Low Light Availability Associated with American Beech Is the Main Factor for Reduced Sugar Maple Seedling Survival and Growth Rates in a Hardwood Forest of Southern Quebec

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Light Availability
2.4. Planting Experiment
2.5. Foliage of Naturally Growing Seedling
2.6. Soils
2.7. Statistics
3. Results
3.1. Stand Characteristics and Light Environment
3.2. Soil Properties
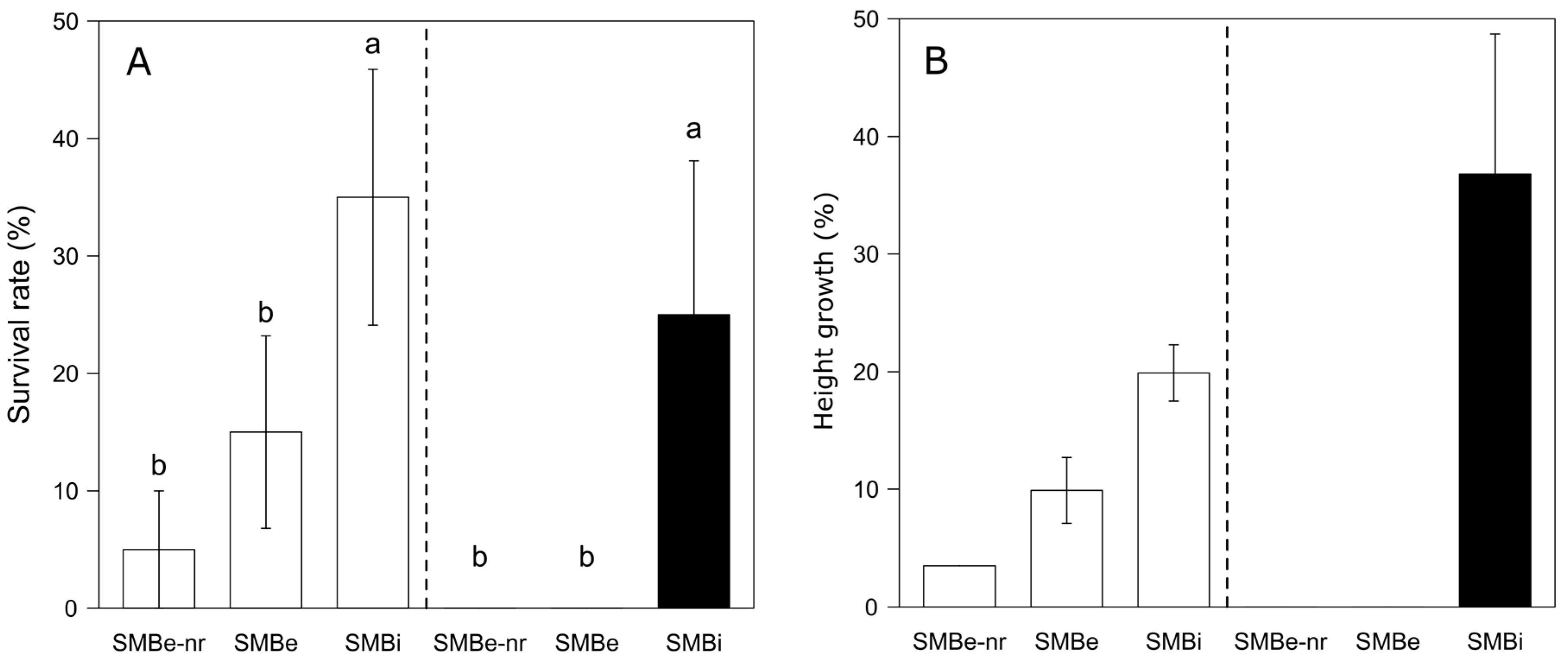
3.3. Survival and Growth of Planted Sugar Maple Seedlings
3.4. Foliar Nutrition of Natural Sugar Maple Seedlings
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Seymour, R.S. The northeastern region. In Regional Silviculture of the United States; Barrett, J.W., Ed.; Wiley and Sons: New York, NY, USA, 1995; Volume 3, pp. 31–79. [Google Scholar]
- Takahashi, K.; Arii, K.; Lechowicz, M.J. Codominance of Acer saccharum and Fagus grandifolia: The role of Fagus root sprouts along a slope gradient in an old-growth forest. J. Plant Res. 2010, 123, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Fain, J.J.; Volk, T.A.; Fahey, T.J. Fifty years of change in an upland forest in south-central New York: General patterns. Bull. Torrey Bot. Club 1994, 121, 130–139. [Google Scholar] [CrossRef]
- Brisson, J.; Bergeron, Y.; Bouchard, A.; Leduc, A. Beech-maple dynamics in an old-growth forest in southern Québec, Canada. Écoscience 1994, 1, 40–46. [Google Scholar] [CrossRef]
- Poulson, T.L.; Platt, W.J. Replacement patterns of beech and sugar maple in Warren Woods, Michigan. Ecology 1996, 77, 1234–1253. [Google Scholar] [CrossRef]
- Foré, S.A.; Vankat, J.L.; Schaefer, R.L. Temporal variation in the woody understory of an old-growth Fagus-Acer forest and implications for overstory recruitment. J. Veg. Sci. 1997, 8, 607–614. [Google Scholar] [CrossRef]
- Duchesne, L.; Ouimet, R.; Moore, J.-D.; Paquin, R. Changes in structure and composition of maple-beech stands following sugar maple decline in Québec, Canada. For. Ecol. Manag. 2005, 208, 223–236. [Google Scholar] [CrossRef]
- Beaudet, M.; Brisson, J.; Gravel, D.; Messier, C. Effect of a major canopy disturbance on the coexistence of Acer saccharum and Fagus grandifolia in the understorey of an old-growth forest. J. Ecol. 2007, 95, 458–467. [Google Scholar] [CrossRef]
- Gravel, D.; Beaudet, M.; Messier, C. Sapling age structure and growth series reveal a shift in recruitment dynamics of sugar maple and American beech over the last 40 years. Can. J. For. Res. 2011, 41, 873–880. [Google Scholar] [CrossRef]
