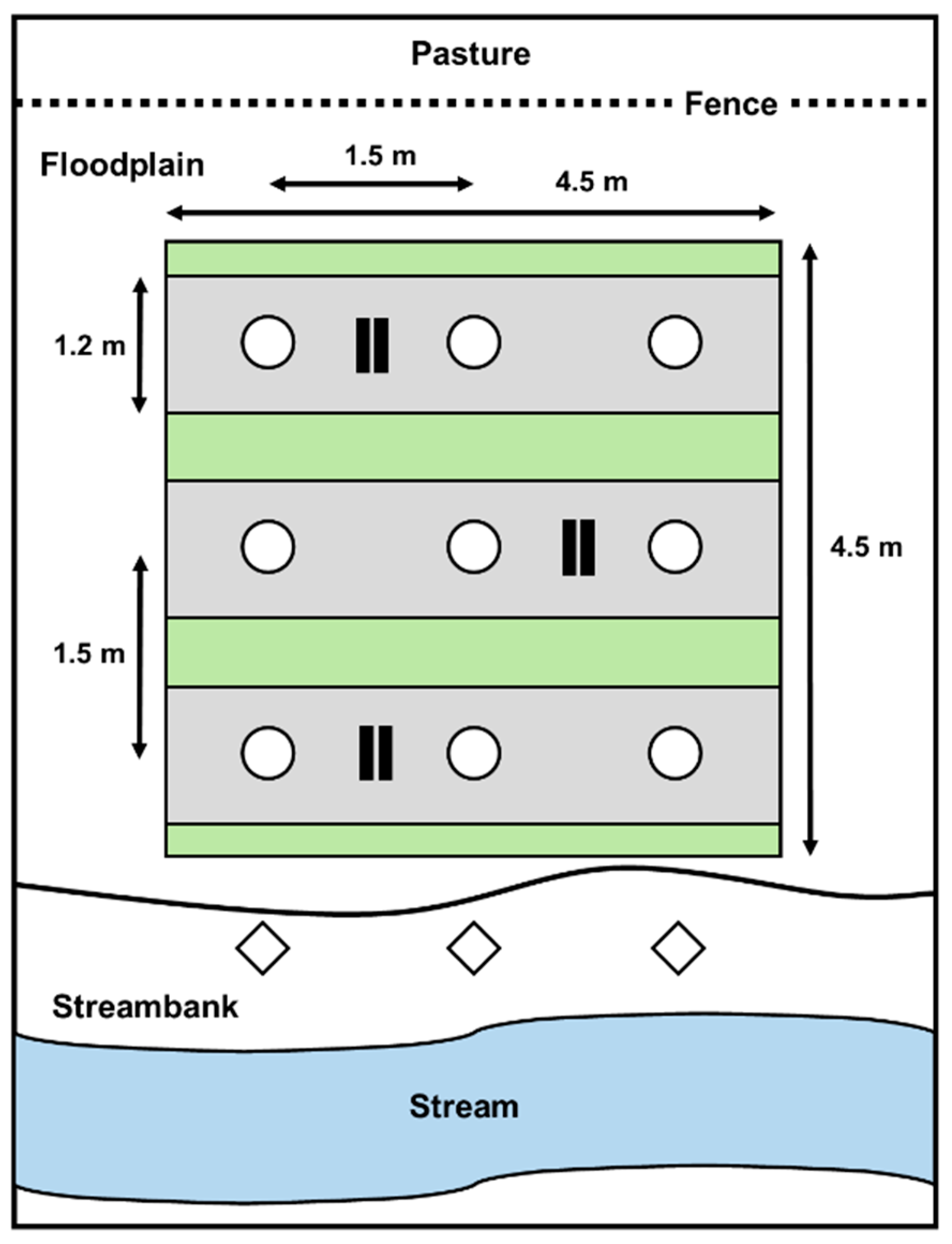
2.2. Experimental Design
2.3. Measurement of Soil Nutrient Dynamics
2.4. Growth and Survival Measurements
2.5. Statistical Analyses
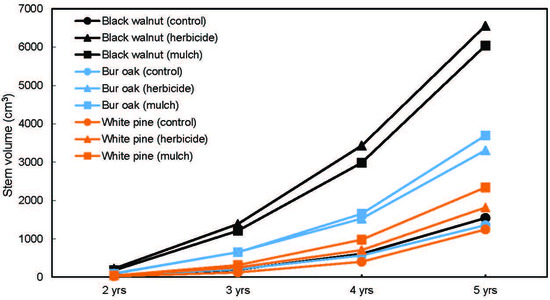
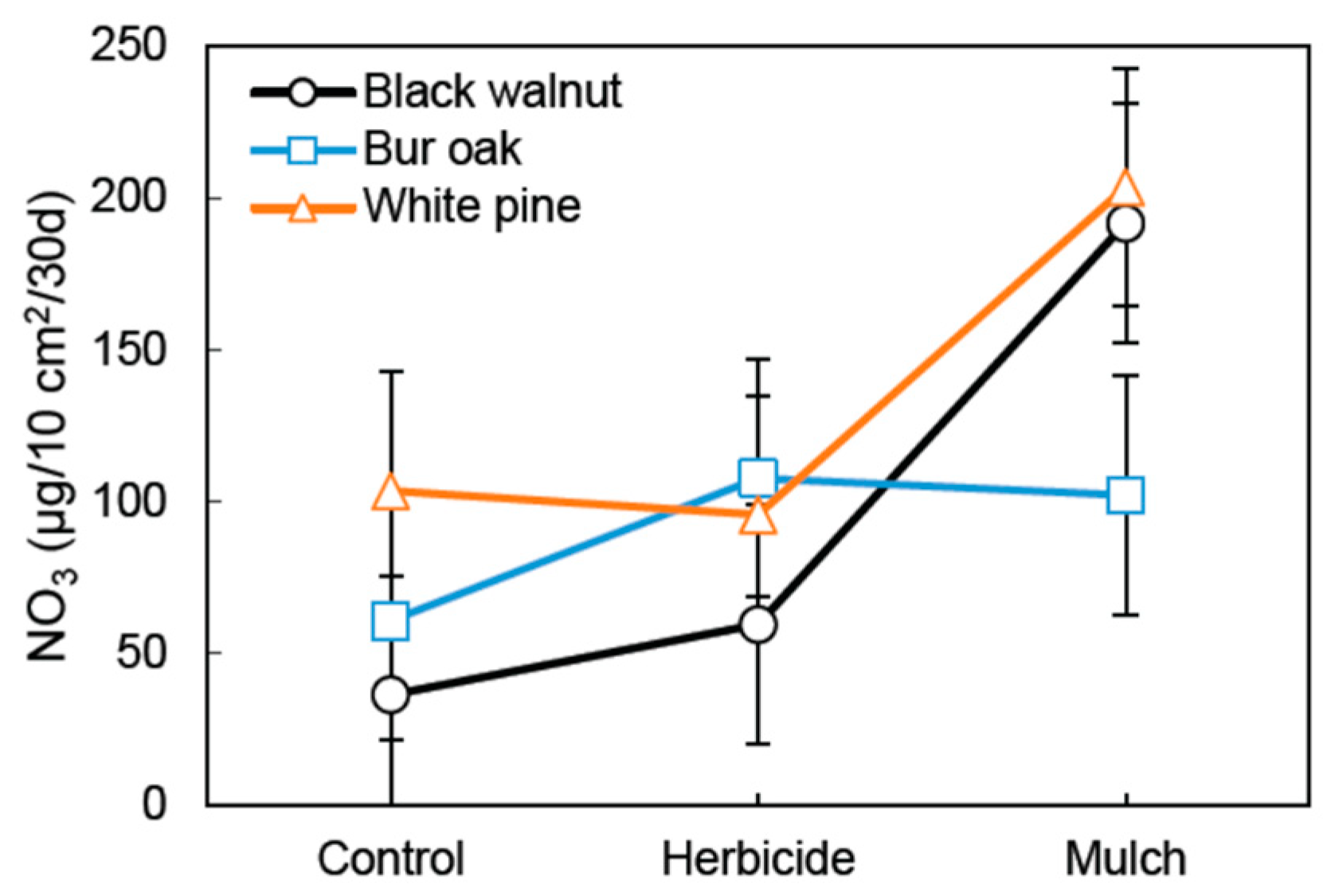
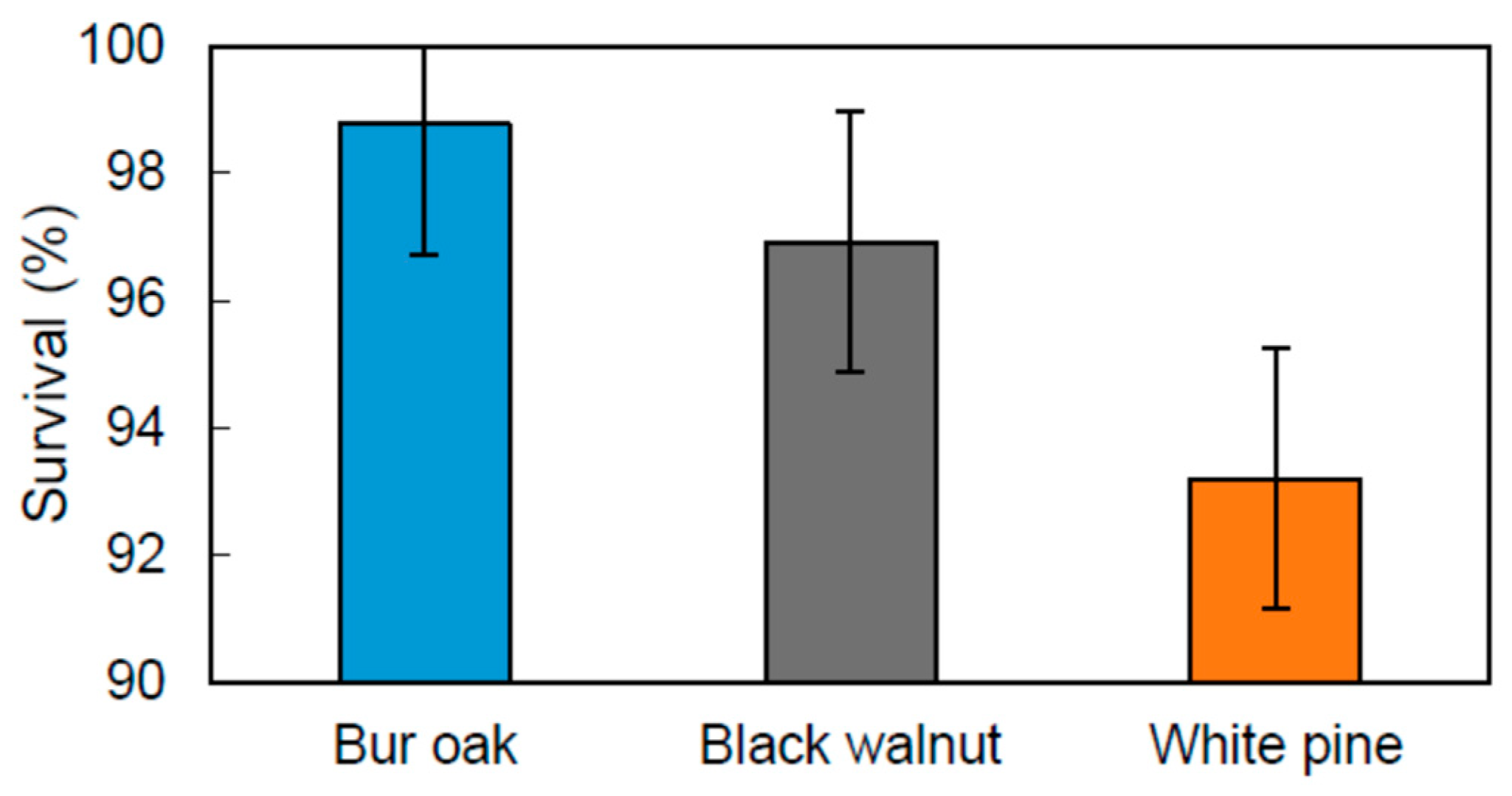
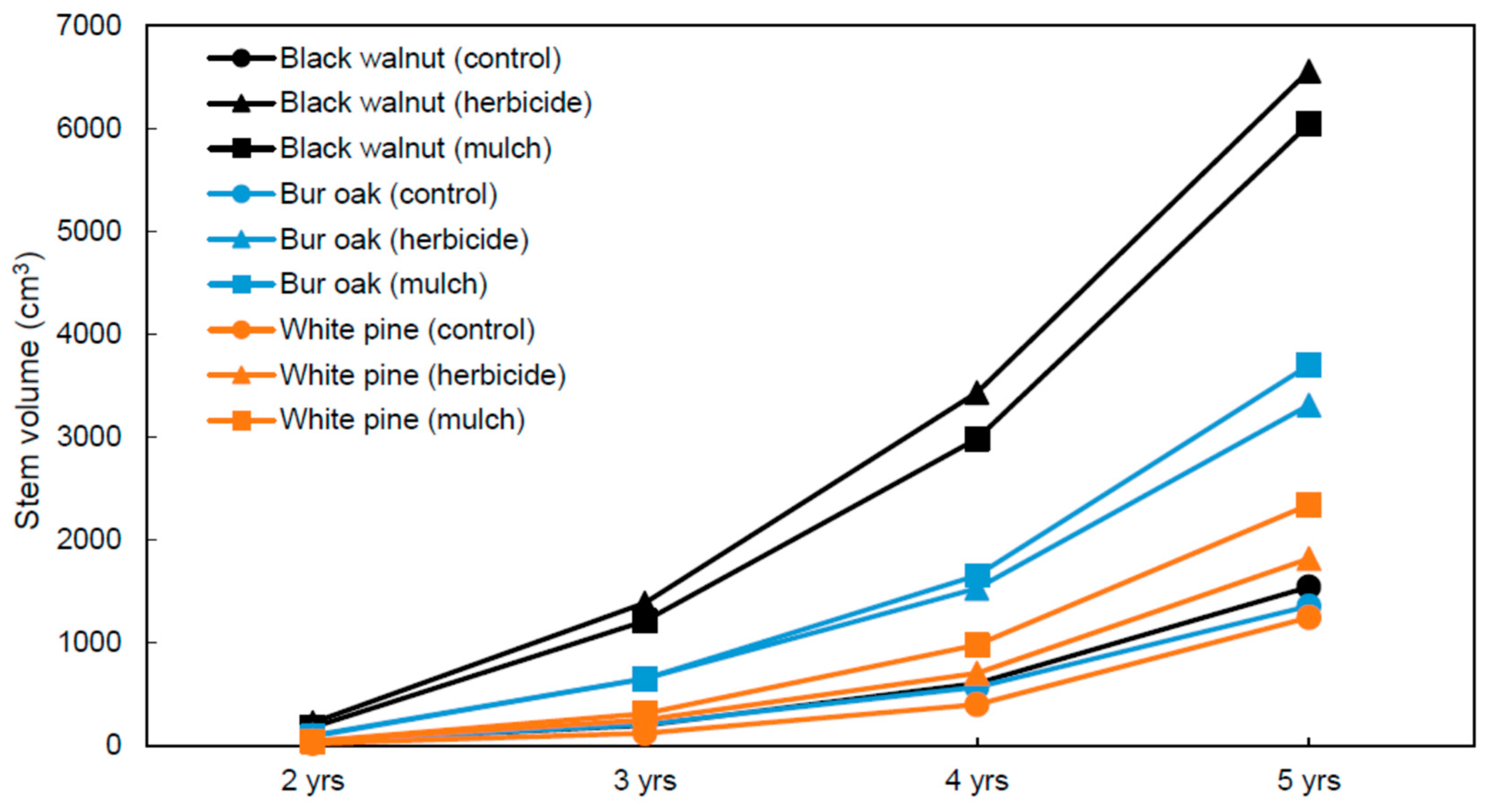
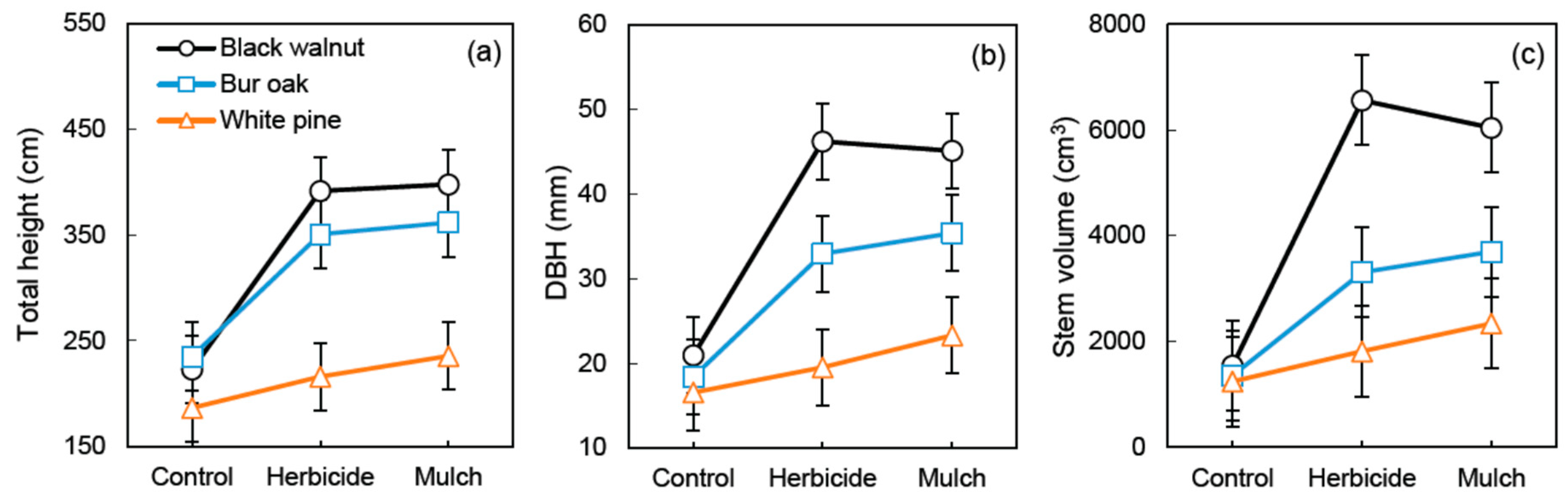
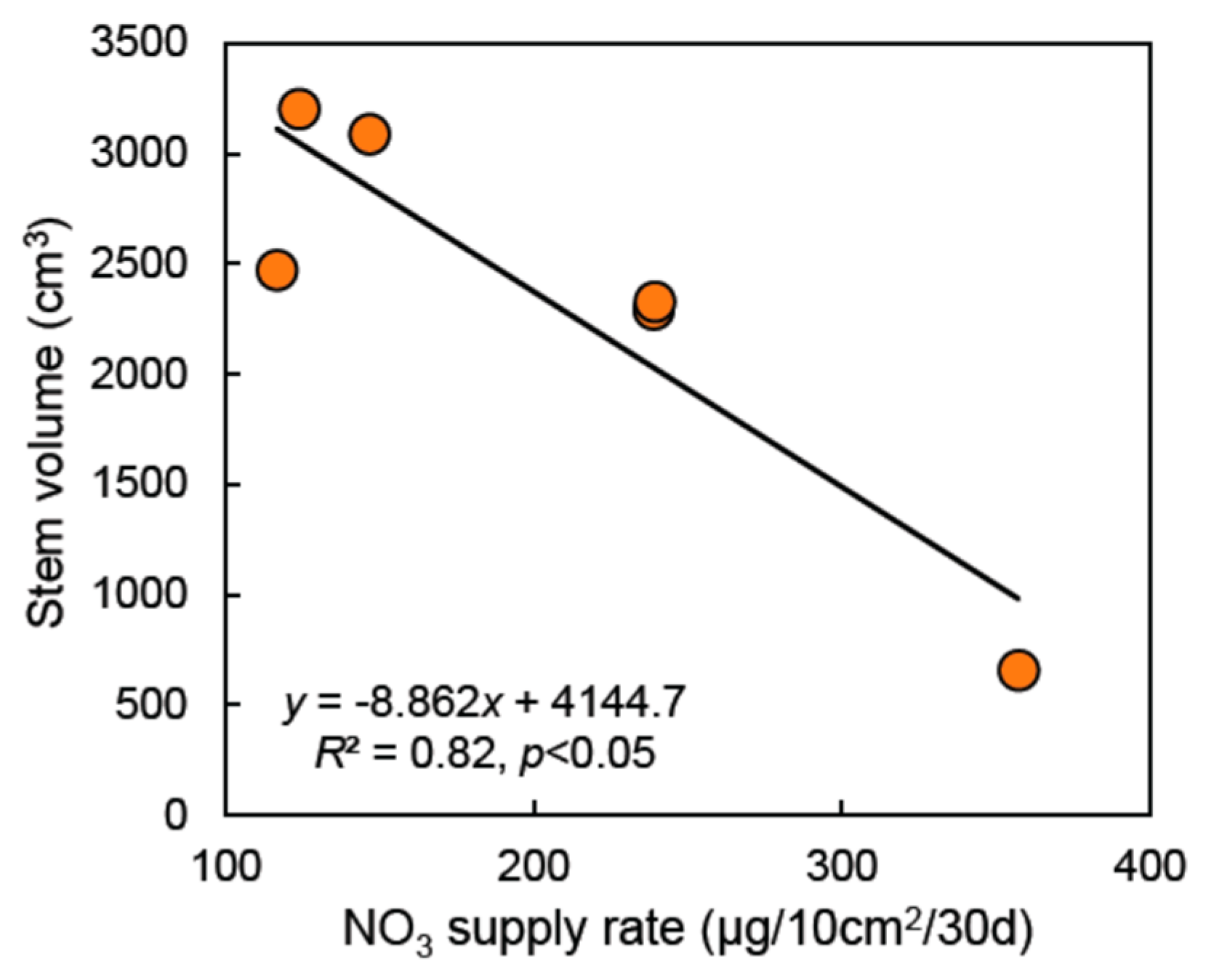
3. Results
3.1. Soil Nutrient Dynamics
3.2. Tree Growth and Survival
4. Discussion
4.1. Vegetation Treatments and Tree Species Affect Riparian Buffer Growth and Soil Nutrient Dynamics
4.2. Selecting a Vegetation Treatment for the Establishment of Trees in Riparian Buffers
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Muscutt, A.D.; Harris, G.L.; Bailey, S.W.; Davies, D.B. Buffer zones to improve water quality: A review of their potential use in UK agriculture. Agric. Ecosyst. Environ. 1993, 45, 59–77. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Blaine, J.G. River conservation, restoration, and preservation: Rewarding private behavior to enhance the commons. Freshw. Sci. 2016, 35, 755–763. [Google Scholar] [CrossRef]
- Dosskey, M.G.; Vidon, P.; Gurwick, N.P.; Allan, C.J.; Duval, T.P.; Lowrance, R. The role of riparian vegetation in protecting and improving chemical water quality in streams. JAWRA 2010, 46, 261–277. [Google Scholar]
- Mander, Ü.; Hayakawa, Y.; Kuusemets, V. Purification processes, ecological functions, planning and design of riparian buffer zones in agricultural watersheds. Ecol. Eng. 2005, 24, 421–432. [Google Scholar] [CrossRef]
- Tabacchi, E.; Lambs, L.; Guilloy, H.; Planty-Tabacchi, A.M.; Muller, E.; Décamps, H. Impacts of riparian vegetation on hydrological processes. Hydrol. Proc. 2000, 14, 2959–2976. [Google Scholar] [CrossRef]
- Boutin, C.; Jobin, B.; Bélanger, L. Importance of riparian habitats to flora conservation in farming landscapes of southern Québec, Canada. Agric. Ecosyst. Environ. 2003, 94, 73–87. [Google Scholar] [CrossRef]
- Jobin, B.; Bélanger, L.; Boutin, C.; Maisonneuve, C. Conservation value of agricultural riparian strips in the Boyer River watershed, Québec (Canada). Agric. Ecosyst. Environ. 2004, 103, 413–423. [Google Scholar] [CrossRef]
- Lovell, S.T.; Sullivan, W.C. Environmental benefits of conservation buffers in the United States: Evidence, promise, and open questions. Agric. Ecosyst. Environ. 2006, 112, 249–260. [Google Scholar] [CrossRef]
- Baxter, C.V.; Fausch, K.D.; Saunders, W.C. Tangled webs: Reciprocal flows of invertebrate prey link streams and riparian zones. Freshw. Biol. 2005, 50, 201–220. [Google Scholar] [CrossRef]
