
Contrasting Effects of Fire Severity on the Regeneration of Pinus halepensis Mill. and Resprouter Species in Recently Thinned Thickets


Abstract
:1. Introduction
2. Material and Methods
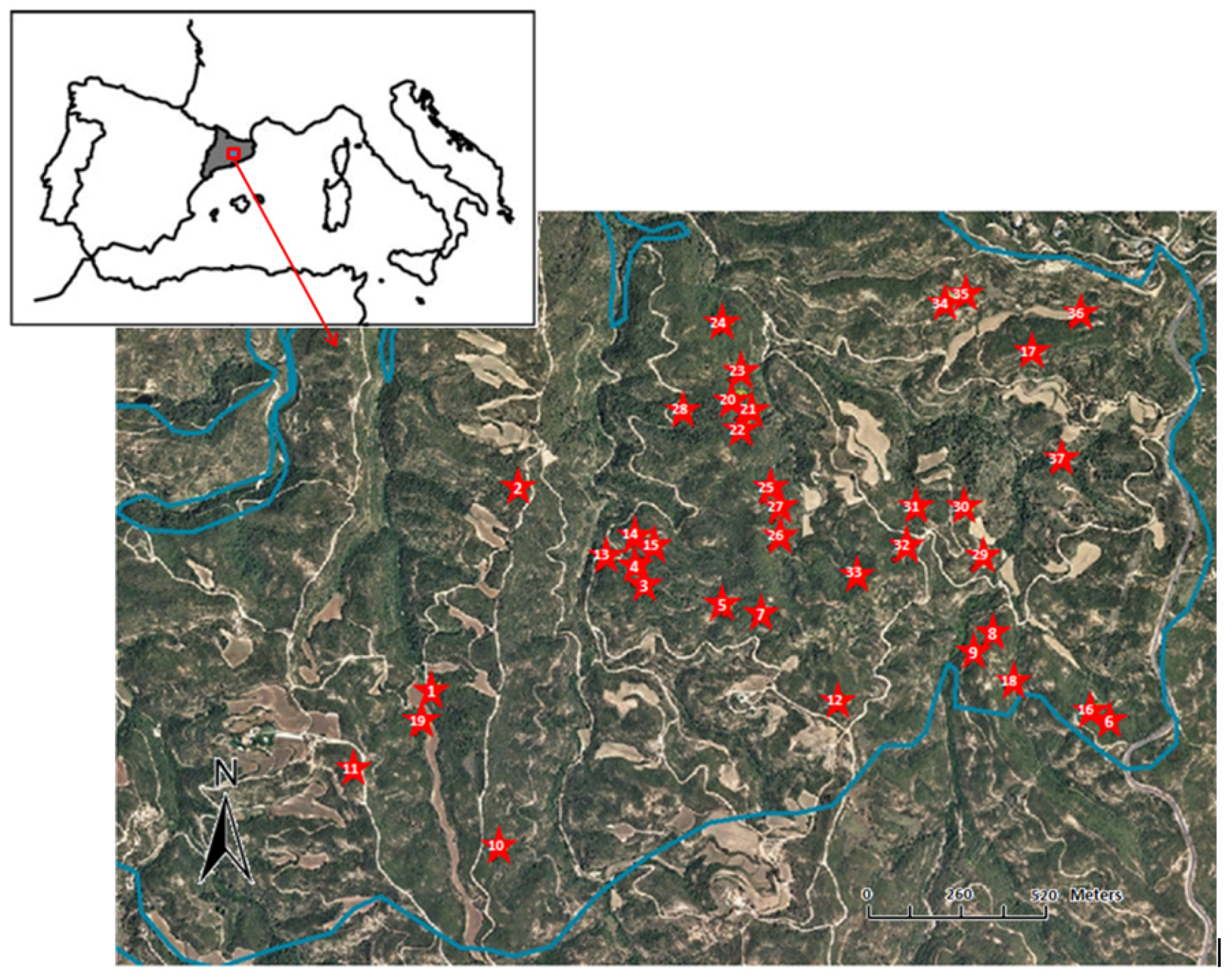
2.1. Study Area and Species
2.2. Experimental Design and Sampling
2.3. Data Analysis
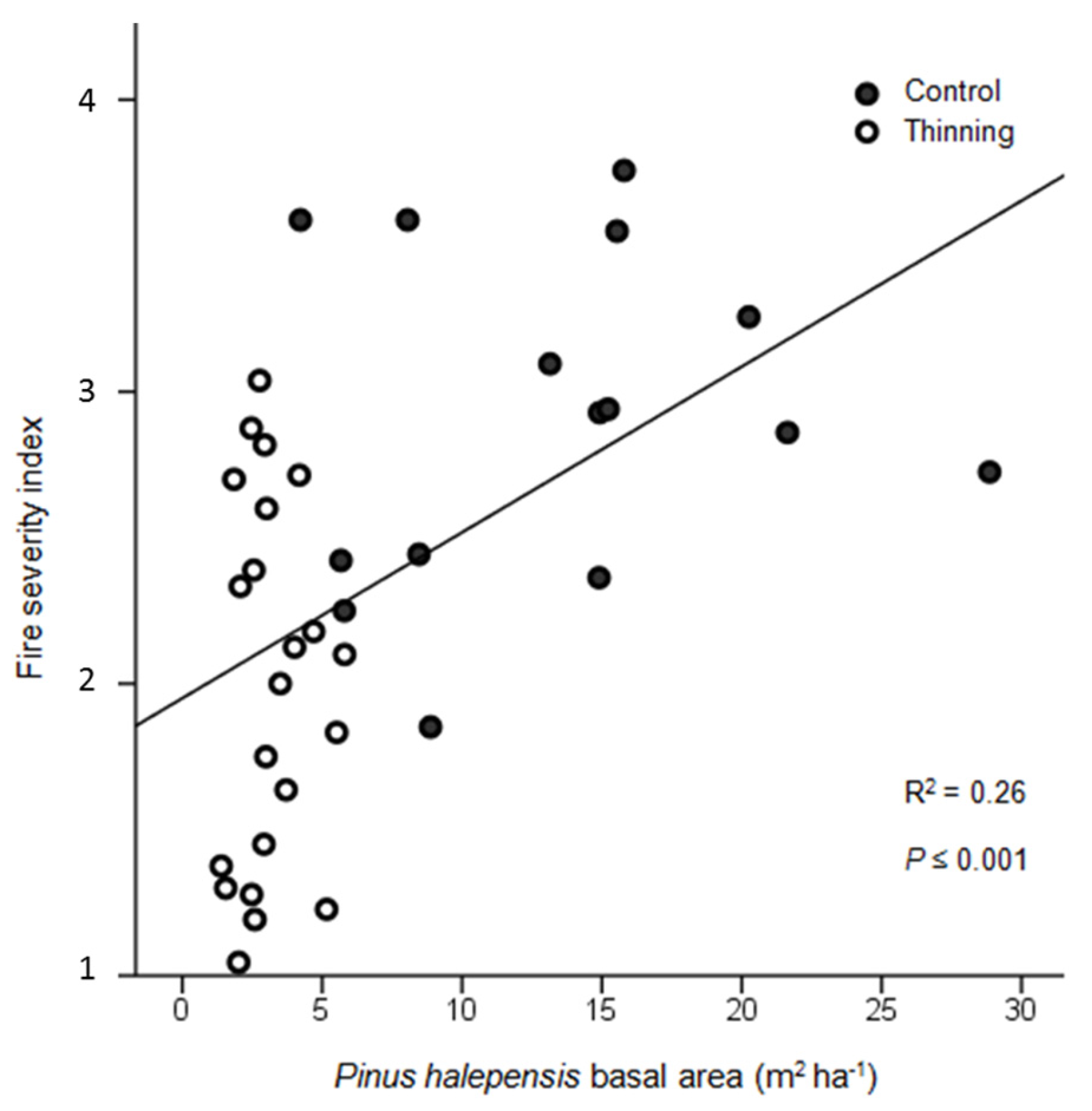
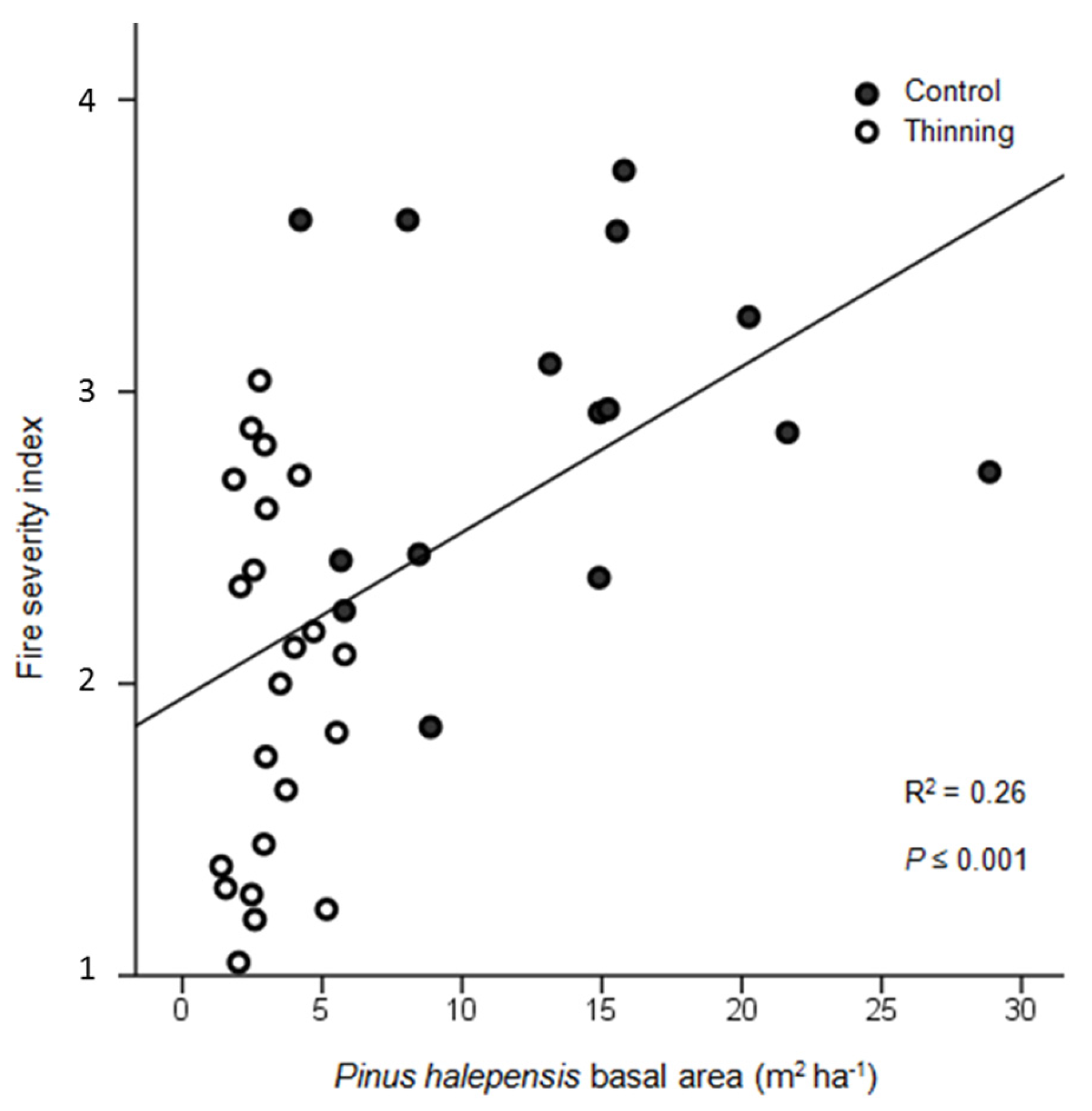
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Treatment | Elevation (m) | Aspect (°) | Slope (°) | Density (pines·ha−1) | Basal Area (m2·ha−1) | Height (m) | Seedling Density (seedlings·ha−1) |
|---|---|---|---|---|---|---|---|---|
| 1 | Control | 485 | 280 | 19 | 8315 | 5.67 | 3.5 ± 0.2 | 260 |
| 2 | Control | 504 | 305 | 14 | 1754 | 8.87 | 5.5 ± 0.1 | 3118 |
| 3 | Control | 480 | 30 | 26 | 2209 | 8.05 | 5.6 ± 0.0 | 10,134 |
| 4 | Control | 469 | 10 | 20 | 5587 | 21.63 | 8.5 ± 0.1 | 7535 |
| 5 | Control | 503 | 225 | 14 | 10,654 | 20.25 | 6.7 ± 0.1 | 1559 |
| 6 | Control | 513 | 20 | 28 | 13,252 | 14.90 | 4.7 ± 0.0 | 2079 |
| 7 | Control | 501 | 215 | 22 | 16,630 | 15.79 | 4.5 ± 0.1 | 1039 |
| 8 | Control | 480 | 300 | 15 | 15,591 | 28.86 | 5.9 ± 0.1 | 6496 |
| 9 | Control | 479 | 310 | 16 | 18189 | 15.54 | 5.0 ± 0.0 | 2598 |
| 10 | Control | 523 | 90 | 19 | 1299 | 5.79 | 4.1 ± 0.0 | 1039 |
| 11 | Control | 551 | 15 | 18 | 7276 | 14.92 | 4.2 ± 0.0 | 0 |
| 12 | Control | 506 | 230 | 25 | 6236 | 8.46 | 4.0 ± 0.1 | 260 |
| 13 | Control | 466 | 290 | 23 | 5457 | 13.14 | 5.0 ± 0.0 | 1299 |
| 14 | Control | 461 | 230 | 18 | 3313 | 4.22 | 5.0 ± 0.0 | 780 |
| 15 | Control | 463 | 255 | 26 | 5457 | 15.21 | 5.7 ± 0.1 | 520 |
| 16 | Thinned | 444 | 15 | 29 | 715 | 2.94 | 3.7 ± 0.1 | 1819 |
| 17 | Thinned | 482 | 184 | 23 | 585 | 2.55 | 4.9 ± 0.1 | 0 |
| 18 | Thinned | 487 | 50 | 27 | 1039 | 1.39 | 3.3 ± 0.1 | 780 |
| 19 | Thinned | 232 | 90 | 26 | 780 | 4.01 | 5.1 ± 0.1 | 1039 |
| 20 | Thinned | 482 | 55 | 27 | 585 | 2.48 | 5.7 ± 0.1 | 6236 |
| 21 | Thinned | 473 | 105 | 24 | 650 | 1.55 | 3.7 ± 0.1 | 0 |
| 22 | Thinned | 482 | 50 | 26 | 520 | 3.50 | 7.0 ± 0.1 | 260 |
| 23 | Thinned | 482 | 60 | 27 | 715 | 5.16 | 7.0 ± 0.1 | 0 |
| 24 | Thinned | 473 | 5 | 26 | 974 | 5.80 | 6.4 ± 0.1 | 1039 |
| 25 | Thinned | 500 | 95 | 22 | 650 | 1.85 | 5.3 ± 0.2 | 0 |
| 26 | Thinned | 505 | 80 | 28 | 520 | 2.99 | 5.7 ± 0.1 | 520 |
| 27 | Thinned | 497 | 15 | 28 | 585 | 2.07 | 4.8 ± 0.1 | 260 |
| 28 | Thinned | 483 | 240 | 30 | 715 | 3.71 | 4.2 ± 0.1 | 260 |
