Abstract
Interior Douglas-fir is a prevalent forest type throughout the central Rocky Mountains. Past management actions, specifically fire suppression, have led to an expansion of this forest type. Although Douglas-fir forests cover a broad geographic range, few studies have described the interactive effects of various disturbance agents on forest health conditions. In this paper, we review pertinent literature describing the roles, linkages, and mechanisms by which disturbances, including insect outbreaks, pathogens, fire, and other abiotic factors, affect the development, structure, and distribution of interior montane forests primarily comprised of Douglas-fir. We also discuss how these effects may influence important resource values such as water, biodiversity, wildlife habitat, timber, and recreation. Finally, we identify gaps where further research may increase our understanding of these disturbance agents, their interacting roles, and how they influence long-term forest health.
1. Introduction
Disturbances exert strong influences over forest development and are expressed on a wide range of temporal and spatial scales [1,2,3]. Over the past century land management practices including timber harvesting, livestock grazing, and fire suppression have greatly altered disturbance regimes across the western USA. In mixed conifer forests the consequences have been increased tree densities, unnatural fuel accumulations and the expansion of fire-intolerant species (Figure 1) [4,5]. Particularly, this is the case where stand conditions are dissimilar to fire-adapted forests that historically had short-interval, low-severity surface fire regimes (e.g., ponderosa pine Pinus ponderosa Laws.) [6,7].

Figure 1.
Increased dead and down fuel following windthrow and Douglas-fir beetle colonization in a mixed conifer forest in the Dixie National Forest, Utah, USA (Photo: A. Giunta).
For example, the absence of frequent surface fires in some locations allowed for the expansion of shade-tolerant white-fir (Abies concolor Lindl.) and interior Douglas-fir (Pseudotsuga menziesii var. glauca Mirb. Franco) into open ponderosa pine stands. These species now form dense understories, effectively lowering canopy base heights, increasing ladder fuels, and elevating the hazard of high-severity fires [6,8].
Logging activities, including widespread clear-cutting during the 19th and early 20th century, throughout the interior west also created landscapes comprised of forests similar in size and age [9]. These forests are now reaching maturity, resulting in stands which are now suitable habitats for bark beetles [10]. In addition, warmer climate conditions favoring bark beetle success have led to an expansion of recent outbreaks, which have increased in severity and hectares infested [11,12]. A rise in bark beetle activity since the early 1990s has occurred across a range of forest types from low-elevation pinyon pine (Pinus edulis Engelm.) [13,14] to upper-elevation lodgepole pine (Pinus contorta Dougl. var. latifolia Engelm.) [15,16] and Engelmann spruce (Picea engelmannii Parry ex Engelm.) forests [17,18]. In interior Douglas-fir forests, the primary insect pest is the Douglas-fir beetle (DFB; Dendroctonus pseudotsugae Hopkins, Curculionidae: Scolytinae), which utilizes Douglas-fir exclusively [19,20].
Since the implementation of the Forest Ecosystems and Atmospheric Research Act of 1988 [21] and the Healthy Forest Restoration Act of 2003 [22], studies investigating the roles of both natural and anthropogenic disturbances on forest health degradation and associated impacts on wildlife habitat, timber production, water quality, recreation, aesthetics, grazing, and biodiversity have increased [23,24]. More recently, interest has grown in understanding how multiple and different disturbances will interact and affect a landscape [25]. In the Rocky Mountains, multiple studies on the interactive effects of fire and insects have been conducted with an emphasis in subalpine spruce-fir forests [26,27,28,29,30].
The complexity of the interactions between multiple disturbance agents in interior Douglas-fir forests and subsequent forest health effects has not readily been quantified or assessed. Thus, from a management perspective, it is important to understand how the potential interactions of multiple disturbances affect ecosystem patterns and processes, and how these in turn affect the vulnerability and susceptibility of forests within ecosystems to subsequent disturbances [31].
In this paper, we used the published literature to construct a synthesis of disturbance agents that primarily regulate vegetative dynamics within interior Douglas-fir forests in the central Rocky Mountains. These disturbances include DFB, western spruce budworm (WSBW; Choristoneura freeman Freeman, Lepidoptera: Tortricidae) and Douglas-fir dwarf mistletoe (Arceuthobium douglasii Engelm.). We focus on how the interactions of these disturbance agents influence the distribution, development, structure and health of interior Douglas-fir forests within the central Rocky Mountains. We start with a discussion of interior Douglas-fir forest ecology and the effects of abiotic disturbance agents: fire, wind, snow avalanches, and their effects on these forests. We then discuss the role of biotic agents including DFB, WSBW, Douglas-fir dwarf mistletoe, root diseases, and anthropogenic influences (e.g., fire management, logging) and how each affects the health of these forests and their role as inciting agents to other disturbances. Finally, we identify gaps in our understanding of these agents, their interactions, and their relationship to managing forest health. This information is designed to assist land managers with making ecologically-based decisions and devising appropriate strategies for long-term management of interior Douglas-fir forests.
2. Interior Douglas-Fir Forests
The composition and structure of interior Douglas-fir forests are rich and diverse due to the influences of a unique suite of biogeoclimatic, genetic, and disturbance factors [32]. The complex plant community assemblages in these forests can also be attributed in part to the broad ecological amplitude of the dominant overstory species Douglas-fir, which is one of the most widely distributed conifers in western North America [33,34,35] (Figure 2).
