
Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance


Abstract
:1. Introduction
1.1. Ecological Resilience in Shrub Systems
1.2. Climate and Fire Severity in the North American Monsoon Region
2. Materials and Methods
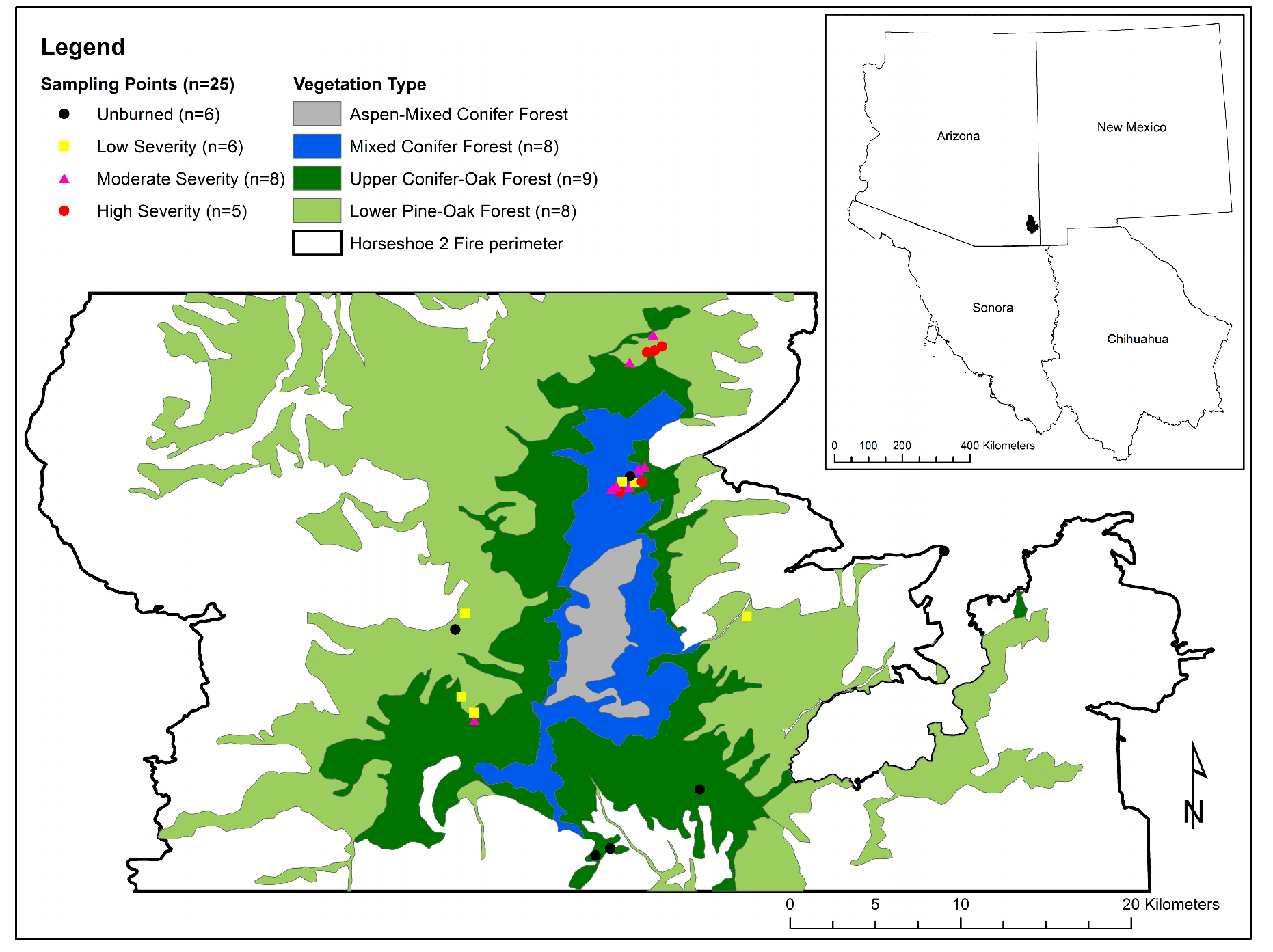
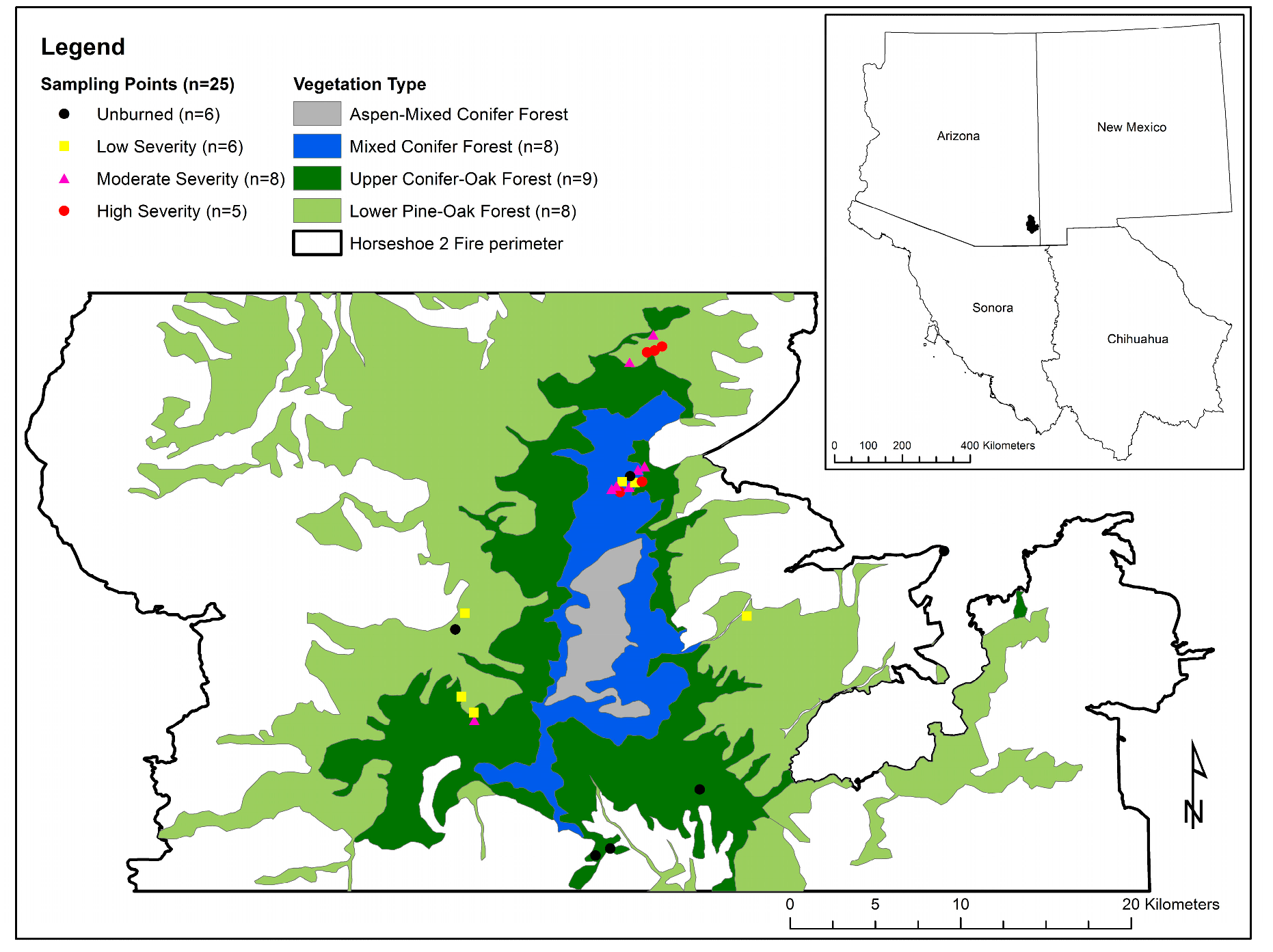
2.1. Study Area
2.2. Shrub Measurements
2.3. Shrub Classification
2.4. Statistical Testing and Regression Analysis
3. Results
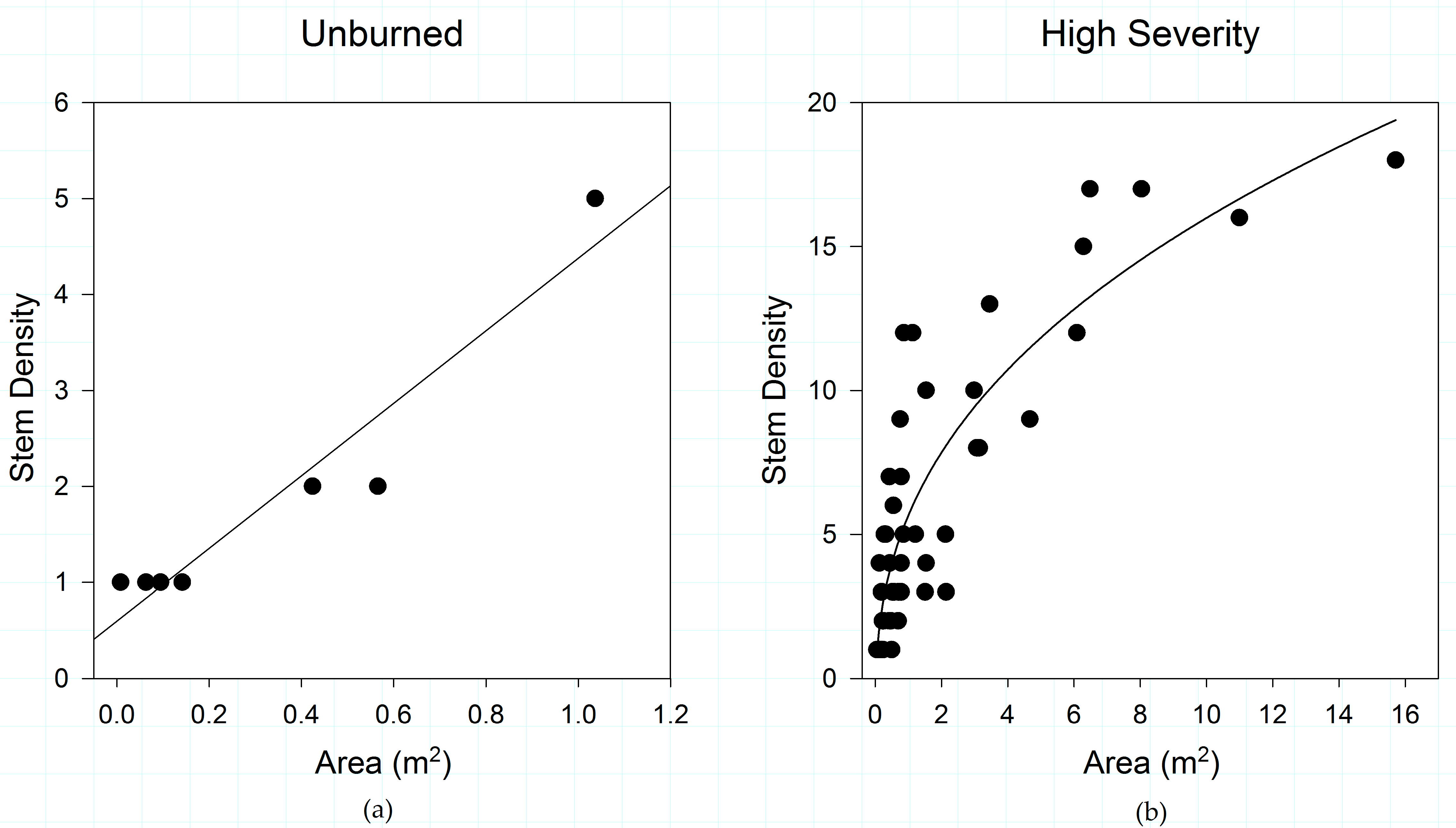
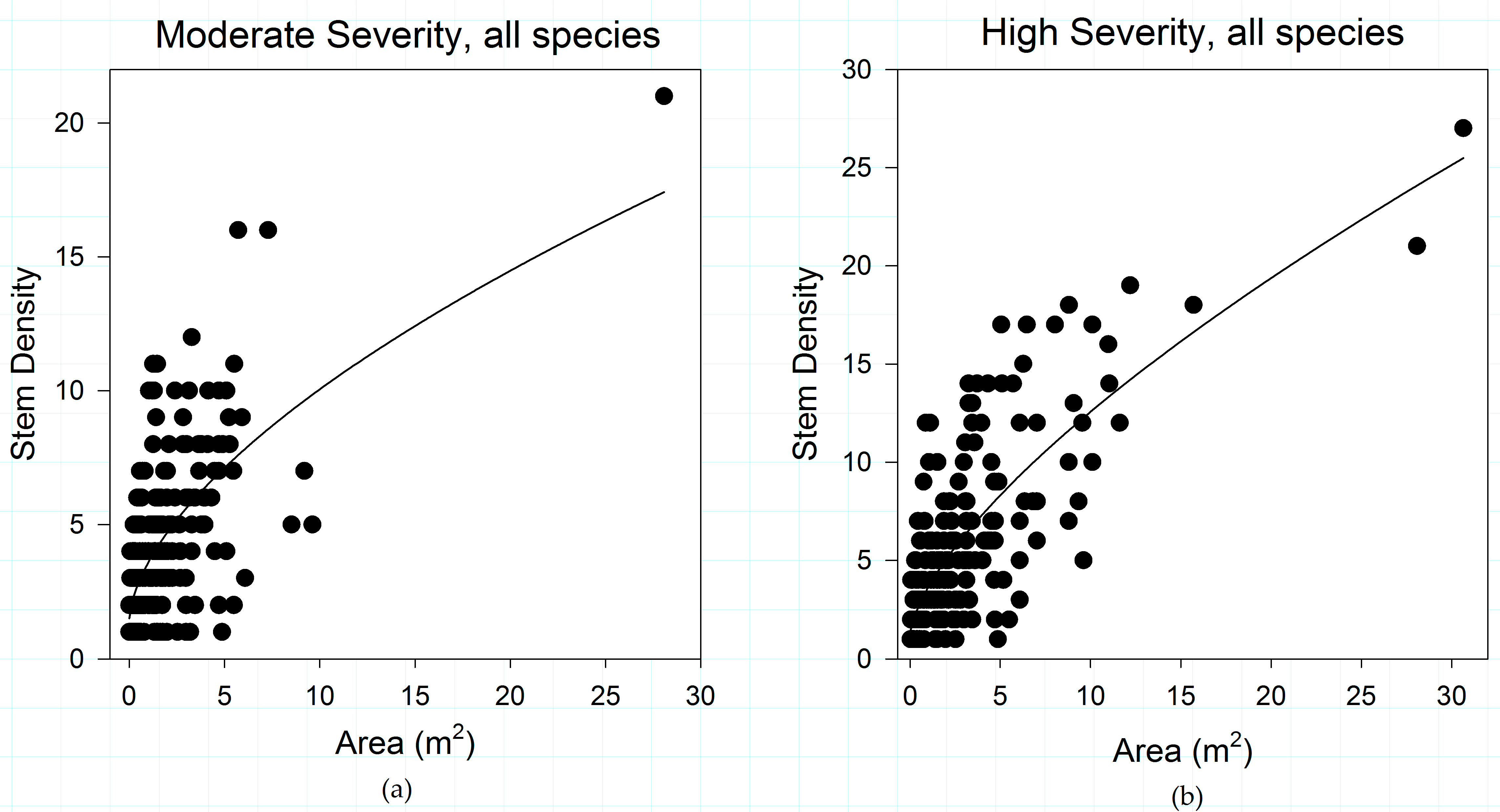
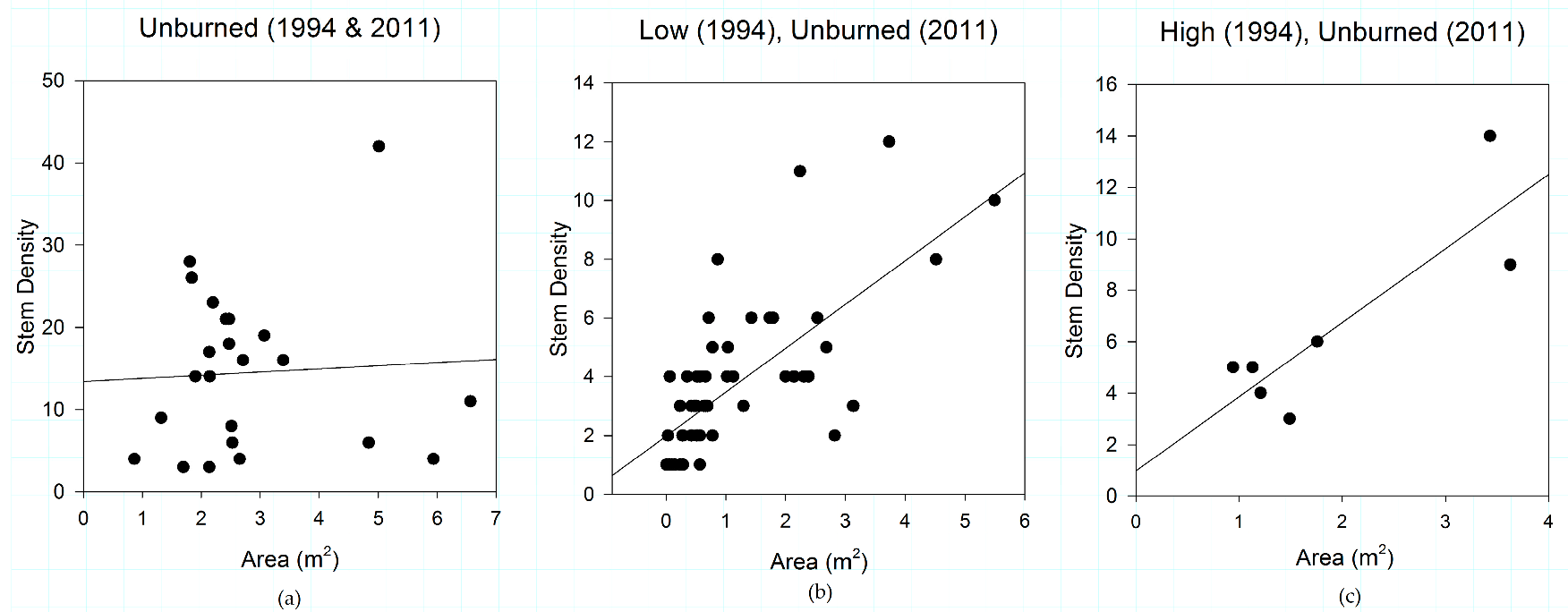
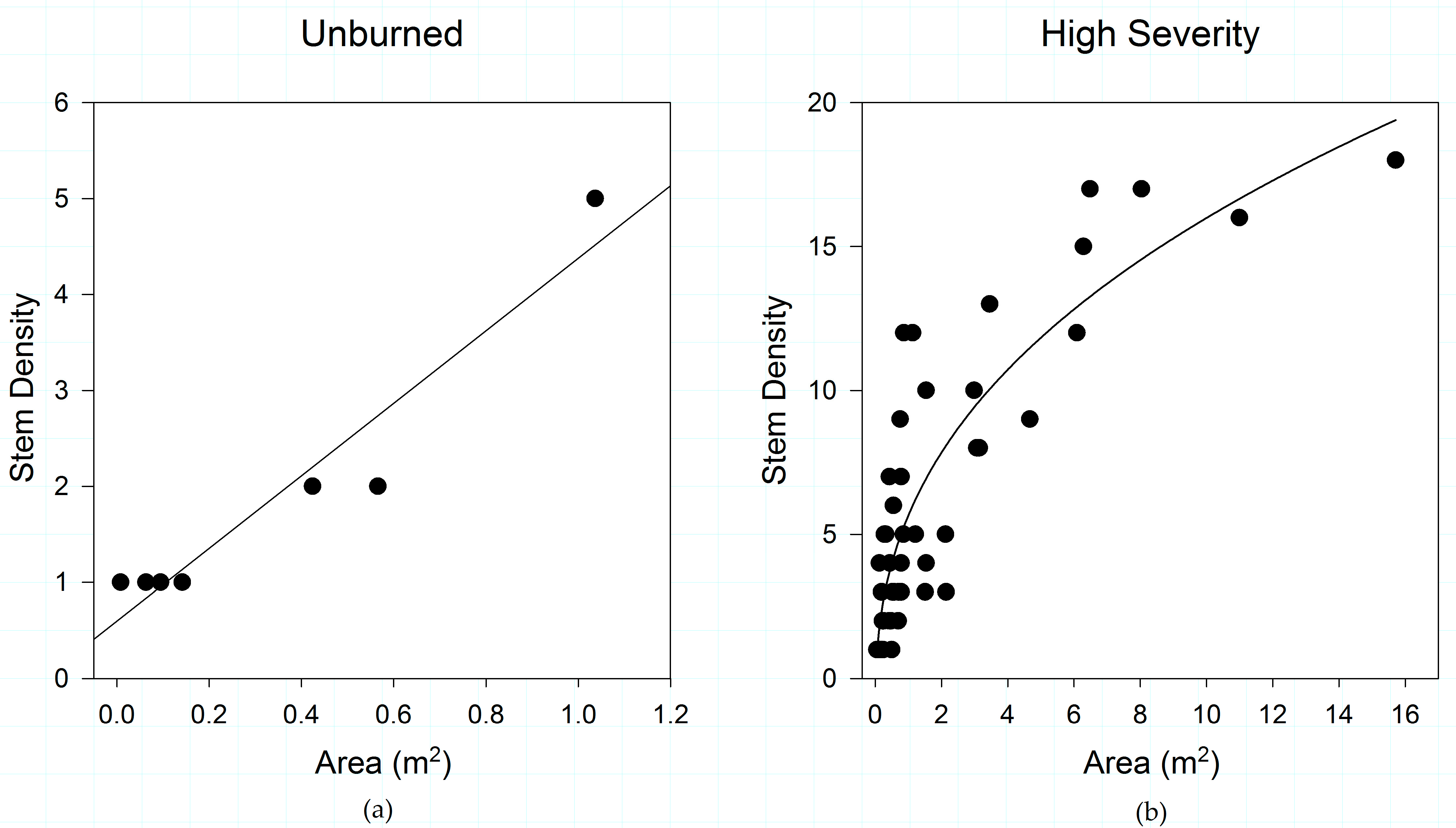
3.1. Fire Severity Influences Shrub Patch Size and Stem Density
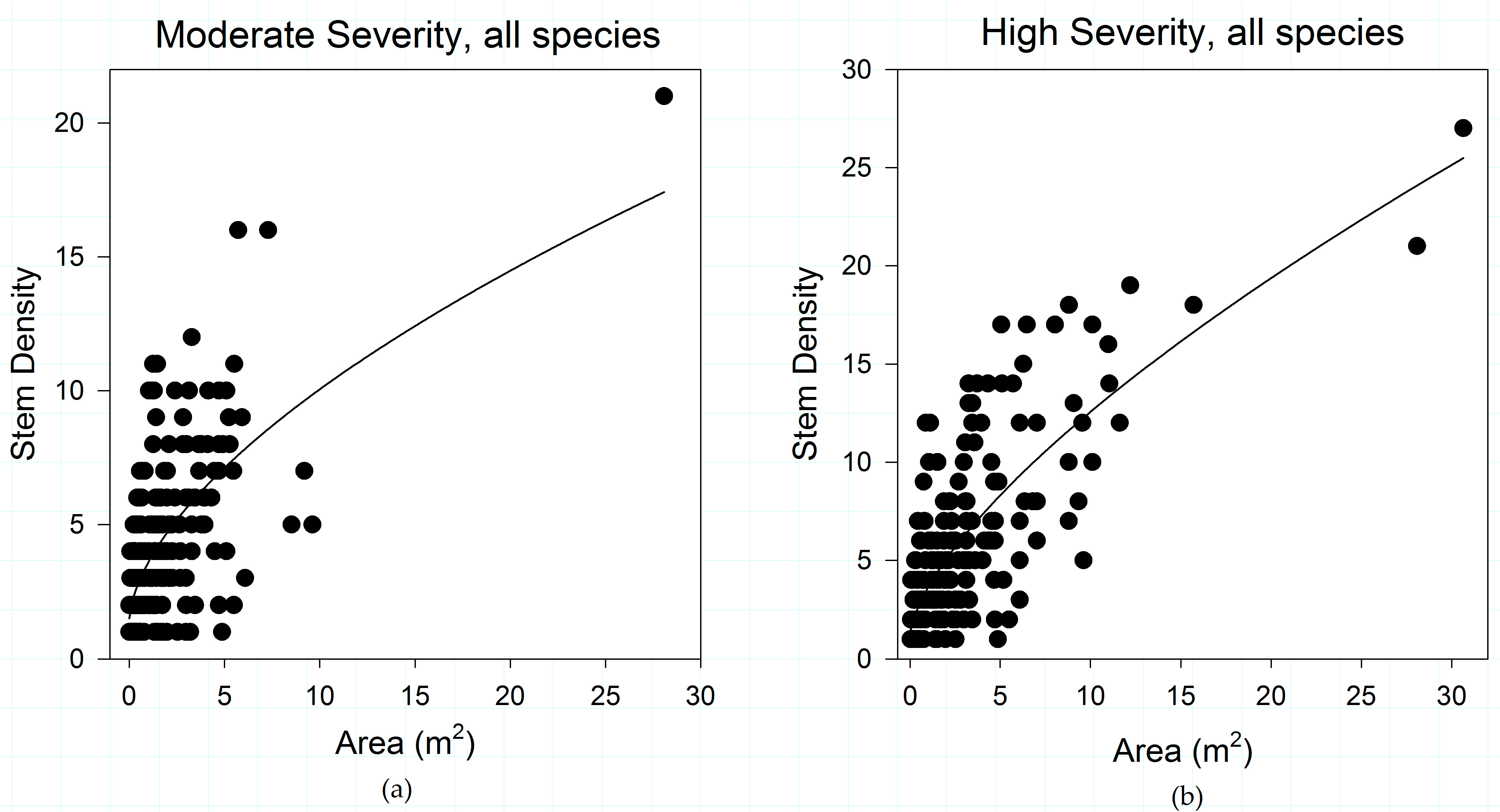
3.2. Shrub Variability across Vegetation Types
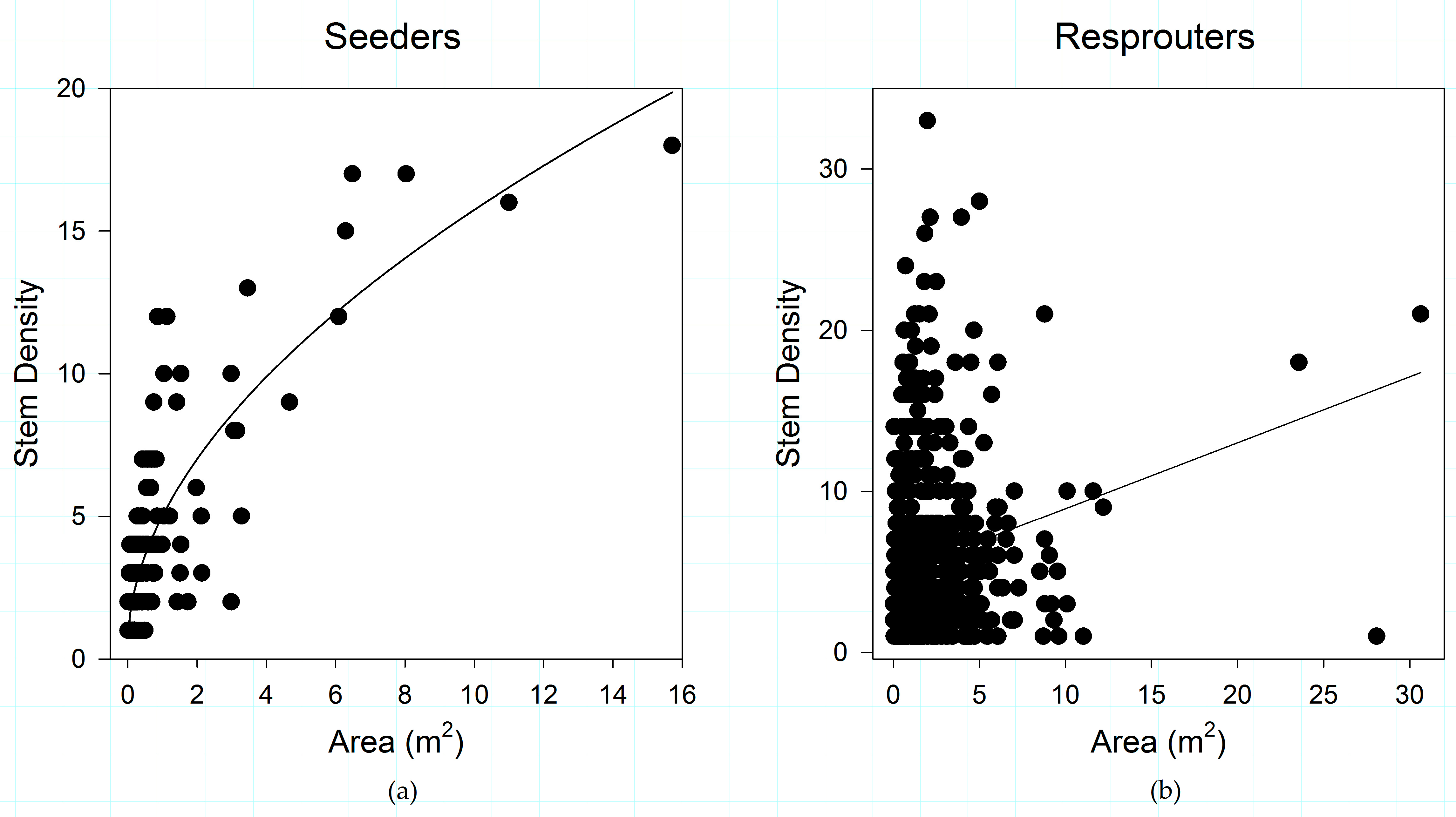
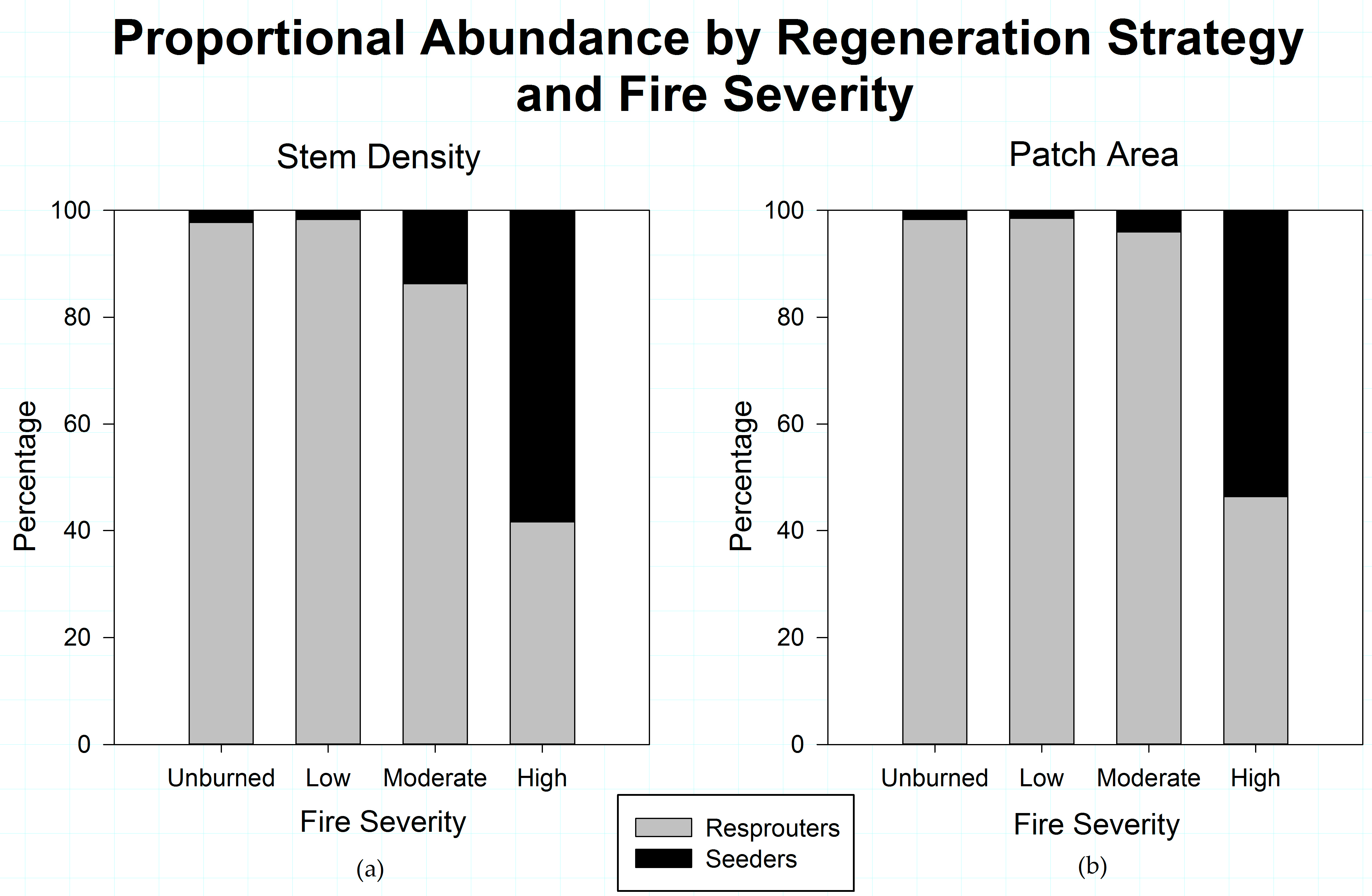
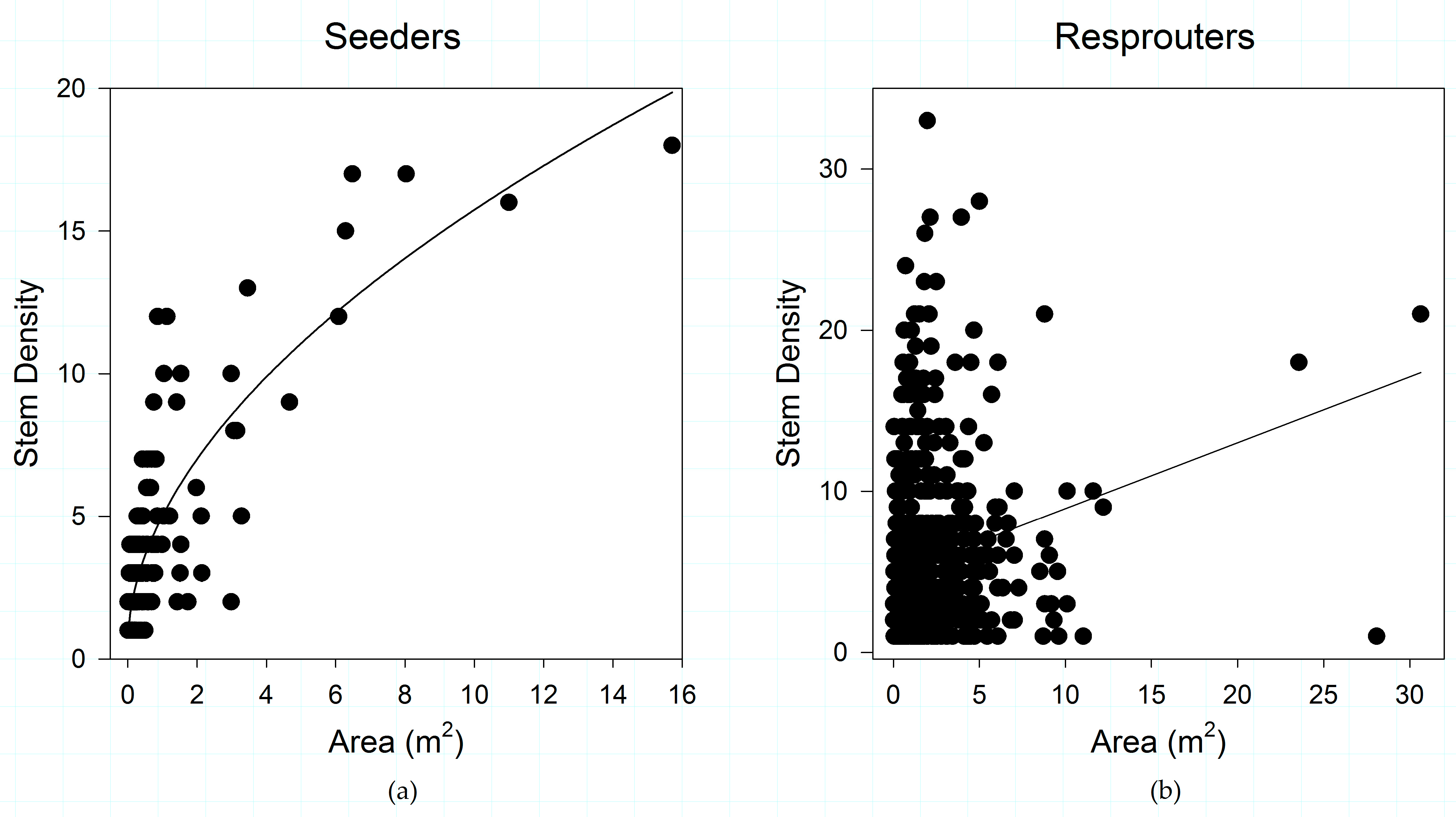
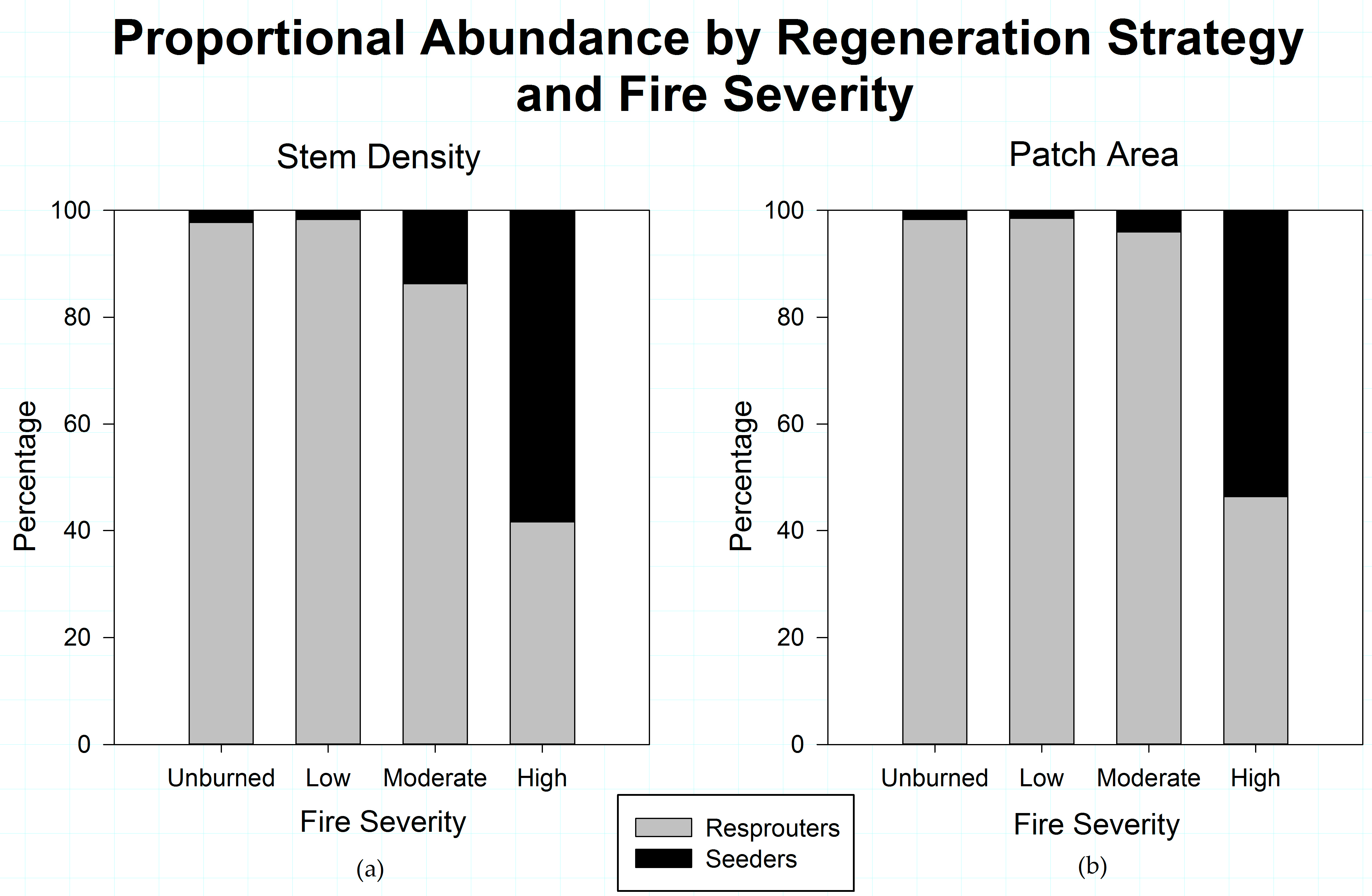
3.3. Post-Disturbance Regeneration Strategies Affect Shrub Patch Dynamics
3.4. Vegetation Type Does Not Predict Variability in Shrub Structure
3.5. Fire Severity Influences Shrub Structure
4. Discussion
4.1. Fire Effects on Shrub Patch Structure
4.2. Post-Fire Regeneration Strategy Affects Post-Disturbance Shrub Patch Dynamics
4.3. Climate Influences on Fire Effects on Shrub Species
4.4. Tipping Points between Forest and Shrubfields
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Species by Post-Fire Regeneration Strategy | Number of Patches (n = 867) |
|---|---|
| Obligate Seeder | 197 |
