Responses of Soil Organic Carbon Sequestration Potential and Bacterial Community Structure in Moso Bamboo Plantations to Different Management Strategies in Subtropical China


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Sampling and Analysis
2.3. Physicochemical and Biochemical Analyses of Soil
2.4. Fourier Transform Infrared Analysis
2.5. DNA Extraction, PCR Amplification, and Illumina Sequencing
2.6. 16S rRNA Data Processing
2.7. Statistical Analysis
3. Results
3.1. Physical and Chemical Properties of Soil
3.2. Soil Organic Carbon Structure
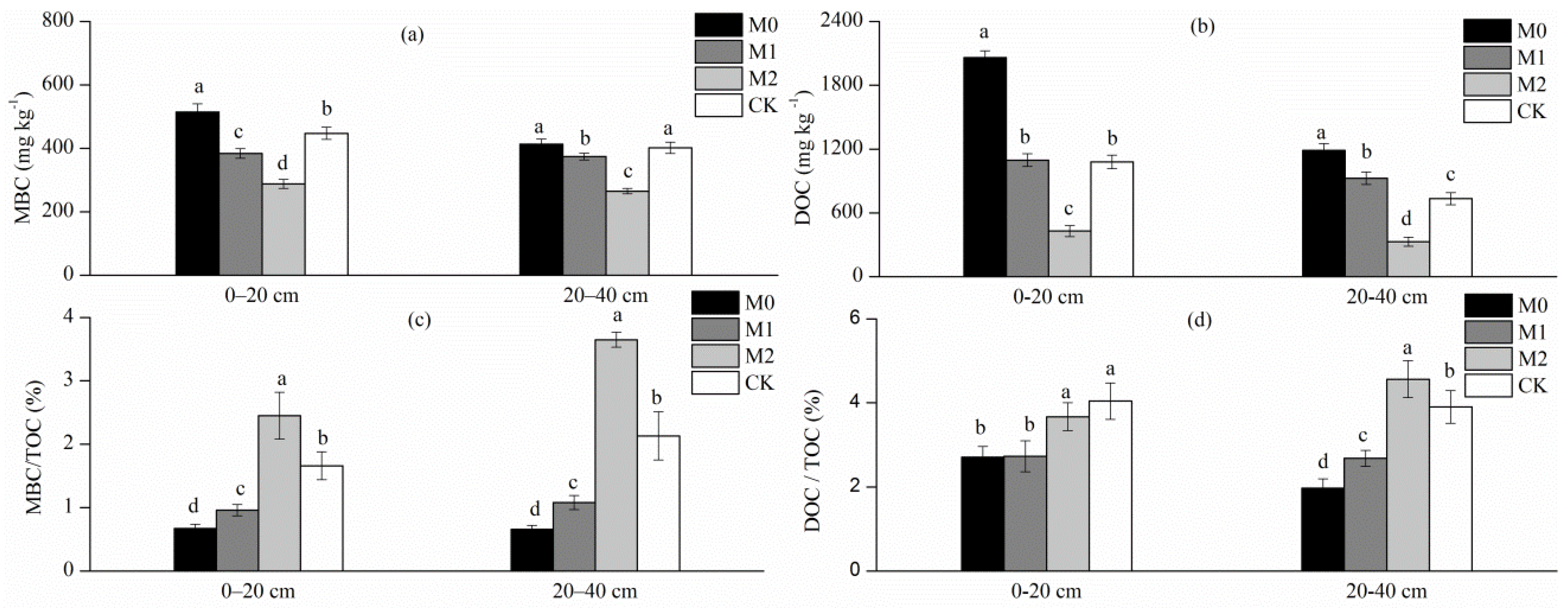
3.3. SOC Mineralization
3.4. Bacterial Community Differences
3.5. Relationships between Soil Carbon-Related Properties and Bacterial Communities
4. Discussion
4.1. Effects of Forest Management Strategies on Soil Properties
4.2. Effects of Forest Management Strategy on the Diversity of Soil Bacterial Communities
4.3. Effects of Changes in Soil Microbiological Properties on Carbon Sequestration Potential and Ecological Indications
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- FAO. Global Forest Resources Assessment; FAO Forestry Paper: Rome, Italy, 2010. [Google Scholar]
- Qin, H.; Niu, L.; Wu, Q.; Chen, J.; Li, Y.; Liang, C.; Xu, Q.; Fuhrmann, J.J.; Shen, Y. Bamboo forest expansion increases soil organic carbon through its effect on soil arbuscular mycorrhizal fungal community and abundance. Plant Soil 2017, 420, 407–421. [Google Scholar] [CrossRef]
- Yen, T.M.; Lee, J.S. Comparing aboveground carbon sequestration between Moso bamboo (phyllostachys heterocycla) and china fir (cunninghamia lanceolata) forests based on the allometric model. For. Ecol. Manag. 2011, 261, 995–1002. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, X.; Zhang, Y.; Booth, T.; He, X. Changes of carbon stocks in bamboo stands in china during 100 years. For. Ecol. Manag. 2009, 258, 1489–1496. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; Gu, H.; Qi, L. Management practices amplify the effects of n deposition on leaf litter decomposition of the Moso bamboo forest. Plant Soil 2015, 395, 391–400. [Google Scholar] [CrossRef]
- Lin, Y.T.; Jangid, K.; Whitman, W.B.; Coleman, D.C.; Chiu, C.Y. Change in bacterial community structure in response to disturbance of natural hardwood and secondary coniferous forest soils in central Taiwan. Microb. Ecol. 2011, 61, 429–437. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Zhang, J.; Meng, M.; Guo, X.; Wu, Y.; Liu, X.; Zhao, K.; Ding, L.; Shao, Y.; Fu, W. Forest-type shift and subsequent intensive management affected soil organic carbon and microbial community in southeastern China. Eur. J. For. Res. 2017, 136, 689–697. [Google Scholar] [CrossRef]