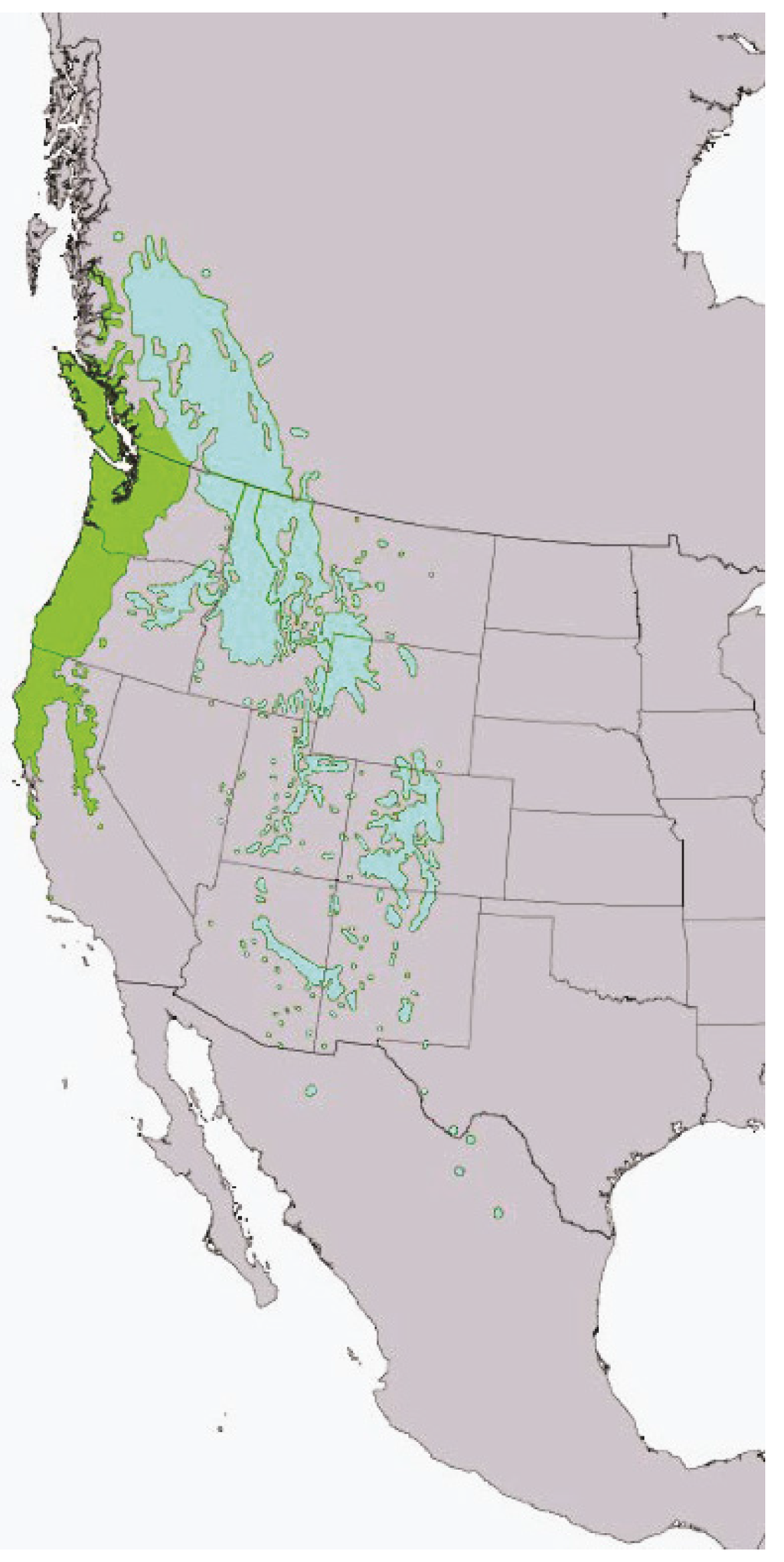
Figure 2.
Geographical distribution of coastal Douglas-fir (Pseudotsuga menziesii var. menziesii) outlined in green, and interior Douglas-fir outlined in blue. Digital representation from, [36].
Douglas-fir is highly adaptive to an array of site conditions that range across xeric to mesic gradients [35,37]. The geographic extent of the interior variety of this species extends from north-central British Columbia (55° N) to central Mexico (19° N) and is well established across an elevation range between 580 and 3500 m [38,39]. Throughout this range, climate and soil largely influence the site conditions of where this species will grow [40]. At southern latitudes, interior Douglas-fir distribution is limited by moisture availability, and is often restricted to north slopes at middle to high elevations in predominantly mesic sites [41,42]. For example, in the Santa Catalina Mountains of Arizona, Douglas-fir is the dominant conifer species above 2450 m [43].
In the northern portion of its range, the majority of the precipitation falls as snow, while in its southern distribution within the US (southern Utah, Arizona, New Mexico), precipitation is most abundant during the growing season, due to the influence of monsoonal moisture [44]. At northern latitudes, its growth is influenced by the length of the growing season and limited by cold temperatures [45]. The overall climate experienced by interior Douglas-fir throughout the central Rocky Mountains is characterized as a continental climate consisting of long, cold winters and hot, dry summers.
Unlike coastal Douglas-fir which is considered moderately shade-intolerant and is succeeded by more shade-tolerant western hemlock (Tsuga heterophylla Raf. Sarg.) and western red cedar (Thuja plicata Donn ex. D. Don), interior Douglas-fir is considered fairly shade-tolerant and is generally considered a climax species [46].
In the central Rocky Mountains, interior Douglas-fir is largely distributed within the mid-elevation montane zone which ranges between 900 and 1500 m [45]. In the overlapping montane and subalpine zones, Douglas-fir intermixes with spruce-fir forests dominated by subalpine-fir (Abies lasiocarpa Hook. Nutt.) and Engelmann spruce, with scattered pockets of limber pine (Pinus flexilis James) and Great Basin bristlecone pine (Pinus longaeva Bailey). At mid-elevations, Douglas-fir occurs with lodgepole pine and white fir [47,48]. At the lower end of its elevation range, Douglas-fir is often dispersed with ponderosa pine and woodlands comprised of piñon pine (Pinus edulis Engelm.), juniper (Juniperus spp.), bigtooth maple (Acer grandidentatum Nutt.), and Gambel oak (Quercus gambelii Nutt.) [49]. Common understory associates include ninebark (Physocarpus malvaceus Greene, Kuntze), mountain snowberry (Symphoricarpos oreophilus A. Gray) [50,51], chokecherry (Prunus virginiana L.), big sagebrush (Artemisia tridentata Nutt.), serviceberry (Amelanchier alnifolia Medik.), and currants (Ribes spp.) [47,51,52].
