Improved Water Consumption Estimates of Black Locust Plantations in China’s Loess Plateau




Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Continuous Field Measurements
2.2.1. Weather Stations
2.2.2. Quantifying Total Evapotranspiration
2.2.3. Quantifying Understory Transpiration
2.2.4. Quantifying Overstory Transpiration
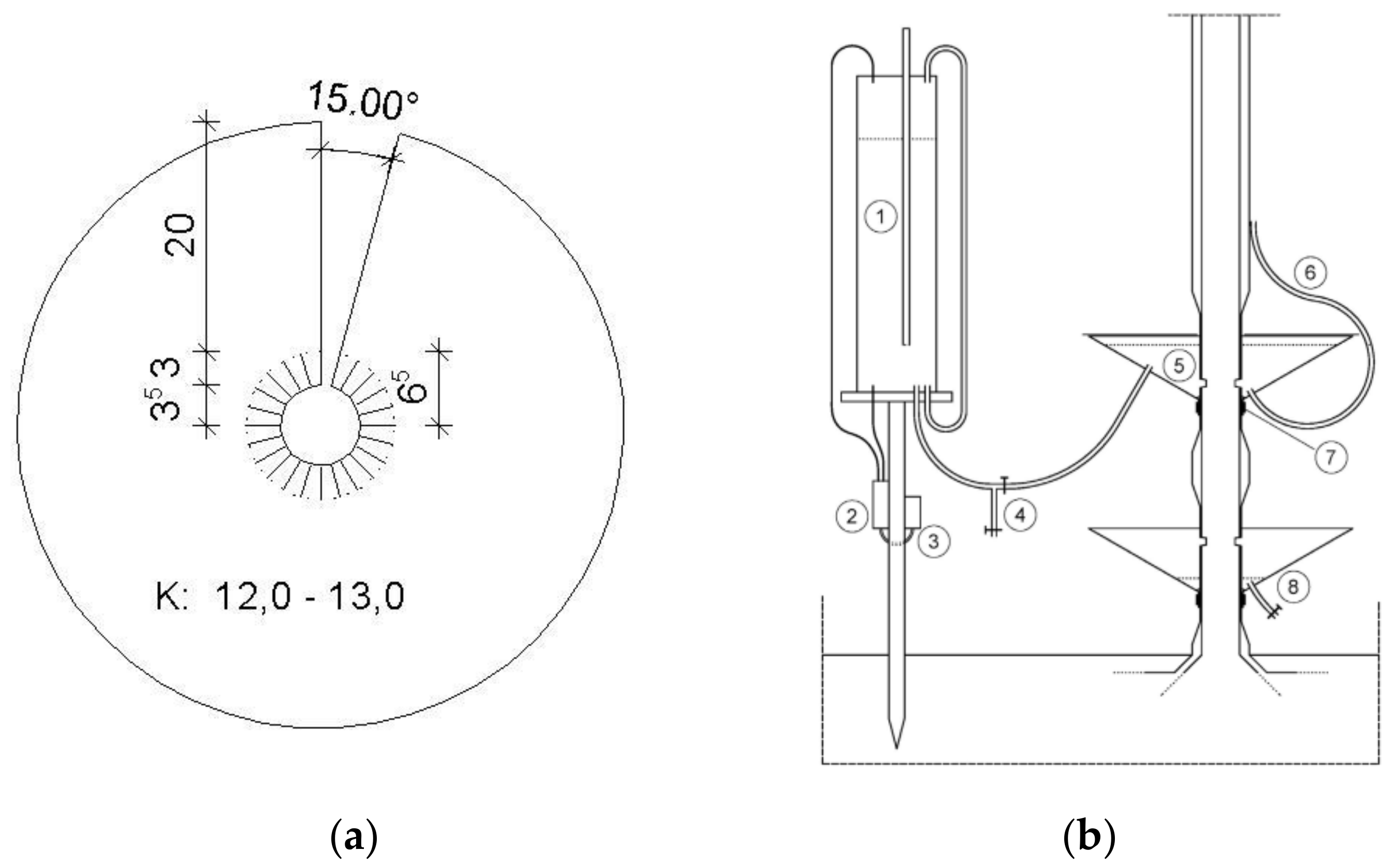
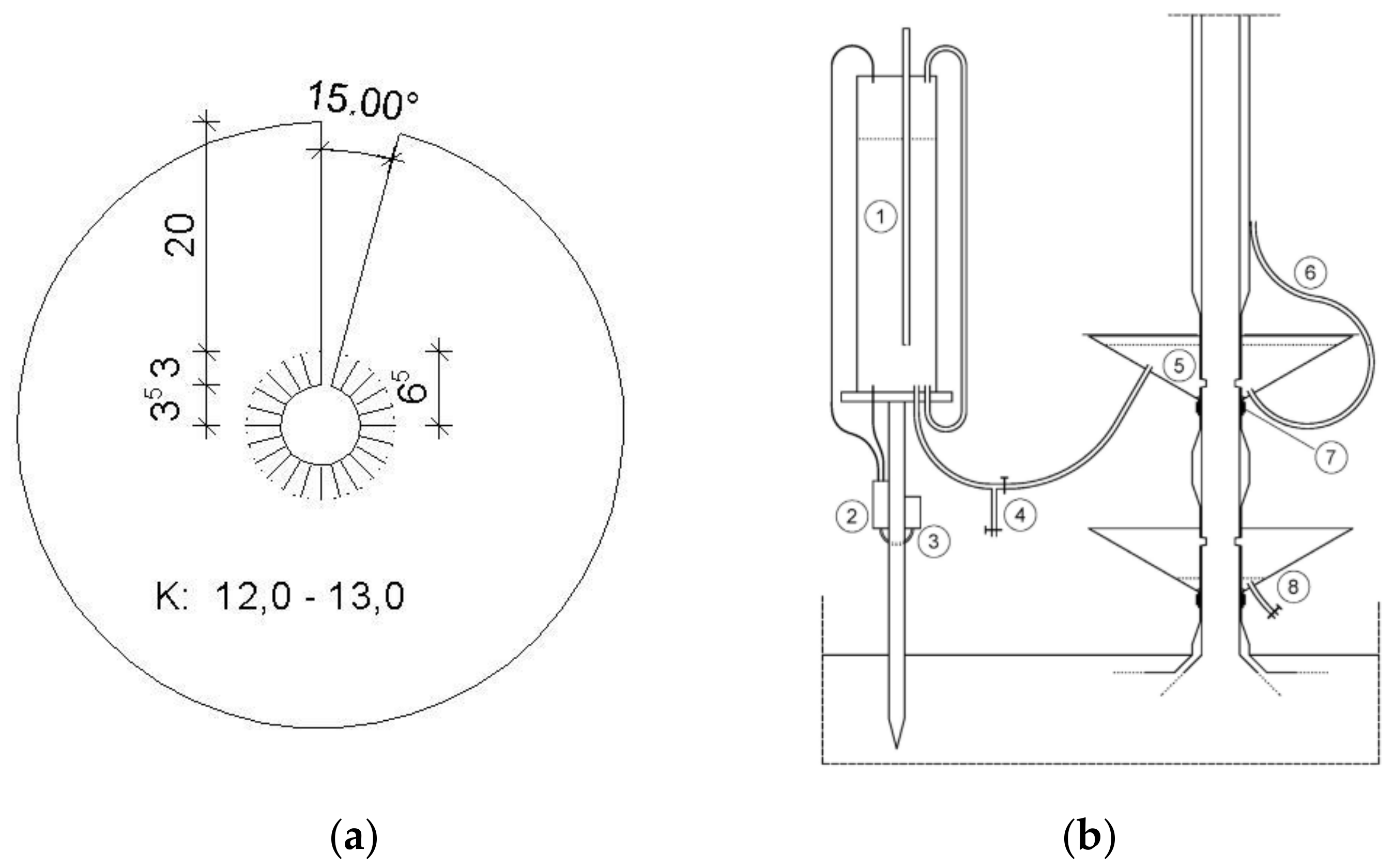
2.3. Calibration Experiment and Cut Tree Technique
3. Results and Discussion
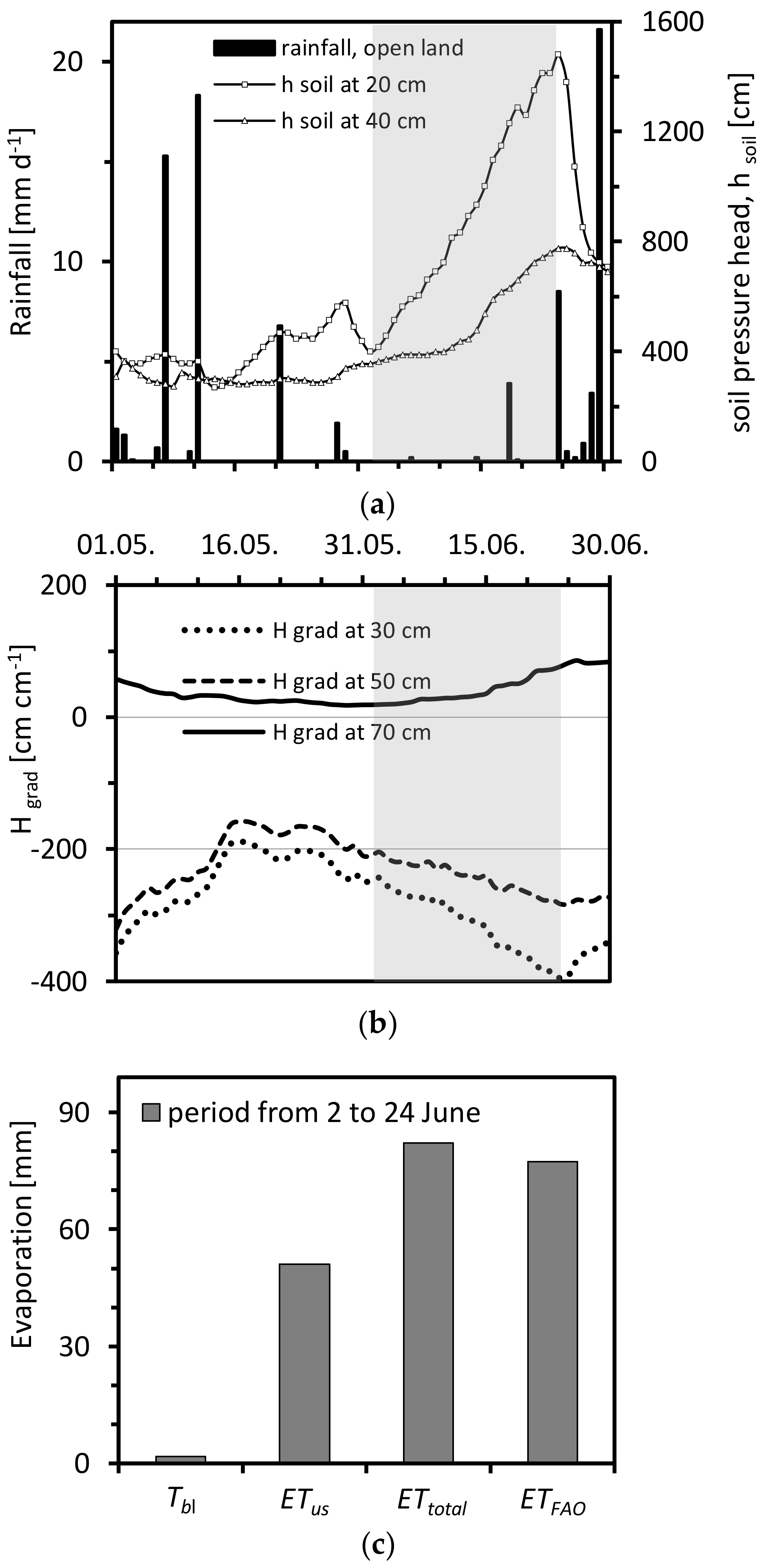
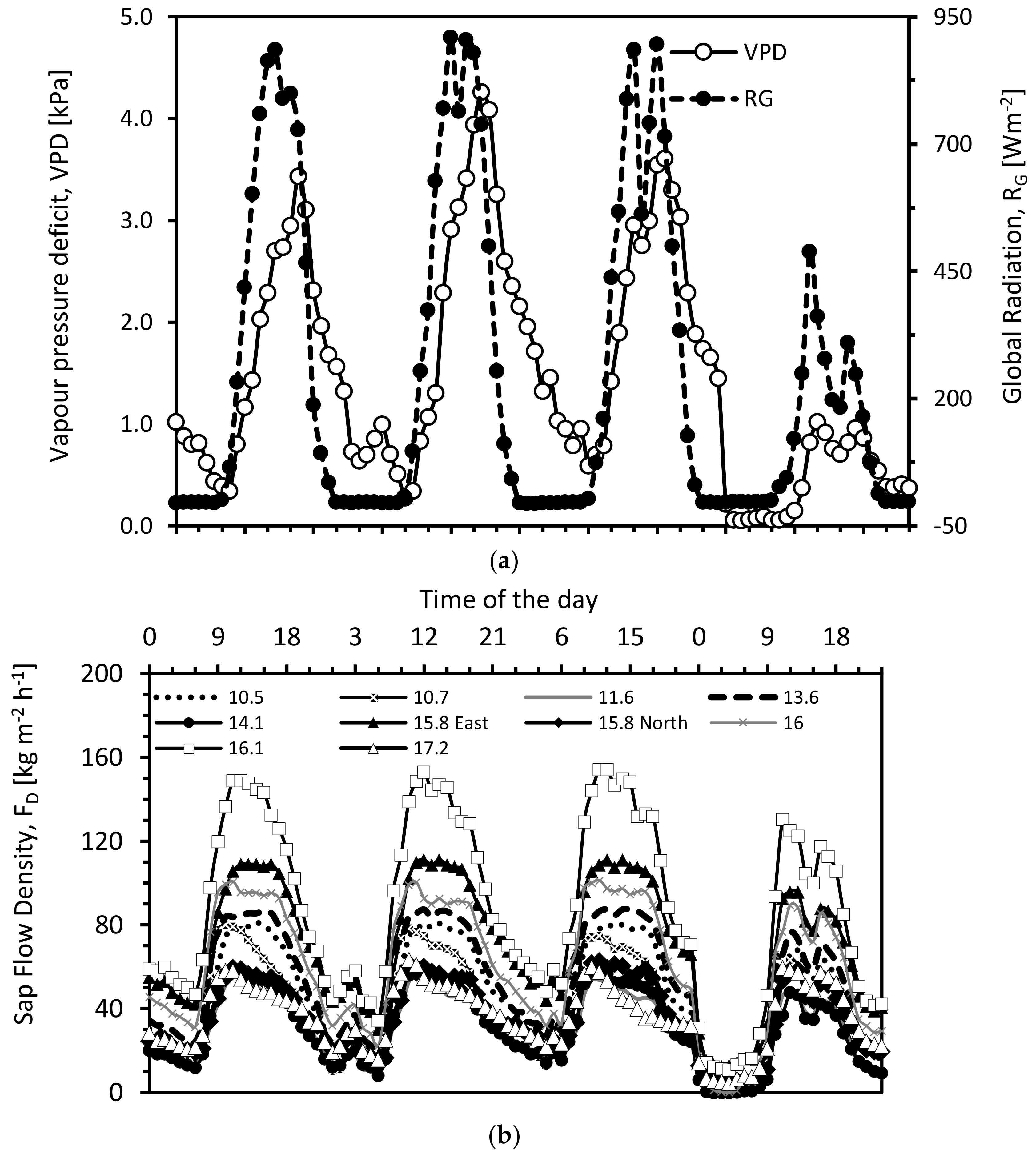
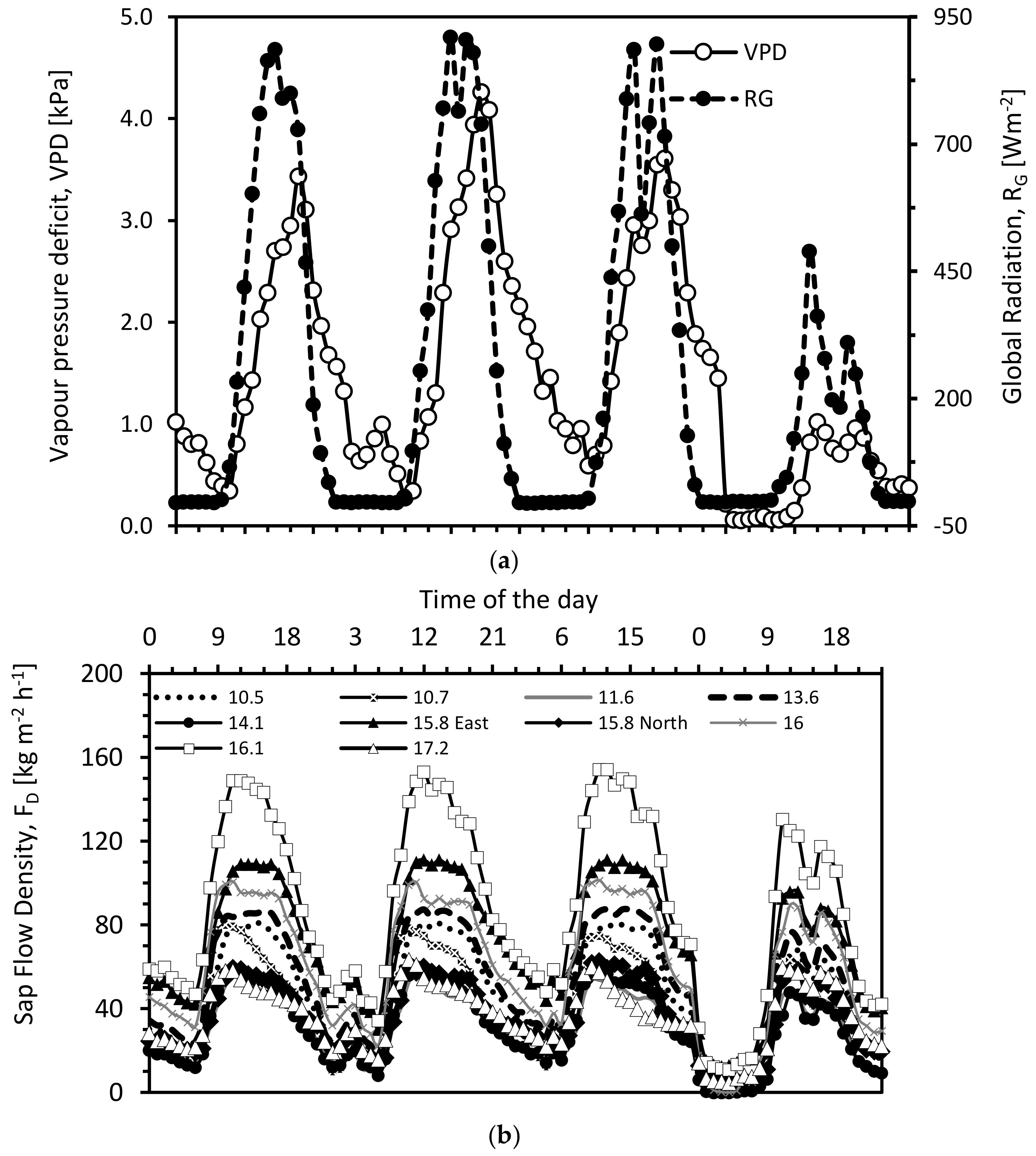
3.1. Diurnal Cycle of Transpiration Estimated Using Granier’s Original Calibration and Soil Water Balance Method
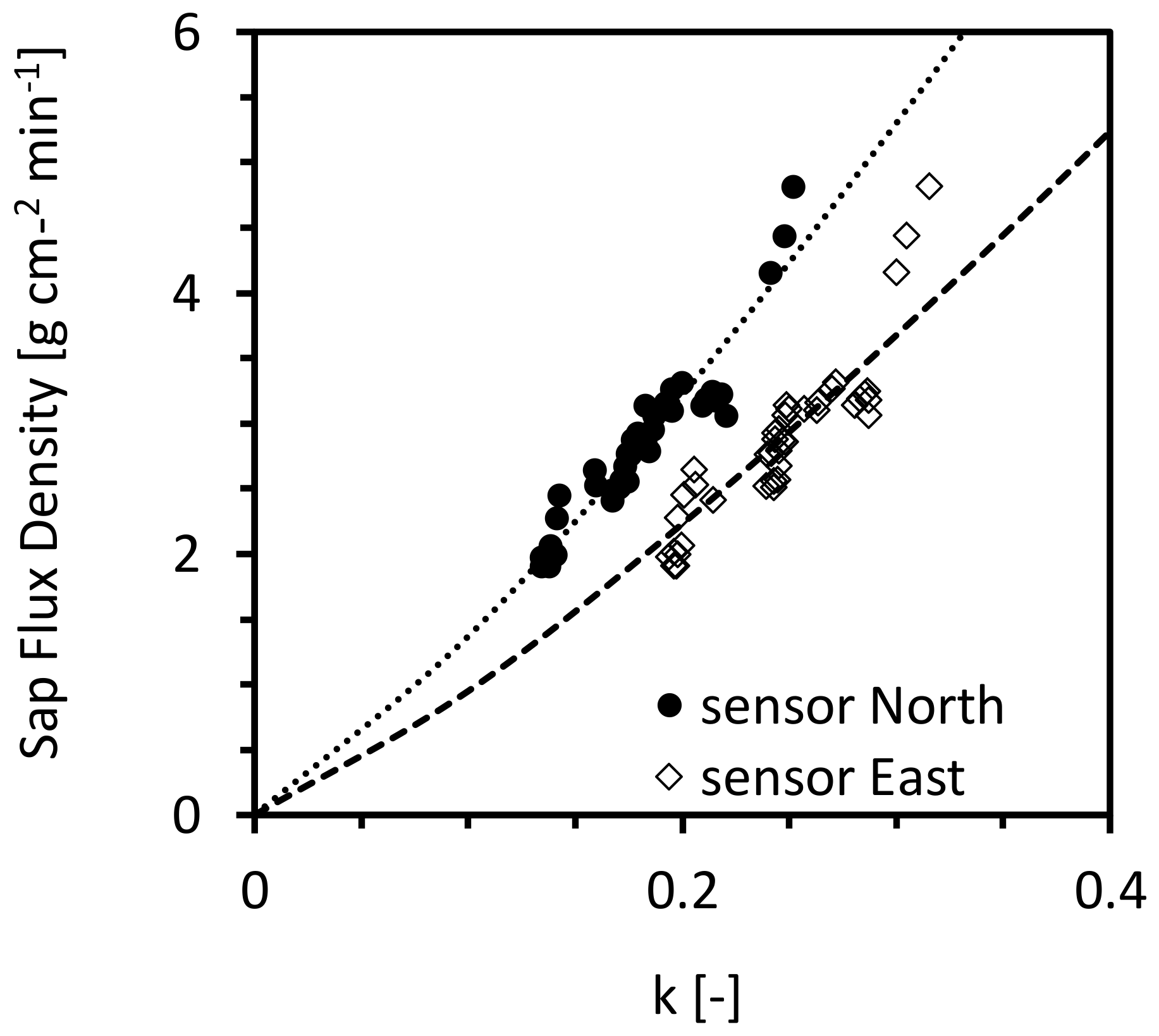
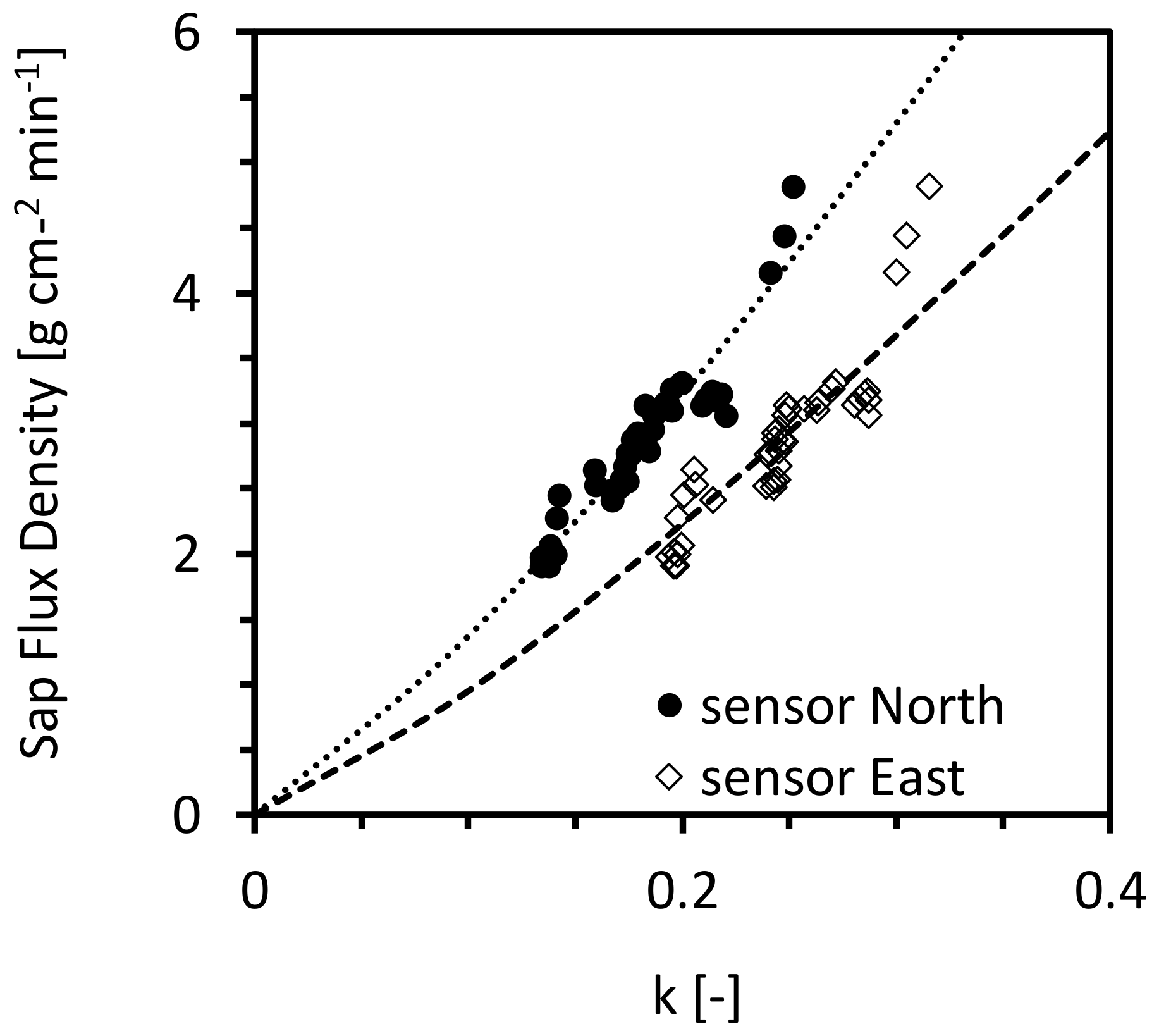
3.2. Tree-Specific Calibration of Granier’s Sensor System
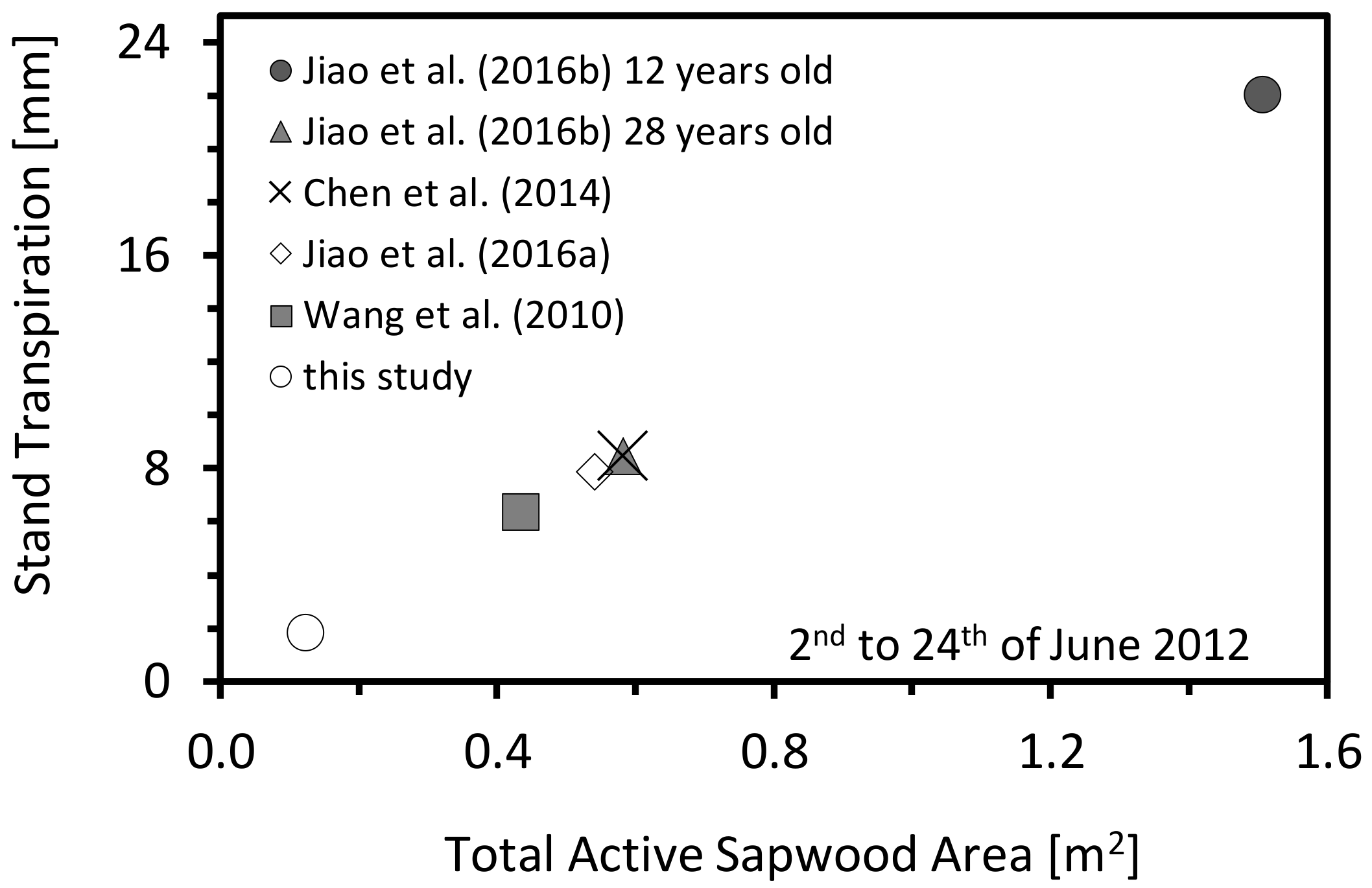
3.3. Validate the Accuracy of the New Calibration
4. Summary and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kim, H.K.; Park, J.; Hwang, I. Investigating water transport through the xylem network in vascular plants. J. Exp. Bot. 2014, 65, 1895–1904. [Google Scholar] [CrossRef] [PubMed]
- Vandegehuchte, M.W.; Steppe, K. Sap-flux density measurement methods: Working principles and applicability. Funct. Plant Biol. 2013, 40, 213–223. [Google Scholar] [CrossRef]
- Köstner, B.; Granier, A.; Cermák, J. Sapflow measurements in forest stands: Methods and uncertainties. In Annales des Sciences Forestières; EDP Sciences: Les Ulis, France, 1998; Volume 55, pp. 13–27. [Google Scholar]
- Regalado, C.M.; Ritter, A. An alternative method to estimate zero flow temperature differences for Granier’s thermal dissipation technique. Tree Physiol. 2007, 27, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Rabbel, I.; Diekkrüger, B.; Voigt, H.; Neuwirth, B. Comparing Tmax Determination Approaches for Granier-Based Sapflow Estimations. Sensors 2016, 16, 2042. [Google Scholar] [CrossRef] [PubMed]
- Oren, R.; Phillips, N.; Ewers, B.E.; Pataki, D.E.; Megonigal, J.P. Sap-flux-scaled transpiration responses to light, vapor pressure deficit, and leaf area reduction in a flooded Taxodium distichum forest. Tree Physiol. 1999, 19, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Clearwater, M.J.; Meinzer, F.C.; Andrade, J.L.; Goldstein, G.; Holbrook, N.M. Potential errors in measurement of nonuniform sap flow using heat dissipation probes. Tree Physiol. 1999, 19, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Paudel, I.; Kanety, T.; Cohen, S. Inactive xylem can explain differences in calibration factors for thermal dissipation probe sap flow measurements. Tree Physiol. 2013, 33, 986–1001. [Google Scholar] [CrossRef] [PubMed]