| 29 | Thinned | 457 | 120 | 23 | 780 | 2.47 | 3.9 ± 0.1 | 260 |
| 30 | Thinned | 453 | 80 | 16 | 909 | 4.69 | 6.0 ± 0.1 | 520 |
| 31 | Thinned | 468 | 315 | 10 | 650 | 2.92 | 4.7 ± 0.06 | 0 |
| 32 | Thinned | 503 | 75 | 14 | 844 | 2.76 | 3.7 ± 0.1 | 0 |
| 33 | Thinned | 525 | 305 | 18 | 974 | 3.01 | 5.3 ± 0.1 | 260 |
| 34 | Thinned | 466 | 150 | 16 | 455 | 4.18 | 7.6 ± 0.1 | 0 |
| 35 | Thinned | 473 | 160 | 14 | 390 | 5.52 | 7.7 ± 0.1 | 260 |
| 36 | Thinned | 482 | 350 | 25 | 715 | 2.01 | 5.2 ± 0.2 | 2339 |
| 37 | Thinned | 459 | 15 | 20 | 844 | 2.59 | 4.2 ± 0.1 | 260 |
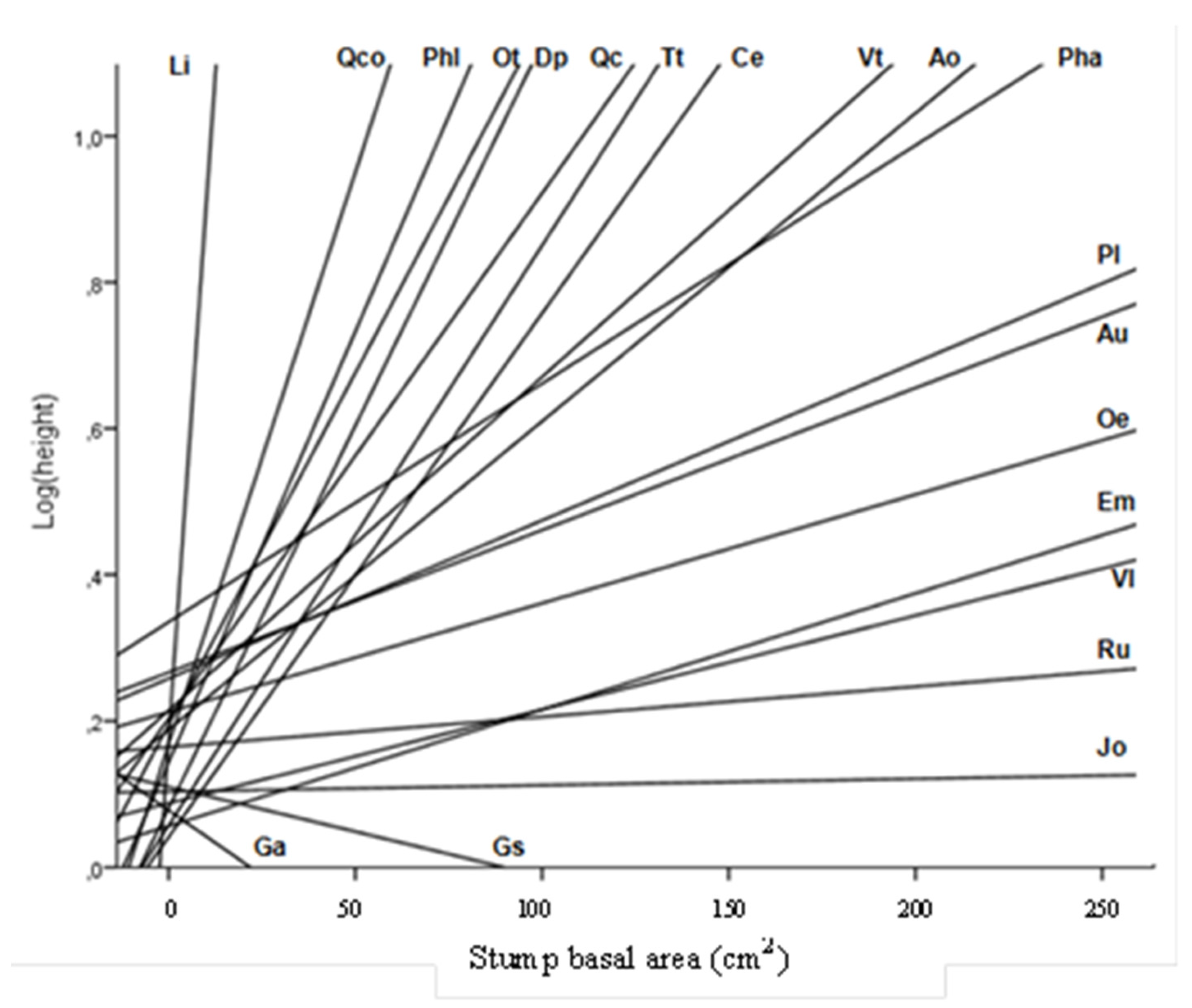
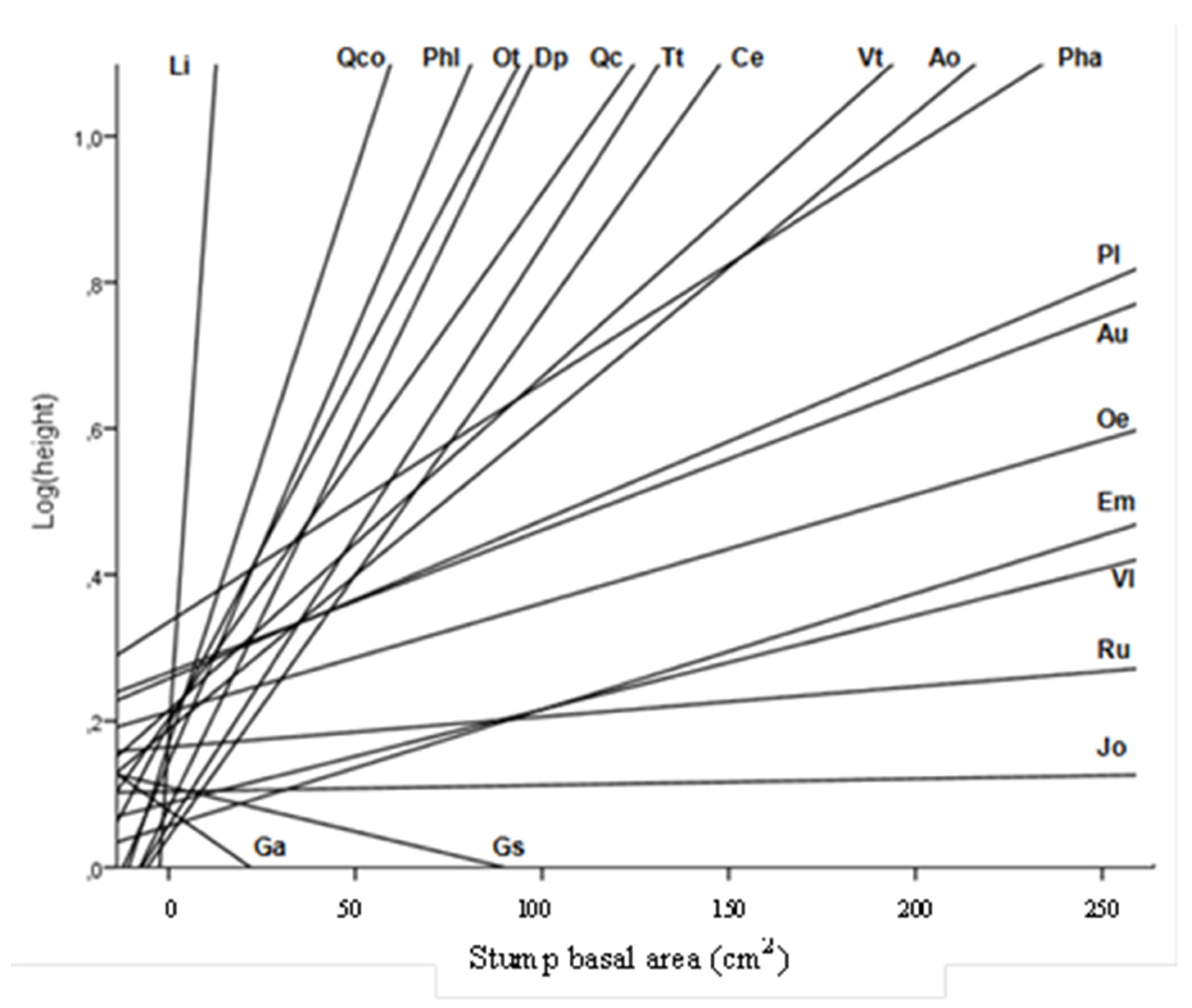
| Species | n | Total Height (cm) | Cover (cm2) | Biovolume (dm3) |
|---|---|---|---|---|
| A. ovalis | 11 | 37 ± 1.0 | 4149 ± 239 | 301 ± 18.40 |
| A. unedo | 30 | 46 ± 0.8 | 352 ± 111 | 231 ± 8.04 |
| C. emerus | 14 | 5 ± 0.1 | 73 ± 5 | 0.5 ± 0.04 |
| D. pentaphyllum | 20 | 11 ± 0.2 | 251 ± 6 | 3.48 ± 0.13 |
| E. multifolora | 234 | 7 ± 0.1 | 76 ± 3 | 0.69 ± 0.03 |
| G. alypum | 29 | 7 ± 0.1 | 64 ± 2 | 0.59 ± 0.03 |
| G. scorpius | 164 | 12 ± 0.2 | 139 ± 5 | 2.28 ± 0.16 |
| J. oxycedrus | 22 | 11 ± 0.1 | 153 ± 5 | 2.03 ± 0.08 |
| L. implexa | 61 | 28 ± 0.8 | 445 ± 21 | 22.94 ± 2.25 |
| O. europaea | 13 | 26 ± 0.4 | 503 ± 15 | 16.50 ± 0.57 |
| P. angustifolia | 12 | 28 ± 0.6 | 640 ± 32 | 33.80 ± 2.61 |
| P. latifolia | 21 | 51 ± 0.9 | 4718 ± 222 | 396.36 ± 23.07 |
| P. lentiscus | 75 | 22 ± 0.4 | 392 ± 18 | 14.28 ± 0.95 |
| Q. cerrioides | 29 | 34 ± 0.4 | 2118 ± 67 | 90.13 ± 3.47 |
| Q. coccifera | 87 | 29 ± 0.7 | 482 ± 21 | 23.29 ± 1.46 |
| R. ulmifolius | 56 | 22 ± 0.4 | 596 ± 30 | 22.70 ± 1.59 |
| T. tinctoria | 18 | 17 ± 0.4 | 1370 ± 82 | 42.12 ± 3.14 |
| V. lantana | 28 | 8 ± 0.1 | 89 ± 3 | 1.07 ± 0.05 |
| V. tinus | 61 | 10 ± 0.2 | 65 ± 3 | 0.92 ± 0.06 |
References
- Pausas, J.G. Changes in fire and climate in the eastern Iberian Peninsula (Mediterranean Basin). Clim. Chang. 2004, 63, 337–350. [Google Scholar] [CrossRef]
- Buhk, C.; Meyn, A.; Jentsch, A. The challenge of plant regeneration after fire in the Mediterranean Basin: scientific gaps in our knowledge on plant strategies and evolution of traits. Plant Ecol. 2007, 192, 1–19. [Google Scholar] [CrossRef]
