1. Introduction
Urban forests are characterized by unique soils and urban morphologies, heterogeneous vegetation structure and composition, and novel assemblages of native and exotic tree species [
1,
2,
3]. The spatial and temporal characteristics of urban forest structure are driven by biophysical factors such as topography, climate, biogeochemical cycles, and disturbances such as drought [
2,
4,
5]. They are also affected by socioeconomic factors such as management and planning regimes, people’s preferences, and socio-political budgets and directives [
6,
7,
8]. As all these drivers alter the structure and composition of an urban forest, so too are ecological processes affected and subsequently the provision of ecosystem services [
6]. A few studies have examined urban forests, their functions, and mortality with some examples from Mediterranean climate urban forests in California (United States, US), Spain, and Italy [
9,
10,
11,
12]. However, other than these few studies, there is little information on how and why urban forest structure in South America, the global south, and Mediterranean climates changes across space and time.
Urban forests are a key component in cities and provider of ecosystem services as they influence the well-being of urban inhabitants [
11,
13]. They are able to store and sequester carbon, regulate hydrologic cycles, ameliorate climate, remove air pollutants, provide habitat for fauna and space for recreation and spiritual enjoyment, among other services [
13,
14]. However, multi-scale economic and socio-ecological drivers can alter urban forest structure, thus understanding the spatio-temporal dynamics of urban forest structure and composition can provide insights on their management and planning [
15].
Tree planting preferences and management decisions by both private individuals and communities can influence urban forest cover and density, growth, mortality, and distribution [
6]. Similarly, species composition, age diversity, condition, site characteristics, and socio-ecological disturbances have an effect on growth of the urban forest [
2,
4,
7,
8], affecting several ecosystem processes, disservices and services. The type of land use and building density (
i.e., urban morphology) and socioeconomic status in particular, affect the structure and distribution of the urban forest, leading to changes in biomass with consequences for carbon storage and sequestration [
5,
6,
10,
12]. Growth, mortality, and regeneration in both urban and natural forests depend on environmental factors such as rainfall, temperature, and soil conditions; however, urban tree growth and biomass have been shown to be often greater in urban than natural landscapes [
16]. Other anthropogenic factors affecting growth and mortality include exposure to pollution, artificial irrigation, and vegetation management and maintenance practices [
7,
8,
16,
17].
Irrigation becomes extremely influential in Mediterranean semi-arid and arid cities where frequent droughts and water scarcity during growing periods can cause reduced growth, basal area, and biomass, and can increase mortality rates [
15]. These structural effects can be measured in growth rates, biomass, basal area, and mortality rates, and are key for analyzing temporal patterns of carbon sequestration and storage [
17], tree wood waste biomass [
5], and overall effects of climate on urban forest structure [
6]. Research on urban tree growth, biomass, and basal area is scarce and, to our knowledge, limited to a number of species mainly from North America [
10,
16,
18] and a few studies from other regions [
19,
20,
21]. Further information on drivers of urban tree growth and mortality can improve decision-making on tree selection, urban forest carbon accounting, and benefit estimates [
10,
17,
22,
23].
Temporal studies of urban forest ecosystems are not common and are mainly based on coarse resolution information derived from satellite imagery [
7,
24,
25,
26,
27]. Most research for monitoring the urban forest has been based on land use/land cover change analyses, with several examples from the US [
5,
7,
16], Europe [
24], and Asia [
25], with few examples from Latin America [
26] and Africa [
27]. Research based on inventories and field data are particularly scarce, aside from monitoring of planted street trees [
10,
20] and citation therein], and a few examples from the subtropics [
5,
8,
16,
28] and humid temperate areas of North America [
7] exist. However, most of these mentioned studies are located in temperate and industrialized regions, with a scarce few studies from elsewhere [
2,
15] such as those in South America and from cities in Mediterranean climates. The use of permanent urban forest plots for monitoring the structure and composition of the urban forest is increasingly being used for not only recognizing necessary changes in planning and management goals, but also for distinguishing the most effective practices that maximize the provision of ecosystem services [
4,
7]. Nevertheless, such research and information is rare for urban forests in South America and in Mediterranean climates, such as south and western Australia [
15], southern Europe [
20,
23], and southern Africa [
27].