- Tardy, V.; Spor, A.; Mathieu, O.; Lévèque, J.; Terrat, S.; Plassart, P.; Regnier, T.; Bardgett, R.D.; van der Putten, W.H.; Roggero, P.P.; et al. Shifts in microbial diversity through land use intensity as drivers of carbon mineralization in soil. Soil Biol. Biochem. 2015, 90, 204–213. [Google Scholar] [CrossRef]
- Chen, F.; Zheng, H.; Zhang, K.; Ouyang, Z.; Lan, J.; Li, H.; Shi, Q. Changes in soil microbial community structure and metabolic activity following conversion from native Pinus massoniana plantations to exotic eucalyptus plantations. For. Ecol. Manag. 2013, 291, 65–72. [Google Scholar] [CrossRef]
- Xiao, W.; Feng, S.; Liu, Z.; Su, Y.; Zhang, Y.; He, X. Interactions of soil particulate organic matter chemistry and microbial community composition mediating carbon mineralization in karst soils. Soil Biol. Biochem. 2017, 107, 85–93. [Google Scholar] [CrossRef]
- Tang, X.; Fan, S.; Qi, L.; Guan, F.; Du, M.; Zhang, H. Soil respiration and net ecosystem production in relation to intensive management in Moso bamboo forests. Catena 2016, 137, 219–228. [Google Scholar] [CrossRef]
- Xu, Q.; Jiang, P.; Xu, Z. Soil microbial functional diversity under intensively managed bamboo plantations in southern China. J. Soils Sediments 2008, 8, 177–183. [Google Scholar] [CrossRef]
- Liu, J.; Jiang, P.; Wang, H.; Zhou, G.; Wu, J.; Yang, F.; Qian, X. Seasonal soil CO2 efflux dynamics after land use change from a natural forest to Moso bamboo plantations in subtropical China. For. Ecol. Manag. 2011, 262, 1131–1137. [Google Scholar] [CrossRef]
- Sun, D.D.; Xu, Q.F.; Tian, T.; Liu, B.R. Investigation on soil microbial biomass and structure in Phyllostachys edulis plantations with increasing cultivation time. Sci. Silvae Sin. 2011, 47, 181–186. [Google Scholar]
- Lin, Y.T.; Tang, S.L.; Pai, C.W.; Whitman, W.B.; Coleman, D.C.; Chiu, C.Y. Changes in the soil bacterial communities in a cedar plantation invaded by Moso bamboo. Microb. Ecol. 2014, 67, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Liu, E.; Yan, C.; Tian, J.; Zhang, H.; Zhang, Y. Impact of no tillage vs. Conventional tillage on the soil bacterial community structure in a winter wheat cropping succession in northern china. Eur. J. Soil Biol. 2017, 80, 35–42. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, B.; Li, Q.; Yang, H.; Xu, K. Analysis of soil degradation causes in Phyllostachys edulis forests with different mulching years. Forests 2018, 9, 149. [Google Scholar] [CrossRef]
- Zech, W.; Senesi, N.; Guggenberger, G.; Kaiser, K.; Lehmann, J.; Miano, T.; Miltner, A.; Schroth, G. Factors controlling humification and mineralization of soil organic matter in the tropics. Geoderma 1997, 79, 117–161. [Google Scholar] [CrossRef]