- Ford, C.R.; Hubbard, R.M.; Kloeppel, B.D.; Vose, J.M. A comparison of sap flux-based evapotranspiration estimates with catchment-scale water balance. Agric. For. Meteorol. 2007, 145, 176–185. [Google Scholar] [CrossRef]
- Wullschleger, S.D.; Childs, K.W.; King, A.W.; Hanson, P.J. A model of heat transfer in sapwood and implications for sap flux density measurements using thermal dissipation probes. Tree Physiol. 2011, 31, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Vergeynst, L.L.; Vandegehuchte, M.W.; McGuire, M.A.; Teskey, R.O.; Steppe, K. Changes in stem water content influence sap flux density measurements with thermal dissipation probes. Trees 2014, 28, 949–955. [Google Scholar] [CrossRef]
- Hölttä, T.; Linkosalo, T.; Riikonen, A.; Sevanto, S.; Nikinmaa, E. An analysis of Granier sap flow method, its sensitivity to heat storage and a new approach to improve its time dynamics. Agric. For. Meteorol. 2015, 211, 2–12. [Google Scholar] [CrossRef]
- Steppe, K.; De Pauw, D.J.; Doody, T.M.; Teskey, R.O. A comparison of sap flux density using thermal dissipation, heat pulse velocity and heat field deformation methods. Agric. For. Meteorol. 2010, 150, 1046–1056. [Google Scholar] [CrossRef]
- Bush, S.E.; Hultine, K.R.; Sperry, J.S.; Ehleringer, J.R. Calibration of thermal dissipation sap flow probes for ring-and diffuse-porous trees. Tree Physiol. 2010, 30, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Luo, Y.; Shao, M.; Li, X.; Sun, L.; Jia, X. Environmental controls on sap flow in black locust forest in Loess Plateau, China. Sci. Rep. 2017, 7, 13160. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Aubrey, D.P.; Teskey, R.O. A simple calibration improved the accuracy of the thermal dissipation technique for sap flow measurements in juvenile trees of six species. Trees 2012, 26, 631–640. [Google Scholar] [CrossRef]
- Domec, J.-C.; Sun, G.; Noormets, A.; Gavazzi, M.J.; Treasure, E.A.; Cohen, E.; Swenson, J.J.; McNulty, S.G.; King, J.S. A comparison of three methods to estimate evapotranspiration in two contrasting loblolly pine plantations: Age-related changes in water use and drought sensitivity of evapotranspiration components. For. Sci. 2012, 58, 497–512. [Google Scholar] [CrossRef]
- Schütt, P.; Weisgerber, H.; Schuck, H.J.; Lang, U.M.; Stimm, B.; Roloff, A. Enzyklopädie der Laubbäume: Die große Enzyklopädie; Nikol Verlag Barkhausenweg: Hamburg, Germany, 2006; ISBN 978-3-937872-39-1. [Google Scholar]