The aim of this study is to analyze the spatio-temporal dynamics of a South American urban forest with a Mediterranean climate using tree and plot-level data from permanent monitoring plots. Additionally, to our knowledge, such an approach would make this study one of the first of its kind in South America. Our specific study objectives are to analyze temporal changes from 2002 to 2014 and spatial differences in urban forest structural characteristics from Santiago, Chile including: (1) overall tree population growth, mortality, and ingrowth; (2) basal area, biomass, and tree cover change dynamics; (3) correlates of structural change; and (4) spatial patterns across the study area. We propose to address these research questions using statistical and spatial analyses of field data collected during 2002 and 2014. This type of study can be used to better understand the effects of urbanization and land use change on the structure of not only Latin American but Mediterranean climate urban forests as well. Findings can also be compared to similar plot based studies from other urban forests. Similarly, structural information, such as growth rates and biomass change, is key towards assessing ecosystem services and disservices that are frequently being used in urban planning, sustainability and climate change initiatives, and land management decisions.
4. Discussion
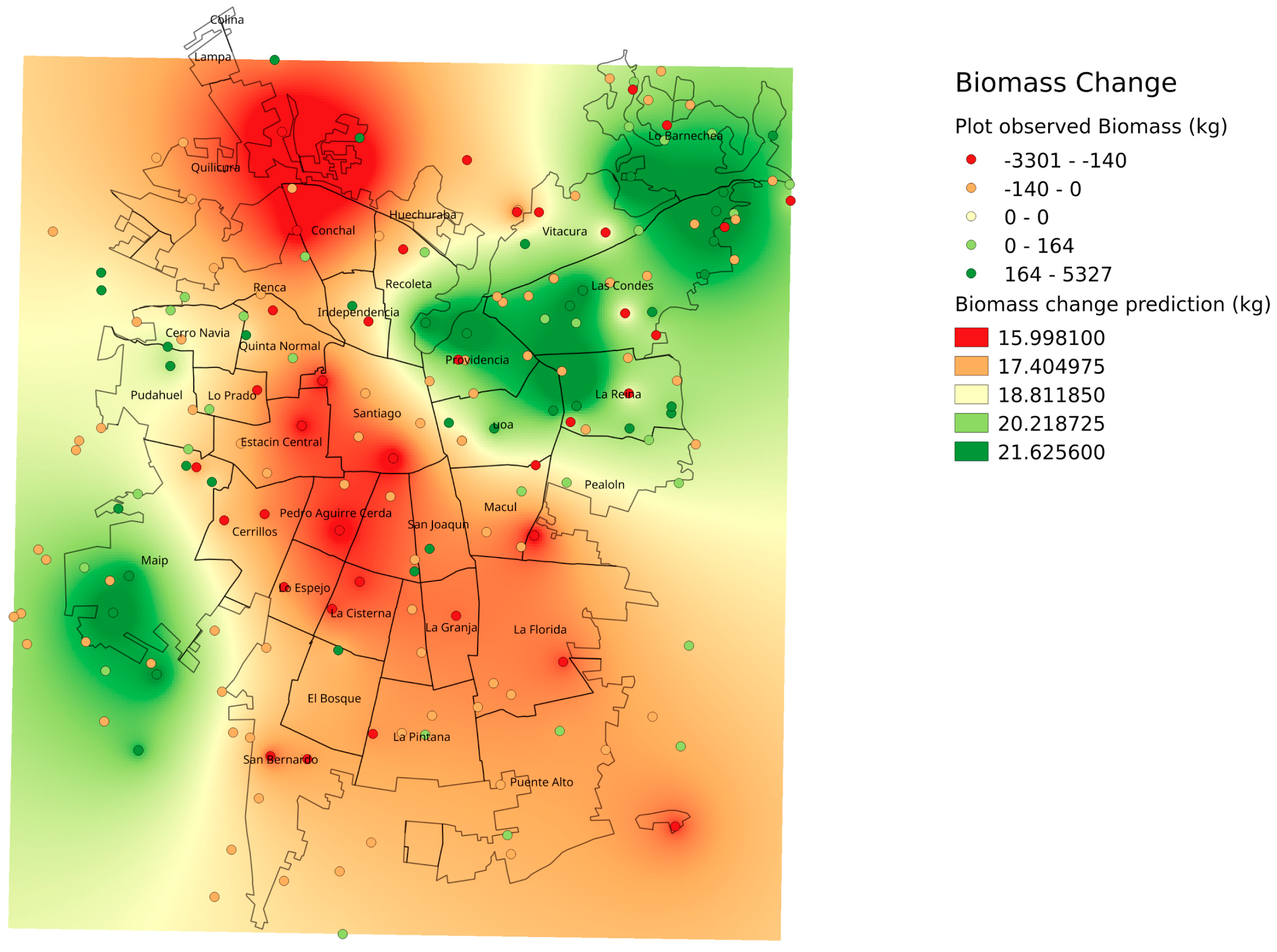
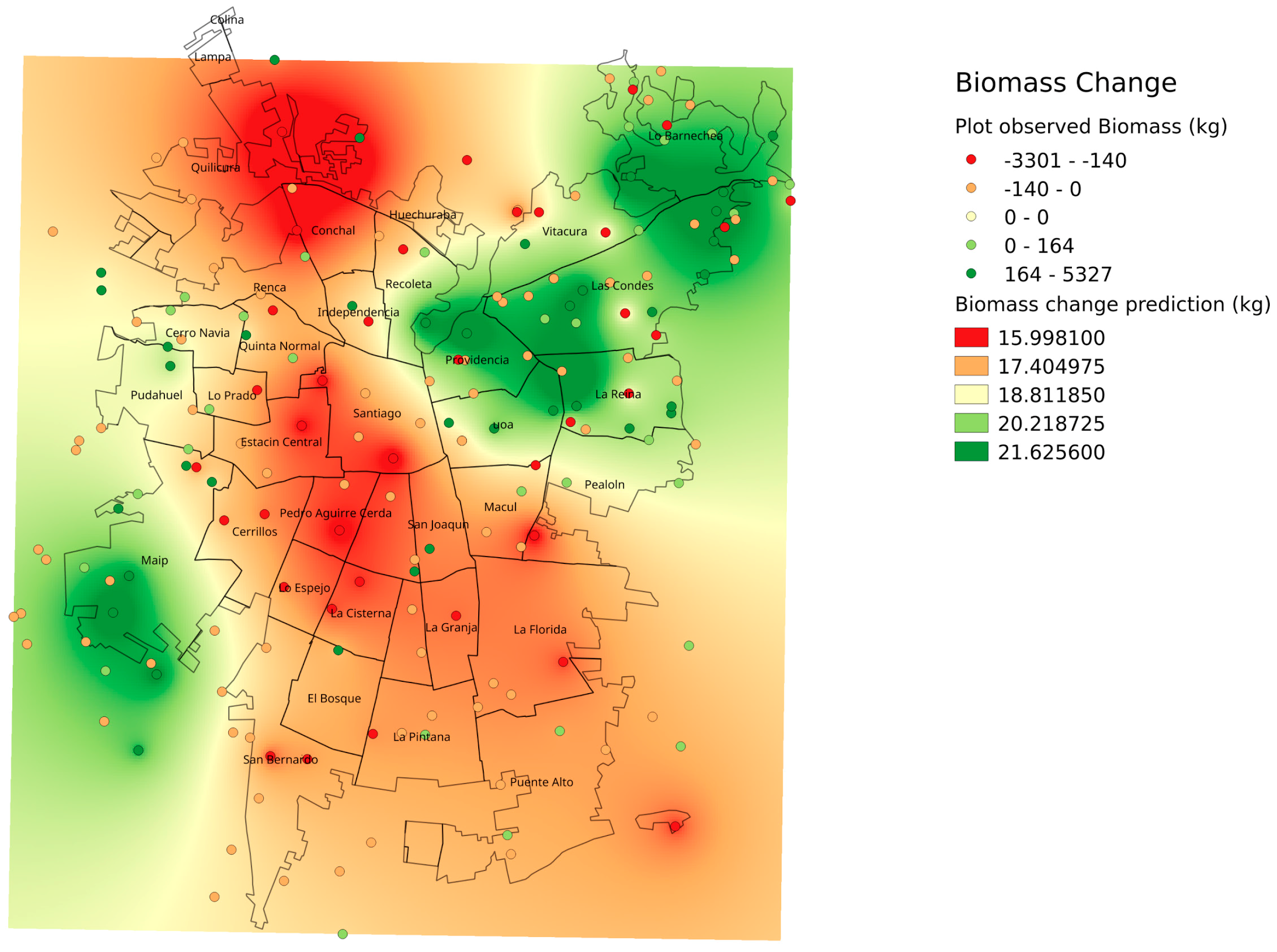
Although no clear evidence can be observed in the 2002 and 2014 Landsat imagery (
Figure 1), the Santiago Metropolitan Area has undergone noticeable urbanization in the form of infill and land use changes since 2002 [
30]. Rapid economic development and recent large-scale infrastructure projects such as new transportation rights of way and hubs, tunnels, and building-housing projects [
35] has negatively affected the structure of Santiago’s urban forest. These changes have occurred for most of the city with the exception of the most affluent areas, the northeast and few areas in the oldest sections of the city center [
35]. Santiago’s semi-arid climate and relative drought related water scarcity explains its low tree cover; however, even small changes can have noticeable effects in terms of the urban forest structure and subsequent functions and