- Gross, C.D.; James, J.N.; Turnblom, E.C.; Harrison, R.B. Thinning treatments reduce deep soil carbon and nitrogen stocks in a coastal pacific northwest forest. Forests 2018, 9, 238. [Google Scholar] [CrossRef]
- Chang, E.-H.; Tian, G.; Chiu, C.-Y. Soil microbial communities in natural and managed cloud montane forests. Forests 2017, 8, 33. [Google Scholar] [CrossRef]
- He, D.; Ruan, H. Long term effect of land reclamation from lake on chemical composition of soil organic matter and its mineralization. PLoS ONE 2014, 9, e99251. [Google Scholar] [CrossRef] [PubMed]
- Ellerbrock, R.H.; Gerke, H.H. Characterizing organic matter of soil aggregate coatings and biopores by fourier transform infrared spectroscopy. Eur. J. Soil Sci. 2004, 55, 219–228. [Google Scholar] [CrossRef]
- Rodriguez-Navarro, A.B.; Romanek, C.S.; Alvarez-Lloret, P.; Gaines, K.F. Effect of in ovo exposure to PCBs and Hg on clapper rail bone mineral chemistry from a contaminated salt marsh in coastal Georgia. Environ. Sci. Technol. 2006, 40, 4936–4942. [Google Scholar] [CrossRef] [PubMed]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Guan, D.; Zhou, B.; Zhao, B.; Ma, M.; Qin, J.; Jiang, X.; Chen, S.; Cao, F.; Shen, D.; et al. Influence of 34-years of fertilization on bacterial communities in an intensively cultivated black soil in northeast China. Soil Biol. Biochem. 2015, 90, 42–51. [Google Scholar] [CrossRef]
- West, N.J.; Lepere, C.; Manes, C.L.; Catala, P.; Scanlan, D.J.; Lebaron, P. Distinct spatial patterns of SAR11, SAR86, and actinobacteria diversity along a transect in the ultra-oligotrophic South Pacific Ocean. Front. Microbiol. 2016, 7, 234. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.-J.; Su, J.-Q.; Li, H.; Zhu, Y.-G.; Cao, Z.-H. Bacterial succession along a long-term chronosequence of paddy soil in the yangtze river delta, china. Soil Biol. Biochem. 2017, 104, 59–67. [Google Scholar] [CrossRef]
- Song, X.; Zhou, G.; Jiang, H.; Yu, S.; Fu, J.; Li, W.; Wang, W.; Ma, Z.; Peng, C. Carbon sequestration by chinese bamboo forests and their ecological benefits: Assessment of potential, problems, and future challenges. Environ. Rev. 2011, 19, 418–428. [Google Scholar] [CrossRef]
- Balota, E.L.; Calegari, A.; Nakatani, A.S.; Coyne, M.S. Benefits of winter cover crops and no-tillage for microbial parameters in a brazilian oxisol: A long-term study. Agric. Ecosyst. Environ. 2014, 197, 31–40. [Google Scholar] [CrossRef]