- Schultz, R.C.; Isenhart, T.M.; Simpkins, W.W.; Colletti, J.P. Riparian forest buffers in agroecosystems—Lessons learned from the Bear Creek Watershed, central Iowa, USA. Agrofor. Syst. 2004, 61–62, 35–50. [Google Scholar]
- Sweeney, B.W.; Czapka, S.J. Riparian forest restoration: Why each site needs an ecological prescription. For. Ecol. Manag. 2004, 192, 361–373. [Google Scholar] [CrossRef]
- Burns, R.M.; Honkala, B.H. Silvics of North America; Forest Service Agriculture: Washington, DC, USA, 1990. [Google Scholar]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Potential for hybrid poplar riparian buffers to provide ecosystem services in three watersheds with contrasting agricultural land use. Forests 2016, 7, 37. [Google Scholar] [CrossRef]
- Rheinhardt, R.; Brinson, M.; Meyer, G.; Miller, K. Carbon storage of headwater riparian zones in an agricultural landscape. Carbon Bal. Manag. 2012, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Truax, B.; Gagnon, D.; Lambert, F.; Fortier, J. Riparian buffer growth and soil nitrate supply are affected by tree species selection and black plastic mulching. Ecol. Eng. 2017, 106, 82–93. [Google Scholar] [CrossRef]
- Wendel, G.W.; Clay Smith, H. Eastern white pine. In Silvics of North America: 1. Conifers. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Andrews, D.M.; Barton, C.D.; Czapka, S.J.; Kolka, R.K.; Sweeney, B.W. Influence of tree shelters on seedling success in an afforested riparian zone. New For. 2010, 39, 157–167. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Czapka, S.J.; Yerkes, T. Riparian forest restoration: Increasing success by reducing plant competition and herbivory. Rest. Ecol. 2002, 10, 392–400. [Google Scholar] [CrossRef]
- Davies, R.J. The importance of weed control and the use of tree shelters for establishing broadleaved trees on grass-dominated sites in England. Forestry 1985, 58, 167–180. [Google Scholar] [CrossRef]
- Balandier, P.; Collet, C.; Miller, J.H.; Reynolds, P.E.; Zedaker, S.M. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 2006, 79, 3–27. [Google Scholar] [CrossRef]
- Fortier, J.; Gagnon, D.; Truax, B.; Lambert, F. Biomass and volume yield after 6 years in multiclonal hybrid poplar riparian buffer strips. Biomass Bioenergy 2010, 34, 1028–1040. [Google Scholar] [CrossRef]
- Borin, M.; Vianello, M.; Morari, F.; Zanin, G. Effectiveness of buffer strips in removing pollutants in runoff from a cultivated field in North-East Italy. Agric. Ecosyst. Environ. 2005, 105, 101–114. [Google Scholar] [CrossRef]