- Vítková, M.; Müllerová, J.; Sádlo, J.; Pergl, J.; Pyšek, P. Black locust (Robinia pseudoacacia) beloved and despised: A story of an invasive tree in Central Europe. For. Ecol. Manag. 2017, 384, 287–302. [Google Scholar] [CrossRef]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Hejda, M.; Arianoutsou, M.; Essl, F.; Jarošík, V.; Pergl, J.; Winter, M.; Anastasiu, P. Alien flora of Europe: Species diversity, temporal trends, geographical patterns and research needs. Preslia 2008, 80, 101–149. [Google Scholar]
- Cao, S.; Chen, L.; Shankman, D.; Wang, C.; Wang, X.; Zhang, H. Excessive reliance on afforestation in China’s arid and semi-arid regions: Lessons in ecological restoration. Earth-Sci. Rev. 2011, 104, 240–245. [Google Scholar] [CrossRef]
- Wang, J.J.; Hu, C.X.; Bai, J.; Gong, C.M. Carbon sequestration of mature black locust stands on the Loess Plateau, China. Plant Soil Environ. 2015, 61, 116–121. [Google Scholar]
- Zhang, J.-G.; Guan, J.-H.; Shi, W.-Y.; Yamanaka, N.; Du, S. Interannual variation in stand transpiration estimated by sap flow measurement in a semi-arid black locust plantation, Loess Plateau, China. Ecohydrology 2015, 8, 137–147. [Google Scholar] [CrossRef]
- Jiao, L.; Lu, N.; Sun, G.; Ward, E.J.; Fu, B. Biophysical controls on canopy transpiration in a black locust (Robinia pseudoacacia) plantation on the semi-arid Loess Plateau, China. Ecohydrology 2016, 9, 1068–1081. [Google Scholar] [CrossRef]
- Jiao, L.; Lu, N.; Fu, B.; Gao, G.; Wang, S.; Jin, T.; Zhang, L.; Liu, J.; Zhang, D. Comparison of transpiration between different aged black locust (Robinia pseudoacacia) trees on the semi-arid Loess Plateau, China. J. Arid Land 2016, 8, 604–617. [Google Scholar] [CrossRef]
- Granier, A. Une nouvelle méthode pour la mesure du flux de sève brute dans le tronc des arbres. In Annales des Sciences Forestières; EDP Sciences: Les Ulis, France, 1985; Volume 42, pp. 193–200. [Google Scholar]
- Renninger, H.J.; Schäfer, K.V.R. Comparison of tissue heat balance-and thermal dissipation-derived sap flow measurements in ring-porous oaks and a pine. Front. Plant Sci. 2012, 3, 103. [Google Scholar] [CrossRef] [PubMed]
- Jian, S.; Zhao, C.; Fang, S.; Yu, K. Effects of different vegetation restoration on soil water storage and water balance in the Chinese Loess Plateau. Agric. For. Meteorol. 2015, 206, 85–96. [Google Scholar] [CrossRef]
- Wang, Y.-L.; Liu, G.-B.; Kume, T.; Otsuki, K.; Yamanaka, N.; Du, S. Estimating water use of a black locust plantation by the thermal dissipation probe method in the semiarid region of Loess Plateau, China. J. For. Res. 2010, 15, 241–251. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, Z.; Zeppel, M.; Liu, C.; Guo, J.; Zhu, J.; Zhang, X.; Zhang, J.; Zha, T. Response of transpiration to rain pulses for two tree species in a semiarid plantation. Int. J. Biometeorol. 2014, 58, 1569–1581. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zhang, Y.; An, J.; Liu, Q.; Lang, Y. Sap flow of black locust in response to environmental factors in two soils developed from different parent materials in the lithoid mountainous area of North China. Trees 2018, 1–14. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, K.; Lin, Y.; Shi, W.; Song, Y.; He, X. Balancing green and grain trade. Nat. Geosci. 2015, 8, 739–741. [Google Scholar] [CrossRef]
