
Soil Enzyme Activities in Pinus tabuliformis (Carriére) Plantations in Northern China

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Soil Sampling and Physico-Chemical Analyses
2.3. Extracellular Enzyme Activity Assays
2.4. Data Analysis
3. Results
3.1. Seasonal Response of Soil Enzyme Activity to Pine after Thinning
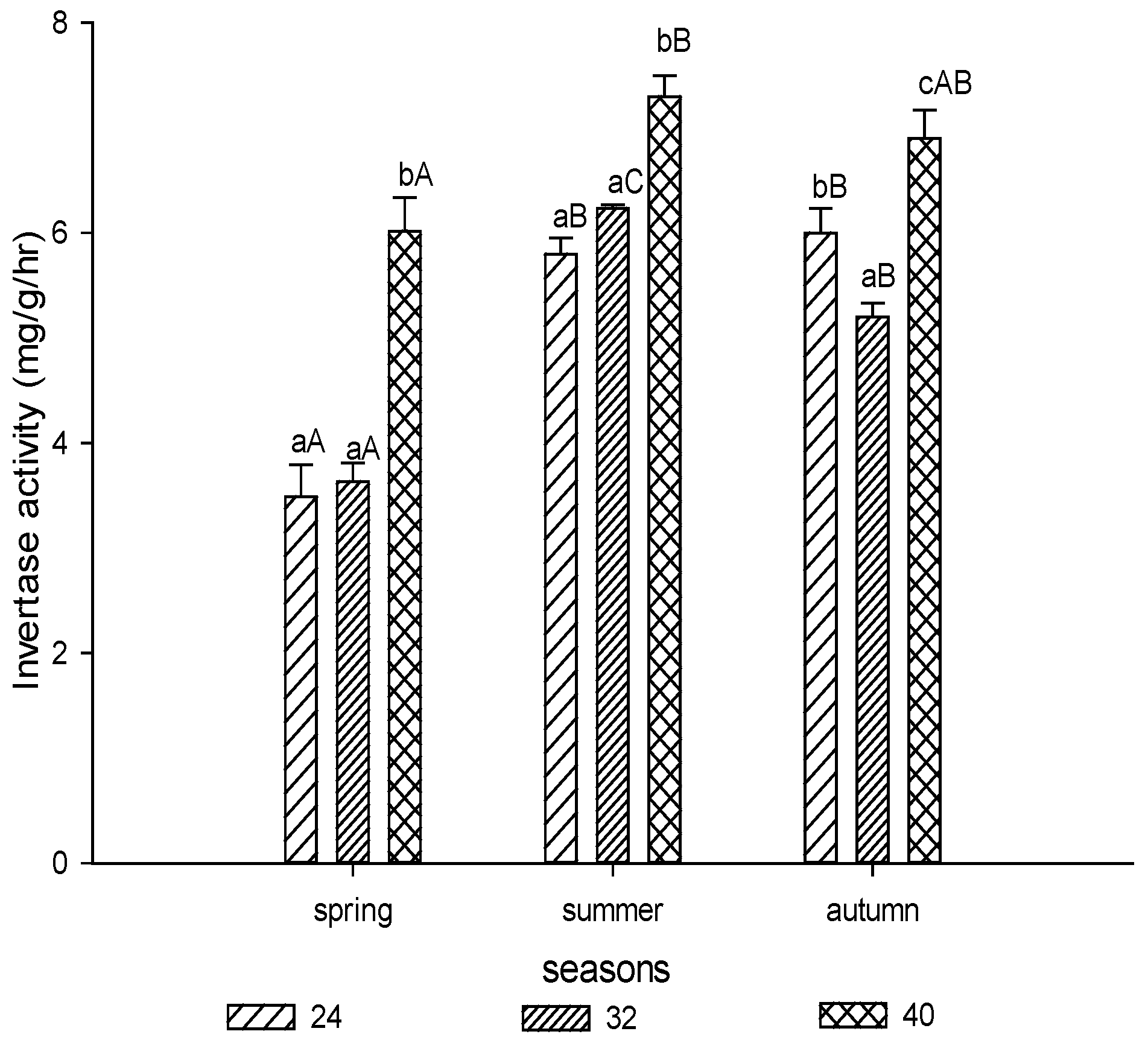
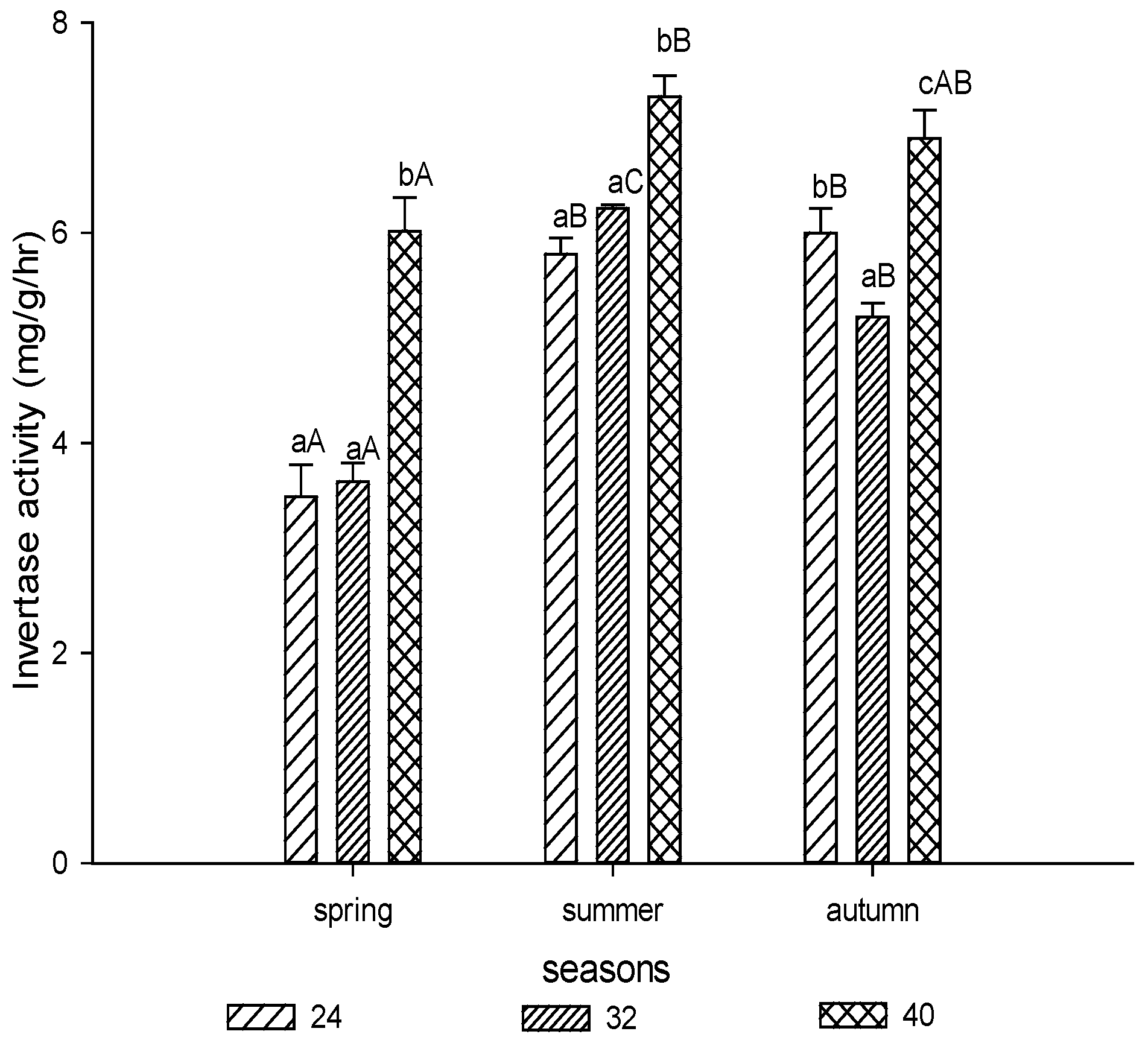
3.1.1. Invertase Activity
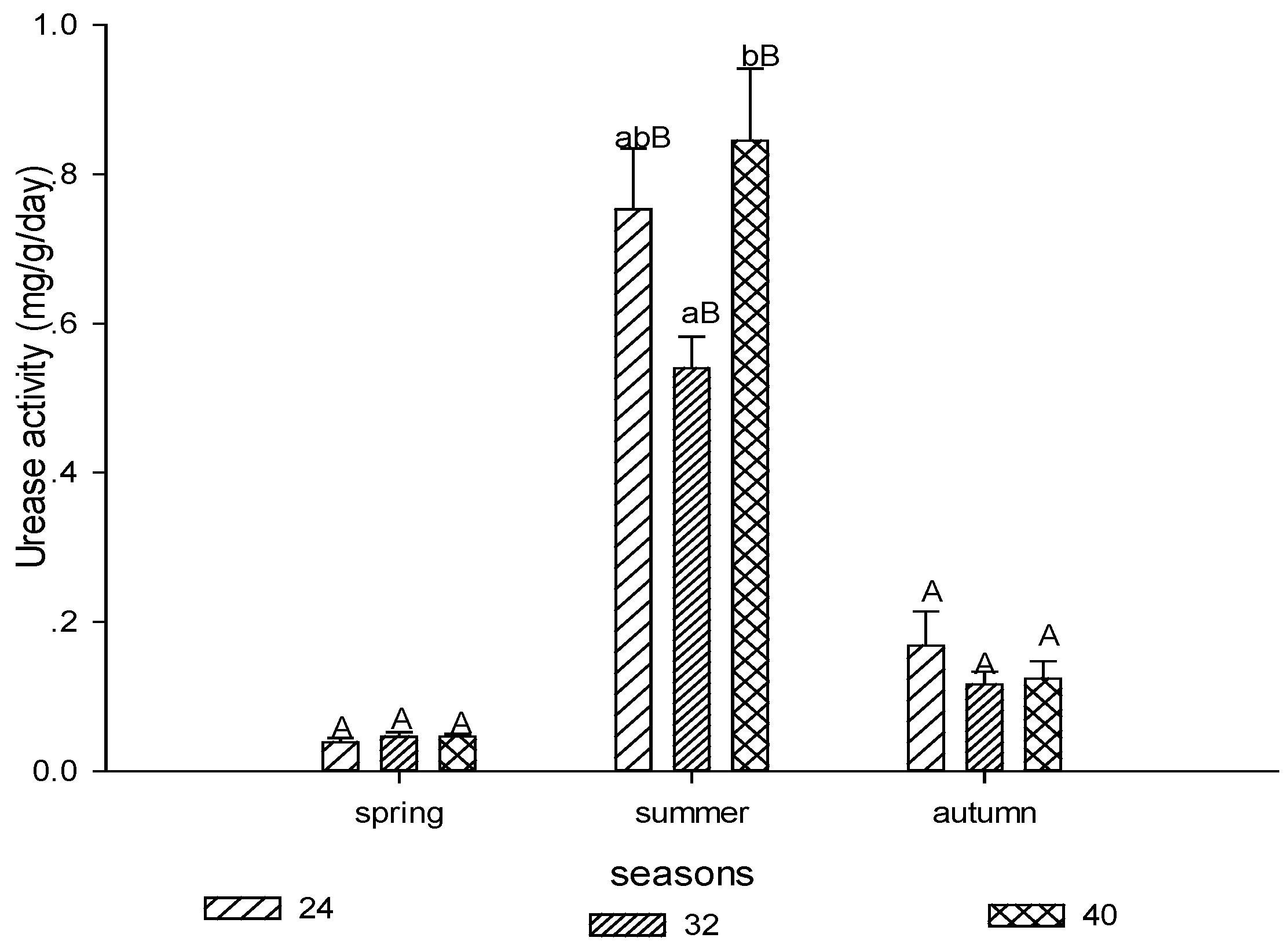
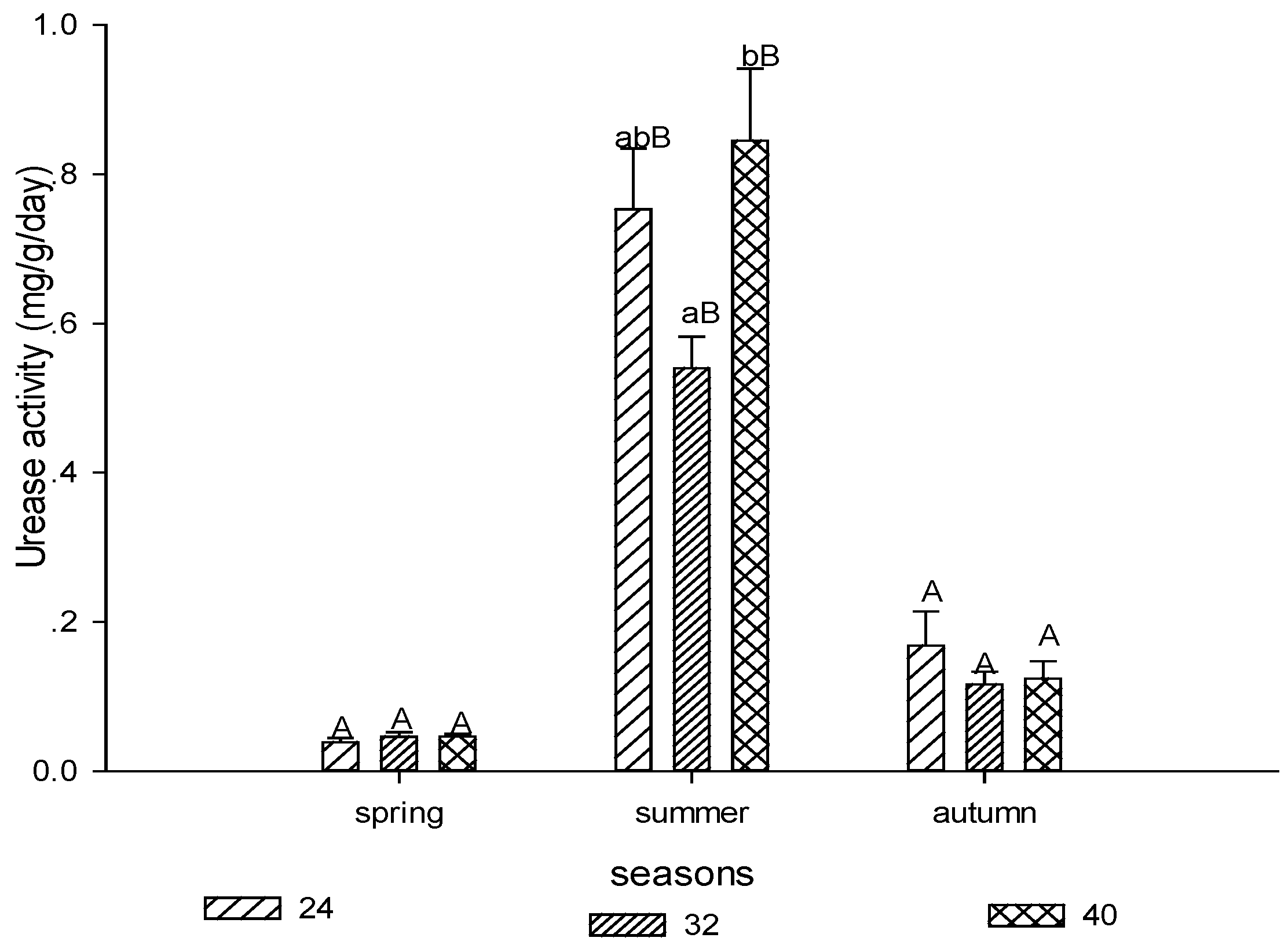
3.1.2. Urease Activity
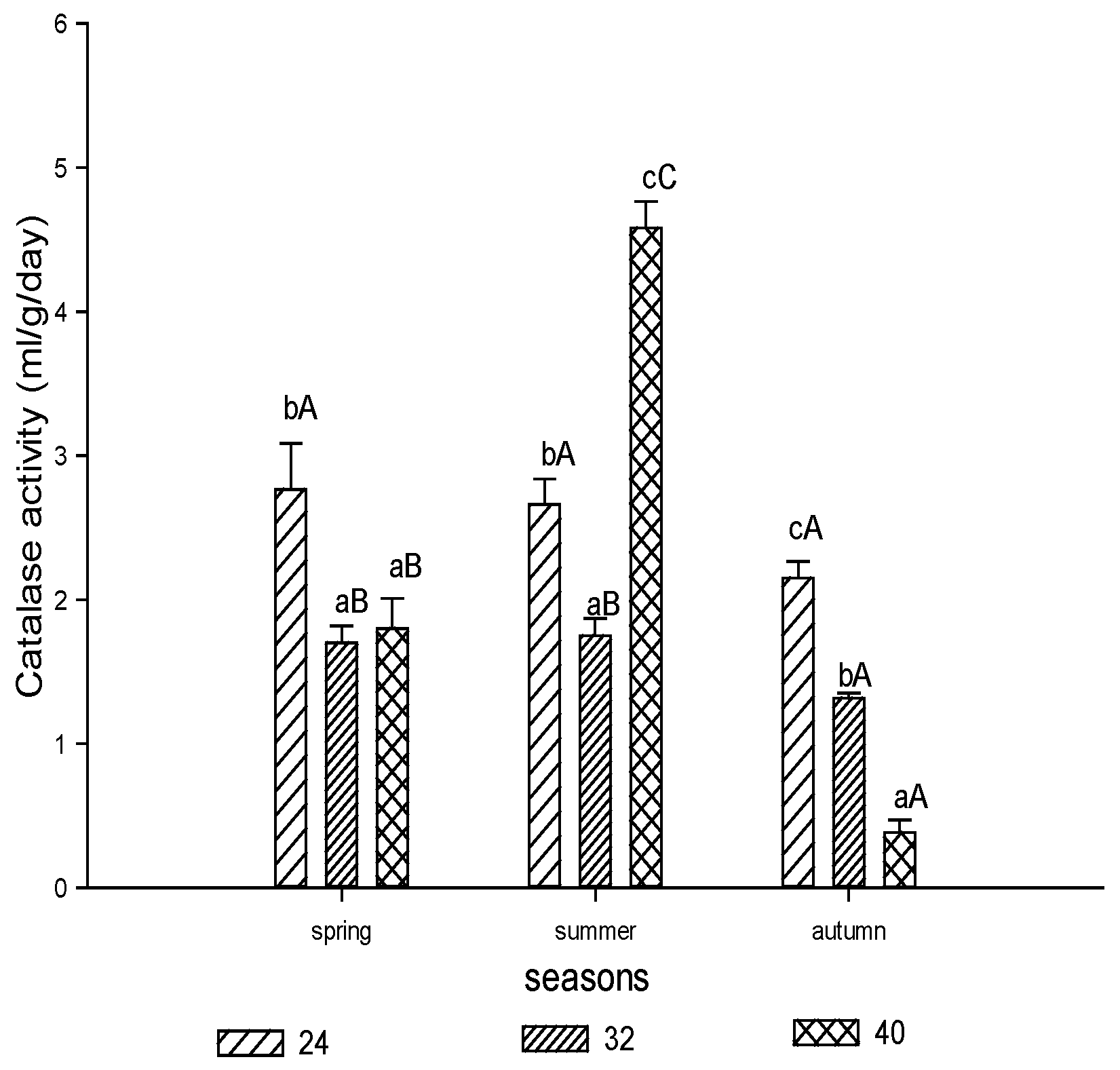
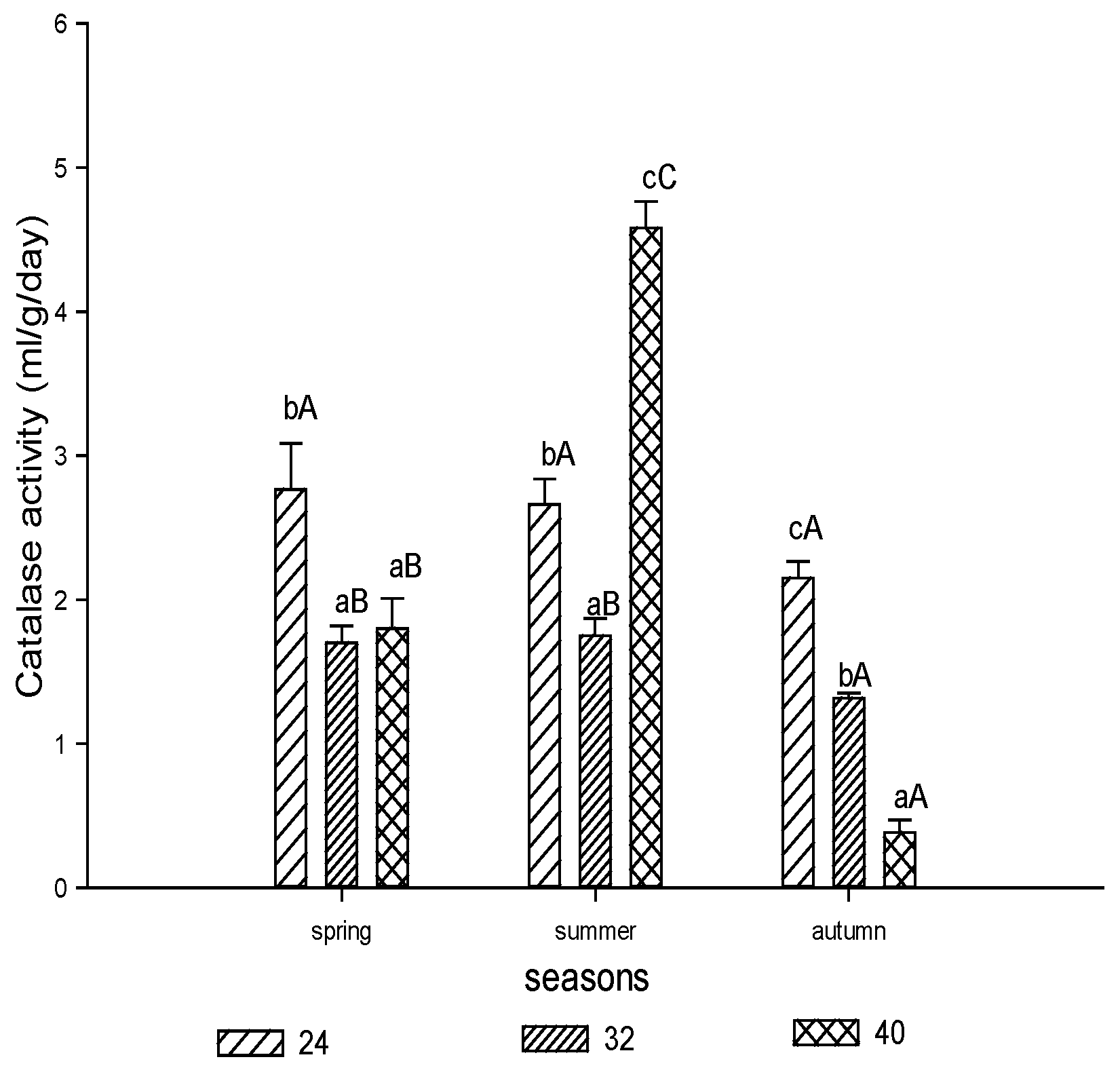
3.1.3. Catalase Activity
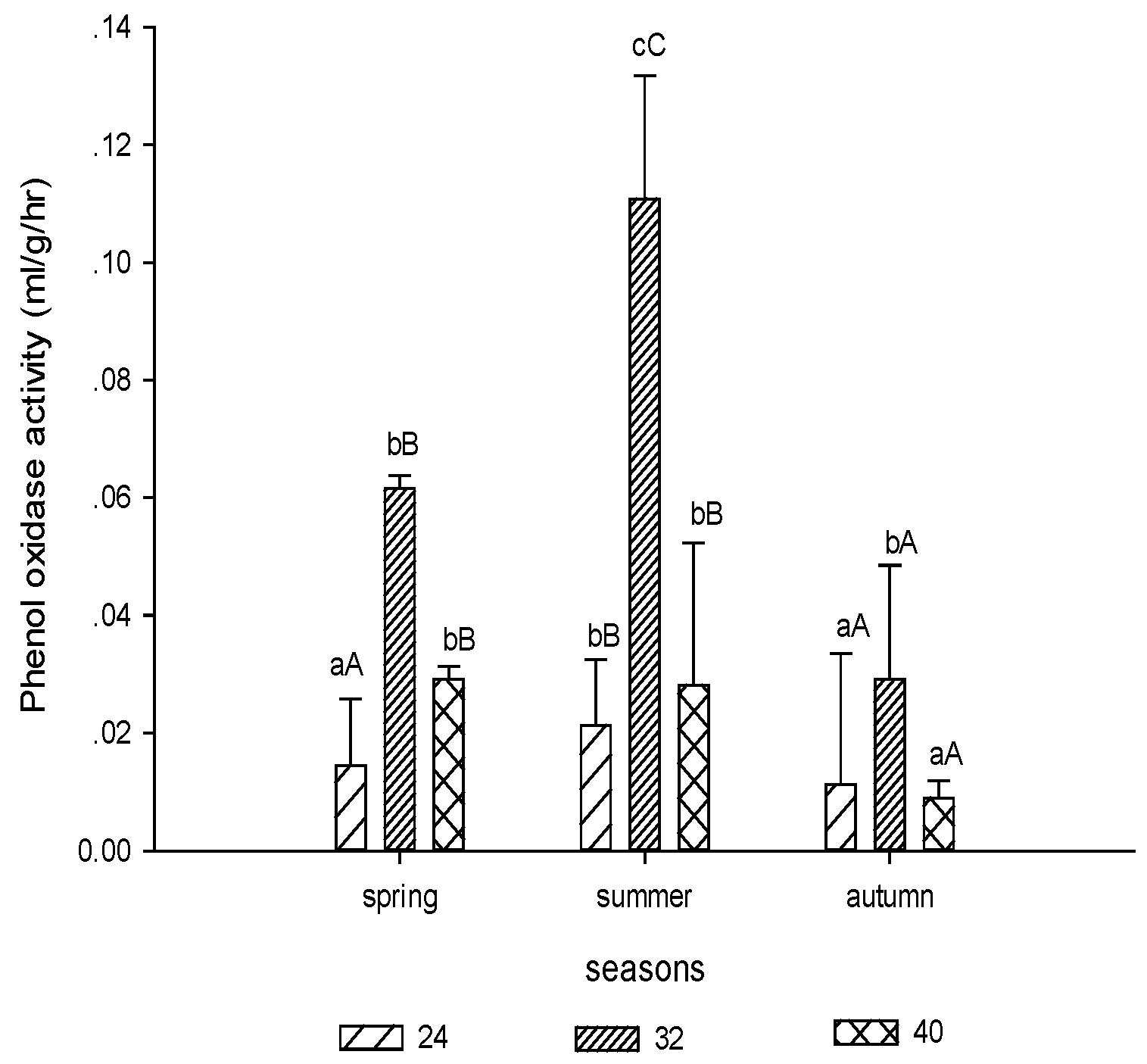
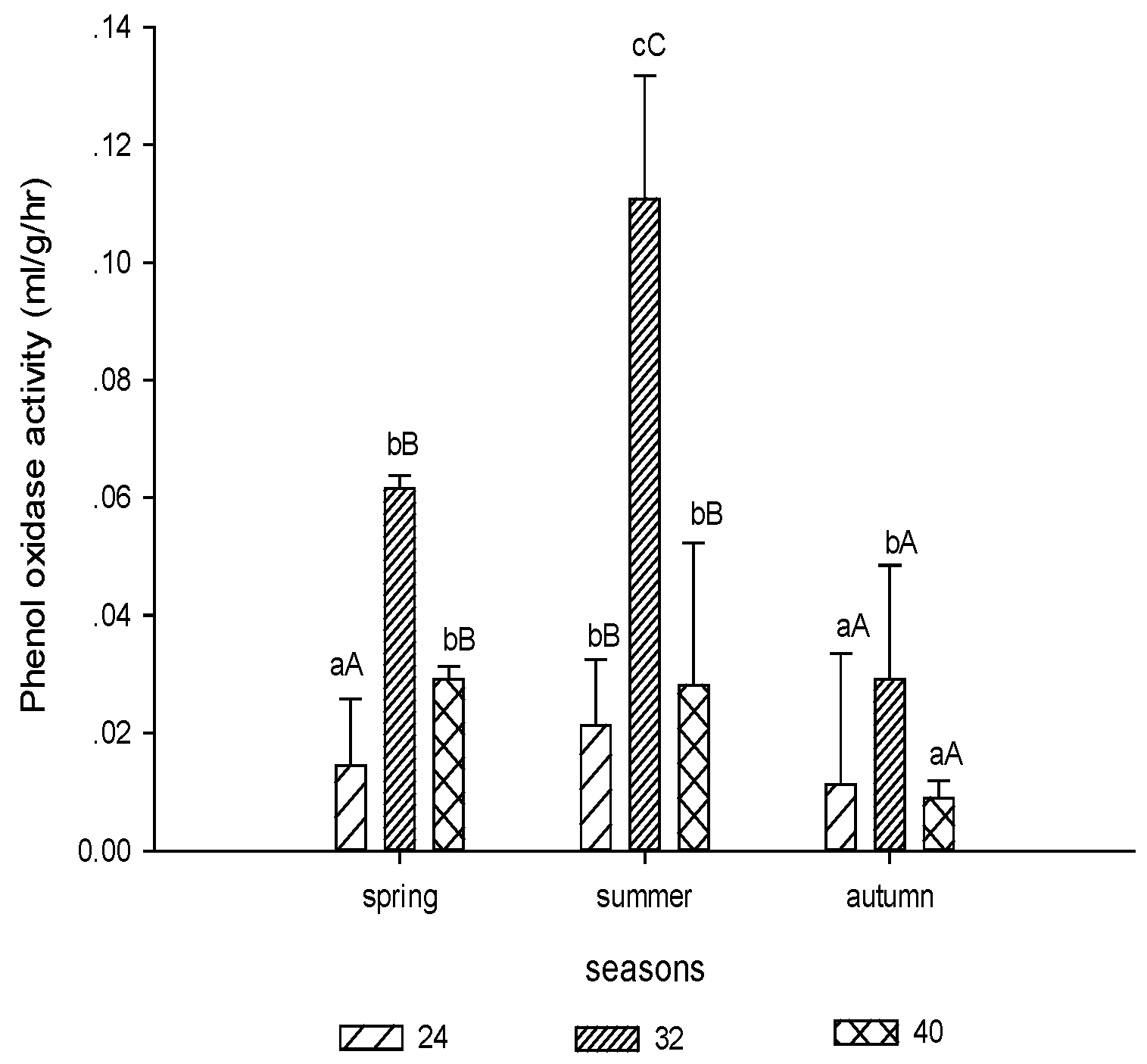
3.1.4. Phenol Oxidase Activity
3.2. Relationships Among Soil Enzyme Activity
3.3. Soil Enzymes at Different Soil Depths
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dick, R.P. A review: Long-term effects of agricultural systems on soil biochemical and microbial parameters. Agric. Ecosyst. Environ. 1992, 40, 25–36. [Google Scholar] [CrossRef]
- Ming, L.; Pang, J.F.; Li, X.Y.; Liu, P.; Wan, L.Y. Microbiological characteristics and enzymes activity of the forest soil in Picea schrenkiana var. tianshanican in Xinjiang. Chin. J. Ecol. 1997, 16, 26–30. [Google Scholar]
- Bergstrom, D.W.; Monreal, C.M.; King, D.J. Sensitivity of soil enzyme activities to conservation practices. Soil Sci. Soc. Am. J. 1998, 62, 1286–1295. [Google Scholar] [CrossRef]
- Dick, R.P. Soil enzyme activities as indicators of soil quality. In Soil Enzymes; Doran, J.W., Coleman, D.C., Bezdicek, D.F., Stewart, B.A., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 107–124. [Google Scholar]
- Dick, W.A.; Tabatabai, W.A. Significance and potential use of soil enzymes. In Soil Microbial Ecology; Metting, F.B., Ed.; Marcel Decker: New York, NY, USA, 1992; pp. 95–130. [Google Scholar]
- Boerner, R.E.J.; Brinkman, J.A.; Smith, A. Seasonal variations in enzyme activity and organic carbon in soil of a burned and unburned hardwood forest. Soil Biol. Biochem. 2005, 37, 1419–1426. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Moorhead, D. Resource allocation to extracellular enzyme production: A model for nitrogen and phosphorus control of litter decomposition. Soil Biol. Biochem. 1994, 26, 1305–1311. [Google Scholar] [CrossRef]