- Lambert, F.; Truax, B.; Gagnon, D.; Chevrier, N. Growth and N nutrition, monitored by enzyme assays, in a hardwood plantation: Effects of mulching materials and glyphosate application. For. Ecol. Manag. 1994, 70, 231–244. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D.; Chevrier, N. Nitrate reductase activity in relation to growth and soil N-forms in red oak and red ash planted in three different environments: Forest, clear-cut and field. For. Ecol. Manag. 1994, 64, 71–82. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D. Effects of straw and black plastic mulching on the initial growth and nutrition of butternut, white ash and bur oak. For. Ecol. Manag. 1993, 57, 17–27. [Google Scholar] [CrossRef]
- Munson, A.D.; Margolis, H.A.; Brand, D.G. Intensive silvicultural treatment: Impacts on soil fertility and planted conifer response. Soil Sci. Soc. Am. J. 1993, 57, 246–255. [Google Scholar] [CrossRef]
- Black, A.; Greb, B. Nitrate accumulation in soils covered with plastic mulch. Agron. J. 1962, 54, 366. [Google Scholar] [CrossRef]
- Kwabiah, A.B. Growth and yield of sweet corn (Zea mays L.) cultivars in response to planting date and plastic mulch in a short-season environment. Sci. Hortic. 2004, 102, 147–166. [Google Scholar] [CrossRef]
- Dreyer, E.; Le Roux, X.; Montpied, P.; Daudet, F.A.; Masson, F. Temperature response of leaf photosynthetic capacity in seedlings from seven temperate tree species. Tree Physiol. 2001, 21, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, W.R.; Naeem, H. Effect of weed control methods on growth of five temperate agroforestry tree species in Saskatchewan. For. Chron. 2017, 93, 271–281. [Google Scholar] [CrossRef]
- Smaill, S.J.; Ledgard, N.; Langer, E.R.; Henley, D. Establishing native plants in a weedy riparian environment. N. Z. J. Mar. Freshw. Res. 2011, 45, 357–367. [Google Scholar] [CrossRef]
- Von Althen, F.W. Effects of weed control on the survival and growth of planted black walnut, white ash and sugar maple. For. Chron. 1971, 47, 223–226. [Google Scholar] [CrossRef][Green Version]
- Bauer, G.A.; Berntson, G.M. Ammonium and nitrate acquisition by plants in response to elevated CO2 concentration: The roles of root physiology and architecture. Tree Physiol. 2001, 21, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Beckjord, P.R.; Adams, R.E.; Smith, D.W. Effects of nitrogen fertilization on growth and ectomycorrhizal formation of red oak. For. Sci. 1980, 26, 529–536. [Google Scholar]