- Zhang, L.; Podlasly, C.; Feger, K.-H.; Wang, Y.; Schwärzel, K. Different land management measures and climate change impacts on the runoff–A simple empirical method derived in a mesoscale catchment on the Loess Plateau. J. Arid Environ. 2015, 120, 42–50. [Google Scholar] [CrossRef]
- Sun, G.; Zhou, G.; Zhang, Z.; Wei, X.; McNulty, S.G.; Vose, J.M. Potential water yield reduction due to forestation across China. J. Hydrol. 2006, 328, 548–558. [Google Scholar] [CrossRef]
- Feng, X.; Fu, B.; Piao, S.; Wang, S.; Ciais, P.; Zeng, Z.; Lü, Y.; Zeng, Y.; Li, Y.; Jiang, X. Revegetation in China’s Loess Plateau is approaching sustainable water resource limits. Nat. Clim. Chang. 2016, 6, 1019–1022. [Google Scholar] [CrossRef]
- Zimmermann, M.H. Hydraulic architecture of some diffuse-porous trees. Can. J. Bot. 1978, 56, 2286–2295. [Google Scholar] [CrossRef]
- Vertessy, R.A.; Hatton, T.J.; Reece, P.; O’sullivan, S.K.; Benyon, R.G. Estimating stand water use of large mountain ash trees and validation of the sap flow measurement technique. Tree Physiol. 1997, 17, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Braun, P.; Schmid, J. Sap flow measurements in grapevines (Vitis vinifera L.) 2. Granier measurements. Plant Soil 1999, 215, 47–55. [Google Scholar] [CrossRef]
- Yu, M.; Zhang, L.; Xu, X.; Feger, K.-H.; Wang, Y.; Liu, W.; Schwärzel, K. Impact of land-use changes on soil hydraulic properties of Calcaric Regosols on the Loess Plateau, NW China. J. Plant Nutr. Soil Sci. 2015, 178, 486–498. [Google Scholar] [CrossRef]
- Du, S.; Wang, Y.-L.; Kume, T.; Zhang, J.-G.; Otsuki, K.; Yamanaka, N.; Liu, G.-B. Sapflow characteristics and climatic responses in three forest species in the semiarid Loess Plateau region of China. Agric. For. Meteorol. 2011, 151, 1–10. [Google Scholar] [CrossRef]
- Schwärzel, K.; Menzer, A.; Clausnitzer, F.; Spank, U.; Häntzschel, J.; Grünwald, T.; Köstner, B.; Bernhofer, C.; Feger, K.-H. Soil water content measurements deliver reliable estimates of water fluxes: A comparative study in a beech and a spruce stand in the Tharandt forest (Saxony, Germany). Agric. For. Meteorol. 2009, 149, 1994–2006. [Google Scholar] [CrossRef]
- Bogena, H.R.; Herbst, M.; Huisman, J.A.; Rosenbaum, U.; Weuthen, A.; Vereecken, H. Potential of wireless sensor networks for measuring soil water content variability. Vadose Zone J. 2010, 9, 1002–1013. [Google Scholar] [CrossRef]