- Cusack, D.F.; Torn, M.S.; McDowell, W.H.; Silver, W.L. The response of heterotrophic activity and carbon cycling to nitrogen additions and warming in two tropical soils. Glob. Chang. Biol. 2010, 16, 2555–2572. [Google Scholar] [CrossRef]
- Waldrop, M.; Zak, D.R.; Sinsabaugh, R.L. Nitrogen deposition modifies soil carbon storage through changes in microbial enzymatic activity. Ecol. Appl. 2004, 14, 1172–1177. [Google Scholar] [CrossRef]
- Trasar-Cepeda, C.; Leirós, M.C.; Gil-Sotres, F. Hydrolytic enzyme activities in agricultural and forest soils. Some implications for their use as indicators of soil quality. Soil Biol. Biochem. 2008, 40, 2146–2155. [Google Scholar] [CrossRef]
- Skujins, J.; Burns, R.G. Extracellular enzymes in soil. CRC Crit. Rev. Microbiol. 1976, 4, 383–421. [Google Scholar] [CrossRef] [PubMed]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Cessherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Bandick, A.K.; Dick, R. Field management effects on soil enzyme activities. Soil Biol. Biochem. 1999, 31, 1471–1479. [Google Scholar] [CrossRef]
- Dick, R.P.; Sandor, J.A.; Eash, N.S. Soil enzyme activities after 1500 years of terrace agriculture in the Colca Valley, Peru. Agric. Ecosyst. Environ. 1994, 50, 123–131. [Google Scholar] [CrossRef]
- Kandeler, E.; Eder, G. Effect of cattle slurry in grassland on microbial biomass and on activities of various enzymes. Biol. Fertil. Soils 1993, 16, 249–254. [Google Scholar] [CrossRef]
- Nuñez-Regueira, L.; Rodríquez-Añon, J.A.; Proupín-Castiñeras, J.; Núñez-Fernández, O. Microcalorimetric study of changes in the microbial activity in a humic Cambisol alter reforestation with eucalyptus in Galicia (NW Spain). Soil Biol. Biochem. 2006, 38, 115–124. [Google Scholar] [CrossRef]
- Zinn, Y.L.; Resck, D.V.S.; da Silva, J.E. Soil organic carbon as affected by afforestation with Eucalyptus and Pinus in the Cerrado region of Brazil. For. Ecol. Manag. 2002, 166, 285–294. [Google Scholar] [CrossRef]
- Resck, D.V.S.; Vasconcellos, C.A.; Vilela, L.; Macedo, M.C.M. Impact of conversion of Brazilian Cerrados to cropland and pasturland on soil carbon pool dynamics. In Global Climate Change and Tropical Ecosystems. Advances in Soil Science; Lal, R., Kimble, J.M., Stewart, B.A, Eds.; Lewis Publishers: Boca Raton, FL, USA, 2000; pp. 169–196. [Google Scholar]