services. The tree scarcity in semi-arid Santiago resulted in different patterns than previous studies that used similar field sampling approaches from wet and humid urban forests in temperate and subtropical North America (e.g., Syracuse, USA [
7]; Gainesville, USA [
16]; Orlando, USA [
33] and San Juan, Puerto Rico [
28]); making this study unique.
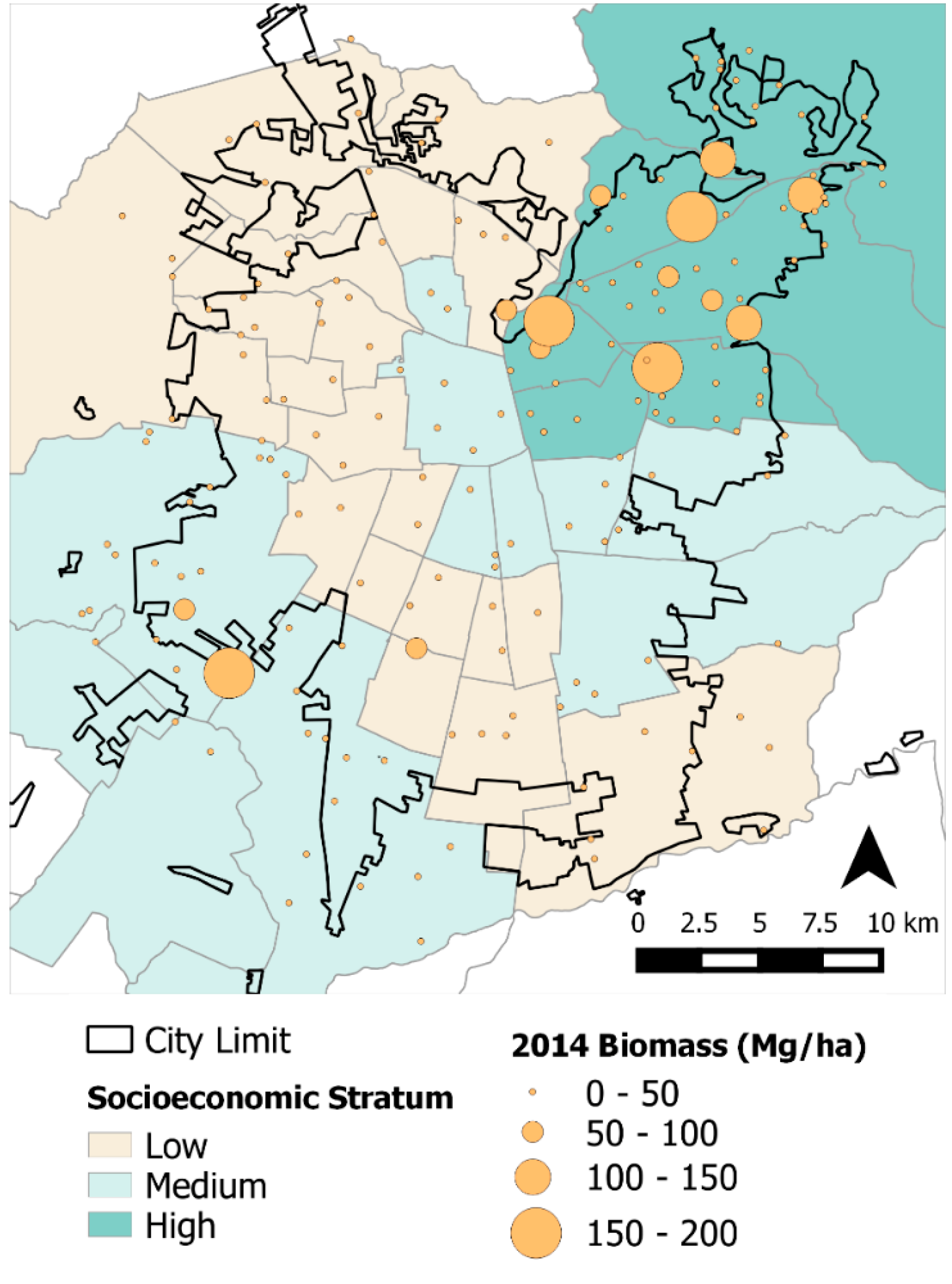
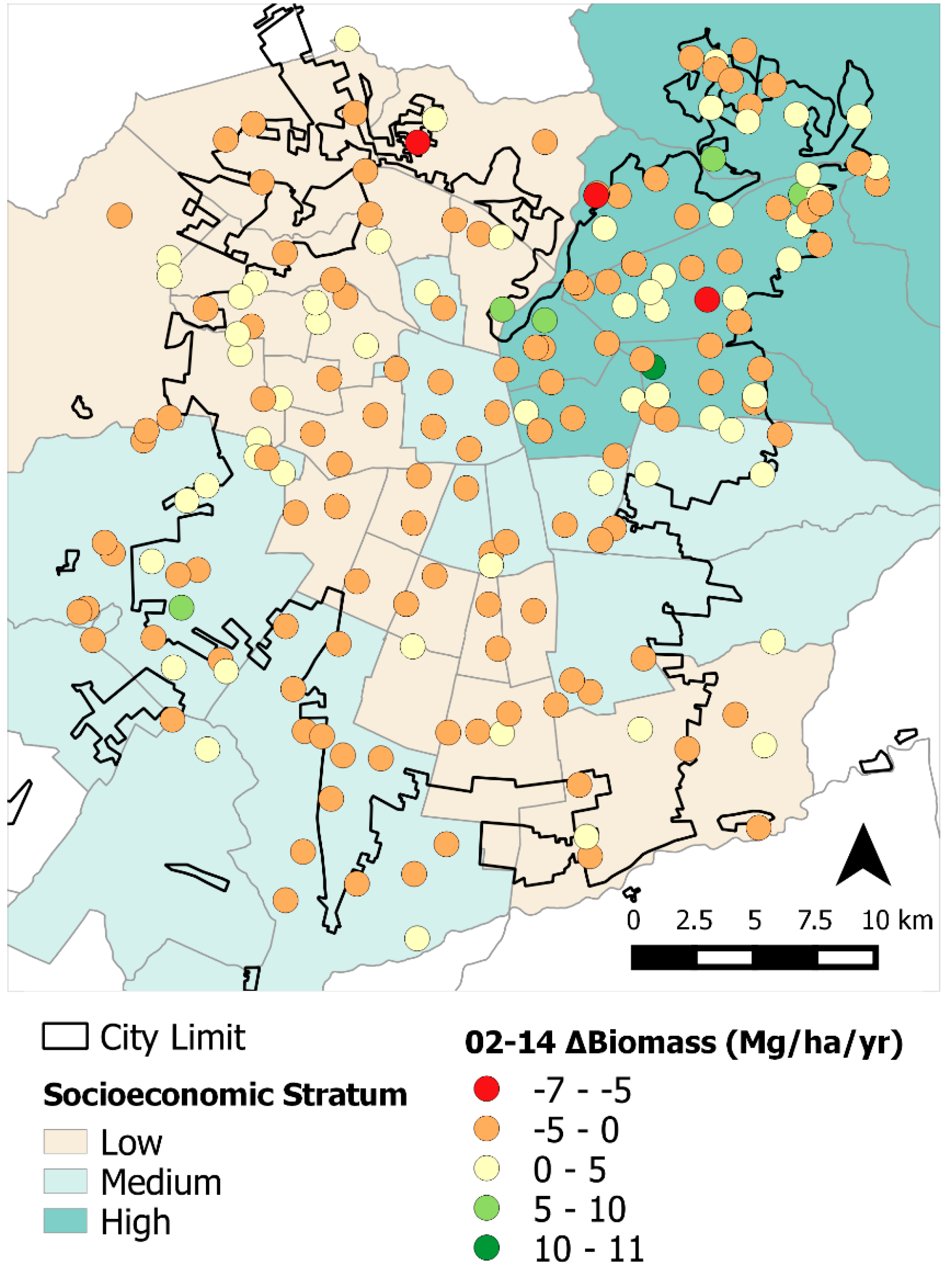
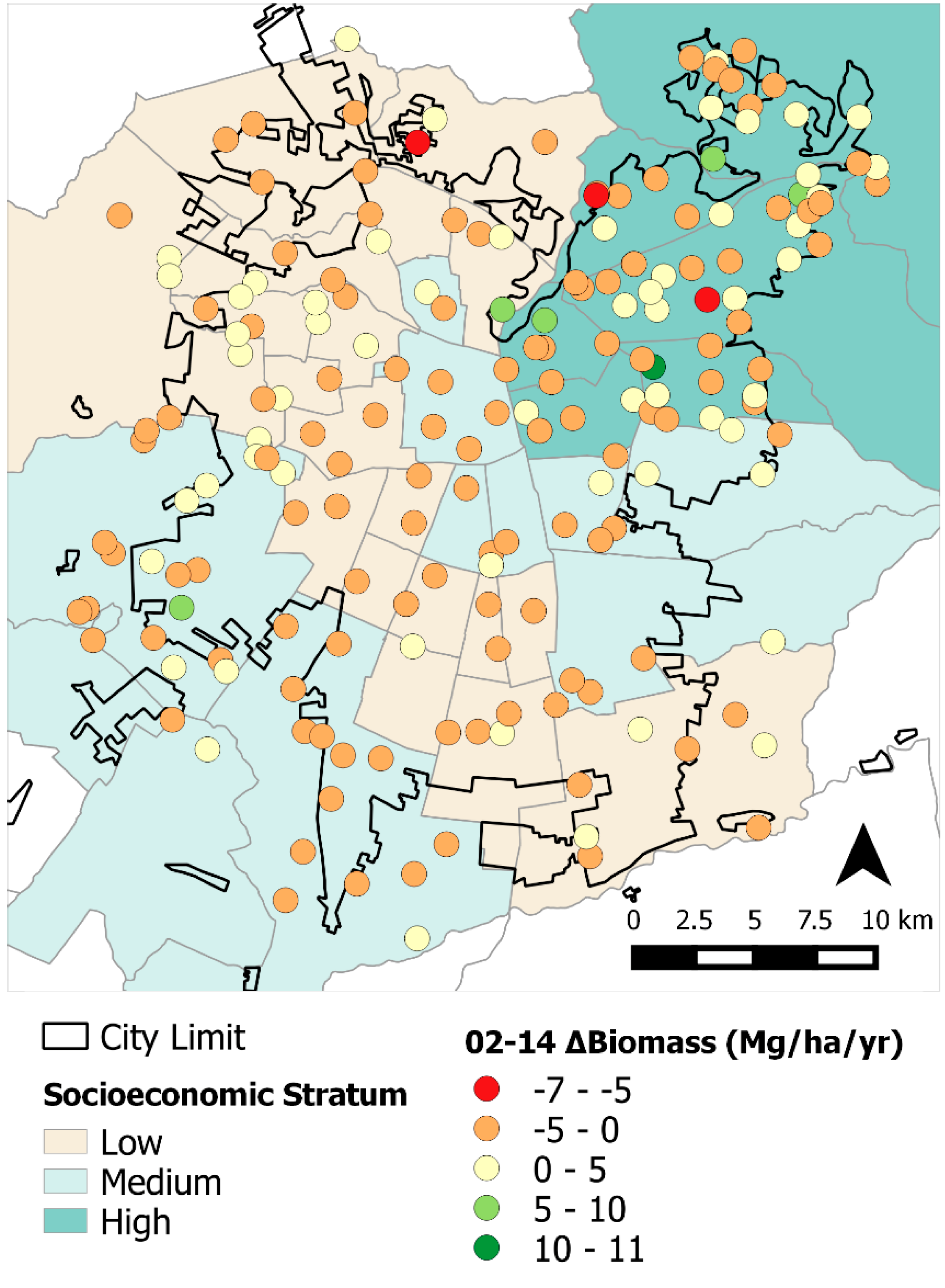
These socio-ecological and urbanization effects were observed in the results from both our statistical and spatial analyses. This is particularly evident from the differences in tree cover, losses in biomass throughout the study area, and BA changes among socioeconomic strata (
Table 3). More affluent
comunas are mostly located in the northeastern section of the city and, because of greater resources, have more tree cover, larger trees (
i.e., greater BA), more abundant trees, and therefore, greater ΔBiomass (0.29 Mg·ha
−1·year
−1) [
6,
15,
36]. Conversely, plots on the medium and lower socioeconomic strata had losses in overall tree cover and negative ΔBiomass. These plots generally correspond to areas with higher building densities and