- Gebauer, G.; Rehder, H.; Wollenweber, B. Nitrate, nitrate reduction and organic nitrogen in plants from different ecological and taxonomic groups of Central Europe. Oecologia 1988, 75, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Walters, M.B.; Willis, J.L.; Gottschalk, K.W. Seedling growth responses to light and mineral N form are predicted by species ecologies and can help explain tree diversity. Can. J. For. Res. 2014, 44, 1356–1368. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Caraco, N.F.; Correll, D.L.; Howarth, R.W.; Sharpley, A.N.; Smith, V.H. Nonpoint pollution of surface waters with phosphorus and nitrogen. Ecol. Appl. 1998, 8, 559–568. [Google Scholar] [CrossRef]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Biomass carbon, nitrogen and phosphorus stocks in hybrid poplar buffers, herbaceous buffers and natural woodlots in the riparian zone on agricultural land. J. Environ. Manag. 2015, 154, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Fortier, J.; Gagnon, D.; Truax, B.; Lambert, F. Understory plant diversity and biomass in hybrid poplar riparian buffer strips in pastures. New For. 2011, 42, 241–265. [Google Scholar] [CrossRef]
- Neilen, A.D.; Chen, C.R.; Parker, B.M.; Faggotter, S.J.; Burford, M.A. Differences in nitrate and phosphorus export between wooded and grassed riparian zones from farmland to receiving waterways under varying rainfall conditions. Sci. Total Environ. 2017, 598, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Kasirajan, S.; Ngouajio, M. Polyethylene and biodegradable mulches for agricultural applications: A review. Agron. Sustain. Dev. 2012, 32, 501–529. [Google Scholar] [CrossRef]
- Nicodemus, M.; Salifu, K.; Jacobs, D. Nitrate reductase activity and nitrogen compounds in xylem exudate of Juglans nigra seedlings: Relation to nitrogen source and supply. Tree Struct. Funct. 2008, 22, 685–695. [Google Scholar] [CrossRef]
- Robitaille, A.; Saucier, J.-P. Paysages Régionaux du Québec Méridional; Les Publications du Québec: Ste-Foy, QC, Canada, 1998. [Google Scholar]
- Westveld, M. Natural forest vegetation zones of New England. J. For. 1956, 54, 332–338. [Google Scholar]
- Government of Canada. Station Results—1981–2010 Climate Normals and Averages. Available online: http://climate.weather.gc.ca/climate_normals/station_select_1981_2010_e.html?searchType=stnProv&lstProvince=QC (accessed on 16 February 2017).
- Cann, D.B.; Lajoie, P. Études Des Sols Des Comtés de Stanstead, Richmond, Sherbrooke et Compton Dans la Province de Québec; Ministère de l’Agriculture: Ottawa, ON, Canada, 1943.
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Root biomass and soil carbon distribution in hybrid poplar riparian buffers, herbaceous riparian buffers and natural riparian woodlots on farmland. SpringerPlus 2013, 2. [Google Scholar] [CrossRef] [PubMed]
- Simavi, M.A. Effet de Plantations de Bandes Riveraines d’arbres sur l’abondance et la Répartition de la Faune Aquatique dans des Ruisseaux Dégradés de Milieux Agricoles dans Les Cantons-de-l’Est. M.Sc. Thesis, Université du Québec à Montréal, Montréal, QC, Canada, 2012. [Google Scholar]