- Qu, W.; Bogena, H.R.; Huisman, J.A.; Vereecken, H. Calibration of a novel low-cost soil water content sensor based on a ring oscillator. Vadose Zone J. 2013, 12. [Google Scholar] [CrossRef]
- Khalil, M.; Sakai, M.; Mizoguchi, M.; Miyazaki, T. Current and prospective applications of zero flux plane (ZFP) method. J. Jpn. Soc. Soil Phys. 2003, 95, 75–90. [Google Scholar]
- Podlasly, C.; Schwärzel, K. Development of a continuous closed pipe system for controlling soil temperature at the lower boundary of weighing field lysimeters. Soil Sci. Soc. Am. J. 2013, 77, 2157–2163. [Google Scholar] [CrossRef]
- Sano, Y.; Okamura, Y.; Utsumi, Y. Visualizing water-conduction pathways of living trees: Selection of dyes and tissue preparation methods. Tree Physiol. 2005, 25, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Clausnitzer, F.; Köstner, B.; Schwärzel, K.; Bernhofer, C. Relationships between canopy transpiration, atmospheric conditions and soil water availability—Analyses of long-term sap-flow measurements in an old Norway spruce forest at the Ore Mountains/Germany. Agric. For. Meteorol. 2011, 151, 1023–1034. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56. FAO Rome 1998, 300, D05109. [Google Scholar]
- Wang, L.; Wei, S.; Horton, R.; Shao, M. Effects of vegetation and slope aspect on water budget in the hill and gully region of the Loess Plateau of China. Catena 2011, 87, 90–100. [Google Scholar] [CrossRef]
- McCulloh, K.; Sperry, J.S.; Lachenbruch, B.; Meinzer, F.C.; Reich, P.B.; Voelker, S. Moving water well: Comparing hydraulic efficiency in twigs and trunks of coniferous, ring-porous, and diffuse-porous saplings from temperate and tropical forests. New Phytol. 2010, 186, 439–450. [Google Scholar] [CrossRef] [PubMed]
- Kume, T.; Otsuki, K.; Du, S.; Yamanaka, N.; Wang, Y.-L.; Liu, G.-B. Spatial variation in sap flow velocity in semiarid region trees: Its impact on stand-scale transpiration estimates. Hydrol. Process. 2012, 26, 1161–1168. [Google Scholar] [CrossRef]
- Umebayashi, T.; Utsumi, Y.; Koga, S.; Inoue, S.; Fujikawa, S.; Arakawa, K.; Matsumura, J.; Oda, K. Conducting pathways in north temperate deciduous broadleaved trees. IAWA J. 2008, 29, 247–263. [Google Scholar] [CrossRef]
- Zimmerman, M.H.; Brown, C.L. Trees: Structure and Function; Springer-Verlag: New York, NY, USA, 1971. [Google Scholar]
- Huber, B.; Schmidt, E. Weitere thermoelektrische Untersuchungen über den Transpirationsstrom der Bäume. Tharandter Forstl. Jahrb. 1936, 87, 369–412. [Google Scholar]