Commercially, interior Douglas-fir forests are an important resource for the forest products industry, providing lumber, plywood, house logs, and fuel wood [40,53]. Interior Douglas-fir forest communities also provide a critical wildlife habitat for a variety of bird species. These include Ruby Crowned Kinglets (Regulus calendula), Evening Grosbeaks (Coccothraustes vespertinus), Western Flycatchers (Empidonax occidentalis), and Northern Goshawks (Accipiter gentilis), which require habitats associated with mature forests [53,54].
3. Abiotic Disturbance Agents
3.1. Fire
Wildfires are one of the most important disturbance agents strongly influencing vegetative patterns across North America [55,56,57]. The effects of fire over a landscape are measured using a multitude of parameters including frequency, intensity, severity, and the spatial and temporal extent of a burn [58,59,60,61]. Collectively, these measures constitute the basis for describing an environments’ fire regime [62]. The most common method for classifying a fire regime is through a severity index which qualitatively describes how fire intensity affects an ecosystem, and is often related to the amount of biomass lost above and below ground [63]. High-severity fire regimes are characterized as those where fire transitions from surface fuels into the crowns of trees, consuming a majority of overstory vegetation [59]. Fire of this type is termed crowning when the fire is actively spreading from tree crown to tree crown [64,65]. In contrast, low-severity fire regimes are typified by frequent (4–30 year) low-intensity fires where surface fuels, including litter, moss, and herbaceous material, are charred or consumed while overstory canopy is minimally damaged or killed [63].
Interior Douglas-fir forests including those mixed with ponderosa pine throughout the central Rocky Mountains are characterized by a mixed-severity fire regime [37,66], one of the most complex and under-studied fire regimes in the western US [67,68]. Under this classification, forest stands experience natural fires across severity levels that range from low to medium to high [35], and with a variable fire return interval between 30 and 100 years [69]. The complexity of the fire regime is driven by the combined influence of both frequent low-severity surface fires and infrequent high-severity stand-replacing fires that create forest stand mosaics across the landscape varying in tree age and density [70,71,72]. Throughout the central Rocky Mountains, two prominent mixed conifer forest types dominate the landscape. The warm-dry type experiences more frequent non-lethal fires, and the cool-moist type experiences infrequent lethal fires that create even-age patches [73]. Overall, “individual mixed-severity fires typically leave a patchy, erratic pattern of mortality on the landscape, which fosters development of highly diverse communities” [69] (p. 226).
3.1.1. Direct Fire Effects
The direct effect of fire leads to either instantaneous tree mortality during initial fire passage, or delayed mortality resulting from severe injury through damage to foliage, cambium, fine roots, and conductive tissues, affecting physiological processes which are important for tree growth and development [74,75]. In the forest canopy, two types of crown damage determine the likelihood of fire-induced tree mortality. These include crown scorch, where the foliage is killed by hot gases above the flames, and crown consumption, where foliage and occasionally small twigs directly support combustion [76]. Two important parameters for predicting post-fire tree mortality associated with crown scorch include crown scorch volume and crown scorch height. Crown scorch volume is measured as the percent of the crown scorched [77,78,79] and crown scorch height is the level where heat is lethal to living foliage [76]. Scorch height is dependent upon fireline intensity, wind speed, and air temperature. The physiological effects of crown scorch can lead to a decrease in carbohydrate production, further weakening a tree’s response to stress and lowering its resistance to insects, drought, and other disturbances [80].
Heat-induced damage to tree boles can also affect a tree’s likelihood of survival after a wildfire [80,81]. Bole charring resulting in cambial death is dependent upon both the amount of heat received by a tree and the insulating capacity of the bark [81]. Older, large-diameter trees tend to have thicker bark with a greater capacity for absorbing heat, thus providing greater resistance to injury [82,83]. Bark of mature Douglas-fir is often comprised of a high percentage of cork, which can aid in the thermal diffusion of heat [81]. Douglas-fir stands with a greater proportion of large-diameter trees are likely to survive low-intensity fires. In the event bole scorch does not produce a fatal response through cambial injury, partial basal girdling and root damage may lead to moisture stress and reduced resistance to insects and diseases [79,84].
Sustained smoldering combustion of litter, duff, or downed woody material within surface and ground fuel layers can lead to root injury and mortality. Soil temperature, soil moisture, root spatial distribution, heat residence time, and fuel loading greatly influence the degree of root damage during a burn [75,85]. Temperatures as low as 48–60 °C have been attributed to root desiccation or death [86,87]. Swezy and Agee [88] found that prescribed surface fires in ponderosa pine stands in Oregon, USA , led to lethal temperatures (greater than 60 °C) that penetrated five centimeters in soil depth, affecting the greatest concentration of fine root mass (1–2 cm diameter). Other studies also concluded that both low- and high-severity burns reduce overall fine root mass [89,90]. Although these studies were conducted in ponderosa pine stands, interior Douglas-fir have shallow lateral roots that are also susceptible to fire damage [91]. Loss of root biomass can have significant implications for decreased essential nutrients and water, and can increase stress and susceptibility of affected stands to insects and diseases [92,93]. Furthermore, root systems anchor soil to prevent erosion, and a reduction or loss of root systems can increase runoff [94].
3.1.2. Indirect Fire Effects
Beyond direct mortality and consumption of forest biomass, fire can have many indirect effects on interior Douglas-fir forests. The disparity in fire severity is related to site influences including topography, aspect, and fuel loading [95], and will create unique fuel complexes in each stand influenced by microsite temperature, precipitation, fuel moisture content, stand densities, and the presence or absence of ladder fuels [96,97]. Variable fire intensities within the mixed-severity regime drive the composition of forests comprised of seral, fire-dependent species and mature fire-resistant species forming multistoried, mixed-aged stands [69,98]. At a landscape scale, even-age forest structures are most common where stand-replacing fires are prevalent, and between these even-age patches are mixed-aged stands where frequent surface fires are dominant [99].