comunas with lesser economic resources ([
12];
Figure 4). Spatially, plots in the northeastern section of the city had greater tree biomass; however, in the far southwest section of study area we also found high biomass areas corresponding to agricultural land from the peri-urban section (
Figure 2). Rainfall gradients across the study areas could also be playing a role in structural differences as upper elevation plots in the Andean piedmont receive more rain, but management and maintenance regimes (
i.e., irrigation and fertilization) could be masking the effects of precipitation on tree growth and mortality.
Poor correlations between surface cover and BA and biomass might be a result of having small sample sizes and a number of plots with both high cement cover and high biomass and others with high grass cover and low biomass. High cement cover and high biomass are typical of large street trees near building areas. Using the sum of the different vegetation cover (horizontal) and height (vertical) estimates might increase the correlations, as observed by [
33]. We noted that cover types are regularly used as indicators of urbanization, planting space, and urban forest structure and function [
7,
16], but given our semi-arid and high building densities, this might have resulted in different patterns.
We acknowledge that our sample size, in terms of plot density and tree numbers, was low relative to similar urban forest studies using fixed area plots [
7,
16,
28,
33]. Factors—not analyzed in this study—such as climate change and socio-ecological dynamics—could have also driven these changes. Additionally, our use of forest grown allometric equations on urban trees can often lead to errors of up to 40% in biomass estimates [
5]. Similarly, the sample size for conifers and palms was very low, seasonal stem shrinkage and swelling is likely [
4,
16], and this was also confounded by the difficulties in measuring palms using measurement techniques developed for single stem temperate trees [
4,
16,
28]. However, we note that all these cited studies had the same limitations. Indeed, given the context of our study area’s: size, socio-political dynamics, and limited infrastructure that characterizes cities from developing countries, we feel that our results do provide a better understanding of urban forests outside the frequently studied areas of the United States and Canada, giving more insights to understudied areas such as Latin America [
1,
13,
32], Mediterranean climates [
12,
28,
32], and Australia [
3,
15].
When comparing urban forest structure results such as annual mortality to other studies using inventory data, we can distinguish the effect of climate and management. Mortality of urban trees was apparently more related to management than to climate; Santiago showed much lower mortality rates (3.0%) than subtropical San Juan, Puerto Rico [
28], with values between 30%–40%. Conversely a subtropical city in the United States, Gainesville [
16] had an average mortality rate between 10%–19%. Looking at growth rates, climate appears to be a stronger driver than management; values for subtropical cities of San Juan and Gainesville are closer to each other (0.7–1.0 cm·year
−1), while Santiago had lower average values between 0.1 to 0.4 cm·year
−1. In terms of the temporal changes to urban forest structure, there are no similarities to relevant studies [
7,
28].