- Taneda, H.; Sperry, J.S. A case-study of water transport in co-occurring ring-versus diffuse-porous trees: Contrasts in water-status, conducting capacity, cavitation and vessel refilling. Tree Physiol. 2008, 28, 1641–1651. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira Reis, F.; Campostrini, E.; de Sousa, E.F.; e Silva, M.G. Sap flow in papaya plants: Laboratory calibrations and relationships with gas exchanges under field conditions. Sci. Hortic. 2006, 110, 254–259. [Google Scholar] [CrossRef]
- Hultine, K.R.; Nagler, P.L.; Morino, K.; Bush, S.E.; Burtch, K.G.; Dennison, P.E.; Glenn, E.P.; Ehleringer, J.R. Sap flux-scaled transpiration by tamarisk (Tamarix spp.) before, during and after episodic defoliation by the saltcedar leaf beetle (Diorhabda carinulata). Agric. For. Meteorol. 2010, 150, 1467–1475. [Google Scholar] [CrossRef]
- Jacobsen, A.L.; Pratt, R.B.; Tobin, M.F.; Hacke, U.G.; Ewers, F.W. A global analysis of xylem vessel length in woody plants. Am. J. Bot. 2012, 99, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.H.; Jeje, A.A. Vessel-length distribution in stems of some American woody plant. Can. J. Bot. 1981, 59, 1882–1892. [Google Scholar]
- Nagata, A.; Kose, K.; Terada, Y. Development of an outdoor MRI system for measuring flow in a living tree. J. Magn. Reson. 2016, 265, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Urban, L.; Zhao, P. Granier’s thermal dissipation probe (TDP) method for measuring sap flow in trees: Theory and practice. Acta Bot. Sin. 2004, 46, 631–646. [Google Scholar]
- Granier, A.; Anfodillo, T.; Sabatti, M.; Cochard, H.; Dreyer, E.; Tomasi, M.; Valentini, R.; Bréda, N. Axial and radial water flow in the trunks of oak trees: A quantitative and qualitative analysis. Tree Physiol. 1994, 14, 1383–1396. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y. The hydrological influence of black locust plantations in the loess area of northwest China. Hydrol. Process. 1992, 6, 241–251. [Google Scholar]
- Wu, Y.Z.; Huang, M.B.; Warrington, D.N. Black locust transpiration responses to soil water availability as affected by meteorological factors and soil texture. Pedosphere 2015, 25, 57–71. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Area | Understory | MAT [°C] | MAP [mm year−1] | LAImax | Mean DBH [cm] | Number of Trees [ha−1] | AS/AG [m2ha−1] | Sapwood Thickness [mm] | TBL [mm d−1] | Relationship between DBH and AS | Source |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mt. Gonglushan, Yan’an 36°25′24″ N 109°31′ 32″ E | Grass and a few scattered shrubs | 10.6 | 498 | 2.89 | 9.3 | 3100 | 5.10 | 5–10 (mean 7.3) | 0.41 | AS = 0.546DBH1.508 | [29] |
| Mt. Gonglushan, Yan’an 36°25ʹ40″ N 109°31′ 53″ E | Small trees and shrubs | 10.6 | 498 | 2.89 | 9.3 | N.S. | N.S. | 7.2–9.2 | N.S. | N.S. | [40] |