Human activities during Euro-American settlement in the western US altered fire regimes in interior Douglas-fir forests. Throughout the past century, fire suppression actions stemming from fire exclusion policies dating back to the early 20th century have reshaped the landscape. These actions led to extended fire-free periods in montane forests which have allowed understory conifers to develop in formerly open stands [100] (Figure 3).

Figure 3.
(A) Dense regeneration of white-fir and interior Douglas-fir saplings in the understory of an unmanaged interior Douglas-fir stand in the Dixie National Forest, Utah, USA (Photo A: A. Giunta); (B) A managed interior Douglas-fir stand in the Ashley National Forest, Utah, USA (Photo B: D. Malesky, USDA Forest Service, Forest Health Protection, Ogden, UT, USA).
In a fire reconstruction study in southwestern Montana, an increase in Douglas-fir density within grassland-sage communities coincided with the exclusion of surface fires due to anthropogenic influences starting in the mid-1880s [101]. The escalation in understory conifer growth has contributed to an increase in ladder fuels, providing a mechanism for the transition of surface fires into forest canopies, increasing the potential for crowning and the occurrence of high-severity, stand-replacing fires [69]. Fire exclusion–induced changes in forest composition, structure, and fuel loads affect interior Douglas-fir stands and may alter historic low-severity fire regimes [102]. Episodic droughts coupled with dense canopy cover, close intercrown distances and large fuel accumulations create environments that are conducive to extreme fire weather favoring the initiation and spread of crown fires.
3.2. Additional Abiotic Disturbances
Wind and snow avalanches are two additional natural disturbance agents that influence forest composition, structure, and forest soils [103,104,105,106]. Primary effects of wind include damage and breakage to the tops of crowns, branch breakage, uprooting, and snapping of trees [107]. Wind events can create canopy gaps that vary in size from a few individual trees to landscape scale (hundreds of hectares) [108]. Gap openings increase available light used by surrounding trees for increased growth, or benefit suppressed trees in the understory [105]. Furthermore, windthrown trees create, “suitable bark beetle habitat, increase fuel loads, and limit mobility of wildlife and forest recreationists” [109] (p. 446). Uprooted trees also expose soil and creates heterogeneity in soil properties [110].
Forest snow avalanches are typically small in size, but large infrequent events can be destructive [111]. Avalanche paths form where there is an abundance of snowfall through natural storm deposition or wind transport in steep terrain (greater than 30° slope angles), allowing for the release and acceleration of a snowslide [112,113]. Much like wind, snow avalanches affect forests through the breakage of stems and branches, uprooting, and the creation of severe wounds on the uphill side of trees [114,115,116]. Large infrequent avalanches often kill overstory trees with little impact on regeneration and can create a rapid buildup of large, coarse, woody material that gets deposited throughout an avalanche run-out zone [117,118,119].
Although these two disturbance agents are most closely associated with subalpine and tree line forest zones [120,121], wind events and avalanches do at times affect the interior Douglas-fir zone. McGregor et al. [122] reported a strong wind event in November 1981, which blew down thousands of trees in forests in Idaho, USA. Many windthrown Douglas-fir were selected for use in trials of the DFB anti-aggregate pheromone MCH (3-methyl-2-cyclohexen-1-one) following this event. Within the Wasatch Mountains of Utah, USA, snow avalanches that occurred in interior Douglas-fir forests created extensive debris piles that subsequently became infested by DFB. Trapping and MCH application by US Forest Service Forest Health Protection staff were employed to prevent DFB spread into neighboring stands [123,124,125] (Figure 4).

Figure 4.
(A) Snow avalanche path through an interior Douglas-fir forest in the Ashley National Forest, UT, USA (Photo A: D. Blackford, USDA Forest Service, Forest Health Protection, Ogden, UT, USA); (B) Interior Douglas-fir debris in the avalanche runout zone, Ashley National Forest, UT, USA. (Photo B: D. Blackford).
4. Biotic Disturbances
4.1. Douglas-Fir Beetle
Forest stand development patterns in montane interior Douglas-fir forests are not regulated by fire alone. Bark beetles are also a major disturbance agent that have a large ecological role in reshaping forests [12,24]. Interactions between bark beetles and their hosts have coevolved over the past 200 million years [126]. Insect-induced tree mortality influences the development, senescence, and rebirth of stands, which in turn affects energy flows and nutrient cycles [127]. Dendroctonus species (Coleoptera: Curculionidae, Scolytinae) are particularly capable of reshaping stand structure, composition, and function [128,129,130]. Endemic populations attack old, large, and weakened trees, which removes trees from the overstory and promotes the recruitment of the next generation of trees in a stand [12]. Periodically, epidemic populations occur and are able to kill live, healthy trees in great numbers [131,132].
Within interior Douglas-fir forests, the most prevalent bark beetle species is the DFB [19,20,123,133]. Density-independent factors that influence the population dynamics of this insect include the availability and suitability of host trees, weather conditions, and disturbances (e.g., windthrow, avalanches) that produce downed host material [134,135]. Often, freshly felled or downed trees greater than 20 cm in diameter, contain sufficiently thick phloem with essential nutrients that are required for successful brood production [136,137]. Newly felled trees typically lack effective defense mechanisms including a decrease or cessation of resin production which makes them attractive targets. Resin is a key compound containing monoterpenes and sesquiterpenes that entrap or elevate toxin levels fatal to beetles or pathogens vectored by beetles [138,139]. In previous studies, trees with low oleoresin pressure have been associated with an increased susceptibility to bark beetle attacks [140]. Consequently, high DFB population increases are often related to disturbances including windthrow or snow avalanches that produce an abundance of downed trees [141], or trees physiologically weakened by drought [142], fire [84,143], ice damage [144], defoliation [145], and diseases [146,147]. Evidence of successful host colonization is determined by the presence of entrance holes, emergence holes, egg galleries, and frass accumulations on the bole and near the base of trees [133,148,149].
