Do Common Silvicultural Treatments Affect Wood Density of Mediterranean Montane Pines?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of Trials and Data Collection
2.2. Measurements with X-ray Microdensitometry
2.3. Statistical Analysis
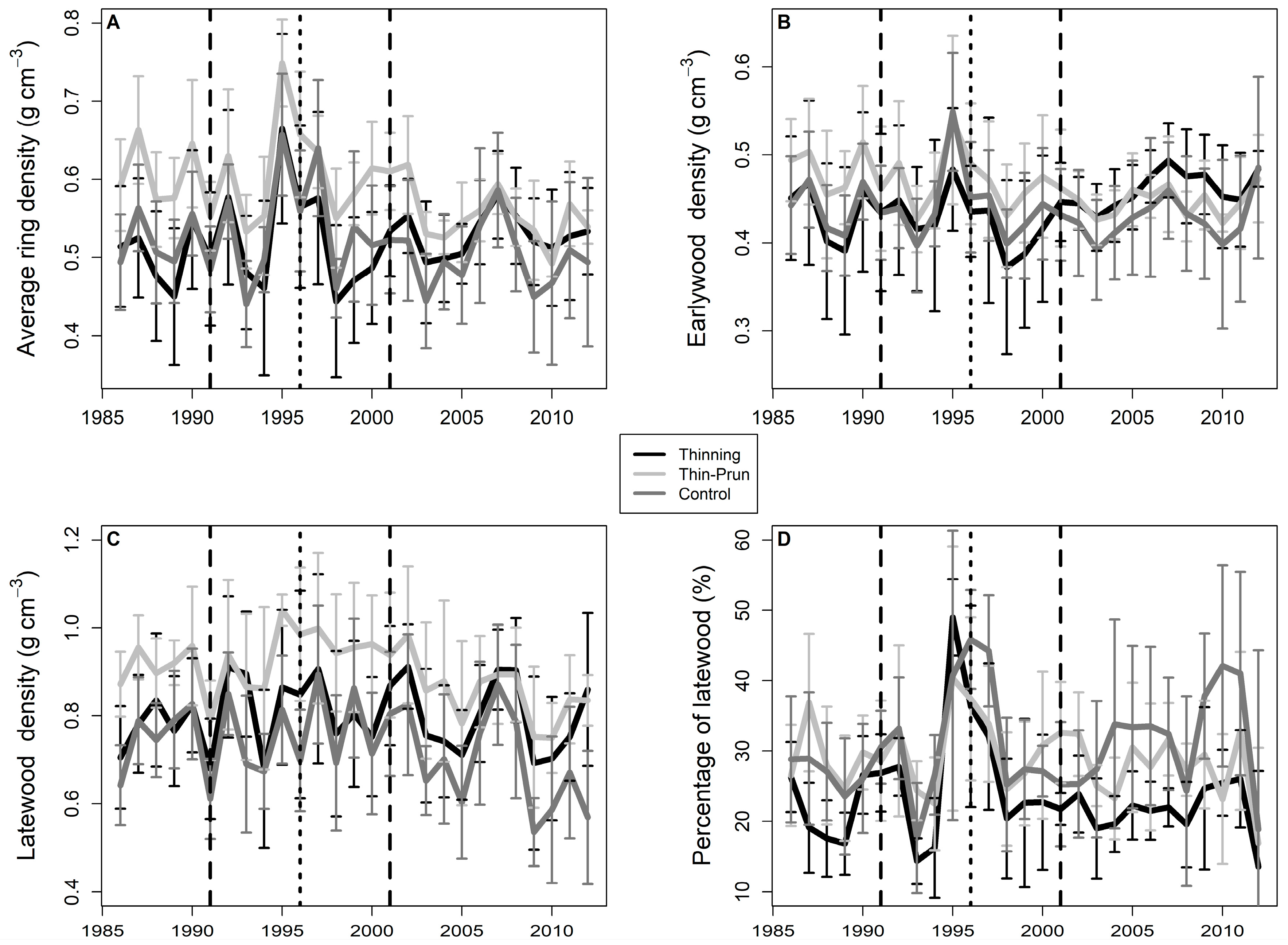
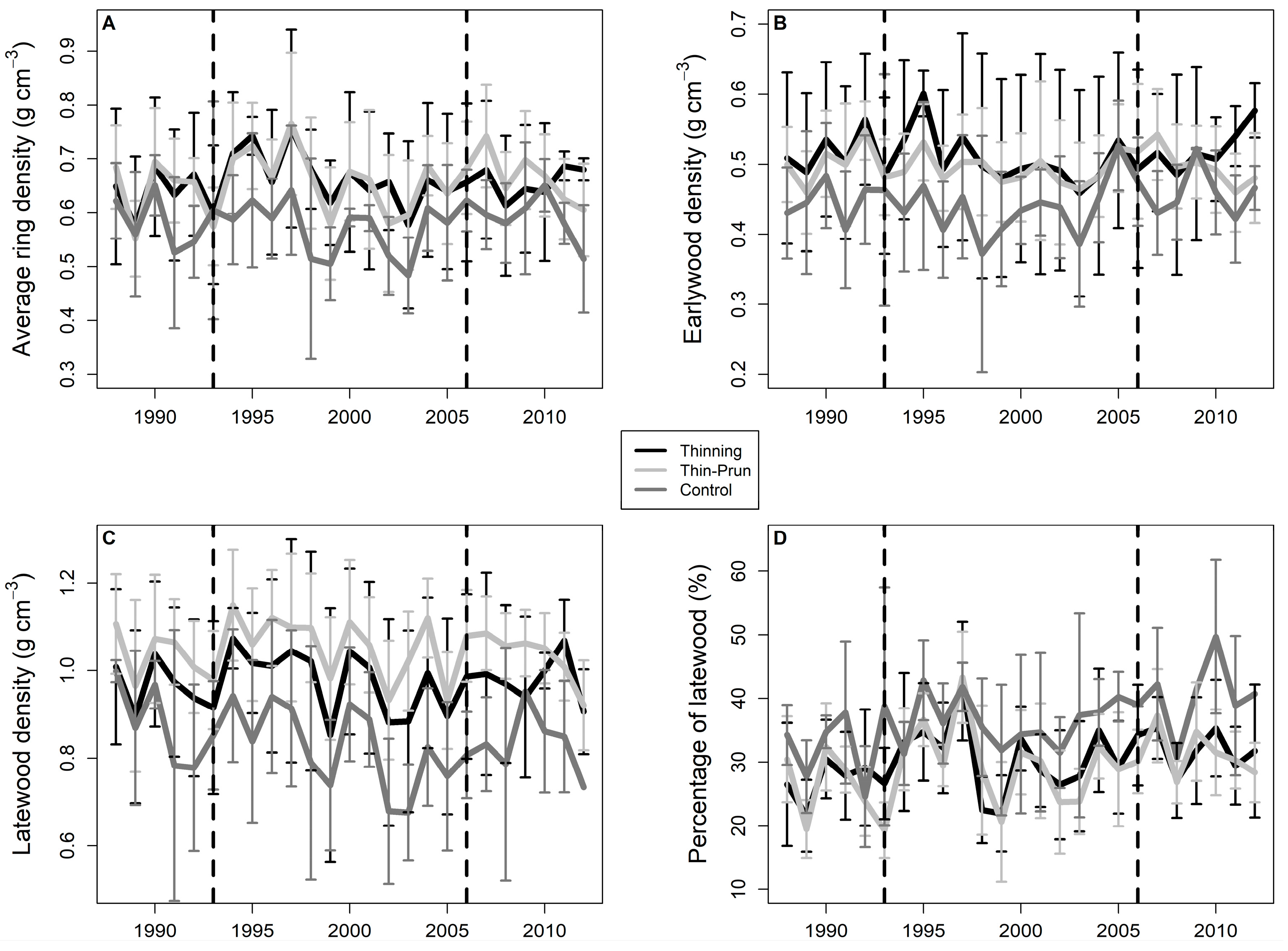
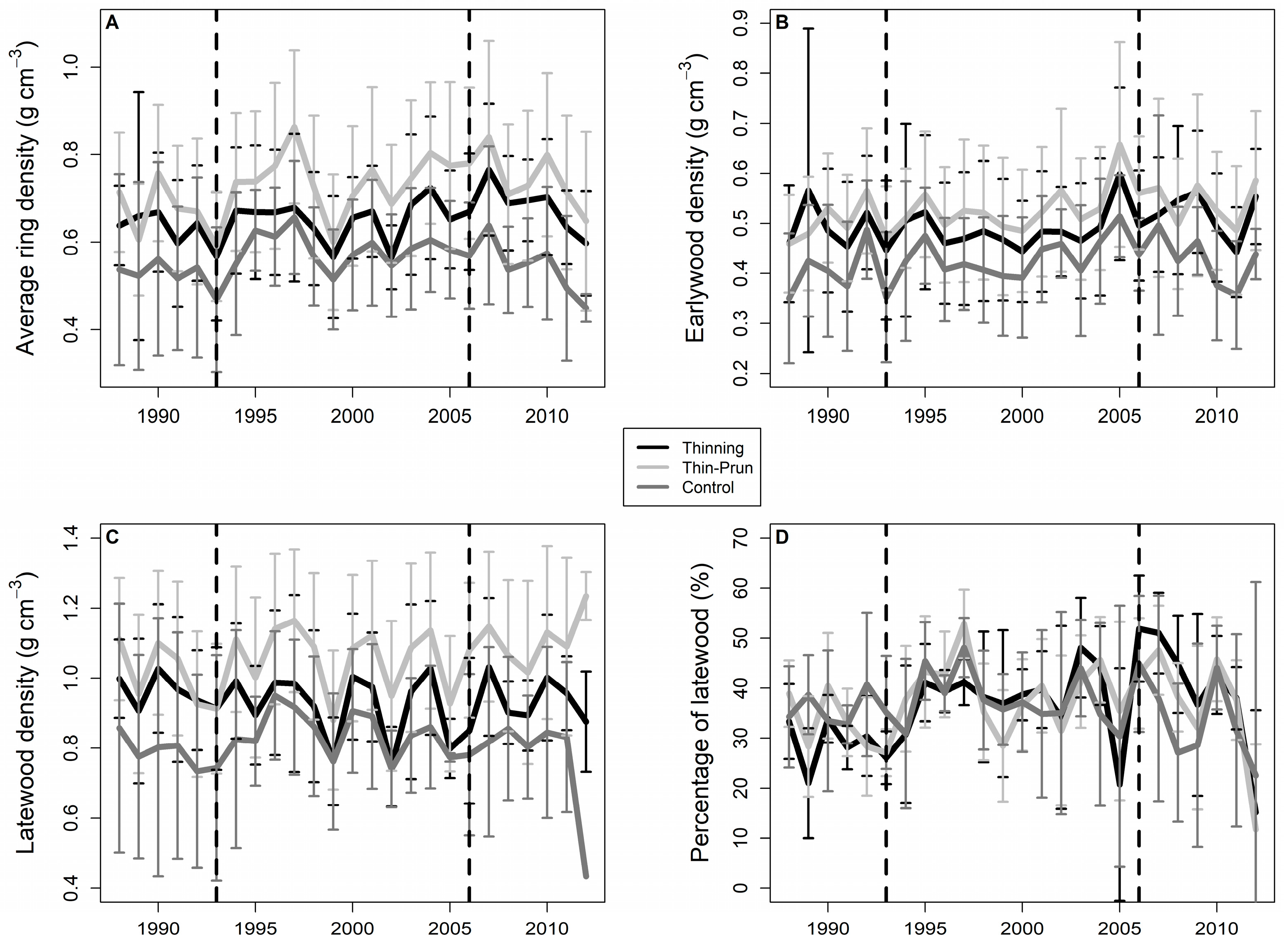
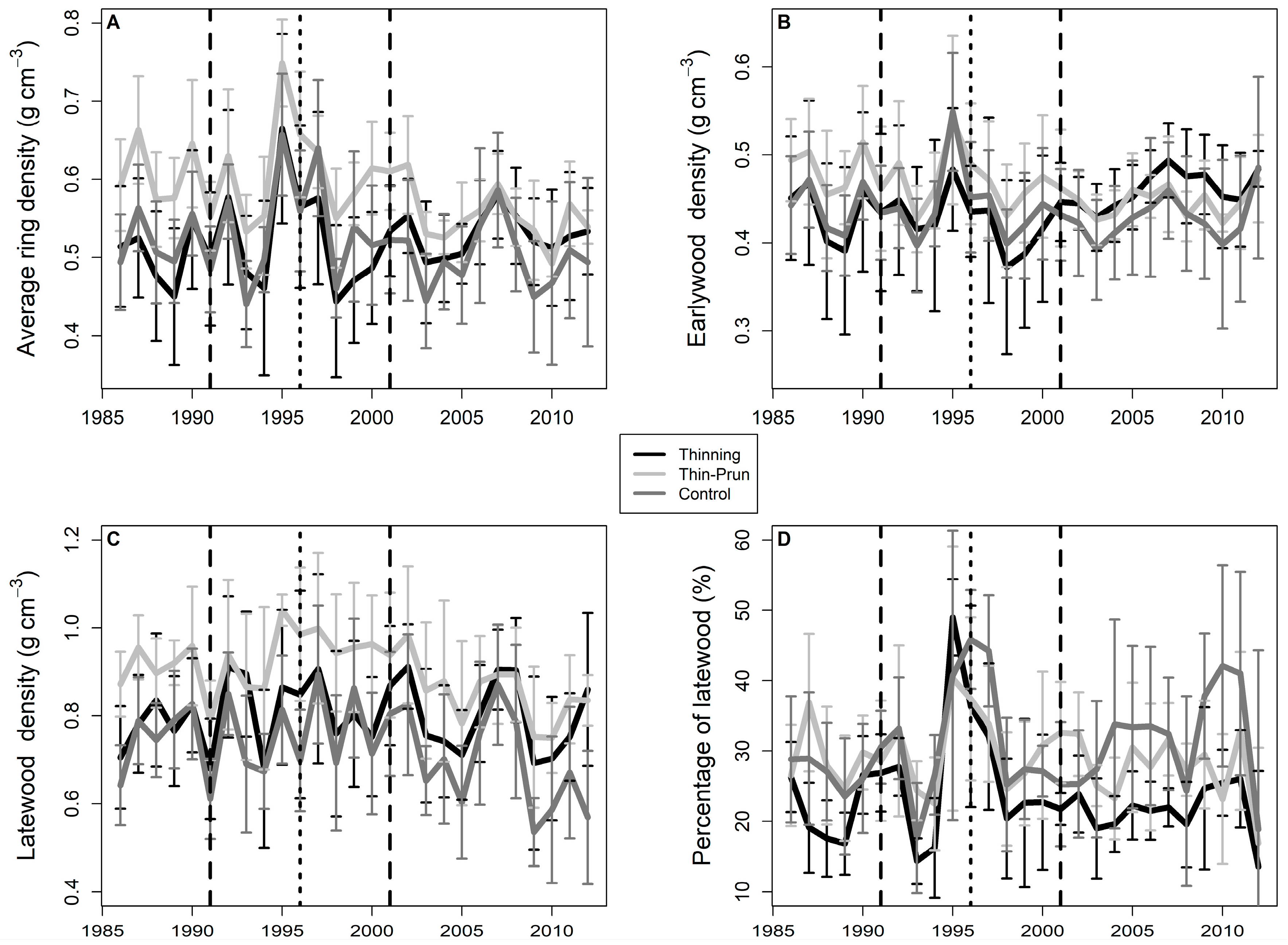
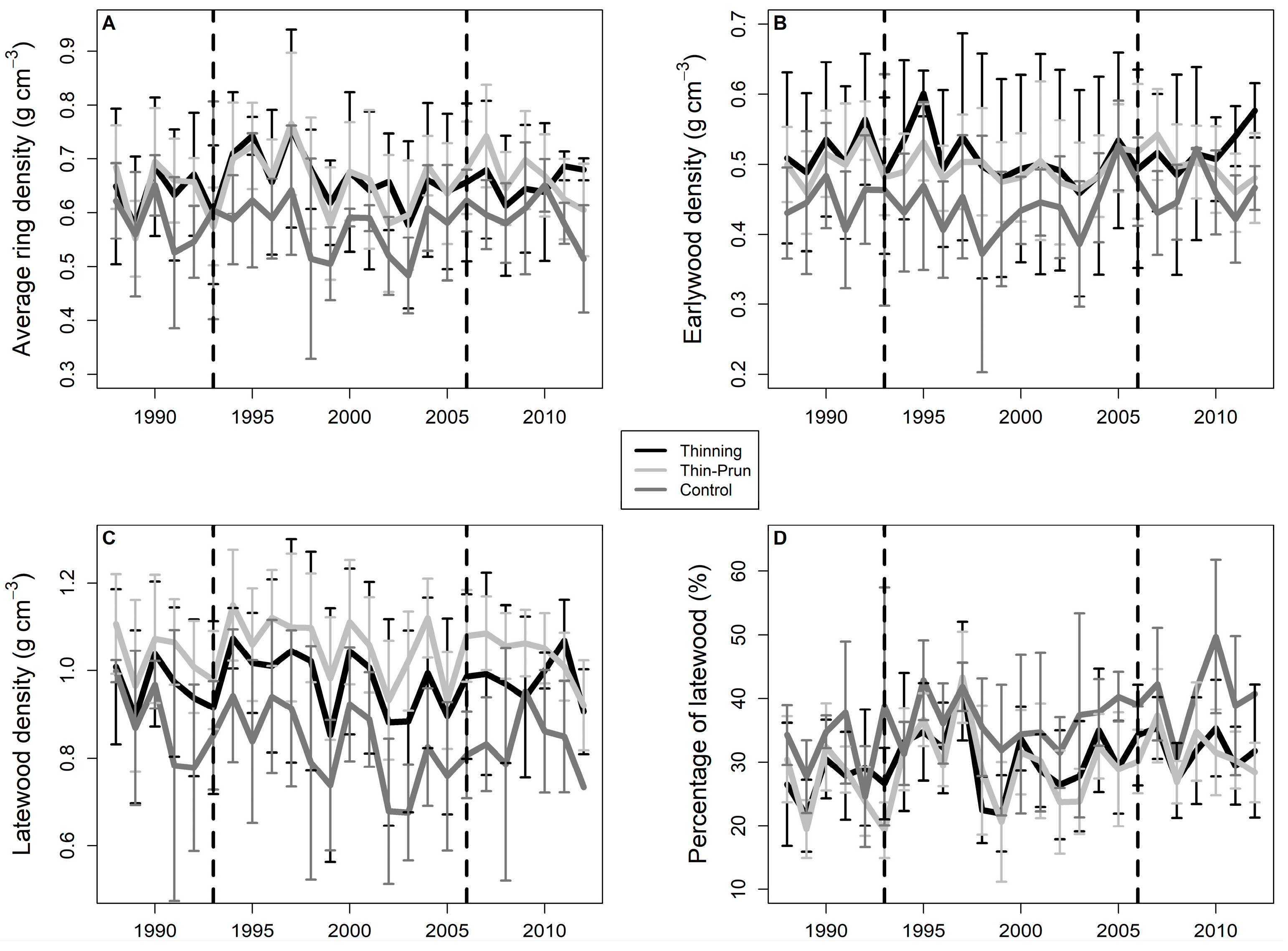
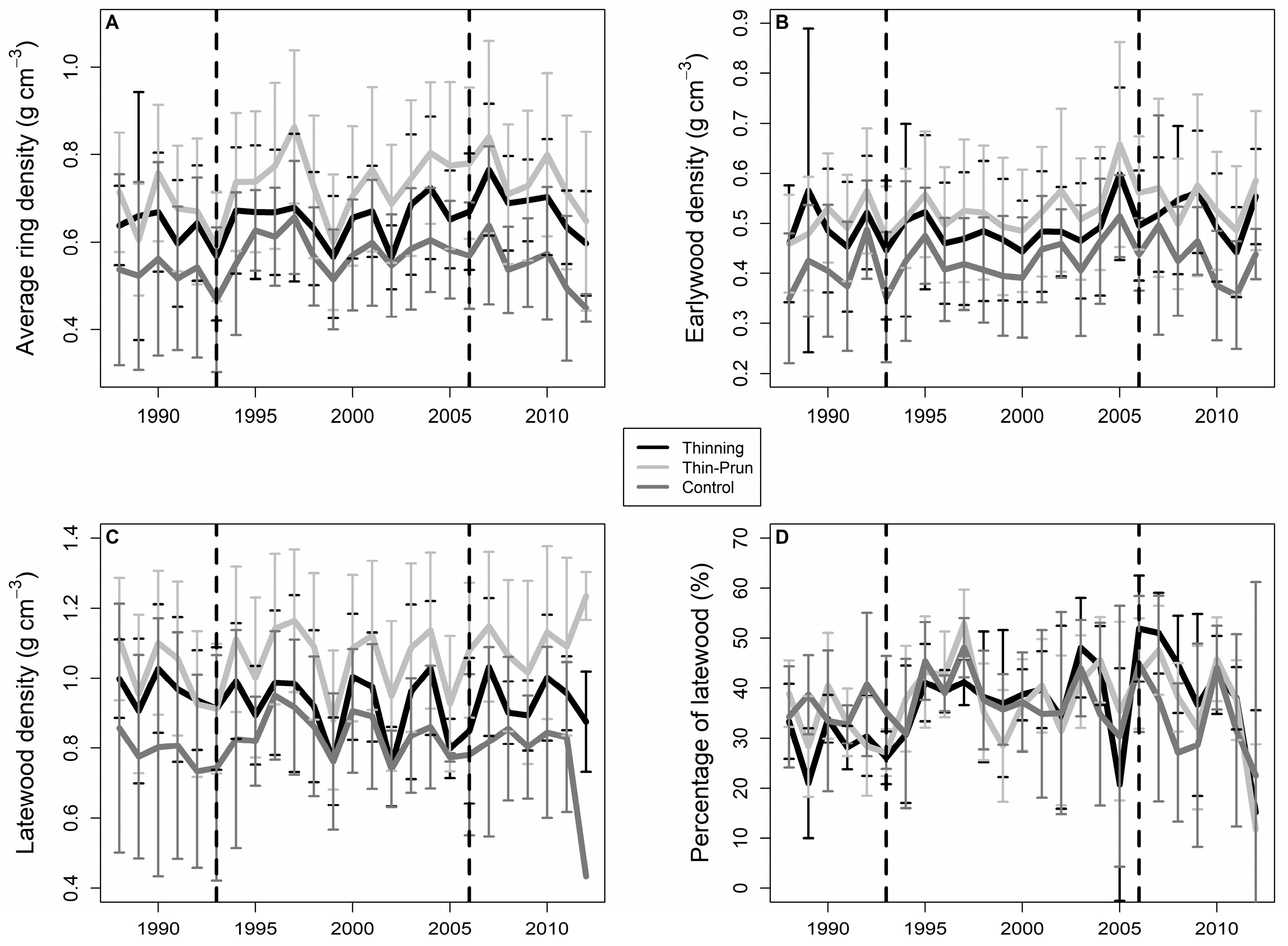