- Bauhaus, J.; Paré, D.; Côté, L. Effect of tree species, stand age, and soil type on soil microbial biomass and its activity in a southern boreal forest. Soil Biol. Biochem. 1998, 30, 1077–1089. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.A.; Fang, J.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Piao, S.L.; Fang, J.Y.; Ciais, P.; Peylin, P.; Huang, Y.; Sitch, S.; Wang, T. The carbon balance of terrestrial ecosystems in China. Nature 2009, 458, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.; Kjøller, A.; Struwe, S. Microbial enzyme activities in leaf litter, humus and mineral soil layers of European forests. Soil Biol. Biochem. 2004, 36, 1527–1537. [Google Scholar] [CrossRef]
- Gartner, T.B.; Treseder, K.K.; Malcolm, G.M.; Sinsabaugh, R.T. Extracellular enzyme activity in the mycorrhizospheres of a boreal forest chronosequence. Pedobiologia 2012, 55, 121–127. [Google Scholar] [CrossRef]
- Higuchi, T. Lignin biochemistry: Biosynthesis and biodegradation. Wood Sci. Technol. 1990, 24, 23–63. [Google Scholar] [CrossRef]
- Holden, S.R.; Gutierrez, A.; Treseder, K.K. Changes in soil fungal communities, extracellular enzyme activities, and litter decomposition along a fire chronosequence in Alaskan boreal forests. Ecosystems 2013, 16, 34–46. [Google Scholar] [CrossRef]
- Tan, X.; Chang, S.X.; Kabzems, R. Soil compaction and forest floor removal reduced microbial biomass and enzyme activities in a boreal aspen forest soil. Biol. Fertil. Soils 2008, 44, 471–479. [Google Scholar] [CrossRef]
- Yuan, B.C.; Yue, D.X. Soil microbial and enzymatic activities across a chronosequence of Chinese pine plantation development on the loess plateau in China. Pedosphere 2012, 22, 1–12. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L. Phenol oxidase, peroxidase, and organic matter dynamics of soil. Soil Biol. Biochem. 2010, 42, 391–404. [Google Scholar] [CrossRef]
- Pancholy, S.K.; Rice, E.L. Soil enzymes in relation to old field succession: Amylase, cellulose, invertase, dehydrogenase, and urease. Soil Sci. Soc. Am. J. 1973, 37, 47–50. [Google Scholar] [CrossRef]
- Conrad, J.P. The occurrence and origin of urease like activities in soils. Soil Sci. 1942, 54, 367–380. [Google Scholar] [CrossRef]