| Ceanothus fendleri (Rhamnaceae) | 143 |
| Ptelea trifoliata (Rutaceae) | 15 |
| Ribes pinetorum (Grossulariaceae) | 38 |
| Sambucus coerulea (Adoxaceae) | 1 |
| Obligate Resprouter | 670 |
| Arbutus arizonica (Ericaceae) | 1 |
| Cercocarpus betuloides (Rosaceae) | 2 |
| Mimosa biuncifera (Fabaceae) | 34 |
| Physocarpus monogynus (Rosaceae) | 2 |
| Quercus arizonica (Fagaceae) | 239 |
| Quercus emoryi (Fagaceae) | 40 |
| Quercus gambelii (Fagaceae) | 27 |
| Quercus hypoleucoides (Fagaceae) | 219 |
| Quercus rugosa (Fagaceae) | 11 |
| Frangula betulaefolia (Rhamnaceae) | 6 |
| Robinia neomexicana (Fabaceae) | 83 |
| Symphoricarpos rotundifolius (Caprifoliaceae) | 6 |
Appendix B
References
- Buhk, C.; Meyn, A.; Jentsch, A. The Challenge of Plant Regeneration after Fire in the Mediterranean Basin: Scientific Gaps in our Knowledge on Plant Strategies and Evolution of Traits. Plant Ecol. 2007, 192, 1–19. [Google Scholar] [CrossRef]
- Pratt, R.B.; Jacobsen, A.L.; Ramirez, A.R.; Helms, A.M.; Traugh, C.A.; Tobin, M.F.; Heffner, M.S.; Davis, S.D. Mortality of Resprouting Chaparral Shrubs after a Fire and during a Record Drought: Physiological Mechanisms and Demographic Consequences. Glob. Chang. Biol. 2014, 20, 893–907. [Google Scholar] [CrossRef] [PubMed]
- Stevens-Rumann, C.; Morgan, P. Repeated Wildfires Alter Forest Recovery of mixed-conifer Ecosystems. Ecol. Appl. 2016, 26, 1842–1853. [Google Scholar] [CrossRef] [PubMed]