- Wang, H.; Liu, S.-R.; Mo, J.-M.; Wang, J.-X.; Makeschin, F.; Wolff, M. Soil organic carbon stock and chemical composition in four plantations of indigenous tree species in subtropical China. Ecol. Res. 2010, 25, 1071–1079. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, A.; Yang, W.; Zhang, J. Accumulation of organic components and its association with macroaggregation in a sandy loam soil following conservation tillage. Plant Soil 2017, 416, 1–15. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Yu, X. Decline of soil fertility during forest conversion of secondary forest to chinese fir plantations in subtropical china. Land Degrad. Dev. 2011, 22, 444–452. [Google Scholar] [CrossRef]
- Guo, X.; Chen, H.Y.H.; Meng, M.; Biswas, S.R.; Ye, L.; Zhang, J. Effects of land use change on the composition of soil microbial communities in a managed subtropical forest. For. Ecol. Manag. 2016, 373, 93–99. [Google Scholar] [CrossRef]
- Jangid, K.; Williams, M.A.; Franzluebbers, A.J.; Sanderlin, J.S.; Reeves, J.H.; Jenkins, M.B.; Endale, D.M.; Coleman, D.C.; Whitman, W.B. Relative impacts of land-use, management intensity and fertilization upon soil microbial community structure in agricultural systems. Soil Biol. Biochem. 2008, 40, 2843–2853. [Google Scholar] [CrossRef]
- Schneider, D.; Engelhaupt, M.; Allen, K.; Kurniawan, S.; Krashevska, V.; Heinemann, M.; Nacke, H.; Wijayanti, M.; Meryandini, A.; Corre, M.D.; et al. Impact of lowland rainforest transformation on diversity and composition of soil prokaryotic communities in Sumatra (Indonesia). Front. Microbiol. 2015, 6, 1339. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, M.; Flower, C.; Knight, K.; Gonzalez-Meler, M. Evidence of ash tree (Fraxinus spp.) specific associations with soil bacterial community structure and functional capacity. Forests 2018, 9, 187. [Google Scholar] [CrossRef]
- Oni, O.E.; Schmidt, F.; Miyatake, T.; Kasten, S.; Witt, M.; Hinrichs, K.U.; Friedrich, M.W. Microbial communities and organic matter composition in surface and subsurface sediments of the Helgoland mud area, North sea. Front. Microbiol. 2015, 6, 1290. [Google Scholar] [CrossRef] [PubMed]
- Nemergut, D.R.; Townsend, A.R.; Sattin, S.R.; Freeman, K.R.; Fierer, N.; Neff, J.C.; Bowman, W.D.; Schadt, C.W.; Weintraub, M.N.; Schmidt, S.K. The effects of chronic nitrogen fertilization on alpine tundra soil microbial communities: Implications for carbon and nitrogen cycling. Environ. Microbiol. 2008, 10, 3093–3105. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, K.S.; Craine, J.M.; Fierer, N. Consistent effects of nitrogen amendments on soil microbial communities and processes across biomes. Glob. Chang. Biol. 2012, 18, 1918–1927. [Google Scholar] [CrossRef]
- Yan, J.; Wang, L.; Hu, Y.; Tsang, Y.F.; Zhang, Y.; Wu, J.; Fu, X.; Sun, Y. Plant litter composition selects different soil microbial structures and in turn drives different litter decomposition pattern and soil carbon sequestration capability. Geoderma 2018, 319, 194–203. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Land Uses | pH | Bulk Density | TOC | TN | C/N | TP | DON |