| Caijiachuan catchment, Ji county 36°14′ 27″ to 36°18′23″ N 110°39′45″ to 110°47′45″ E | N.S. | 10.0 | 579 | N.S. | 6.9 | 850 | 1.57 | N.S. | <0.2 | AS = 3.4DBH−8.7 | [30] |
| Mt. Gonglushan, Yan’an 36°25′24″ N 109°31′32″ E | Grass and a few scattered shrubs | 10.6 | 498 | 2.73–3.14 | 9.3 | 3100 | 5.09 | 5–10 | 0.32 to 0.49 | AS = 0.546DBH1.508 | [23] |
| Yangjuangou, Yan’an 36°42′ N 109°31′ E | Patches of liana and herbs | 9.8 | 531 | 2.32–2.98 | 9.9–10.8 | 1300 | 3.16 to 3.60 | N.S. | 0.14 to 0.23 | AS = 0.61DBH1.55 | [24] |
| Yangjuangou, Yan’an 36°42′ N 109°31′ E | N.S. | 9.8 | 531 | 2.77 (12 years old) 2.38 (28 years old) | 6.94 (12 years old) 8.93 (28 years old) | 2500 (12 years old) 1200 (28 years old) | 3.16 to 3.60 | 15.4 (12 years old) 8.4 (28 years old) | 0.22 (12 years old) 0.39 (28 years old) | AS = 0.28DBH2.25 (12 years old) AS = 0.25DBH1.81 (28 years old) | [25] |
| Yeheshan forest reserve, Fufeng county 34°31′46″ N 107°54′40″ E | Grass | 12.7 | 580 | 2.4–.8 | 6.0−11 | 2450 (15 years old) | 5.13 to 5.30 | 10 ± 2 | 0.12−0.16 * | AS = 0.4024DBH1.9 | [15] |
| Zhonggou, 35°20′ N 107°31′ E | Grass and a few scattered shrubs | 10.2 | 509 | 2.8 | 10.7 | 1006 (33 years old) | 0.61 | 3.43 | 0.08 * | AS = 0.0604DBH1.882 | This study |
| Tree Species | Characteristics of the Tree and Stem Segments | a [kg m−2 s−1] | b [–] | Calibration Set-Up | Source |
|---|---|---|---|---|---|
| Elaeagnus Angustifolia (Russian Olive) | Diameter 4.36 cm, water-conducting sapwood area 1.70 cm2 | 9.30 | 1.65 | Lab experiments on cut stem segments: water was pulled through stem by applying varying pressure gradients, whereby the pressure was increased in several smaller steps ranging from 0.001 to 0.14 MPa m−1 | [14] |
| Gleditsia Triacanthos (Honey Locust) | Diameter 5.06 cm, water-conducting sapwood area 0.73 cm2 | 30.7 | 1.40 | ||
| Qercus Gambelii (Gambel Oak) | Diameter 4.37 cm, water-conducting sapwood area 0.35 cm2 | 58.1 | 1.88 | ||
| Sophora Japonica (Japanese Pagoda) | Diameter of stem segments 4.47 cm, water-conducting sapwood area 0.51 cm2 | 11.90 | 1.24 | ||
| Quercus Pedunuculata (English Oak) | Diameter of stem segments between 4 and 5 cm | 0.119 | 1.231 | Lab experiment on cut stem segments: water was forced through stem segments under pressure, flow rate was varied by adjusting the water pressure | [26] |
| Quercus Petraea (Sessile Oak) | Diameter of the stem 13.7 cm | 0.119 | 1.231 | Cut tree experiment: water flow through the tree due to transpiration | [62] |
| Quercus Alba (White Oak) | Juvenile trees, diameter between 6.5 and 9 cm | 0.128 | 1.47 | Potometer experiments: juvenile trees were cut and placed in containers, water flow through the cut stems due to transpiration | [16] |