- Shakesby, R.; Doerr, S. Wildfire as a Hydrological and Geomorphological Agent. Earth-Sci. Rev. 2006, 74, 269–307. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a Life History Strategy in Woody Plant Communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an Evolutionary Pressure Shaping Plant Traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef] [PubMed]
- Ackerly, D. Functional Strategies of Chaparral Shrubs in Relation to Seasonal Water Deficit and Disturbance. Ecol. Monogr. 2004, 74, 25–44. [Google Scholar] [CrossRef]
- Clarke, P.J.; Lawes, M.; Midgley, J.; Lamont, B.; Ojeda, F.; Burrows, G.; Enright, N.; Knox, K. Resprouting as a Key Functional Trait: How Buds, Protection and Resources Drive Persistence after Fire. New Phytol. 2013, 197, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Barton, A.M. Intense Wildfire in Southeastern Arizona: Transformation of a Madrean oak-pine Forest to Oak Woodland. For. Ecol. Manag. 2002, 165, 205–212. [Google Scholar] [CrossRef]
- Barton, A. Fire Adaptations in Pines and Oaks: Tree Population Responses to Fire Suppression in Arizona's Madrean Forests. Gen. Tech. Rep. INT-GTR-320 (USA) 1995, 159–163. [Google Scholar]
- Guiterman, C.H.; Margolis, E.Q.; Swetnam, T.W. Dendroecological Methods for Reconstructing High-Severity Fire in Pine-Oak Forests. Tree-Ring Res. 2015, 71, 67–77. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and Stability of Ecological Systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Gunderson, L.H. Ecological resilience—In Theory and Application. Annu. Rev. Ecol. Syst. 2000, 31, 425–439. [Google Scholar] [CrossRef]