|---|---|---|---|---|---|---|---|
| (g cm−3) | (g kg−1) | (g kg−1) | (mg kg−1) | (mg kg−1) | |||
| 0–20 cm | |||||||
| M0 | 4.57 ± 0.1 B | 0.33 ± 0.02 Db | 75.0 ± 4.8 Aa | 8.7 ± 0.5 Aa | 8.76 ± 0.2 A | 134 ± 8 Ab | 212 ± 12 Aa |
| M1 | 4.97 ± 0.1 A | 0.58 ± 0.06 Cb | 40.4 ± 3.5 Ba | 5.8 ± 0.5 Ba | 7.02 ± 0.2 C | 118 ± 7 B | 93 ± 7 Ba |
| M2 | 4.60 ± 0.1 B | 1.18 ± 0.04 Ab | 12.0 ± 2.1 Da | 1.8 ± 0.3 Da | 6.71 ± 0.2 C | 76 ± 3 Ca | 42 ± 6 Da |
| CK | 4.52 ± 0.1 B | 0.85 ± 0.08 Bb | 27.0 ± 2.8 Ca | 3.4 ± 0.3 Ca | 7.85 ± 0.3 B | 80.0 ± 6 Ca | 68 ± 8 Ca |
| 20–40 cm | |||||||
| M0 | 4.51 ± 0.1 BC | 0.50 ± 0.08 Da | 60.5 ± 3.7 Ab | 7.0 ± 0.5 Ab | 8.65 ± 0.4 A | 192 ± 7 Aa | 102 ± 8 Ab |
| M1 | 4.92 ± 0.0 A | 0.72 ± 0.03 Ca | 34.8 ± 3.1 Bb | 4.7 ± 0.4 Bb | 7.38 ± 0.2 B | 122 ± 7 B | 49 ± 5 Bb |
| M2 | 4.60 ± 0.0 B | 1.45 ± 0.06 Aa | 7.3 ± 0.6 Cb | 1.1 ± 0.1 Db | 6.43 ± 0.3 C | 61 ± 4 Db | 19 ± 4 Db |
| CK | 4.44 ± 0.1 C | 1.05 ± 0.04 Ba | 19.1 ± 2.3 Db | 2.4 ± 0.3 Cb | 7.90 ± 0.3 B | 102 ± 6 Cb | 30 ± 5 Cb |
| Land Uses | 1073 | 1158 | 1375 | 1644 | 2851 | 2920 | Hydrophobicity |
|---|---|---|---|---|---|---|---|
| 0–20 cm | |||||||
| M0 | 1.66 ± 0.08 A | 0.53 ± 0.04 A | 0.19 ± 0.01 A | 10.23 ± 0.20 B | 4.25 ± 0.07 A | 7.25 ± 0.07 A | 1.12 ± 0.04 A |
| M1 | 1.08 ± 0.04 C | 0.43 ± 0.01 B | 0.15 ± 0.01 B | 11.71 ± 0.12 A | 3.00 ± 0.01 B | 5.44 ± 0.06 B | 0.72 ± 0.01 B |
| M2 | 0.43 ± 0.02 D | 0.13 ± 0.01 D | 0.06 ± 0.01 D | 4.71 ± 0.13 D | 0.62 ± 0.03 D | 1.34 ± 0.03 D | 0.42 ± 0.01 D |
| CK | 1.19 ± 0.01 B | 0.31 ± 0.01 C | 0.10 ± 0.01 C | 8.05 ± 0.16 C | 1.91 ± 0.04 C | 2.92 ± 0.04 C | 0.60 ± 0.01 C |
| 20–40 cm | |||||||
| M0 | 0.99 ± 0.05 B | 0.37 ± 0.02 A | 0.20 ± 0.04 A | 8.41 ± 0.17 B | 3.10 ± 0.11 A | 5.15 ± 0.11 B | 0.99 ± 0.02 A |
| M1 | 1.33 ± 0.06 A | 0.38 ± 0.01 A | 0.12 ± 0.01 B | 11.30 ± 0.22 A | 3.04 ± 0.06 A | 5.46 ± 0.09 A | 0.75 ± 0.03 B |
| M2 | 0.38 ± 0.02 D | 0.04 ± 0.00 C | 0.04 ± 0.02 C | 6.24 ± 0.11 D | 0.10 ± 0.01 C | 0.30 ± 0.02 D | 0.06 ± 0.01 D |
| CK | 0.74 ± 0.03 C | 0.17 ± 0.01 B | 0.06 ± 0.01 C | 8.08 ± 0.06 C | 1.21 ± 0.05 B | 2.32 ± 0.03 C | 0.44 ± 0.03 C |
| Land Use | C0 (mg C kg−1) | k (day−1) | R2 | Cumulative C Mineralization (mg CO2-C kg−1 soil) | C Mineralization Rate (μg C g−1 SOC d−1) |
|---|---|---|---|---|---|
| 0–20 cm | |||||
| CK | 386.8 ± 16.9 Ca | 0.08 | 0.99 | 374.7 ± 0.06 Ca | 277.6 ± 9.7 Ba |
| M0 | 844.2 ± 36.6 Aa | 0.064 | 0.98 | 805 ± 0.58 Aa | 214.7 ± 8.5 Ca |
| M1 | 466.6 ± 26.3 Ba | 0.08 | 0.99 | 460.2 ± 0.44 Ba | 227.8 ± 9.2 Ca |