- Lyons, J.; Thimble, S.W.; Paine, L.K. Grass versus trees: Managing riparian areas to benefit streams of central North America. JAWRA 2000, 36, 919–930. [Google Scholar] [CrossRef]
- Wallace, J.B.; Eggert, S.L.; Meyer, J.L.; Webster, J.R. Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science 1997, 277, 102–104. [Google Scholar] [CrossRef]
- Farrar, J.L. Les Arbres du Canada; Fides et le Service Canadien des Forêts, Ressources naturelles Canada: St-Laurent, QC, Canada, 2006. [Google Scholar]
- Williams, R.D. Black walnut. In Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Kabrick, J.M.; Dey, D.C.; Van Sambeek, J.W.; Coggeshall, M.V.; Jacobs, D.F. Quantifying flooding effects on hardwood seedling survival and growth for bottomland restoration. New For. 2012, 43, 695–710. [Google Scholar] [CrossRef]
- Johnson, P.S. Bur oak. In Silvics of North America: 2. Hardwoods. Agriculture Handbook 654; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990. [Google Scholar]
- Qian, P.; Schoenau, J.J.; Huang, W.Z. Use of ion exchange membranes in routine soil testing. Commun. Soil Sci. Plant Anal. 1992, 23, 1791–1804. [Google Scholar] [CrossRef]
- Fortier, J.; Truax, B.; Gagnon, D.; Lambert, F. Mature hybrid poplar riparian buffers along farm streams produce high yields in response to soil fertility assessed using three methods. Sustainability 2013, 5, 1893–1916. [Google Scholar] [CrossRef]
- West, P. Tree and Forest Measurement; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Perron, J.-Y. Inventaire forestier. In Manuel de Foresterie; Les Presses de l’Université Laval: Ste-Foy, QC, Canada, 1996; pp. 390–473. [Google Scholar]
- Petersen, R.G. Design and Analysis of Experiments; Marcel-Dekker: New York, NY, USA, 1985. [Google Scholar]
- Warton, D.I.; Hui, F.K.C. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Day, R.W.; Quinn, G.P. Comparisons of treatments after an analysis of variance in ecology. Ecol. Monogr. 1989, 59, 433–463. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Ellison, A.M. A Primer of Ecological Statistics; Sinauer Associated, Inc.: Sunderland, MA, USA, 2004. [Google Scholar]
- Bentrup, G. Conservation Buffers: Design Guidelines for Buffers, Corridors, and Greenways; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2008. [Google Scholar]
- Périé, C.; Munson, A.D. Ten-year responses of soil quality and conifer growth to silvicultural treatments. Soil Sci. Soc. Am. J. 2000, 64, 1815–1826. [Google Scholar] [CrossRef]