- Brooks, M.L.; Chambers, J.C. Resistance to Invasion and Resilience to Fire in Desert Shrublands of North America. Rangel. Ecol. Manag. 2011, 64, 431–438. [Google Scholar] [CrossRef]
- Fire Effects Information System (FEIS). Available online: https://www.feis-crs.org/feis/ (accessed on 20 January 2016).
- Fukami, T.; Nakajima, M. Community Assembly: Alternative Stable States or Alternative Transient States? Ecol. Lett. 2011, 14, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.A. Mega-Fires, Tipping Points and Ecosystem Services: Managing Forests and Woodlands in an Uncertain Future. For. Ecol. Manag. 2013, 294, 250–261. [Google Scholar] [CrossRef]
- Falk, D.A. Are Madrean Ecosystems Approaching Tipping Points? Anticipating Interactions of Landscape Disturbance and Climate Change. In Proceedings of the Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III and 7th Conference on Research and Resource Management in the Southwestern Deserts, Tucson, AZ, USA, 1–5 May 2012; pp. 40–47. [Google Scholar]
- Law, R.; Morton, R.D. Alternative Permanent States of Ecological Communities. Ecology 1993, 74, 1347–1361. [Google Scholar] [CrossRef]
- VanderWeide, B.L.; Hartnett, D.C. Fire Resistance of Tree Species Explains Historical Gallery Forest Community Composition. For. Ecol. Manag. 2011, 261, 1530–1538. [Google Scholar] [CrossRef]