- Xun, W.; Huang, T.; Zhao, J.; Ran, W.; Wang, B.; Shen, Q.; Zhang, R. Environmental conditions rather than microbial inoculum composition determin the bacterial composition, microbial biomass, and enzymatic activity of reconstructed soil microbical communities. Soil Biol. Biochem. 2015, 90, 10–18. [Google Scholar] [CrossRef]
- Huo, Y.Z. Beijing Physical Geography; Beijing Normal University Publishing Group: Beijing, China, 1989. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, 10th ed.; USDA National Soil Conservation Service: Washington, DC, USA, 2006; pp. 331–332. [Google Scholar]
- Horwitz, W. Official Methods of Analysis of the Association of Official Analytical Chemists, 13th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1980. [Google Scholar]
- Yan, C.S. Soil Fertility Research Methods; China Agriculture Press: Beijing, China, 1988; pp. 243–279. (In Chinese) [Google Scholar]
- Guan, S.Y. Soil Enzyme and Its Research Approaches; China Agriculture Press: Beijing, China, 1986. (In Chinese) [Google Scholar]
- Trasar-Cepeda, C.; Camina, F.; Leiros, M.C.; Gil-Sotres, F. An improved method to measure catalase activity in soils. Soil Biol. Biochem. 1999, 31, 483–485. [Google Scholar] [CrossRef]
- Levene, H. Robust tests for equality and variances. In Contributions to Probability and Statistics; Olkin, I., Ghurye, S.G., Hoeffding, W., Madow, W.G., Mann, H.B., Eds.; Stanford University Press: Stanford, CA, USA, 1960; pp. 278–292. [Google Scholar]
- Lv, R.H.; Li, G.L.; Liu, Y.; Jin, H.; Lin, N. Study on initial decomposition characteristics of Pinus tabulaeformis needle litter at different stand age. J. Nanjing For. Univ. (Nat. Sci. Ed.) 2013, 37, 37–44. [Google Scholar] [CrossRef]
- Barford, C.; Lajtha, K. Nitrification and nitrate reductase rate along a secondary successional gradient. Plant Soil 1992, 145, 1–10. [Google Scholar] [CrossRef]
- Frankenberger, W.T., Jr.; Johanson, J.B. Factors affecting invertase activity in soil. Plant Soil 1983, 74, 313–323. [Google Scholar] [CrossRef]
- Frankenberger, W.T., Jr.; Johanson, J.B. Effect of pH on enzyme stability in soils. Soil Biol. Biochem. 1982, 14, 433–437. [Google Scholar] [CrossRef]
- Ross, D.J. Some enzyme and respiratory activities of tropical soils from New Hebrides. Soil Biol. Biochem. 1973, 5, 559–567. [Google Scholar] [CrossRef]