Studies that use permanent random plots often report number of trees per unit area—as opposed to basal area—as a measure of tree density [
7]; however, multi-stemmed trees with a shrub form or secondary tropical forests and mangroves can confound comparisons, thus our preference for basal area. Values for tree density can vary from 222–328 trees per ha in San Juan in 2001 and 2010, respectively [
28]. While there is an average of 34 trees per ha in a temperate city such as Syracuse, US [
7], in Santiago, a Mediterranean shrubland biome, this value reached 64 trees per ha [
12]. Meanwhile, there was an overall annual net loss of approximately four trees per ha in subtropical Gainesville [
16]. Tucker-Lima
et al. report a basal area of 4.6 m
2·ha
−1 in San Juan, which is very similar to our 2014 estimate of 4.8 m
2·ha
−1. Changes in basal area in Santiago of 0.1 m
2·ha
−1 were, however, much lower than in San Juan (1.0 m
2·ha
−1) [
28].
The growing body of literature using similar sized, long-term monitoring plots as utilized in this study can be used to compare trends for urban forests across different regions of the globe [
5,
6,
16,
28,
33,
36]. The characteristics of urban forest growth, mortality, and the effects of site characteristic such as irrigation, and ecological disturbance on these, could be analyzed against field measured plot and site correlates [
5,
16,
20,
21]. However, the presence of palms, multi-stemmed tall shrubs, and size criteria for tree-shrub differentiation across different biomes can confound some of these comparisons. Care is also warranted when comparing shrubland dominated biomes such as Santiago, to dense subtropical secondary and mangrove forests or temperate forests. Similarly, because of the difficulty in sampling large, heterogeneous, urbanized areas, sampling intensities can be low and result in larger uncertainties. This is particularly true in Santiago’s densely built, semi-arid, urban context, which also resulted in a reduced sample of measured and matched trees.
Our study provides one of the few comparative insights into how a South American and Mediterranean urban forest changes across space and time. Future research could analyze spatio-temporal changes in urban forest composition and its subsequent effects such as the spatial dynamics of ecosystem service provision and disservice hotspots [
11,
13,
21,
22,
24,
25,
26]. Plot level structure-function information could also be used to test land management and planning scenario effects on the demand and supply of services [
4,
7,
14,
33]. Quantitative analyses could also determine the socio-ecological causal factors behind, and the drivers related to, changes in urban forests, their processes, and services [
1,
3,
6,
23,
36]. More basic research could use these monitoring sites for better understanding floral diversity and the occurrence of invasives and understory dynamics in urbanized forests, as well as for developing allometric equations [
36,
37,
38,
39]. Permanent plots could also facilitate dendrochronological analysis of different tree species and their growth and mortality as affected by climate change, pollution, maintenance practices, and community preferences.
5. Conclusions
Overall, we found that urban forest mortality, ingrowth, and tree cover in the greater Santiago area remained stable during 2002 to 2014. We also noticed slight losses in basal area and biomass change across the study area. However, there were some noticeable trends during the analysis period in that tree cover increased and was greater in the highest socioeconomic stratum; however, it decreased in the medium and low strata. Similarly, the less affluent central, northwest, and southern plots, in particular, exhibited losses in BA and biomass from 2002 to 2014.
As previously mentioned, other studies have used plot-level data to analyze changes in an urban forest but, to our knowledge, most of these studies are from North America. Here, we describe for the first time the spatio-temporal dynamics of a South American urban forest in a Mediterranean climate. Overall, we found that these urban forests follow similar trends as in other parts of the world. For example, affluent areas tend to have more and better condition urban forests than poorer areas. Thus, barring substantial ecological disturbance (e.g., storms, drought, pests, or urbanization), changes in urban forest structure will primarily depend on human management (e.g., maintenance and irrigation) and biophysical factors (i.e., growing space and climate). Findings could also be used to identify which tree species perform better in terms of growth and basal area. Accordingly, this information could be used to identify tree functional traits that are most associated with ecosystem service provision (e.g., carbon offsets) and disservice minimization (e.g., allergenic tree locations).
However, Latin America and the rest of the developing world are generally characterized by unplanned land uses and marked socioeconomic inequities. Thus, studies such as ours can be used for targeting specific sites for improved management and setting monitoring and evaluation standards for municipalities. Conversely, areas that are maintaining their urban forest and standards—despite limited budgets—could also be identified. Most importantly, issues of environmental justice and resilience to the effects of climate and socio-political changes could be better addressed with this long-term data. Indeed, the growing body of literature calls for the development of an available global urban forest monitoring database that could be used for research by the global scientific community.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}