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Macdonald, E.; Hubert, J. A review of the effects of silviculture on timber quality of Sitka spruce. Forestry 2002, 75, 107–138. [Google Scholar] [CrossRef]
- Hevia, A.; Álvarez-González, J.G.; Majada, J. Effects of pruning on knotty core taper and form of Pinus radiata and Pinus pinaster. Eur. J. Wood Wood Prod. 2016, 74, 741–750. [Google Scholar] [CrossRef]
- Mäkinen, H.; Hynynen, J. Wood density and tracheid properties of Scots pine: Responses to repeated fertilization and timing of the first commercial thinning. Forestry 2014, 87, 437–447. [Google Scholar] [CrossRef]
- Moreno-Fernández, D.; Sánchez-González, M.; Álvarez-González, J.G.; Hevia, A.; Majada, J.P.; Cañellas, I.; Gea-Izquierdo, G. Response to the interaction of thinning and pruning of pine species in Mediterranean mountains. Eur. J. For. Res. 2014, 133, 833–843. [Google Scholar] [CrossRef]
- Montagu, K.; Kearney, D.; Smith, R.G. The biology and silviculture of pruning planted eucalypts for clear wood production—A review. For. Ecol. Manag. 2003, 179, 1–13. [Google Scholar] [CrossRef]