- O’Connell, A.M. Litter decomposition, soil respiration, and soil chemical and biochemical properties at three contrasting sites in karri (Eucalyptus diversicolor F. Muell) forests of south-western Australia. Aust. J. Ecol. 1987, 12, 31–40. [Google Scholar] [CrossRef]
- Page-Dumroese, D.S.; Jurgensen, M.F. Soil carbon and nitrogen pools in mid—To late-successional forest stands of the northwestern United States: Potential impacts of fire. Can. J. For. Res. 2006, 36, 2270–2284. [Google Scholar] [CrossRef]
- Boerner, R.E.J.; Giai, C.; Huang, J.; Miesel, J.R. Initial effects of fire and mechanical thinning on soil enzyme activity and nitrogen transformations in eight North American forest ecosystems. Soil Biol. Biochem. 2008, 40, 3076–3085. [Google Scholar] [CrossRef]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta-analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Gallo, M.E.; Lauber, C.L.; Waldrop, M.P.; Zak, D.R. Extracellular enzyme activities and soil carbon dynamics for northern hardwood forests receiving simulated nitrogen deposition. Biogeochemistry 2005, 75, 201–215. [Google Scholar] [CrossRef]
- A’Bear, A.D.; Joanes, T.H.; Kandeler, E.; Boddy, L. Interactive effects of temperature and soil moisture on fungal-mediated wood decomposition and extracellular enzyme activity. Soil Biol. Biochem. 2014, 70, 151–158. [Google Scholar] [CrossRef]
- Fekete, I.; Kotroczó, Z.; Varga, C.; Hargitai, R.; Townsend, K.; Csanye, G.; Varbiro, G. Variability of organic matter inputs affects soil moisture and soil biological parameters in a European detritus manipulation experiment. Ecosystems 2012, 15, 792–803. [Google Scholar] [CrossRef]
- Baldrian, P.; Merhautová, V.; Cajthaml, T.; Petránková, M.; Snajdr, J. Small-scale distribution of extracellular enzymes, fungal, and bacterial biomass in Quercus petraea forest topsoil. Biol. Fertil. Soils 2010, 46, 717–726. [Google Scholar] [CrossRef]
- Rubén, L.; Jana, V.; Tomáš, V.; Petr, B. The bacterial community inhabiting temperate deciduous forests is vertically stratified and undergoes seasonal dynamics. Soil Biol. Biochem. 2015, 87, 43–50. [Google Scholar] [CrossRef]
- Zsolt Kotroczó, Z.; Veres, Z.; Fekete, I.; Krakomperger, Z.; Toth, J.A.; lajtha, K.; Tothmeresz, B. Soil enzyme activity in response to long-term organic matter manipulation. Soil Biol. Biochem. 2014, 70, 237–243. [Google Scholar] [CrossRef]