- Graves, S.J.; Rifai, S.W.; Putz, F.E. Outer Bark Thickness Decreases More with Height on Stems of Fire-Resistant than Fire-Sensitive Floridian Oaks (Quercus spp.; Fagaceae). Am. J. Bot. 2014, 101, 2183–2188. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.K.; Comrie, A.C. The North American Monsoon. Bull. Am. Meteorol. Soc. 1997, 78, 2197–2213. [Google Scholar] [CrossRef]
- Biswell, H.H. Prescribed Burning in California Wildlands Vegetation Management; University of California Press: Berkeley, CA, USA, 1989. [Google Scholar]
- Trouet, V.; Taylor, A.H.; Carleton, A.M.; Skinner, C.N. Fire-climate Interactions in Forests of the American Pacific Coast. Geophys. Res. Lett. 2006, 33, L18704. [Google Scholar] [CrossRef]
- Swetnam, T.W.; Betancourt, J.L. Mesoscale Disturbance and Ecological Response to Decadal Climatic Variability in the American Southwest. J. Clim. 1998, 11, 3128–3147. [Google Scholar] [CrossRef]
- Westerling, A.L.; Gershunov, A.; Brown, T.J.; Cayan, D.R.; Dettinger, M.D. Climate and Wildfire in the Western United States. Bull. Am. Meteorol. Soc. 2003, 84, 595–604. [Google Scholar] [CrossRef]
- Barger, N.N.; Archer, S.R.; Campbell, J.L.; Huang, C.; Morton, J.A.; Knapp, A.K. Woody Plant Proliferation in North American Drylands: A Synthesis of Impacts on Ecosystem Carbon Balance. J. Geophys. Res. Biogeosci. 2011, 116, G00K07. [Google Scholar] [CrossRef]
- Knapp, A.K.; Briggs, J.M.; Collins, S.L.; Archer, S.R.; Bret-Harte, M.S.; Ewers, B.E.; Peters, D.P.; Young, D.R.; Shaver, G.R.; Pendall, E. Shrub Encroachment in North American Grasslands: Shifts in Growth Form Dominance Rapidly Alters Control of Ecosystem Carbon Inputs. Glob. Chang. Biol. 2008, 14, 615–623. [Google Scholar] [CrossRef]