- Jenkins, J. Hardwood Regeneration Failure in the Adirondacks; Wildlife Conservation Society Working Paper No. 9; Wildlife Conservation Society: Bronx, NY, USA, 1997. [Google Scholar]
- Beaudet, M.; Messier, C.; Paré, D.; Brisson, J.; Bergeron, Y. Possible mechanisms of sugar maple regeneration failure and replacement by beech in the Boisé-des-Muir old-growth forest, Québec. Écoscience 1999, 6, 264–271. [Google Scholar] [CrossRef]
- Hane, E.N. Indirect effects of beech bark disease on sugar maple seedling survival. Can. J. For. Res. 2003, 33, 807–813. [Google Scholar] [CrossRef]
- Canham, C.D. Growth and canopy architecture of shade-tolerant trees: Response to canopy gaps. Ecology 1988, 69, 786–795. [Google Scholar] [CrossRef]
- Beaudet, M.; Messier, C. Growth and morphological responses of yellow birch, sugar maple, and beech seedlings growing under a natural light gradient. Can. J. For. Res. 1998, 28, 1007–1015. [Google Scholar] [CrossRef]
- Kobe, R.K.; Pacala, S.W.; Silander, J.A.; Canham, C.D. Juvenile tree survivorship as a component of shade tolerance. Ecol. Appl. 1995, 5, 517–532. [Google Scholar] [CrossRef]
- Messier, C.; Nikinmaa, E. Effects of light availability and sapling size on the growth, biomass allocation, and crown morphology of understory sugar maple, yellow birch, and beech. Écoscience 2000, 7, 345–356. [Google Scholar] [CrossRef]
- Canham, C.D. Different respones to gaps among shade-tollerant tree species. Ecology 1989, 70, 548–550. [Google Scholar] [CrossRef]
- Nolet, P.; Delagrange, S.; Bouffard, D.; Doyon, F.; Forget, E. The successional status of sugar maple (Acer saccharum), revisited. Ann. For. Sci. 2008, 65, 208. [Google Scholar] [CrossRef]
- Takahashi, K.; Lechowicz, M.J. Do interspecific differences in sapling growth traits contribute to the co-dominance of Acer saccharum and Fagus grandifolia? Ann. Bot. 2008, 101, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Arii, K.; Lechowicz, M.J. The influence of overstory trees and abiotic factors on the sapling community in an old-growth Fagus-Acer forest. Ecoscience 2002, 9, 386–396. [Google Scholar] [CrossRef]
- Duchesne, L.; Ouimet, R. Present-day expansion of American beech in northeastern hardwood forests: Does soil base status matter? Can. J. For. Res. 2009, 39, 2273–2282. [Google Scholar] [CrossRef]
- Kobe, R.K.; Likens, G.E.; Eagar, C. Tree seedling growth and mortality responses to manipulations of calcium and aluminum in a northern hardwood forest. Can. J. For. Res. 2002, 32, 954–966. [Google Scholar] [CrossRef]
- Halman, J.M.; Schaberg, P.G.; Hawley, G.J.; Hansen, C.F.; Fahey, T.J. Differential impacts of calcium and aluminum treatments on sugar maple and american beech growth dynamics. Can. J. For. Res. 2014, 45, 52–59. [Google Scholar] [CrossRef]