- Tao, B.X.; Zhang, J.C.; Yu, Y.C. Season variations of forest soil enzyme activities in the hilly region of southern Jiangsu province. Ecol. Environ. Sci. 2010, 19, 2349–2354. [Google Scholar]
- Weintraub, M.N.; Scott-Denton, L.E.; Schmidt, S.K.; Monson, R.K. The effects of tree rhizodeposition on soil exoenyzme activity, dissolved organic carbon, and nutrient availability in a subalpine forest ecosystem. Oecologia 2007, 154, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Toberman, H.; Freeman, C.; Evans, C.; Fenner, N.; Artz, R.R.E. Summer drought decreases soil fungal diversity and associated phenol oxidase activity in upland Calluna heathland soil. FEMS Microbiol. Ecol. 2008, 66, 426–436. [Google Scholar] [CrossRef] [PubMed]
- Šnajdr, J.; Valášková, V.; Merhautová, V.; Herinková, J.; Cajthaml, T.; Baldrian, P. Spatial variability of enzyme activities and microbial biomass in the upper layers of Quercus petraea forest soil. Soil Biol. Biochem. 2008, 40, 2068–2075. [Google Scholar] [CrossRef]
- Tian, L.; Dell, E.; Shi, W. Chemical composition of dissolved organic matter in agroecosystems: Correlations with soil enzyme activity and carbon and nitrogen mineralization. Appl. Soil Ecol. 2010, 46, 426–435. [Google Scholar] [CrossRef]
- Veres, Z.; Kotroczó, Z.; Fekete, I.; Tóth, J.A.; Lajtha, K.; Townsend, K.; Tothmeresz, B. Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability. Appl. Soil Ecol. 2015, 92, 18–23. [Google Scholar] [CrossRef]
- Decker, K.L.M.; Boerner, R.E.J.; Morris, S.J. Scale-dependent patterns of soil enzyme activity in a forested landscape. Can. J. For. Res. 1999, 29, 232–241. [Google Scholar] [CrossRef]
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setala, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Paterson, E. Comments on the regulatory gate hypothesis and implications for C-cyling in soil. Soil Biol. Biochem. 2009, 41, 1352–1354. [Google Scholar] [CrossRef]