- Lin, C.-J.; Wang, S.-Y.; Yang, T.-H.; Tsai, M.-J. Compressive strength of young Taiwania (Taiwania cryptomerioides) trees grown with different thinning and pruning treatments. J. Wood Sci. 2006, 52, 337–341. [Google Scholar] [CrossRef]
- Beets, P.N.; Kimberley, M.O.; McKinley, R.B. Predicting wood density of Pinus radiata annual growth increments. N. Z. J. For. Sci. 2007, 37, 241–266. [Google Scholar]
- Peltola, H.; Kilpeläinen, A.; Sauvala, K.; Räisänen, T.; Ikonen, V. Effects of early thinning regime and tree status on the radial growth and wood density of Scots pine. Silva Fenn. 2007, 41, 489–505. [Google Scholar] [CrossRef]
- Larjavaara, M.; Muller-Landau, H.C. Rethinking the value of high wood density. Funct. Ecol. 2010, 24, 701–705. [Google Scholar] [CrossRef]
- Niklas, K.J.; Spatz, H.-C. Worldwide correlations of mechanical properties and green wood density. Am. J. Bot. 2010, 97, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Elliott, G. Wood Density in Conifers, Technical Communication No. 8; Commonwealth Forestry Bureau: Oxford, UK, 1970.