- Notaro, M.; Mauss, A.; Williams, J.W. Projected Vegetation Changes for the American Southwest: Combined Dynamic Modeling and bioclimatic-envelope Approach. Ecol. Appl. 2012, 22, 1365–1388. [Google Scholar] [CrossRef] [PubMed]
- Petrie, M.; Collins, S.; Swann, A.; Ford, P.; Litvak, M. Grassland to Shrubland State Transitions Enhance Carbon Sequestration in the Northern Chihuahuan Desert. Glob. Chang. Biol. 2015, 21, 1226–1235. [Google Scholar] [CrossRef] [PubMed]
- Huxman, T.E.; Wilcox, B.P.; Breshears, D.D.; Scott, R.L.; Snyder, K.A.; Small, E.E.; Hultine, K.; Pockman, W.T.; Jackson, R.B. Ecohydrological Implications of Woody Plant Encroachment. Ecology 2005, 86, 308–319. [Google Scholar] [CrossRef]
- Scott, R.L.; Huxman, T.E.; Williams, D.G.; Goodrich, D.C. Ecohydrological Impacts of woody-plant Encroachment: Seasonal Patterns of Water and Carbon Dioxide Exchange within a Semiarid Riparian Environment. Glob. Chang. Biol. 2006, 12, 311–324. [Google Scholar] [CrossRef]
- Barton, A.M. Pines Versus Oaks: Effects of Fire on the Composition of Madrean Forests in Arizona. For. Ecol. Manag. 1999, 120, 143–156. [Google Scholar] [CrossRef]
- Caprio, A.C.; Zwolinski, M.J. Fire and Vegetation in a Madrean Oak Woodland, Santa Catalina Mountains, Southeastern Arizona; USDA For. Serv. Gen. Tech. Rep. RM-218; USDA Forest Service: Fort Collins, CO, USA, 1994; pp. 389–398.