- De Las Heras, J.; Moya, D.; Vega, J.A.; Daskalakou, E.; Vallejo, R.; Grigoriadis, N.; Tsitsoni, T.; Baeza, J.; Valdecantos, A.; Fernández, C.; et al. Post-Fire management of serotinous pine forests. In Post-Fire Management and Restoration of Southern European Forests; Moreira, F., Arianoutsou, M., Corona, P., de las Heras, J., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 24, pp. 121–150. [Google Scholar]
- Hanes, T.L. Succession after fire in the chaparral of southern California. Ecol. Monogr. 1971, 41, 27–52. [Google Scholar] [CrossRef]
- Rodrigo, A.; Retana, J.; Picó, F.X. Direct regeneration is not the only response of Mediterranean forests to large fires. Ecology 2004, 85, 716–729. [Google Scholar] [CrossRef]
- Calvo, A.; Cerdà, A. An example of the changes in the hydrological and erosional response of soil after a forest fire, Pedralba (Valencia), Spain. In Soil Erosion as a Consequence of Forest Fires; Sala, M., Rubio, J.L., Eds.; Geoforma Ediciones: Logroño, Spain, 1994; pp. 99–110. [Google Scholar]
- De las Heras, J.; Moya, D.; Lopez-Serrano, F.R.; Rubio, E. Carbon sequestration of naturally regenerated Aleppo pine stands in response to early thinning. New For. 2013, 44, 457–470. [Google Scholar] [CrossRef]
- Alfaro-Sanchez, R.; Lopez-Serrano, F.R.; Rubio, E.; Moya, D.; Sanchez-Salguero, R.; de las Heras, J. Biomass storage in low timber productivity Mediterranean forests managed after natural post-fire regeneration in south-eastern Spain. Eur. J. For. Res. 2014, 133, 793–807. [Google Scholar] [CrossRef]
- Verkaik, I.; Espelta, J.M. Post-Fire regeneration thinning, cone production, serotiny and regeneration age in Pinus halepensis. For. Ecol. Manag. 2006, 231, 155–163. [Google Scholar] [CrossRef]
- Moya, D.; Espelta, J.M.; Verkaik, I.; López-Serrano, F.; de Las Heras, J. Tree density and site quality influence on Pinus halepensis Mill. reproductive characteristics after large fires. Ann. For. Sci. 2007, 64, 649–656. [Google Scholar] [CrossRef]
- Zedler, P.H. Fire frequency in southern California shrublands: Biological effects and management options. In Bushfires in California Wildlands: Ecology and Resource Management; Keeley, J.E., Scott, T., Eds.; International Association of Wildland Fire: Fairfield, CA, USA, 1995; pp. 101–112. [Google Scholar]
- Ne’eman, G.; Goubitz, S.; Nathan, R. Reproductive traits of Pinus halepensis in the light of fire—A critical review. Plant Ecol. 2004, 171, 69–79. [Google Scholar] [CrossRef]
- Alfaro-Sanchez, R.; Lopez-Serrano, F.R.; Rubio, E.; Sanchez-Salguero, R.; Moya, D.; Hernandez-Tecles, E.; de las Heras, J. Response of biomass allocation patterns to thinning in Pinus halepensis differs under dry and semiarid Mediterranean climates. Ann. For. Sci. 2015, 72, 595–607. [Google Scholar] [CrossRef]