- Jackson, C.R.; Liew, K.C.; Yule, C.M. Structural and functional changes with depth in microbial communities in a tropical Malaysian peat swamp forest. Microb. Ecol. 2009, 57, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Luis, P.; Kellner, H.; Zimdars, B.; Langer, U.; Martin, F.; Buscot, F. Patchiness and spatial distribution of laccase genes of ectomycorrhizal, saprotrophic, and unknown basidiomycetes in the upper horizons of a mixed forest Cambisol. Micrb. Ecol. 2005, 50, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Page-Dumroese, D.S.; Jurgensen, M.F.; Elliot, W.; Rice, T.; Nesser, J.; Collins, T.; Meurisse, R. Soil quality standards and guidelines for forest sustainability in northwestern North America. For. Ecol. Manag 2000, 138, 445–462. [Google Scholar] [CrossRef]
- Gao, X.B.; Liu, Z.W.; Pan, K.W.; Du, H.X.; Zhang, L.P. Preliminary studies on soil enzymes activities of typical forest ecosystem in Minjiang Areas. J. Northwest For. Uni. 2005, 20, 1–5. [Google Scholar]
- Awad, Y.M.; Blagodatskaya, E.; Ok, Y.S.; Kuzyakov, Y. Effect of polyacrylamide, biopolymer, and biochar on decomposition of soil organic matter and plant residues as determined by 14C and enzyme activities. Eur. J. Soil Biol. 2012, 48, 1–10. [Google Scholar] [CrossRef]
- Fontaine, S.; Mariotti, A.; Abbadie, L. The priming effect of organic matter: A question of microbial competition? Soil Biol. Biochem 2003, 35, 837–843. [Google Scholar] [CrossRef]
- John, D.; Baileya, J.D.; Mayrsohnb, C.; Doescherc, P.A.; Pierre, E.S.; Tappeiner, J.C. Understory vegetation in old and young Douglas-fir forests of western Oregon. For. Ecol. Manag. 1998, 112, 289–302. [Google Scholar] [CrossRef]
- Fekete, I.; Varga, C.; Kotroczó, Z.; Tóth, J.; Varbiro, G. The relation between various detritus inputs and soil enzyme activities in a Central European deciduous forest. Geoderma 2011, 167, 15–21. [Google Scholar] [CrossRef]
- Courty, P.E.; Pouysegur, R.; Bue´e, M.; Garbaye, J. Laccase and phosphatase activities of the dominant ectomycorrhizal types in a lowland oak forest. Soil Biol. Biochem. 2006, 38, 1219–1222. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tree Attributes | Site Attributes | ||||||
|---|---|---|---|---|---|---|---|
| Time Since Thinning (years) | Height (m) | Diameter at Breast Height (cm) | Elevation (m) | Depth of Mineral Soil (cm) | pH | Organic Matter (g/kg) | Nitrogen - (g/kg) |
| 24 | 6.79 | 10.36 | 879 | 53 | 5.05 ± 0.18 | 16.4 ± 1.0 | 0.348 ± 0.085 |
| 32 | 8.03 | 12.40 | 882 | 53 | 5.44 ± 0.08 | 26.9 ± 1.2 | 0.372 ± 0.017 |
| 40 | 9.21 | 14.12 | 910 | 52 | 5.45 ± 0.04 | 29.3 ± 1.3 | 0.432 ± 0.023 |