- Zobel, B.J.; van Buijtenen, J.P. Wood Variation. Its Causes and Control, 1st ed.; Springer: New York, NY, USA, 1989. [Google Scholar]
- Fries, A.; Ericsson, T. Genetic parameters for early wood and latewood densities and development with increasing age in Scots pine. Ann. For. Sci. 2009, 66, 404. [Google Scholar] [CrossRef]
- Camarero, J.J.; Fernández-Pérez, L.; Kirdyanov, A.V.; Shestakova, T.A.; Knorre, A.A.; Kukarskih, V.V.; Voltas, J. Minimum wood density of conifers portrays changes in early season precipitation at dry and cold Eurasian regions. Trees 2017, 31, 1423–1437. [Google Scholar] [CrossRef]
- Sousa, V.B.; Louzada, J.L.; Pereira, H. Age trends and within-site effects in wood density and radial growth in Quercus faginea mature trees. For. Syst. 2016, 25, 1–9. [Google Scholar] [CrossRef]
- Rodríguez Trobajo, E.; Ortega Quero, M. Tendencias radiales de la densidad y sus componentes en Pinus nigra Arn. de la Península Ibérica. Investig. Agrar. Sist. Y Recur. For. 2006, 15, 120–133. [Google Scholar] [CrossRef]
- Jyske, T.; Mäkinen, H.; Saranpää, P. Wood density within Norway spruce stems. Silva Fenn. 2008, 42, 439–455. [Google Scholar] [CrossRef]
- Guller, B.; Isik, K.; Cetinay, S. Variations in the radial growth and wood density components in relation to cambial age in 30-year-old Pinus brutia Ten. at two test sites. Trees 2012, 26, 975–986. [Google Scholar] [CrossRef]
- Kennedy, R. Coniferous wood quality in the future: Concerns and strategies. Wood Sci. Technol. 1995, 29, 321–338. [Google Scholar] [CrossRef]
- Passialis, C.; Kiriazakos, A. Juvenile and mature wood properties of naturally-grown fir trees. Eur. J. Wood Wood Ind. 2004, 62, 476–478. [Google Scholar] [CrossRef]
- Gartner, B.L.; Robbins, J.M.; Newton, M. Effects of pruning on wood density and tracheid length in young Douglas-fir. Wood Fiber Sci. 2005, 37, 304–313. [Google Scholar]
- Panshin, C.; De Zeeuw, A. Textbook of Wood Technology, 4th ed.; McGraw-Hill Book Co.: New York, NY, USA, 1980. [Google Scholar]
- Rozenberg, P.; Franc, A.; Bastien, C.; Cahalan, C. Improving models of wood density by including genetic effects: A case study in Douglas-fir. Ann. For. Sci. 2001, 58, 385–394. [Google Scholar] [CrossRef]
- Olivar, J.; Rathgeber, C.; Bravo, F. Climate Change, tree-ring width and wood density of pines in mediterranean environments. IAWA J. 2015, 36, 257–269. [Google Scholar] [CrossRef]
- Muller-Landau, H.C. Interspecific and inter-site variation in wood specific gravity of tropical trees. Biotropica 2004, 36, 20–32. [Google Scholar] [CrossRef]
- King, D.A.; Davies, S.J.; Tan, S.; Noor, N.S.M. The role of wood density and stem support costs in the growth and mortality of tropical trees. J. Ecol. 2006, 94, 670–680. [Google Scholar] [CrossRef]
- Mäkinen, H.; Saranpää, P.; Linder, S. Wood-density variation of Norway spruce in relation to nutrient optimization and fibre dimensions. Can. J. For. Res. 2006, 32, 185–194. [Google Scholar] [CrossRef]
- Alteyrac, J.; Zhang, S.; Cloutier, A.; Ruel, J.-C. Influence of stand density on ring width and wood density at different sampling heights in black spruce (Picea mariana (Mill.) B.S.P.). Wood Fiber Sci. 2007, 37, 83–94. [Google Scholar]
- Pape, R. Effects of thinning regime on the wood properties and stem quality of Picea abies. Scand. J. For. Res. 1999, 14, 38–50. [Google Scholar] [CrossRef]
- Carson, S.D.; Cown, D.; McKinley, R.; Moore, J. Effects of site, silviculture and seedlot on wood density and estimated wood stiffness in radiata pine at mid-rotation. N. Z. J. For. Sci. 2014, 44, 26. [Google Scholar] [CrossRef]
- Gómez, J.A.; Gutiérrez, A.; Baonza, M.V. Influencia de la intensidad de clara en el crecimiento, la producción y las características de la madera en una masa artificial de Pinus sylvestris L. Investig. Agrar. Sist. Y Recur. For. 1996, 5, 257–273. [Google Scholar]
- Mäkinen, H.; Hynynen, J.; Penttilä, T. Effect of thinning on wood density and tracheid properties of Scots pine on drained peatland stands. Forestry 2015, 359–367. [Google Scholar] [CrossRef]