- Alfaro-Sanchez, R.; Camarero, J.J.; Lopez-Serrano, F.R.; Sanchez-Salguero, R.; Moya, D.; de las Heras, J. Positive coupling between growth and reproduction in young post-fire Aleppo pines depends on climate and site conditions. Int. J. Wildland Fire 2015, 24, 507–517. [Google Scholar] [CrossRef]
- Francos, M.; Ubeda, X.; Tort, J.; Panareda, J.M.; Cerdà, A. The role of forest fire severity on vegetation recovery after 18 years. Implications for forest management of Quercus suber L. in Iberian Peninsula. Glob. Planet. Chang. 2016, 145, 11–16. [Google Scholar] [CrossRef]
- González-Ochoa, A.I.; López-Serrano, F.R.; de Las Heras, J. Does post-fire forest management increase tree growth and cone production in Pinus halepensis? For. Ecol. Manag. 2004, 188, 235–247. [Google Scholar] [CrossRef]
- Moya, D.; de las Heras, J.; López-Serrano, F.R.; Leone, V. Optimal intensity and age of management in young aleppo pine stands for post-fire resilience. For. Ecol. Manag. 2008, 255, 3270–3280. [Google Scholar] [CrossRef]
- Alfaro-Sanchez, R.; Sanchez-Salguero, R.; de las Heras, J.; Hernández-Tecles, E.; Moya, D.; Lopez-Serrano, F.R. Vegetation dynamics of managed Mediterranean forests 16 year after large fires in southeastern Spain. Appl. Veg. Sci. 2015, 18, 272–282. [Google Scholar] [CrossRef]
- Palmero-Iniesta, M. Fire Behavior in Pinus Halepensis Thickets: Effects of Thinning and Woody Debris Decomposition in Different Climatic Scenarios. Master’s Thesis, Master en Incendios Forestales: Ciencia y gestión integral, Universitat de Lleida, Lleida, Spain, September 2016. [Google Scholar]
- Domènech-Jardí, R.; Pastor-Ferrer, E.; Agueda-Costafreda, A.; Sans, A.; Navascués, P.; Planas-Cuchi, E. Modelos de combustible para caracterizar el comportamiento de los incendios en regenerados clareados de pino carrasco. Montes 2013, 115, 22–29. [Google Scholar]
- Fulé, P.Z.; Waltz, A.E.M.; Covington, W.W.; Heinlein, T.A. Measuring forest restoration effectiveness in reducing hazardous fuels. J. For. 2001, 99, 24–29. [Google Scholar]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Brown, R.T.; Agee, J.K.; Franklin, J.F. Forest restoration and fire: Principles in the context of place. Conserv. Biol. 2004, 18, 903–912. [Google Scholar] [CrossRef]