Stand and bark beetle population dynamics are often highly interrelated [150]. Coulson [150] stated, “Tree age, diameter, and phloem thickness are all correlated and in turn are related to beetle survival, i.e., the large-diameter trees with thick phloem accommodate large beetle populations and have high survival rates” (p. 433). In stands with mixed diameter classes, the percentage of trees killed during outbreaks is related to tree diameter, with the greatest number of trees being killed in the diameter class representing the highest basal area [151].
Unlike lodgepole pine or spruce/fir forests which typically form dense, uniform, even-age forests in the central Rocky Mountains as a result of stand-replacing fire or logging, the intrinsic characteristics of interior Douglas-fir forests combined with the generally less aggressive nature of DFB limit landscape-scale DFB-induced mortality. Throughout the central Rocky Mountains, interior Douglas-fir stands often occur in mixed-species stands, or in small groups surrounded by non-host trees at the edge of their upper and lower elevation limits. Thus, suitable DFB hosts are typically distributed unevenly throughout a forest [152]. Where Douglas-fir is a dominant overstory component, past logging and fire history have created forest mosaics in which Douglas-fir oscillates in age and density, limiting the extent of potential hosts [135]. Typically, small groups of trees are attacked [153]. However, under certain conditions, drought coupled with a supply of recently downed trees can facilitate the development of DFB populations from endemic into epidemic levels. At this stage of an outbreak, DFB are able to overcome host resistance and initiate attacks on standing live trees, where groups of 100 or more can become infested [129,153] (Figure 5).

Figure 5.
Interior Douglas-fir stand conditions before (A); during (B); and following (C) Douglas-fir beetle colonization in Utah, USA. (A) An uninfested phase—green (G); (B) currently infested phase—red (R); and (C) older mortality phase—gray (GY). (Photo A: M. Jenkins, B: A. Giunta, C: M. Jenkins).
A stand hazard rating system developed by Weatherby and Thier [154] for southern Idaho, USA, suggests that the highest potential for tree mortality due to a DFB outbreak is in stands with basal areas greater than 23.2 m2/ha, a proportion of Douglas-fir greater than 50%, an average stand age above 120 years, and an average diameter at breast height greater than 50 cm.
4.2. Western Spruce Budworm
Insect defoliators have an important influence on the condition of interior Douglas-fir forests. Some of the most important impacts of insect defoliation are tree mortality, rotation delays, and increased susceptibility to secondary insects and disease [155]. The WSBW is considered one of the most widespread and destructive defoliators in western coniferous forests, particularly where Douglas-fir and true firs are the primary tree species in a stand [156,157,158]. Depending on environmental and biological conditions, the timing of WSBW outbreaks is highly variable. The periodicity and duration of outbreaks can range from two to over 35 years [159,160].
The life history requirements for WSBW are highly interdependent upon forest stand structure and conditions. This insect preferentially feeds on the current years’ growth where larvae penetrate swelling buds that have the highest food quality and offer the best protection from predators [145,161]. Bud phenology, specifically bud development and the timing of budburst, can greatly influence WSBW population dynamics. Trees that exhibit delayed budburst have been associated with a reduction in WSBW success as new bud formation occurs after second instar larvae emerge from hibernation and initiate feeding [162]. Douglas-fir forests with a large proportion of trees that are genetically predisposed to delayed budburst will likely have a greater resistance to WSBW infestations. Site location can also affect biological processes that influence WSBW survival. Sites with warm soils and warmer microclimates have been linked to earlier budburst timing [163]. Dry sites situated along south and west aspects where earlier bud development coincides with larvae feeding after winter emergence can be associated with higher larvae survival. Often, stands with fewer host trees have been shown to have lower levels of mortality within various size classes [132].
Multi-age, multi-level forest canopies in stands dominated by host trees provide optimal WSBW habitat as second instar larvae are dependent upon a successful canopy descent to reach host resources [164]. Weather factors including wind also exert a large control over the success rate of locating a suitable host. During the past 50 years, singular overlapping and repeated outbreaks of WSBW have greatly altered the structure and composition of montane forests along the Colorado Front Range [132].
The greatest impact of WSBW within infested stands is on subcanopy and understory layers where larvae feed on host regeneration within the understory (Figure 6). Conifer seedlings and saplings have relatively few needles and buds, and new growth can become deformed or killed by only a few larvae [157]. In one study, Hadley and Veblen [165] used dendrochronological analysis to reconstruct past WSBW and DFB attacks throughout the Colorado Front Range. Results from their study indicated WSBW outbreaks were responsible for high (greater than 50%) mortalities of seedlings, saplings, and small-diameter trees. Future regeneration within a stand is further impeded by WSBW feeding on developing cones and seeds [166]. Frank and Jenkins [167] found that a higher percentage of larvae feed on seed cones as opposed to pollen cones. This could affect future regeneration since Douglas-fir is known to have infrequent cone crops every two to seven years at lower elevations [168], and every one in 11 years at higher elevations [169].

Figure 6.
Western spruce budworm defoliation on interior Douglas-fir saplings in southern Idaho, USA. (Photo: Carl Jorgensen, USDA Forest Service, Forest Health Protection, Boise, ID, USA).
Western spruce budworm also negatively affects overstory host trees. Consecutive years of feeding can lead to decreased stem growth, top kill, and, in some cases, tree mortality [170]. A loss in tree volume due to decreased growth rates can lead to an overall decrease in a stand’s basal area, which could impact timber harvest projections if merchantable stands were to become infested.