- Jaakkola, T.; Mäkinen, H.; Saranpää, P. Wood density of Norway spruce: Responses to timing and intensity of first commercial thinning and fertilisation. For. Ecol. Manag. 2006, 237, 513–521. [Google Scholar] [CrossRef]
- Jaakkola, T.; Mäkinen, H.; Saranpää, P. Wood density in Norway spruce: Changes with thinning intensity and tree age. Can. J. For. Res. 2005, 35, 1767–1778. [Google Scholar] [CrossRef]
- Moschler, W.W.; Dougal, E.F.; McRael, D.D. Density and growth ring characteristics of Pinus taeda L. following thinning. Wood Fiber Sci. 1989, 21, 313–319. [Google Scholar]
- Todaro, L.; Macchioni, N. Wood properties of young Douglas-fir in Southern Italy: Results over a 12-year post-thinning period. Eur. J. For. Res. 2010, 130, 251–261. [Google Scholar] [CrossRef]
- Schneider, R.; Zhang, S.Y.; Swift, D.E.; Bégin, J.; Lussier, J.-M. Predicting selected wood properties of jack pine following commercial thinning. Can. J. For. Res. 2008, 38, 2030–2043. [Google Scholar] [CrossRef]
- Johansson, C.J. Grading of timber with respect to mechanical properties. In Timber Engineering; Thelandersson, S., Larsen, H.J., Eds.; John Wiley & Sons: Chichester, UK, 2003. [Google Scholar]
- Cown, J. Effects of severe thinning and pruning treatments on the intrinsic wood properties of young radiata pine. N. Z. J. For. Sci. 1973, 3, 379–389. [Google Scholar]
- Larson, P.R. The indirect effect of drought on tracheid diameter in Red pine. For. Sci. 1963, 9, 52–62. [Google Scholar]
- Mutz, R.; Guilley, E.; Sauter, U.H.; Nepveu, G. Modelling juvenile-mature wood transition in Scots pine (Pinus sylvestris L.) using nonlinear mixed-effects models. Ann. For. Sci 2004, 61, 831–841. [Google Scholar] [CrossRef]
- Koga, S.; Zhang, S.; Bégin, J. Effects of precommercial thinning on annual radial growth and wood density in balsam fir (Abies balsamea). Wood Fiber Sci. 2002, 34, 625–642. [Google Scholar]
- Hevia, A.; Álvarez-González, J.G.; Majada, J. Comparison of pruning effects on tree growth, productivity and dominance of two major timber conifer species. For. Ecol. Manag. 2016, 374, 82–92. [Google Scholar] [CrossRef]
- Hermoso, E.; Fernández-Golfín, J.; Díez, R. Madera estructural de pino silvestre: Caracterización mecánica. Investig. Agrar. Sist. Y Recur. For. 2002, 11, 425–440. [Google Scholar]
- Trasobares, A.; Pukkala, T. Optimising the management of uneven-aged Pinus sylvestris L. and Pinus nigra Arn. mixed stands in Catalonia, north-east Spain. Ann. For. Sci. 2004, 61, 747–758. [Google Scholar] [CrossRef]
- Del Río, M.; Montero, G.; Ortega, C. Respuesta de los distintos regímenes de claras a los daños causados por la nieve en masas de Pinus sylvestris en el Sistema Central. Investig. Agrar. Sist. Y Recur. For. 1997, 6, 103–117. [Google Scholar]
- Pretzsch, H.; Biber, P. Size-symmetric versus size-asymmetric competition and growth partitioning among trees in forest stands along an ecological gradient in central Europe. Can. J. For. Res. 2010, 40, 370–384. [Google Scholar] [CrossRef]
- Schweingruber, F.H. Tree Rings and Environment Dendroecology; Paul Haupt: Bern, Germany, 1996. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree ring dating and measurements. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- Littell, R.; Pendergast, J.; Natarajan, R. Tutorial in biostatistics: Modelling covariance structure in the analysis of repeated measures data. Stat. Med. 2000, 19, 1793–1819. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Viena, Austria, 2017. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models; R Core Team: Viena, Austria, 2017. [Google Scholar]
- Tong, Q.; Tanguay, F.; Zhang, S. Impact of commercial thinning on annual radial growth and wood density in plantation-grown black spruce. Wood Fiber Sci. 2011, 43, 311–325. [Google Scholar]
- Cregg, B.; Dougherty, P.; Hennessey, T. Growth and wood quality of young loblolly pine trees in relation to stand density and climatic factors. Can. J. For. Res. 1988, 18, 851–858. [Google Scholar] [CrossRef]
- Amateis, R.L.; Burkhart, H.E. Growth of young loblolly pine trees following pruning. For. Ecol. Manag. 2011, 262, 2338–2343. [Google Scholar] [CrossRef]