- Pausas, J.G.; Ribeiro, E.; Vallejo, R.V. Post-Fire regeneration variability of Pinus halepensis in the eastern Iberian Peninsula. For. Ecol. Manag. 2004, 203, 251–259. [Google Scholar] [CrossRef]
- Eugenio, M.; Verkaik, I.; Lloret, F.; Espelta, J.M. Recruitment and growth decline in Pinus halepensis populations after recurrent wildfires in Catalonia (NE Iberian Peninsula). For. Ecol. Manag. 2006, 231, 47–54. [Google Scholar]
- Turner, M.G.; Romme, W.H.; Gardner, R.H. Prefire heterogeneity, fire severity, and early postfire plant reestablishment in subalpine forests of Yellowstone National Park, Wyoming. Int. J. Wildland Fire 1999, 9, 21–36. [Google Scholar] [CrossRef]
- Pausas, J.G.; Ouadah, N.; Ferran, A.; Gimeno, T.; Vallejo, R.V. Fire severity and seedling establishment in Pinus halepensis woodlands, eastern Iberian Peninsula. Plant Ecol. 2003, 169, 205–213. [Google Scholar] [CrossRef]
- Schwartz, N.B.; Urban, D.L.; White, P.S.; Moody, A.; Klein, R.N. Vegetation dynamics vary across topographic and fire severity gradients following prescribed burning in Great Smoky Mountains National Park. For. Ecol. Manag. 2016, 365, 1–11. [Google Scholar] [CrossRef]
- Ninyerola, M.; Pons, X.; Roure, J.M. A methodological approach of climatological modelling of air temperature and precipitation through GIS techniques. Int. J. Climatol. 2000, 20, 1823–1841. [Google Scholar] [CrossRef]
- Panareda-Clopés, J.M.; Nuet-Badia, J. Tipología y cartografía corológica de las plantas vasculares de Montserrat (Cordillera Prelitoral Catalana). Rev. Geogr. 1993, 27, 33–58. [Google Scholar]
- Panetsos, K.P. Monograph of Pinus halepensis Mill. and P. brutia Ten. Ann. For. Zagreb. 1981, 9, 39–77. [Google Scholar]
- Llobet, S. Propuesta de modelo silvícola para montes de pino carrasco (Pinus halepensis). In Modelos Silvícolas en Bosques Privados Mediterraneos; Colección Documentos de Trabajo, Serie_Territorio 5; Diputació de Barcelona: Barcelona, Spain, 2008; pp. 109–145. [Google Scholar]
- Omi, P.N.; Kalabokidis, K.D. Fire damage on extensively vs. intensively managed forest stands within the North Fork Fire, 1988. Northwest Sci. 1991, 65, 149–157. [Google Scholar]
- Weatherspoon, C.P.; Skinner, C.N. An assessment of factors associated with damage to tree crowns from the 1987 wildfires in northern California. For. Sci. 1995, 41, 430–451. [Google Scholar]
- Habrouk, A.; Retana, J.; Espelta, J.M. Role of heat tolerance and cone protection of seeds in the response of three pine species to wildfires. Plant Ecol. 1999, 145, 91–99. [Google Scholar] [CrossRef]