- Vitousek, P.M.; Porder, S.; Houlton, B.Z.; Chadwick, O.A. Terrestrial phosphorus limitation: Mechanisms, implications, and nitrogen-phosphorus interactions. Ecol. Appl. 2010, 20, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W. Sulfur cycling in forests. Biogeochemie 1984, 1, 29–43. [Google Scholar] [CrossRef]
- Sæbø, A.; Fløistad, I.S.; Netland, J.; Skúlason, B.; Edvardsen, Ø.M. Weed control measures in Christmas tree plantations of Abies nordmanniana and Abies lasiocarpa on agricultural land. New For. 2009, 38, 143–156. [Google Scholar] [CrossRef]
- Laungani, R.; Knops, J.M.H. Species-driven changes in nitrogen cycling can provide a mechanism for plant invasions. Proc. Natl. Acad. Sci. USA 2009, 106, 12400–12405. [Google Scholar] [CrossRef] [PubMed]
- Paris, P.; Cannata, F.; Olimpieri, G. Influence of alfalfa (Medicago sativa L.) intercropping and polyethylene mulching on early growth of walnut (Juglans spp.) in central Italy. Agrofor. Syst. 1995, 31, 169–180. [Google Scholar] [CrossRef]
- Von Althen, F.W. Revitalizing a black walnut plantation through weed control and fertilization. For. Chron. 1985, 61, 71–74. [Google Scholar] [CrossRef]
- Garrett, H.E.; Jones, J.E.; Kurtz, W.B.; Slusher, J.P. Black walnut (Juglans nigra L.) agroforestry—Its design and potential as a land-use alternative. For. Chron. 1991, 67, 213–218. [Google Scholar] [CrossRef]
- Von Althen, F.W. Afforestation of former farmland with high-value hardwoods. For. Chron. 1991, 67, 209–212. [Google Scholar] [CrossRef]
- Aber, J.D.; Magill, A.; Boone, R.; Melillo, J.M.; Steudler, P. Plant and soil responses to chronic nitrogen additions at the Harvard forest, Massachusetts. Ecol. Appl. 1993, 3, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Reich, P.B.; Schoettle, A.W. Role of phosphorus and nitrogen in photosynthetic and whole plant carbon gain and nutrient use efficiency in eastern white pine. Oecologia 1988, 77, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Steele, K.L.; Kabrick, J.M.; Dey, D.C.; Jensen, R.G. Restoring riparian forests in the Missouri Ozarks. North. J. Appl. For. 2013, 30, 109–117. [Google Scholar] [CrossRef]
- Truax, B.; Gagnon, D.; Lambert, F.; Fortier, J. Multiple-use zoning model for private forest owners in agricultural landscapes: A case study. Forests 2015, 6, 3614–3664. [Google Scholar] [CrossRef]
- Danner, B.T.; Knapp, A.K. Growth dynamics of oak seedlings (Quercus macrocarpa Michx. and Quercus muhlenbergii Engelm.) from gallery forests: Implications for forest expansion into grasslands. Trees 2001, 15, 271–277. [Google Scholar] [CrossRef]
- Beketov, M.A.; Kefford, B.J.; Schäfer, R.B.; Liess, M. Pesticides reduce regional biodiversity of stream invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef] [PubMed]