- Gutiérrez Oliva, A.; Baonza Merino, V.; Fernández-Golfín Seco, J.; Conde García, M.; Hermoso Prieto, E. Effect of growth conditions on wood density of Spanish Pinus nigra. Wood Sci. Technol. 2006, 40, 190–204. [Google Scholar] [CrossRef]
- Pazdrowski, W. The proportion and some selected physical and mechanical properties of juvenile, maturing and adult wood of black pine and Scots pine. Electron. J. Pol. Agric. Univ. 2004, 7, 1–6. [Google Scholar]
- Gartner, B.L.; North, E.M.; Johnson, G.R.; Singleton, R. Effects of live crown on vertical patterns of wood density and growth in Douglas-fir. Can. J. For. Res. 2002, 32, 439–447. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Treatment | First Thinning | Second Thinning | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Age | Ho | G | Dg | N | %G | Year | Age | Ho | G | Dg | N | %G | |
| Pinus sylvestris | ||||||||||||||
| C | 1991 | 37 | 11.2 | 38.5 | 14.5 | 2340 | 0.7 * | 2001 | 47 | 13.3 | 46.4 | 17.2 | 2005 | 9.9 * |
| T | 1991 | 37 | 10.6 | 34.1 | 14.6 | 2040 | 28.2 | 2001 | 47 | 13.0 | 31.2 | 20.7 | 930 | 18.6 |
| TPB | 1991 | 37 | 11.0 | 33.9 | 14.4 | 2080 | 34.8 | 2001 | 47 | 13.2 | 29.0 | 21.2 | 825 | 14.4 |
| Pinus nigra nigra | ||||||||||||||
| C | 1993 | 26 | 13.8 | 36.2 | 18.2 | 1400 | 0.0 | 2006 | 39 | 17.7 | 44.1 | 21.2 | 1245 | 0.0 |
| T | 1993 | 26 | 13.4 | 36.0 | 17.8 | 1450 | 41.9 | 2006 | 39 | 17.2 | 31.7 | 23.6 | 725 | 17.6 |
| TPB | 1993 | 26 | 13.8 | 35.8 | 17.7 | 1460 | 40.2 | 2006 | 39 | 17.5 | 32.2 | 23.3 | 765 | 16.5 |
| Pinus nigra salzmannii | ||||||||||||||
| C | 1993 | 31 | 12.3 | 30.9 | 15.7 | 1600 | 0.0 | 2006 | 44 | 14.8 | 37.2 | 18.1 | 1445 | 0.0 |
| T | 1993 | 31 | 12.2 | 30.1 | 15.6 | 1580 | 24.8 | 2006 | 44 | 15.0 | 32.1 | 19.6 | 1070 | 16.9 |
| TPB | 1993 | 31 | 12.4 | 32.4 | 16.6 | 1500 | 30.4 | 2006 | 44 | 14.9 | 31.1 | 20.9 | 905 | 16.9 |
| Species | RD | ED | LD | LWP |
|---|---|---|---|---|
| Pinus sylvestris | ||||
| C | 0.519 (0.088) | 0.433 (0.067) | 33.04 (12.78) | 0.722 (0.165) |
| T | 0.527 (0.090) | 0.443 (0.068) | 24.62 (10.46) | 0.807 (0.163) |
| TPB | 0.578 (0.073) | 0.458 (0.056) | 29.52 (9.21) | 0.890 (0.149) |
| Pinus nigra nigra | ||||
| C | 0.589 (0.093) | 0.445 (0.068) | 37.65 (9.04) | 0.784 (0.240) |
| T | 0.656 (0.121) | 0.507 (0.112) | 30.92 (8.46) | 0.965 (0.212) |
| TPB | 0.659 (0.101) | 0.494 (0.066) | 29.98 (8.25) | 1.046 (0.123) |
| Pinus nigra salzmannii | ||||
| C | 0.566 (0.129) | 0.422 (0.105) | 36.44 (15.54) | 0.834 (0.200) |
| T | 0.658 (0.130) | 0.490 (0.118) | 37.87 (14.31) | 0.934 (0.176) |
| TPB | 0.734 (0.170) | 0.527 (0.125) | 37.74 (13.29) | 1.066 (0.217) |
| Variable | AM5 | Year | Treatment | Treatment × Year |
|---|---|---|---|---|
| Pinus sylvestris | ||||
| RD | <0.0001 | 0.1553 | 0.4975 | 0.4064 |
| ED | <0.0001 | 0.8673 | 0.4257 | 0.3127 |
| LD | <0.0001 | 0.1272 | 0.7550 | 0.7323 |
| LWP | 0.0115 | 0.8462 | 0.4831 | 0.3908 |
| Pinus nigra nigra | ||||
| RD | <0.0001 | 0.8225 | 0.9718 | 0.9708 |
| ED | <0.0001 | 0.0465 | 0.3806 | 0.1945 |
| LD | 0.0019 | 0.0001 | 0.2650 | 0.0627 |
| LWP | 0.4618 | 0.1221 | 0.5939 | 0.4983 |
| Pinus nigra salzmannii | ||||
| RD | <0.0001 | 0.1222 | 0.2480 | 0.0987 |
| ED | <0.0001 | 0.7420 | 0.1584 | 0.0273 |
| LD | <0.0001 | 0.0154 | 0.1332 | 0.0144 |
| LWP | 0.0547 | 0.1746 | 0.4859 | 0.3936 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Fernández, D.; Hevia, A.; Majada, J.; Cañellas, I. Do Common Silvicultural Treatments Affect Wood Density of Mediterranean Montane Pines? Forests 2018, 9, 80. https://doi.org/10.3390/f9020080
Moreno-Fernández D, Hevia A, Majada J, Cañellas I. Do Common Silvicultural Treatments Affect Wood Density of Mediterranean Montane Pines? Forests. 2018; 9(2):80. https://doi.org/10.3390/f9020080
Chicago/Turabian StyleMoreno-Fernández, Daniel, Andrea Hevia, Juan Majada, and Isabel Cañellas. 2018. "Do Common Silvicultural Treatments Affect Wood Density of Mediterranean Montane Pines?" Forests 9, no. 2: 80. https://doi.org/10.3390/f9020080
APA StyleMoreno-Fernández, D., Hevia, A., Majada, J., & Cañellas, I. (2018). Do Common Silvicultural Treatments Affect Wood Density of Mediterranean Montane Pines? Forests, 9(2), 80. https://doi.org/10.3390/f9020080