Often, the absence and, more importantly, release patterns of growth rings in mature Douglas-fir and other host species (e.g., white fir), coincide with WSBW outbreaks [171,172]. Using tree-ring reconstructions, Swetnam and Lynch [172] found that overstory trees in Devil’s Gulch, located in northern Colorado, USA, experienced 60% mortality from WSBW feeding. In another tree-ring study conducted near Pemberton, British Columbia, Canada, Alfaro et al. [173] found a 39% reduction in the number of host trees per hectare within three years after a WSBW outbreak. Most sampled stands experienced host growth reduction evidenced by reduced tree ring widths during WSBW outbreaks. This study also indicated older, less vigorous stands with suppressed Douglas-fir trees were most susceptible to WSBW infestations. In mixed-species stands such as Douglas-fir/ponderosa pine, WSBW outbreaks tend to shift species dominance towards ponderosa pine [120,165].
4.3. Douglas-Fir Dwarf Mistletoe
Dwarf mistletoes (Arceuthobium spp.) are one of the most important, widespread disease agents in North American conifer forests, and are found throughout montane forest ecosystems [174,175,176]. All mistletoe species are host specific and slow spreading, making stand composition, tree size, and structure important for their persistence in a forest community [177]. The plants form obligate hemiparisitic relationships with host plants, extracting vital water and minerals through haustorium from their hosts [178,179]. This process depletes essential photosynthetic reserves used for growth and maintenance by host trees [176,180]. Although dwarf mistletoes are capable of complete photosynthesis, upwards of 60% of their carbohydrates can be extracted from their hosts [179]. Tree response to infection results in dense abnormal growth of host twigs that form branch clusters termed witches’ brooms [181]. This irregular growth pattern changes branch structure, function, and can eliminate cone production by infected branches [182,183]. Additional degenerative, induced effects on host plants include stem and height growth reductions, top kill, and reduced forest productivity [181,184].
Douglas-fir dwarf mistletoe is the most damaging species that parasitizes Douglas-fir [174,185]. Spread of this disease is initiated through the movement of the parasite to previously uninfected branches of a single tree or between trees [184]. The female plant produces fruit and seed that mature in fall (September, October). Seeds are under high internal water pressure within the fruit which, when abscised from the parent plant, are explosively propelled through the air at upwards of 22 m per s. Seeds contain a sticky viscin coating which allows them to attach to hosts [186]. Spread rates of Douglas-fir dwarf mistletoe are often accelerated in multi-storied Douglas-fir stands where understory trees receive abundant seed rain from infected overstory trees, as reported in southwestern interior Douglas-fir stands [184]. When Douglas-fir is the climax member of the community, as often is the case in interior Douglas-fir stands, there is typically not a shortage of hosts for Douglas-fir dwarf mistletoe, which can persist unless a severe disturbance leads to the loss of its host species [181]. Seedlings and saplings, especially those with main stem infections, readily succumb to this parasite [186]. The presence of non-host species can slow the spread of the disease agent. Stands with open canopies are often more susceptible compared to dense stands, because stands with high densities create shading conditions which retard Douglas-fir dwarf mistletoe growth [186].
4.4. Root Diseases
Pathogens and, in particular, root diseases are an important component in forest ecosystems, and they exert strong influences over forest dynamics including structure, composition, and function [187,188]. Within a stand, fungi spread via spores transported by rhizomorphs in the soil, or through direct root contact between infected and uninfected hosts [189]. Specifically, fungi infect the cambial tissues of roots and root collars where root tissues have evidence of staining and decay. The foliage of root-diseased trees typically appears chlorotic and thin. Trees lose needles from the lower crown upward, and from the inside (near the stem) outward [190]. Trees sometimes respond to infection by producing copious amounts of resin near the base of the stem [191]. Trees may also produce a stress cone crop in response to infection [189]. Eventually, damage to roots leaves trees girdled and host trees die. Connections between infection sites create root disease centers characterized by circular openings in the main canopy that range from approximately one-tenth of a hectare to 400 or more hectares in size [189]. These “mortality centers” are associated with groups of dying and dead conifers. One issue with root diseases is that they are persistent in a stand, and can survive as saprophytes on dead wood material for decades [189]. The severity of infestation is often amplified by disturbances including fire suppression and logging where fungi can colonize stumps and roots of cut trees and eventually spread to healthy trees [187,190,192].
In the Rocky Mountains, Armillaria spp., and specifically Armillaria ostoyae, is the most important and widespread of all root pathogens [193,194]. It has a broad host range including Abies and Pinus species. In dry, interior conifer forests, it aggressively infects interior Douglas-fir, colonizing and killing healthy trees in all age classes [190]. The patterns of Armillaria spread follow two main pathways: either via distinct mortality patches, circular in nature with mortality mostly confined to the leading edge of the patch, or as dispersed mortality, forming continuous coverage over a site [195,196].
This pathogen greatly affects forest community structure. Once established, a slow progression of the fungi into non-infected portions of a stand creates an initial pulse of mortality. Once trees along the edge die, canopy gaps are created which benefit the regeneration of the next tree cohorts. Seedlings and saplings are also vulnerable to this disease which can retard the development of the stand. Surviving trees, once reaching maturity, become a vital resource that can support further development of the disease which continually cycles its way through a stand, forming a wave pattern of mortality [195].
Species composition within a stand can be affected by the creation of canopy gaps. In the northern Rockies (interior British Colombia, Canada), gaps created by the mortality of interior Douglas-fir associated with Armillaria may become filled with more disease-resistant and shade-tolerant western hemlock, western red cedar, or even subalpine fir, though this species is also susceptible to Armillaria [187,191]. In the central Rocky Mountains, Douglas-fir which is often the climax overstory species in its community would likely continue to persist in a stand.
Once Armillaria is established, infected trees become susceptible to windthrow or fall over on their own from weakened root systems [141,193]. This can lead to an increase in hazardous trees if root disease centers are located in developed recreation areas including campgrounds or trailheads [197]. Furthermore, root disease also affects tree species not infected by creating openings in forests where healthy trees along gap margins can be exposed to high winds and subsequent windthrow [141].
5. Disturbance Interactions
Interactions between disturbance agents including fire, DFB, WSBW, Douglas-fir dwarf mistletoe, and root diseases affect the health of interior Douglas-fir forests, and can predispose forests to subsequent disturbances (Figure 7). Specific disturbance agent interactions are addressed below.