- Duchesne, L.; Ouimet, R.; Houle, D. Basal area growth of sugar maple in relation to acid deposition, stand health, and soil nutrients. J. Environ. Qual. 2002, 31, 1676–1683. [Google Scholar] [CrossRef] [PubMed]
- St. Clair, S.B.; Sharpe, W.E.; Lynch, J.P. Key interactions between nutrient limitation and climatic factors in temperate forests: A synthesis of the sugar maple literature. Can. J. For. Res. 2008, 38, 401–414. [Google Scholar] [CrossRef]
- Long, R.P.; Horsley, S.B.; Hallett, R.A.; Bailey, S.W. Sugar maple growth in relation to nutrition and stress in the northeastern United States. Ecol. Appl. 2009, 19, 1454–1466. [Google Scholar] [CrossRef] [PubMed]
- Côté, B.; Fyles, J.W. Nutrient concentration and acid-base status of leaf litter of tree species characteristic of the hardwood forest of southern Quebec. Can. J. For. Res. 1994, 24, 192–196. [Google Scholar] [CrossRef]
- Hane, E.N.; Hamburg, S.P.; Barber, A.L.; Plaut, J.A. Phytotoxicity of american beech leaf leachate to sugar maple seedlings in a greenhouse experiment. Can. J. For. Res. 2003, 33, 814–821. [Google Scholar] [CrossRef]
- Beaudet, M.; Messier, C. Beech regeneration of seed and root sucker origin: A comparison of morphology, growth, survival, and response to defoliation. For. Ecol. Manag. 2008, 255, 3659–3666. [Google Scholar] [CrossRef]
- Cale, J.A.; Garrison-Johnston, M.T.; Teale, S.A.; Castello, J.D. Beech bark disease in north America: Over a century of research revisited. For. Ecol. Manag. 2017, 394, 86–103. [Google Scholar] [CrossRef]
- Marquis, D.A.; Brenneman, R. The Impact of Deer on Forest Vegetation in Pennsylvania; Northeastern Forest Experimental Station: Broomall, PA, USA, 1981; p. 7. [Google Scholar]
- Long, Z.T.; Pendergast, T.H.; Carson, W.P. The impact of deer on relationships between tree growth and mortality in an old-growth beech-maple forest. For. Ecol. Manag. 2007, 252, 230–238. [Google Scholar] [CrossRef]
- Bose, A.K.; Weiskittel, A.; Wagner, R.G. Occurrence, pattern of change, and factors associated with american beech-dominance in stands of the northeastern USA forest. For. Ecol. Manag. 2017, 392, 202–212. [Google Scholar] [CrossRef]
- Saucier, J.; Robitaille, A.; Grondin, P. Cadre bioclimatique du Québec. In Manuel de Foresterie, 2nd ed.; Écologie Forestière; Ordre Des Ingénieurs Forestiers Du Québec: Québec, QC, USA, 2009; pp. 186–205. [Google Scholar]
- Soil Classification Working Group. The Canadian System of Soil Classification, 3rd ed.; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 1998; p. 187.