| Sources of Variation | Invertase | Urease | Hydrogen Peroxidase | Phenol Oxidase | ||||
|---|---|---|---|---|---|---|---|---|
| MS | p | MS | p | MS | p | MS | p | |
| Time since thinning | 0.743 | 0.031 | 0.028 | 0.037 | 2.093 | 0.001 | 0.007 | 0.0001 |
| Season | 2.199 | 0.044 | 0.028 | 0.036 | 2.920 | 0.002 | 0.003 | 0.0001 |
| Time since thinning × Season | 2.777 | 0.053 | 0.002 | 0.399 | 4.333 | 0.068 | 0.002 | 0.0001 |
| Error | 0.555 | 0.011 | 0.102 | < 0.001 | ||||
| Urease | Invertase | Catalase | Phenol Oxidase | |||||
|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | |
| Urease | 0.941 ** | 0.000 | −0.140 | 0.766 | −0.405 | 0.096 | ||
| Invertase | −0.080 | 0.595 | −0.261 | 0.058 | ||||
| Catalase | 0.861 * | 0.000 | ||||||
| Phenol oxidase | ||||||||
| Time Since Thinning (years) | Depth (cm) | Invertase (mg/g/day) | Urease (mg/g/day) | Catalase (mL/g/day) | Phenol Oxidase (µL/g/h) |
|---|---|---|---|---|---|
| 24 | 0–10 | 5.095 ± 0.138 ** | 0.319 ± 0.044 * | 2.525 ± 0.145 | 35.057 ± 4.016 |
| 24 | 10–20 | 1.973 ± 0.101 | 0.135 ± 0.039 | 2.231 ± 0.243 | 32.818 ± 3.872 |
| 32 | 0–10 | 5.021 ± 0.383 ** | 0.234 ± 0.078 * | 2.588 ± 0.084 | 70.807 ± 5.025 |
| 32 | 10–20 | 2.573 ± 0.243 | 0.169 ± 0.079 | 2.800 ± 0.191 | 66.680 ± 3.852 |
| 40 | 0–10 | 6.739 ± 0.231 * | 0.339 ± 0.130 | 2.255 ± 0.144 * | 15.743 ± 1.682 ** |
| 40 | 10–20 | 4.073 ± 0.160 | 0.248 ± 0.105 | 1.691 ± 0.207 | 7.542 ± 1.003 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Page-Dumroese, D.; Lv, R.; Xiao, C.; Li, G.; Liu, Y. Soil Enzyme Activities in Pinus tabuliformis (Carriére) Plantations in Northern China. Forests 2016, 7, 112. https://doi.org/10.3390/f7060112
Wang W, Page-Dumroese D, Lv R, Xiao C, Li G, Liu Y. Soil Enzyme Activities in Pinus tabuliformis (Carriére) Plantations in Northern China. Forests. 2016; 7(6):112. https://doi.org/10.3390/f7060112
Chicago/Turabian StyleWang, Weiwei, Deborah Page-Dumroese, Ruiheng Lv, Chen Xiao, Guolei Li, and Yong Liu. 2016. "Soil Enzyme Activities in Pinus tabuliformis (Carriére) Plantations in Northern China" Forests 7, no. 6: 112. https://doi.org/10.3390/f7060112
APA StyleWang, W., Page-Dumroese, D., Lv, R., Xiao, C., Li, G., & Liu, Y. (2016). Soil Enzyme Activities in Pinus tabuliformis (Carriére) Plantations in Northern China. Forests, 7(6), 112. https://doi.org/10.3390/f7060112