- Keeley, J.E.; Zedler, P.H. Reproduction of Chaparral Shrubs after Fire: A Comparison of Sprouting and Seeding Strategies. Am. Midl. Nat. 1978, 99, 142–161. [Google Scholar] [CrossRef]
- McLaughlin, S.P. An Overview of the Flora of the Sky Islands, Southeastern Arizona: Diversity, Affinities, and Insularity. In Biodiversity and Management of the Madrean Archipelago: The Sky Islands of Southwestern United States and Northwestern Mexico; USDA Forest Service General Technical Report RM-GTR 264; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 1994; pp. 60–70. [Google Scholar]
- Richardson, D.M. Ecology and Biogeography of Pinus; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Van Devender, T.R.; Avila-Villegas, S.; Emerson, M.; Turner, D.; Flesch, A.D.; Deyo, N.S. Biodiversity in the Madrean Archipelago of Sonora, Mexico. In Proceedings of the Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III, Tucson, AZ, USA, 1–5 May 2012; pp. 10–16. [Google Scholar]
- Sawyer, D.A.; Kinraide, T.B. The Forest Vegetation at Higher Altitudes in the Chiricahua Mountains, Arizona. Am. Midl. Nat. 1980, 104, 224–241. [Google Scholar] [CrossRef]
- Malusa, J.; Falk, D.; Laing, L.; Gebow, B. Mapping Ecological Systems in Southeastern Arizona. In Proceedings of the Merging Science and Management in a Rapidly Changing World: Biodiversity and Management of the Madrean Archipelago III and 7th Conference on Research and Resource Management in the Southwestern Deserts, Tucson, AZ, USA, 1–5 May 2012; pp. A1–A7. [Google Scholar]
- Arizona FireScape. Chiricahua Ecological Systems Index. Available online: https://azfirescape.org/chiricahua/ecosystems_descriptions (accessed on 10 May 2015).
- Kaib, M.; Baisan, C.H.; Grissino-Mayer, H.D.; Swetnam, T.W. Fire History of the Gallery Pine-Oak Forests and Adjacent Grasslands of the Chiricahua Mountains of Arizona; United States Department of Agriculture Forest Service General Technical Report RM; United States Department of Agriculture Forest Service: Fort Collins, CO, USA; 1996; pp. 253–264.
- Morino, K.A.; Baisan, C.H.; Swetnam, T.W. Historical Fire Regimes in the Chiricahua Mountains, Arizona: An Examination of Fire along an Elevation Gradient and in Mixed-Confier Forest: Final Report; Laboratory of Tree-Ring Research, The University of Arizona: Tucson, AZ, USA, 2000. [Google Scholar]
- Swetnam, T.W.; Baisan, C.H.; Caprio, A.C.; McCord, A.; Brown, P.M. Fire and Flood in a Canyon Woodland: The Effects of Floods and Debris Flows on the Past Fire Regime of Rhyolite Canyon, Chiricahua National Monument; Report to the National Park Service; The University of Arizona: Tucson, AZ, USA, 1990; pp. 1–51. [Google Scholar]
- Seklecki, M.T.; Grissino-Mayer, H.D.; Swetnam, T.W. Fire History and the Possible Role of Apache-Set Fires in the Chiricahua Mountains of Southeastern Arizona; United States Department of Agriculture Forest Service General Technical Report RM; United States Department of Agriculture Forest Service: Fort Collins, CO, USA, 1996; pp. 238–246.
- Minor, J.J.; (University of Arizona, Tucson, AZ, USA). Personal communication, 2017.
- Monitoring Trends in Burn Severity: MTBS Project. MTBS Data Access: Fire Level Geospatial Data. Available online: http://mtbs.gov/dataquery/individualfiredata.html (accessed on 20 May 2015).
- LANDFIRE. LANDFIRE: Existing Vegetation Type. Available online: http://landfire.cr.usgs.gov/viewer/ (accessed on 12 May 2015).
- Pavek, D.S. Quercus arizonica. 1994. Available online: https://www.fs.fed.us/database/feis/plants/tree/queari/all.html (accessed on 20 January 2016).
- Aleksoff, K.C. Symphoricarpos oreophilus. 1999. Available online: https://www.fs.fed.us/database/feis/plants/shrub/symore/all.html (accessed on 20 January 2016).
- Simonin, K.A. Quercus gambelii. 2000. Available online: https://www.fs.fed.us/database/feis/plants/tree/quegam/all.html (accessed on 20 January 2016).
- Cook, R.D.; Weisberg, S. Residuals and Influence in Regression; Chapman and Hall: New York, NY, USA, 1982. [Google Scholar]
- Lavorel, S. Ecological Diversity and Resilience of Mediterranean Vegetation to Disturbance. Divers. Distrib. 1999, 5, 3–13. [Google Scholar] [CrossRef]
- Scasta, J.D.; Engle, D.M.; Harr, R.N.; Debinski, D.M. Fire Induced Reproductive Mechanisms of a Symphoricarpos (Caprifoliaceae) Shrub after Dormant Season Burning. Bot. Stud. 2014, 55, 80. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, Á.; Prior, L.D.; Perry, G.L.; Bowman, D.M. High Post-Fire Mortality of Resprouting Woody Plants in Tasmanian Mediterranean-Type Vegetation. Int. J. Wildland Fire 2017, 26, 532–537. [Google Scholar] [CrossRef]
- Keeley, J.E. Role of Fire in Seed Germination of Woody Taxa in California Chaparral. Ecology 1987, 68, 434–443. [Google Scholar] [CrossRef]
- Keeley, J.E.; Fotheringham, C. Role of Fire in Regeneration from Seed. In Seeds: The Ecology of Regeneration in Plant Communities, 2nd ed.; Fenner, M., Ed.; CABI: Wallingford, UK, 2000; pp. 311–330. [Google Scholar]
- Zouhar, K.L. Ceanothus greggii. 2000. Available online: https://www.fs.fed.us/database/feis/plants/shrub/ceagre/all.html (accessed on 20 January 2016).
- Quick, C.R. Notes on the Germination of Ceanothus Seeds. Madroño 1935, 3, 135–140. [Google Scholar]
- Sweeney, J.R. Responses of Vegetation to Fire: A Study of the Herbaceous Vegetation Following Chaparral Fires; University of California Publications in Botany; University of California Press: Berkeley, CA, USA, 1956; Volume 28, pp. 143–216. [Google Scholar]
- Crane, M.F. Sambucus nigra subsp. cerulea. 1989. Available online: https://www.fs.fed.us/database/feis/plants/shrub/samnigc/all.html (accessed on 20 January 2016).