- Collin, A.; Messier, C.; Kembel, S.W.; Bélanger, N. Can sugar maple establish into the boreal forest? Insights from seedlings under various canopies in southern Quebec. Ecosphere 2017, in press. [Google Scholar]
- Frazer, G.W.; Canham, C.; Lertzman, K. Gap Light Analyzer (GLA), Version 2.0; Imaging Software to Extract Canopy Structure and Gap Light Transmission Indices from True-Colour Fisheye Photographs, Users Manual and Program Documentation; Simon Fraser University: Burnaby, BC, Canada; The Institute of Ecosystem Studies: Millbrook, NY, USA, 1999. [Google Scholar]
- Minocha, R.; Martinez, G.; Lyons, B.; Long, S. Development of a standardized methodology for quantifying total chlorophyll and carotenoids from foliage of hardwood and conifer tree species. Can. J. For. Res. 2009, 39, 849–861. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Collin, A.; Messier, C.; Bélanger, N. Conifer presence may negatively affect sugar maple’s ability to migrate into the boreal forest through reduced foliar nutritional status. Ecosystems 2016, 20, 16. [Google Scholar] [CrossRef]
- Hangs, R.D.; Greer, K.J.; Sulewski, C.A. The effect of interspecific competition on conifer seedling growth and nitrogen availability measured using ion-exchange membranes. Can. J. For. Res. 2004, 34, 754–761. [Google Scholar] [CrossRef]
- Moukoumi, J.; Farrell, R.E.; Rees, K.J.C.; Hynes, R.K.; Bélanger, N. Intercropping Caragana arborescens with Salix miyabeana to satisfy nitrogen demand and maximize growth. Bioenergy Res. 2012, 5, 719–732. [Google Scholar] [CrossRef]
- Bilodeau-Gauthier, S.; Paré, D.; Messier, C.; Bélanger, N. Root production of hybrid poplars and nitrogen mineralization improve following mounding of boreal Podzols. Can. J. For. Res. 2013, 43, 1092–1103. [Google Scholar] [CrossRef]
- Morse, C.C.; Yevdokimov, I.V.; DeLuca, T.H. In Situ extraction of rhizosphere organic compounds from contrasting plant communities. Commun. Soil Sci. Plant Anal. 2000, 31, 725–742. [Google Scholar] [CrossRef]
- Graham, H. Modified prussian blue assay for total phenols. J. Agric. Food Chem. 1992, 40, 801–805. [Google Scholar] [CrossRef]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. Nlme: Linear and Nonlinear Mixed Effects Models; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [PubMed]
- Strobl, C.; Malley, J.; Tutz, G. An introduction to recursive partitioning: Rationale, application, and characteristics of classification and regression trees, bagging, and random forests. Psychol. Methods 2009, 14, 323. [Google Scholar] [CrossRef] [PubMed]
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.; Simpson, G.L.; Solymos, P.; Stevens, M.; Wagner, H. Vegan: Community Ecology Package; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Stenberg, P.; Linder, S.; Smolander, H.; Flower-Ellis, J. Performance of the LAI-2000 plant canopy analyzer in estimating leaf area index of some Scots pine stands. Tree Physiol. 1994, 14, 981–995. [Google Scholar] [CrossRef] [PubMed]