| Ulmus Americana (American Elm) | Juvenile trees, diameter between 6.5 and 9 cm | 0.272 | 2.57 | ||
| Qercus Gambelii (Gambel Oak) | 4–7 years old, diameter of the stem segment was between 5 and 6 cm | 9.99 ± 6.18 (ranging from 2.38 to 18.1) | 1.24 ± 0.19 (ranging from 1.05 to 1.50) | Lab experiments on cut stem segments: water was pulled through stem by applying varying pressure gradients, whereby the pressure increased in several smaller steps | [55] |
| Robinia Pseudoacacia (Black Locust) | 15 years old stand, diameter 6–10 cm, 97–102 cm long, sapwood area 12.6–31.5 cm2 | 0.051 | 1.18 | Lab experiments on cut stem segments: water was pulled through stem by applying a series of pressure 0.005–0.04 MPa, whereby each pressure was achieved by varying the height of the reservoir and held for 30 min | [15] |
| Robinia Pseudoacacia (Black Locust) | 33 years old stand, diameter 14.3 cm, 13 m long, active sapwood area 7.16 cm2 | 3.29 (ranging from 2.70 to 3.89) | 1.231 | Cut tree experiment, water flow through the tree due to transpiration | this study |
| Period | Days | Rainfall (Open Land) | ETFAO | ETtotal (Soil Water Balance) | ETus (Lysimeter) | TBL (Sap Flow) | Tresidual (ETtotal − ETus) | Difference between the Two Transpiration Estimates (TBL − Tresidual) |
|---|---|---|---|---|---|---|---|---|
| [mm] | ||||||||
| 2 June 2012–24 June 2012 | 23 | 4.4 | 77.3 | 82.2 | 51.0 | 27.7 (1.2 *) | 31.2 | 11% |
| 29 May 2013–7 June 2013 | 10 | 2.2 | 39.0 | 34.4 | 24.7 | 13.9 (1.4) | 9.7 | 30% |
| 12 June 2013–18 June 2013 | 8 | 0.7 | 25.2 | 23.2 | 17.4 | 7.0 (1.1) | 5.8 | 26% |
| 23 June 2013–30 June 2013 | 8 | 1.1 | 39.0 | 33.2 | 23.5 | 10.4 (1.3) | 9.7 | 7% |
| 28 July 2013–6 August 2013 | 10 | 0.3 | 46.2 | 39.7 | 30.6 | 12.1 (1.2) | 9.1 | 33% |
| 12 August 2013–23 August 2013 | 12 | 0.0 | 51.7 | 41.8 | 29.5 | 14.0 (1.2) | 12.3 | 14% |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwärzel, K.; Zhang, L.; Strecker, A.; Podlasly, C. Improved Water Consumption Estimates of Black Locust Plantations in China’s Loess Plateau. Forests 2018, 9, 201. https://doi.org/10.3390/f9040201
Schwärzel K, Zhang L, Strecker A, Podlasly C. Improved Water Consumption Estimates of Black Locust Plantations in China’s Loess Plateau. Forests. 2018; 9(4):201. https://doi.org/10.3390/f9040201
Chicago/Turabian StyleSchwärzel, Kai, Lulu Zhang, Andreas Strecker, and Christian Podlasly. 2018. "Improved Water Consumption Estimates of Black Locust Plantations in China’s Loess Plateau" Forests 9, no. 4: 201. https://doi.org/10.3390/f9040201
APA StyleSchwärzel, K., Zhang, L., Strecker, A., & Podlasly, C. (2018). Improved Water Consumption Estimates of Black Locust Plantations in China’s Loess Plateau. Forests, 9(4), 201. https://doi.org/10.3390/f9040201