- Espelta, J.M.; Retana, J.; Habrouk, A. Resprouting patterns after fire and response to stool cleaning of two coexisting Mediterranean oaks with contrasting leaf habits on two different sites. For. Ecol. Manag. 2003, 179, 401–414. [Google Scholar] [CrossRef]
- Espelta, J.M.; Barbati, A.; Quevedo, L.; Tárrega, R.; Navascués, P.; Bonfil, C.; Peguero, G.; Fernández-Martínez, M.; Rodrigo, A. Post-Fire management of Mediterranean broadleaved forests. In Post-Fire Management and Restoration of Southern European Forests; Moreira, F., Arianoutsou, M., Corona, P., de las Heras, J., Eds.; Springer: Dordrecht, The Netherlands, 2012; Volume 24, pp. 171–194. [Google Scholar]
- Quevedo, L.; Rodrigo, A.; Espelta, J.M. Post-Fire resprouting ability of 15 non-dominant shrub and tree species in Mediterranean areas of NE Spain. Ann. For. Sci. 2007, 64, 883–890. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R Package Version 1.15.1. 2015. Available online: http://CRAN.R-project.org/package=MuMIn (accessed on 23 March 2016).
- Pollet, J.; Omi, P.N. Effect of thinning and prescribed burning on crown fire severity in ponderosa pine forests. Int. J. Wildland Fire 2002, 11, 1–10. [Google Scholar] [CrossRef]
- Carey, H.; Schumann, M. Modifying wildfire behavior—The effectiveness of fuel treatments: The status of our knowledge. 2003; in press. [Google Scholar]
- Anderson, H.E. Aids to Determining Fuel Models for Estimating Fire Behavior; USDA Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1982.
- Kalabokidis, K.D.; Omi, P.N. Reduction of fire hazard through thinning/residue disposal in the urban interface. Int. J. Wildland Fire. 1998, 8, 29–35. [Google Scholar] [CrossRef]
- Ordóñez, J.L.; Retana, J.; Espelta, J.M. Effects of tree size, crown damage, and tree location on post-fire survival and cone production of Pinus nigra trees. For. Ecol. Manag. 2005, 206, 109–117. [Google Scholar] [CrossRef]
- Herranz, J.; Martínez-Sánchez, J.J.; Martín, A.; Ferrandis, P. Postfire regeneration of Pinus halepensis Miller in a semi-arid area in Albacete province (southeastern Spain). Écoscience 1997, 4, 86–90. [Google Scholar] [CrossRef]
- Tsitsoni, T. Conditions determining natural regeneration after wildfires in the Pinus halepensis (Miller, 1768) forests of Kassandra Peninsula (North Greece). For. Ecol. Manag. 1997, 92, 199–208. [Google Scholar] [CrossRef]