- Nolet, P.; Bouffard, D.; Doyon, F.; Delagrange, S. Relationship between canopy disturbance history and current sapling density of Fagus grandifolia and Acer saccharum in a northern hardwood landscape. Can. J. For. Res. 2008, 38, 216–225. [Google Scholar] [CrossRef]
- Yawney, H.W. The effects of four levels of shade in sugar maple seedling development. In Proceedings of the Fourth North American Forest Biology Workshop, Syracuse, NY, USA, 9–11 August 1976; pp. 189–190. [Google Scholar]
- Bannon, K.; Delagrange, S.; Bélanger, N.; Messier, C. American beech and sugar maple sapling relative abundance and growth are not modified by light availability following partial and total canopy disturbances. Can. J. For. Res. 2015, 45, 632–638. [Google Scholar] [CrossRef]
- Nolet, P.; Delagrange, S.; Bannon, K.; Messier, C.; Kneeshaw, D. Liming has a limited effect on sugar maple—American beech dynamics compared with beech sapling elimination and canopy opening. Can. J. For. Res. 2015, 45, 1376–1386. [Google Scholar] [CrossRef]
- Cleavitt, N.L.; Fahey, T.J.; Battles, J.J. Regeneration ecology of sugar maple (Acer saccharum): Seedling survival in relation to nutrition, site factors, and damage by insects and pathogens. Can. J. For. Res. 2011, 41, 235–244. [Google Scholar] [CrossRef]
- Souto, X.; González, L.; Reigosa, M. Allelopathy in forest environment in Galicia, NW Spain. Allelopath. J. 1995, 2, 67–78. [Google Scholar]
- Coldwell, B.; DeLong, W. Studies of the composition of deciduous forest tree leaves before and after partial decomposition. Sci. Agric. 1950, 30, 456–466. [Google Scholar]
- Smith, W.H. Character and significance of forest tree root exudates. Ecology 1976, 57, 324–331. [Google Scholar] [CrossRef]
- Cale, J.A.; McNulty, S.A.; Teale, S.A.; Castello, J.D. The impact of beech thickets on biodiversity. Biol. Invasions 2013, 15, 699–706. [Google Scholar] [CrossRef]
- Cleavitt, N.L.; Battles, J.J.; Fahey, T.J.; Blum, J.D. Determinants of survival over 7 years for a natural cohort of sugar maple seedlings in a northern hardwood forest. Can. J. For. Res. 2014, 44, 1112–1121. [Google Scholar] [CrossRef]
- Côté, S.D.; Rooney, T.P.; Tremblay, J.-P.; Dussault, C.; Waller, D.M. Ecological impacts of deer overabundance. Ann. Rev. Ecol. Evol. Syst. 2004, 35, 113–147. [Google Scholar] [CrossRef]
- Ohse, B.; Hammerbacher, A.; Seele, C.; Meldau, S.; Reichelt, M.; Ortmann, S.; Wirth, C. Salivary cues: Simulated roe deer browsing induces systemic changes in phytohormones and defence chemistry in wild-grown maple and beech saplings. Funct. Ecol. 2017, 31, 340–349. [Google Scholar] [CrossRef]


{kind=link}
| SMBe-nr | SMBe | SMBi | |
|---|---|---|---|
| Basal area (m2/ha) | 32.5 | 32.5 | 37 |
| Individual contribution to basal area by species (%) | |||
| Acer saccharum | 61.75 | 62.5 | 69 |
| Betula papyfera | 3.75 | 3.75 | 2.25 |
| Betula alleghaniensis | 0 | 0 | 22.5 |
| Fagus grandifolia | 32.5 | 31.75 | 4 |
| Abies balsamea | 2 | 2 | 2.25 |
| Forest Type | Canopy Openness % | LAI | Light Transmitted mol·m2·day−1 |
|---|---|---|---|
| SMBe-nr | 10.72 (±0.52) c | 3.14 (±0.15) a | 4.47 (±0.43) c |
| SMBe | 16.66 (±1.50) b | 2.29 (±0.11) b | 7.14 (±0.79) b |