| M2 | 292.2 ± 11.3 Da | 0.083 | 0.99 | 280.7 ± 0.04 Da | 467.8 ± 10.1 Aa |
| 20–40 cm | |||||
| CK | 180.7 ± 11.8 Db | 0.082 | 0.98 | 169.5 ± 13.4 Cb | 177.5 ± 9.6 Bb |
| M0 | 496.8 ± 20.6 Ab | 0.049 | 0.97 | 423.1 ± 21.5 Ab | 139.9 ± 6.6 Cb |
| M1 | 357.4 ± 18.6 Bb | 0.053 | 0.99 | 320.3 ± 18.4 Bb | 184.1 ± 7.9 Bb |
| M2 | 154.2 ± 10.3 Bb | 0.075 | 0.99 | 137.6 ± 10.6 Db | 377.0 ± 10.6 Ab |
| Land Use Type | OTUs | Coverage | Chao 1 | Shannon |
|---|---|---|---|---|
| 0–20 cm | ||||
| CK | 1638 ± 313 Ba | 0.98 ± 0.01 | 2101 ± 355 Ba | 8.09 ± 0.06 Ba |
| M0 | 837 ± 366 C | 0.99 ± 0.01 | 942 ± 406 C | 8.11 ± 0.58 Ba |
| M1 | 1398 ± 428 BC | 0.99 ± 0.01 | 1673 ± 406 B | 8.54 ± 0.44 B |
| M2 | 4411 ± 53 Aa | 0.91 ± 0.01 | 6043 ± 43 Aa | 10.03 ± 0.04 Aa |
| 20–40 cm | ||||
| CK | 1069 ± 132 Bb | 0.98 ± 0.01 | 1472 ± 154 Bb | 7.51 ± 0.12 Bb |
| M0 | 496 ± 6 C | 0.99 ± 0.01 | 587 ± 54 C | 7.03 ± 0.50 Bb |
| M1 | 1190 ± 398 B | 0.98 ± 0.01 | 1424 ± 560 AB | 7.76 ± 0.84 B |
| M2 | 1885 ± 470 Ab | 0.98 ± 0.01 | 2204 ± 459 Ab | 9.03 ± 0.32 Ab |
| Variables | Chao 1 | Shannon | MBC | MBC/TOC | Hydrophobicity |
|---|---|---|---|---|---|
| 0–20 cm | |||||
| TOC | −0.797 ** | −0.661 * | 0.816 ** | −0.903 ** | 0.991 ** |
| C mineralization rate | 0.733 ** | 0.723 * | −0.820 ** | 0.893 ** | −0.780 ** |
| 20–40 cm | |||||
| TOC | −0.852 ** | −0.672 * | 0.688 * | −0.906 ** | 0.923 ** |
| C mineralization rate | 0.868 ** | 0.970 ** | −0.843 ** | 0.886 ** | −0.947 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.; Zhong, Z.; Zhang, X.; Bian, F.; Du, X. Responses of Soil Organic Carbon Sequestration Potential and Bacterial Community Structure in Moso Bamboo Plantations to Different Management Strategies in Subtropical China. Forests 2018, 9, 657. https://doi.org/10.3390/f9100657
Yang C, Zhong Z, Zhang X, Bian F, Du X. Responses of Soil Organic Carbon Sequestration Potential and Bacterial Community Structure in Moso Bamboo Plantations to Different Management Strategies in Subtropical China. Forests. 2018; 9(10):657. https://doi.org/10.3390/f9100657
Chicago/Turabian StyleYang, Chuanbao, Zheke Zhong, Xiaoping Zhang, Fangyuan Bian, and Xuhua Du. 2018. "Responses of Soil Organic Carbon Sequestration Potential and Bacterial Community Structure in Moso Bamboo Plantations to Different Management Strategies in Subtropical China" Forests 9, no. 10: 657. https://doi.org/10.3390/f9100657
APA StyleYang, C., Zhong, Z., Zhang, X., Bian, F., & Du, X. (2018). Responses of Soil Organic Carbon Sequestration Potential and Bacterial Community Structure in Moso Bamboo Plantations to Different Management Strategies in Subtropical China. Forests, 9(10), 657. https://doi.org/10.3390/f9100657