- Castro, J.; Allen, C.D.; Molina-Morales, M.; Marañon-Jimenez, S.; Sanchez-Miranda, A.; Zamora, R. Salvage Logging Versus the Use of Burnt Wood as a Nurse Object to Promote Post-Fire Tree Seedling Establishment. For. Ecol. Manag. 2011, 19, 537–544. [Google Scholar] [CrossRef]
- Fernandez, C.; Vega, J.A.; Fonturbel, T.; Jimenez, E.; Perez-Gorostiaga, P. Effects of wildfire, salvage logging and slash manipulation on Pinus pinaster Ait. recruitment in Orense (NW Spain). For. Ecol. Manag. 2008, 255, 1294–1304. [Google Scholar] [CrossRef]
- Bonfil, P.; Cortes, P.; Espelta, J.M.; Retana, J. The role of disturbance in the co-existence of the evergreen Quercus ilex and the deciduous Quercus cerrioides. J. Veg. Sci. 2004, 15, 423–430. [Google Scholar] [CrossRef]
- Klimešová, J.; Klimeš, L. Bud banks and their role in vegetative regeneration—A literature review and proposal for simple classification and assessment. Perspect. Plant Ecol. Evol. Syst. 2007, 8, 115–129. [Google Scholar] [CrossRef]
- Peguero, G.; Espelta, J.M. Disturbance intensity and seasonality affect the resprouting ability of the neotropical dry-forest tree Acacia pennatula: Do resources stored below-ground matter? J. Trop. Ecol. 2011, 27, 539–546. [Google Scholar] [CrossRef]
- Manes, F.; Capogna, F.; Puppi, G.; Vitele, M. Ecophysiological characterisation of Phillyrea angustifolia L. and response of resprouts to different fire disturbance intensities. In Fire and Biological Processes; Trabaud, L., Prodon, R., Eds.; Backhuys publishers: Leiden, The Netherlands, 2002; pp. 115–130. [Google Scholar]
- Capogna, F.; Persiani, A.M.; Maggi, O.; Dowgiallo, G.; Puppi, G.; Manes, F. Effects of different fire intensities on chemical and biological soil components and related feedbacks on a Mediterranean shrub (Phillyrea angustifolia L.). Plant Ecol. 2009, 204, 155–171. [Google Scholar] [CrossRef]
- Vilà, M.; Terradas, J. Effects of nutrient availability and neighbours on shoot growth, resprouting and flowering of Erica multiflora. J. Veg. Sci. 1995, 6, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Cotillas, M.; Espelta, J.M.; Sánchez-Costa, E.; Sabaté, S. Aboveground and belowground biomass allocation patterns in two Mediterranean oaks with contrasting leaf habit: An insight into carbon stock in young oak coppices. Eur. J. For. Res. 2016, 135, 243–252. [Google Scholar] [CrossRef]
- Espelta, J.M.; Verkaik, I.; Eugenio, M.; Lloret, F. Recurrent wildfires constrain long-term reproduction ability in Pinus halepensis Mill. Int. J. Wildland Fire 2008, 17, 579–585. [Google Scholar] [CrossRef]