| SMBi | 23.76 (±1.74) a | 1.93 (±0.11) b | 10.59 (±0.87) a |
| SMBe-nr | SMBe | SMBi | |
|---|---|---|---|
| FH horizon | |||
| pH | 4.50 (±0.16) | 4.55 (±0.20) | 4.47 (±0.10) |
| C/N | 19.0 (±1.51) | 20.7 (±1.41) | 22.8 (±1.51) |
| VWC (%) | 20.4 (±0.37) | 20.9 (±0.82) | 20.3 (±1.20) |
| Temperature (°C) † | 7.12 (±0.04) | 7.10 (±0.08) | 7.15 (±0.12) |
| Phenols (µg·g−1 soil) † | 176 (±54.6) a | 64.6 (±9.10) b | 63.5 (±3.15) b |
| B horizon | |||
| pH | 4.81 (±0.06) | 4.84 (±0.08) | 4.98 (±0.10) |
| Clay (%) | 3.13 (±0.44) | 3.17 (±0.78) | 2.20 (±0.41) |
| Silt (%) | 38.0 (±4.76) | 45.2 (±6.97) | 38.4 (±1.60) |
| Sand (%) | 8.7 (±5.03) | 51.6 (±7.73) | 59.4 (±1.96) |
| Soil Solution Ionic Activities (µg·10 cm2·2 month−1) | SMBe-nr | SMBe | SMBi |
|---|---|---|---|
| N † | 33.3 (±17.6) | 35.2 (±19.9) | 58.7 (±35.8) |
| P | 1.99 (±0.81) | 0.97 (±0.27) | 1.85 (±0.70) |
| K | 34.0 (±6.88) | 45.2 (±14.8) | 72.7 (±11.3) |
| Ca | 978 (±119) | 974 (±127) | 787 (±38.4) |
| Mg | 154 (±22.8) | 153 (±26.2) | 147 (±9.01) |
| Al | 43.3 (±4.92) | 47.9 (±8.27) | 97.3 (±27.6) |
| Mn | 7.80 (±2.78) | 8.87 (±0.87) | 13.1 (±4.75) |
| Fe | 6.87 (±0.85) | 5.00 (±1.72) | 7.07 (±1.47) |
| SMBe | SMBi | p-Value | |
|---|---|---|---|
| Foliar nutrients (mg·g−1) | |||
| C | 466 (±3.03) | 468 (±4.58) | 0.516 |
| N | 17.5 (±0.59) | 18.5 (±1.20) | 0.497 |
| P | 0.98 (±0.05) | 1.14 (±0.08) | 0.019 |
| K | 5.80 (±0.87) | 5.87 (±0.66) | 0.393 |
| Ca | 9.22 (±1.03) | 8.89 (±0.46) | 0.777 |
| Mg | 2.03 (±0.25) | 2.05 (±0.26) | 0.857 |
| Leaf pigments (mg·g−1 of Fresh Matter) | |||
| Chlorophyll a | 2.40 (±0.16) | 2.96 (±0.47) | 0.013 |
| Chlorophyll b | 1.47 (±0.11) | 1.68 (±0.23) | 0.015 |
| Total carotenoids | 0.52 (±0.03) | 0.63 (±0.09) | 0.012 |
| Leaf damage (% of total leaf area) | 3.79 (±0.44) | 1.85 (±0.34) | 0.041 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collin, A.; Messier, C.; Kembel, S.W.; Bélanger, N. Low Light Availability Associated with American Beech Is the Main Factor for Reduced Sugar Maple Seedling Survival and Growth Rates in a Hardwood Forest of Southern Quebec. Forests 2017, 8, 413. https://doi.org/10.3390/f8110413
Collin A, Messier C, Kembel SW, Bélanger N. Low Light Availability Associated with American Beech Is the Main Factor for Reduced Sugar Maple Seedling Survival and Growth Rates in a Hardwood Forest of Southern Quebec. Forests. 2017; 8(11):413. https://doi.org/10.3390/f8110413
Chicago/Turabian StyleCollin, Alexandre, Christian Messier, Steven W. Kembel, and Nicolas Bélanger. 2017. "Low Light Availability Associated with American Beech Is the Main Factor for Reduced Sugar Maple Seedling Survival and Growth Rates in a Hardwood Forest of Southern Quebec" Forests 8, no. 11: 413. https://doi.org/10.3390/f8110413
APA StyleCollin, A., Messier, C., Kembel, S. W., & Bélanger, N. (2017). Low Light Availability Associated with American Beech Is the Main Factor for Reduced Sugar Maple Seedling Survival and Growth Rates in a Hardwood Forest of Southern Quebec. Forests, 8(11), 413. https://doi.org/10.3390/f8110413