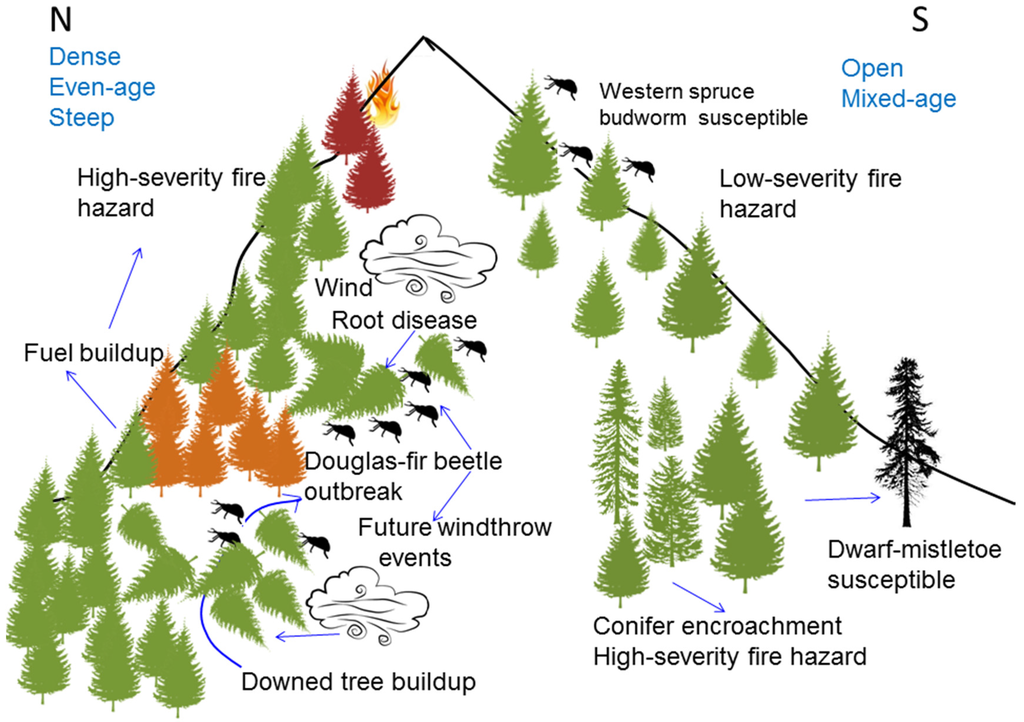
Figure 7.
Conceptual schematic of interacting biotic and abiotic disturbance agents and associated forest health issues in interior Douglas-fir forests. (Illustration: A. Giunta).
5.1. Fire and Douglas-Fir Beetle
Following non-stand-replacing fire events, wildfire impacts can predispose stands to subsequent bark beetle disturbances. Trees impacted by crown scorch, bole charring, and root damage associated with fire become attractive targets for DFB [198,199]. Furniss [137] reported 70% of fire-injured Douglas-fir were infested by DFB following a fire in southern Idaho, USA. The proportion of attacks was highest in trees that experienced low-to-moderate cambium injury. Trees with high levels of cambium injury experienced fewer attacks as phloem and other essential resources were damaged beyond utilization by beetles. In other studies, DFB mass attacked trees with 25%–50% cambial damage and greater-than-50% crown scorch [200,201]. Colonization patterns following fire progress from fire-damaged trees to healthy live trees over time as suitable phloem resources became scarce in successive fire-damaged classes [143,200]. Trees completely defoliated by crown fires also result in the complete burning or severe scorching of the inner bark, especially in thin-barked trees, and were not suitable for bark beetle use [198,199].
Bark beetle attack dynamics affect forest structure differently compared to fire. With low-intensity fire, often smaller-diameter and younger tree cohorts are killed, while larger-diameter trees survive, because “temperatures in the plume at the height of the canopy are too low” [202] (p. 483). Thus, mature trees within a stand continue to produce seeds contributing to regeneration. With bark beetle attacks, the Douglas-fir beetle seeks old, large-diameter trees, which are also mature, seed-producing trees resulting in stands with a younger age class, and often a reduction in reproductive output [203,204,205]. Following low-intensity fire, canopy structure is likely to remain intact aside from occasional torched trees. Surviving trees maintain shade cover, which can increase soil moisture retention benefiting regeneration and other established vegetation. Furthermore, periodic low-intensity fire reduces the amount of surface fuels and decreases the overall fuel load in a forest. In contrast, bark beetle colonization contributes to an increase in litter loading when dead needles begin to fall from a tree, a one- to four-year period post-attack [203]. Increased amounts of coarse woody debris can accumulate on the forest floor following overstory tree mortality. The loss of overstory trees creates canopy openings that favor the growth of herbaceous vegetation and grasses [11,129].
5.2. Douglas-Fir Beetle and Forest Fuel Changes
Only recently have the interactive effects between bark beetle–induced changes and fuel complexes been studied thoroughly. Much of the recent research is associated with forest systems that experience infrequent high-severity fire regimes (e.g., lodgepole pine forests infested by mountain pine beetle [206,207], and Engelmann spruce forests infested by spruce beetle [29,208]. Jenkins et al. [129] provide an extensive review of fuel complex changes during typical bark beetle rotations in these forest types. It has been hypothesized that DFB-induced alterations to fuel complexes will differ from upper elevation forest types due to drier sites, more open stand conditions, lower biomass loads, and lower tree and canopy base heights [37,57,209]. The spatial pattern of DFB-caused tree mortality also complicates how fuel complexes change within stands.
During a DFB outbreak, the most notable changes begin in the canopy where dead tree foliage begins to desiccate, fading from green to yellow and finally to red. Dead needles drop to the forest floor one to four years post-colonization where in increase of fine surface fuels occurs (litter, woody material less than 7.62 cm in diameter) through branch and canopy breakage. With the loss of overstory cover, a slight increase in herbaceous material follows as increased sunlight reaches the forest floor. Over time (20 years or longer), dead, standing beetle-killed snags begin to fall, increasing the amount of large woody material (greater than 7.62 cm) in the surface fuels layer (Figure 8) [37,129,210].