- Espelta, J.M.; Arnan, X.; Rodrigo, A. Non-Fire induced seed release in a weakly serotinous pine: Climatic factors, maintenance costs or both? Oikos 2011, 11, 1752–1760. [Google Scholar] [CrossRef]
- Graham, R.T.; Harvey, A.E.; Jain, T.B.; Tonn, J.R. The Effects of Thinning and Similar Stand Treatments on Fire Behavior in Western Forests; General Technical Report PNW-GTR-463; USDA Forest Service, Pacific Northwest Research Station: Seattle, WA, USA, 1999.



| Characteristics | Total Height | Cover | Biovolume | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| df | F | p | R2 | df | F | p | R2 | df | F | p | R2 | |
| Severity (Sev) | 720.9 | 5.49 | 0.019 | 0.58 m 0.60 c | 709.7 | 7.83 | 0.005 | 0.53 m 0.57 c | 714.5 | 7.99 | 0.005 | 0.57 m 0.60 c |
| Species (Sp) | 843.6 | 4.27 | ≤0.001 | 878.6 | 4.53 | ≤0.001 | 873.2 | 4.57 | ≤0.001 | |||
| Stump n. (Sn) | 930.9 | <0.01 | 0.937 | 930.6 | 0.55 | 0.458 | 930.8 | 0.27 | 0.605 | |||
| Stump s. (Ss) | 929.8 | 16.53 | ≤0.001 | 931.0 | 26.65 | ≤0.001 | 931.0 | 26.28 | ≤0.001 | |||
| Sp x Sev | 769.3 | 1.80 | 0.019 | 831.8 | 1.55 | 0.061 | 821.9 | 1.58 | 0.053 | |||
| Sp x Sn | 926.2 | 1.34 | 0.150 | 926.2 | 1.01 | 0.449 | 926.3 | 1.10 | 0.342 | |||
| Sp x Ss | 927.8 | 3.18 | ≤0.001 | 927.5 | 3.12 | ≤0.001 | 927.6 | 3.37 | ≤0.001 | |||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García‐Jiménez, R.; Palmero‐Iniesta, M.; Espelta, J.M. Contrasting Effects of Fire Severity on the Regeneration of Pinus halepensis Mill. and Resprouter Species in Recently Thinned Thickets. Forests 2017, 8, 55. https://doi.org/10.3390/f8030055
García‐Jiménez R, Palmero‐Iniesta M, Espelta JM. Contrasting Effects of Fire Severity on the Regeneration of Pinus halepensis Mill. and Resprouter Species in Recently Thinned Thickets. Forests. 2017; 8(3):55. https://doi.org/10.3390/f8030055
Chicago/Turabian StyleGarcía‐Jiménez, Ruth, Marina Palmero‐Iniesta, and Josep Maria Espelta. 2017. "Contrasting Effects of Fire Severity on the Regeneration of Pinus halepensis Mill. and Resprouter Species in Recently Thinned Thickets" Forests 8, no. 3: 55. https://doi.org/10.3390/f8030055
APA StyleGarcía‐Jiménez, R., Palmero‐Iniesta, M., & Espelta, J. M. (2017). Contrasting Effects of Fire Severity on the Regeneration of Pinus halepensis Mill. and Resprouter Species in Recently Thinned Thickets. Forests, 8(3), 55. https://doi.org/10.3390/f8030055