- Stone, W.W.; Gilliom, R.J.; Ryberg, K.R. Pesticides in U.S. streams and rivers: Occurrence and trends during 1992–2011. Environ. Sci. Technol. 2014, 48, 11025–11030. [Google Scholar] [CrossRef] [PubMed]
- Myers, J.P.; Antoniou, M.N.; Blumberg, B.; Carroll, L.; Colborn, T.; Everett, L.G.; Hansen, M.; Landrigan, P.J.; Lanphear, B.P.; Mesnage, R.; et al. Concerns over use of glyphosate-based herbicides and risks associated with exposures: A consensus statement. Environ. Health 2016, 15. [Google Scholar] [CrossRef] [PubMed]
- Fortier, J.; Messier, C. Are chemical or mechanical treatments more sustainable for forest vegetation management in the context of the TRIAD? For. Chron. 2006, 82, 806–818. [Google Scholar] [CrossRef]
- Steinmetz, Z.; Wollmann, C.; Schaefer, M.; Buchmann, C.; David, J.; Tröger, J.; Muñoz, K.; Frör, O.; Schaumann, G.E. Plastic mulching in agriculture. Trading short-term agronomic benefits for long-term soil degradation? Sci. Total Environ. 2016, 550, 690–705. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.J.; Liu, W.; Wang, E.; Zhou, T.; Xin, P. Residual plastic mulch fragments effects on soil physical properties and water flow behavior in the Minqin Oasis, Northwestern China. Soil Tillage Res. 2017, 166, 100–107. [Google Scholar] [CrossRef]
- Yan, C.; Liu, E.; Shu, F.; Liu, Q.; Liu, S.; He, W. Review of agricultural plastic mulching and its residual pollution and prevention measures in China. J. Agric. Res. Environ. 2014, 31, 95–102. [Google Scholar]
- Knight, K.W.; Schultz, R.C.; Mabry, C.M.; Isenhart, T.M. Ability of remnant riparian forests, with and without grass filters, to buffer concentrated surface runoff. JAWRA 2010, 46, 311–322. [Google Scholar]
- Hood, W.G.; Naiman, R.J. Vulnerability of riparian zones to invasion by exotic vascular plants. Plant Ecol. 2000, 148, 105–114. [Google Scholar] [CrossRef]
- Pysek, P.; Prach, K. Plant invasions and the role of riparian habitats: A comparison of four species alien to central Europe. J. Biogeogr. 1993, 20, 413–420. [Google Scholar] [CrossRef]
- Maillard, É.; Paré, D.; Munson, A.D. Soil carbon stocks and carbon stability in a twenty-year-old temperate plantation. Soil Sci. Soc. Am. J. 2010, 74, 1775–1785. [Google Scholar] [CrossRef]
- Rolando, A.C.; Baillie, R.B.; Thompson, G.D.; Little, M.K. The risks associated with glyphosate-based herbicide use in planted forests. Forests 2017, 8. [Google Scholar] [CrossRef]
- Environmental Protection Authority (EPA) of South Australia. Safe and Effective Herbicide Use—A Handbook for Near-Water Applications; EPA: Adelaide (SA), Australia, 2017.