- Quick, C.R. Resurgence of a Gooseberry Population after Fire in Mature Timber. J. For. 1962, 60, 100–103. [Google Scholar]
- Sparks, J.C.; Masters, R.E.; Engle, D.M.; Palmer, M.W.; Bukenhofer, G.A. Effects of Late growing-season and Late dormant-season Prescribed Fire on Herbaceous Vegetation in Restored pine-grassland Communities. J. Veg. Sci. 1998, 9, 133–142. [Google Scholar] [CrossRef]
- Knapp, E.E.; Schwilk, D.W.; Kane, J.M.; Keeley, J.E. Role of Burning Season on Initial Understory Vegetation Response to Prescribed Fire in a Mixed Conifer Forest. Can. J. For. Res. 2006, 37, 11–22. [Google Scholar] [CrossRef]
- Garrison, G.A. Carbohydrate Reserves and Response to Use; USDA For. Serv. Gen. Tech. Rep. GTR-INT-1; United States Department of Agriculture Forest Service: Fort Collins, CO, USA, 1972.
- Harrington, M.G. Notes: The Effects of Spring, Summer, and Fall Burning on Gambel Oak in a Southwestern Ponderosa Pine Stand. For. Sci. 1985, 31, 156–163. [Google Scholar]
- Swetnam, T.W.; Betancourt, J.L. Fire-Southern Oscillation Relations in the Southwestern United States. Science 1990, 249, 1017. [Google Scholar] [CrossRef] [PubMed]
- Enright, N.; Fontaine, J.; Westcott, V.; Lade, J.; Miller, B. Fire Interval Effects on Persistence of Resprouter Species in Mediterranean-Type Shrublands. Plant Ecol. 2011, 212, 2071–2083. [Google Scholar] [CrossRef]
- Stone, E.; Stone, M. Root Collar Sprouts in Pine. J. For. 1954, 52, 487–491. [Google Scholar]
- McCune, B. Ecological Diversity in North American Pines. Am. J. Bot. 1988, 353–368. [Google Scholar] [CrossRef]
- Rowe, J.S. Concepts of Fire Effects on Plant Individuals and Species. In The Role of Fire in Northern Circumpolar Ecosystems; Rowe, R.W., MacLean, D.A., Eds.; John Wiley & Sons: Chichester, UK, 1983; pp. 135–154. [Google Scholar]
- Babb, G.D. Sprouting Response of Quercus arizonica and Quercus emoryi Following Fire. Master’s Thesis, The University of Arizona, Tucson, AZ, USA, 1992. [Google Scholar]
- Schwilk, D.W.; Gaetani, M.S.; Poulos, H.M. Oak Bark Allometry and Fire Survival Strategies in the Chihuahuan Desert Sky Islands, Texas, USA. PLoS ONE 2013, 8, e79285. [Google Scholar] [CrossRef] [PubMed]
- Barton, A.M. Factors Controlling Plant Distributions: Drought, Competition, and Fire in Montane Pines in Arizona. Ecol. Monogr. 1993, 63, 367–397. [Google Scholar] [CrossRef]
- Poulos, H.M. A Review of the Evidence for Pine-Oak Niche Differentiation in the American Southwest. J. Sustain. For. 2009, 28, 92–107. [Google Scholar] [CrossRef]
- White, A.S. Presettlement Regeneration Patterns in a Southwestern Ponderosa Pine Stand. Ecology 1985, 66, 589–594. [Google Scholar] [CrossRef]
- Brown, D.J.; Mali, I.; Forstner, M.R. Wildfire and Postfire Restoration Action Effects on Microclimate and Seedling Pine Tree Survivorship. J. Fish Wildl. Manag. 2014, 5, 174–182. [Google Scholar] [CrossRef]

| Fire Severity | Patch Area (m2) | Stem Density (Stems/m2) | ||||||
|---|---|---|---|---|---|---|---|---|
| Mean | Median | Maximum | C.V. | Mean | Median | Maximum | C.V. | |
| Unburned (n = 84) | 1.7 | 1.3 | 6.5 | 0.9 | 4.1 * | 3.1 | 6.4 | 1.2 |
| Low (n = 286) | 1.5 | 0.9 | 26.4 | 1.6 | 3.9 | 4.6 | 2.3 | 0.6 |
| Moderate (n = 312) | 1.4 * | 0.6 | 28.1 | 1.6 | 2.6 * | 4.8 | 0.7 | 0.5 |
| High (n = 184) | 2.7 * | 1.6 | 30.6 | 1.4 | 2.0 * | 2.6 | 0.9 | 0.6 |
| Fire Severity | Stem Density (Stems/m2) | ||||
|---|---|---|---|---|---|
| Unburned | Low | Moderate | High | ||
| Canopy Area (m2) | Unburned | 8.7 | 10.9 ** | 5.6 | |
| Low | 1.5 | 10.0 ** | 7.0 * | ||
| Moderate | 1.4 * | 1.3 * | 8.7 ** | ||
| High | 2.4 * | 2.0 ** | 2.0 ** | ||
| Vegetation Type | Species | Total Shrub Patch Area (m2) | Measured Shrub Cover (%) | Measured Patches (n) |
|---|---|---|---|---|
| Lower Pine-Oak | Quercus arizonica | 435.6 | 47.8% | 199 |
| Quercus hypoleucoides | 325.3 | 35.7% | 139 | |
| Ceanothus fenderli † | 50.7 | 5.6% | 116 | |
| Quercus emoryi | 36.7 | 4.0% | 40 | |
| Quercus rugosa | 23.1 | 2.5% | 6 | |
| Mimosa biuncifera | 21.5 | 2.4% | 34 | |
| Quercus gambelii | 8.0 | 0.9% | 5 | |
| Symphoricarpos rotundifolius | 6.3 | 0.7% | 2 | |
| Robinia neomexicana | 3.7 | 0.4% | 4 | |
| Physocarpus monogynus | 0.2 | 0.1% | 1 | |
| Total | 911.1 | - | 546 | |
| Upper Pine-Oak | Quercus hypoleucoides | 142.0 | 27.0% | 76 |
| Robinia neomexicana | 134.5 | 25.6% | 56 | |
| Quercus arizonica | 93.8 | 17.9% | 39 | |
| Ceanothus fenderli † | 63.3 | 12.1% | 25 | |
| Quercus gambelii | 49.7 | 9.5% | 22 | |
| Quercus rugosa | 21.9 | 4.2% | 5 | |
| Cercocarpus betuloides | 8.9 | 1.7% | 2 | |
| Frangula betulaefolia | 3.8 | 0.7% | 5 | |
| Physocarpus monogynus | 1.7 | 0.3% | 1 | |
| Arbutus arizonica | 1.5 | 0.3% | 1 | |
| Symphoricarpos rotundifolius | 0.7 | 0.1% | 3 | |
| Ribes pinetorum † | 0.2 | 0.1% | 2 | |
| Total | 522 | - | 237 | |
| Mixed Conifer | Robinia neomexicana | 73.7 | 65.0% | 23 |
| Ribes pinetorum † | 17.6 | 16.2% | 35 | |
| Ptelea trifoliata † | 5.6 | 5.7% | 15 | |
| Quercus hypoleucoides | 3.6 | 3.9% | 4 | |
| Sambucus coerulea † | 3.0 | 3.4% | 1 | |
| Frangula betulaefolia | 2.2 | 2.8% | 1 | |
| Ceanothus fenderli † | 0.2 | 1.1% | 1 | |
| Symphoricarpos rotundifolius | 0.05 | 1.0% | 1 | |
| Quercus arizonica | 0.01 | 0.9% | 1 | |
| Total | 106.0 | - | 84 |
| Vegetation Type | Obligate Seeders | Obligate Resprouters | Total n |
|---|---|---|---|
| Lower Pine-Oak | 21.2 | 78.8 * | 546 |
| Upper Pine-Oak | 11.4 | 88.6 * | 237 |
| Mixed Conifer | 63.9 * | 36.1 | 84 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minor, J.; Falk, D.A.; Barron-Gafford, G.A. Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance. Forests 2017, 8, 221. https://doi.org/10.3390/f8070221
Minor J, Falk DA, Barron-Gafford GA. Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance. Forests. 2017; 8(7):221. https://doi.org/10.3390/f8070221
Chicago/Turabian StyleMinor, Jesse, Donald A. Falk, and Greg A. Barron-Gafford. 2017. "Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance" Forests 8, no. 7: 221. https://doi.org/10.3390/f8070221
APA StyleMinor, J., Falk, D. A., & Barron-Gafford, G. A. (2017). Fire Severity and Regeneration Strategy Influence Shrub Patch Size and Structure Following Disturbance. Forests, 8(7), 221. https://doi.org/10.3390/f8070221