- Sweeney, B.W.; Czapka, S.J.; Petrow, L.C.A. How planting method, weed abatement, and herbivory affect afforestation success. South J. Appl. Ecol. 2007, 31, 85–92. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Common Name | Habitat Range | Site Fertility Class | Early Growth | Shade Tolerance | N-Form Preference | Successional Status | Flood Tolerance |
|---|---|---|---|---|---|---|---|
| Black walnut 1 | Well-drained bottomlands | High | Moderatly fast | Low | NH4 | Early | Low |
| Bur oak 1 | Bottomlands, riparian zones, and dry calcareous sites | High | Slow to moderate | Low to intermediate | NO3:NH4 | Early | Intermediate |
| White pine 1 | All types of sites ranging from rocky hill top to peatland | Low to moderate | Slow to moderate | Intermediate | NH4 | Mid | Low |
| Contrast | Species | Treatment 2 | Black Walnut | Bur Oak | White Pine | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cont. | Herb. | Mulch | Cont. | Herb. | Mulch | Cont. | Herb. | Mulch | ||||
| (1) | NH4 pref. 1 vs. oak | Mulch | 0 | 0 | −1 | 0 | 0 | 2 | 0 | 0 | −1 | <0.01 |
| (2) | All | Mulch vs. Herb. | 0 | 1 | −1 | 0 | 1 | −1 | 0 | 1 | −1 | <0.01 |
| (3) | All | Herb. vs. Cont. | 1 | −1 | 0 | 1 | −1 | 0 | 1 | −1 | 0 | 0.37 |
| Treatment | NO3 | NH4 | P | Ca | K | Mg | S |
|---|---|---|---|---|---|---|---|
| Control | 66.9 | 3.23 | 9.13 | 2049 | 201.0 | 434.4 | 53.9 |
| Herbicide | 87.6 | 2.67 | 7.72 | 2161 | 149.5 | 380.6 | 52.0 |
| Plastic mulch | 165.8 | 2.58 | 12.69 | 2292 | 175.4 | 375.4 | 83.4 |
| SED 1 | 22.7 | 0.35 | 2.35 | 95 | 60.7 | 36.2 | 10.0 |
| p-value | <0.001 | 0.13 | 0.11 | <0.05 | 0.70 | 0.21 | <0.01 |
| Contrast | Species | Treatment 1 | Black Walnut | Bur Oak | White Pine | p-Values | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cont. | Herb. | Mulch | Cont. | Herb. | Mulch | Cont. | Herb. | Mulch | ||||
| (1) | B. walnut | Veg. vs. Cont. | 2 | −1 | −1 | 0 | 0 | 0 | 0 | 0 | 0 | <0.001 |
| (2) | B. walnut | Herb. vs. Mulch | 0 | −1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0.55 |
| (3) | B. oak | Veg. vs. Cont. | 0 | 0 | 0 | 2 | −1 | −1 | 0 | 0 | 0 | <0.01 |
| (4) | B. oak | Herb. vs. Mulch | 0 | 0 | 0 | 0 | −1 | 1 | 0 | 0 | 0 | 0.65 |
| (5) | W. pine | Veg. vs. Cont. | 0 | 0 | 0 | 0 | 0 | 0 | 2 | −1 | −1 | 0.25 |
| (6) | W. pine | Herb. vs. Mulch | 0 | 0 | 0 | 0 | 0 | 0 | 0 | −1 | 1 | 0.54 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truax, B.; Fortier, J.; Gagnon, D.; Lambert, F. Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers? Forests 2018, 9, 258. https://doi.org/10.3390/f9050258
Truax B, Fortier J, Gagnon D, Lambert F. Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers? Forests. 2018; 9(5):258. https://doi.org/10.3390/f9050258
Chicago/Turabian StyleTruax, Benoit, Julien Fortier, Daniel Gagnon, and France Lambert. 2018. "Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers?" Forests 9, no. 5: 258. https://doi.org/10.3390/f9050258
APA StyleTruax, B., Fortier, J., Gagnon, D., & Lambert, F. (2018). Black Plastic Mulch or Herbicide to Accelerate Bur Oak, Black Walnut, and White Pine Growth in Agricultural Riparian Buffers? Forests, 9(5), 258. https://doi.org/10.3390/f9050258