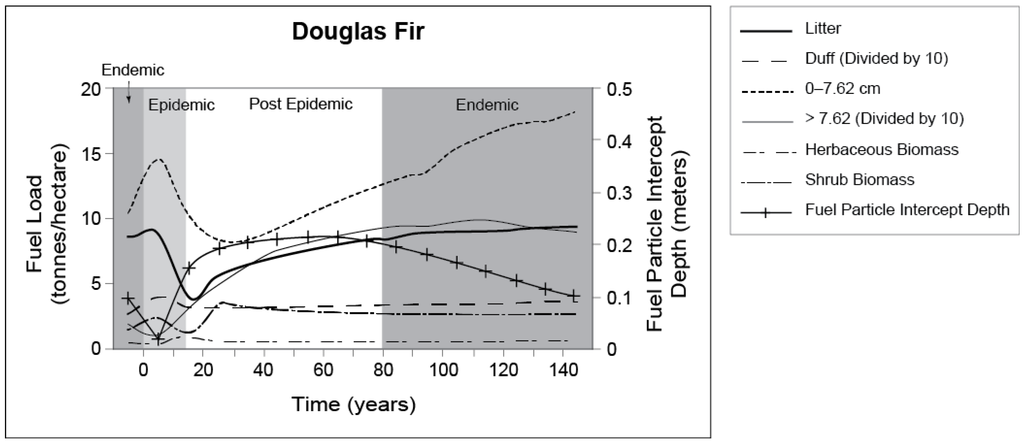
Figure 8.
Select fuel characteristic type changes in interior Douglas-fir forests over the course of a Douglas-fir beetle rotation. (Figures and data from Jenkins et al. [129]).
Donato et al. [37] inventoried changes to surface and aerial fuels in interior Douglas-fir forests across four different DFB outbreak stages. This included green (unattacked), red (recent mortality, one to three years post-attack), gray (older mortality, 4–14 years post-attack), and silver (older mortality, 25–30 years post-attack) across a range of Douglas-fir habitats in the Greater Yellowstone Ecosystem (GYE). The results from their study indicated that significant reductions in available canopy fuel load and bulk density occurred as time increased post-outbreak (4–14 years), while significant changes to surface fuels were minor, aside from an increase in 1000 h fuels during the silver stage. In a similar study conducted in northern Utah, Giunta [210] found that in surface fuels, a significant increase in litter depth (cm) and litter loading (kg/m3) was associated with needle loss during the time bark beetle–killed trees began losing red needles. With canopy loss, an increase in herbaceous biomass followed in stands where the majority of trees were grey as increased levels of light were able to infiltrate the forest floor. In other studies, DFB-induced changes to interior Douglas-fir stands resulted in the basal area of 40%–70%, a reduction in the mean diameter at breast height of 8%–40%, and a three-fold increase in grasses and herbaceous plants in infested stands [21,211].
Alterations to canopy fuels have been attributed to a series of physiological changes directly resulting from the colonization of host trees by beetles and the introduction of the associated blue-stain fungi, principally the species Ophiostoma pseudotsugae (Rumb.) von Arx (Ceratostomella pseudotsugae Rumbold) [212]. Blue-stain fungi penetrate the sapwood and phloem tissues adjacent to larval galleries, contributing to the disruption of water transport between root systems and foliage [139]. Needles desiccate over time and foliage fades from green to red [37]. It is during the red needle stage where the probability of torching and crowning can increase as canopy foliar moisture declines to its lowest moisture content level and dead needles are still retained in the canopy [37,129].
Giunta [210] measured foliar moisture content in infested Douglas-fir trees across all four crown condition classes (green, green-infested, yellow, and red) associated with DFB outbreaks. His findings showed that yellow and red crown condition classes had a significantly (p < 0.0001) lower foliar moisture content based on percent of oven-dry weight compared to green and green-infested foliage.
5.3. Western Spruce Budworm and Fire
Many open-canopy stands that were once maintained by frequent surface fires have been replaced by densely-stocked, closed canopy stands composed of mixed-age shade-tolerant species across the landscape compared to pre-settlement forested landscapes [171,213].
The effects of WSBW on fuel loads, fire occurrence, and fire behavior are not fully understood, but are certainly different from DFB [214]. Western spruce budworm directly alters aerial fuels by consuming the current year’s needles, whereas DFB does not feed on needles, and needle loss occurs once a tree is dead. A reduction in canopy bulk density decreases the likelihood of torching and crown fire initiation and spread [215]. Although no studies on the effect of WSBW on fire and fuel loads have been reported for the central Rocky Mountains, research has been conducted in Douglas-fir stands in the Pacific Northwest, USA. Flower et al. [216] examined dendrochronological records of fire and WSBW outbreaks in interior Douglas-fir forests in Oregon, USA, and western Montana, USA. Their research showed no synchronous pattern between WSBW outbreaks and increased fire occurrence. In another study after a WSBW outbreak in Washington, USA, Hummel and Agee [217] measured decreases in canopy closure and a reduction in the density of small-diameter (less than 20 cm) trees over an eight-year period. Incorporating these inputs from stand data, they used the fire behavior model BEHAVE [218] to simulate fire spread in their study site. Crown characteristics that contributed to crown fire initiation (e.g., canopy base height and canopy bulk density) remained stable and did not indicate any significant increase in crown or torch potential. In a similar study (central Oregon to western Montana, USA), Gavin et al. [214] used the Wildland-Urban Interface Fire Dynamic Simulator (WFDS) to model fine-scale fuel changes associated with WSBW infestations. They found defoliation consistently reduced both the vertical and horizontal spread of crown fire across a range of surface fire intensities. They also discovered that a row of defoliated trees released substantially less heat compared to a row of non-defoliated trees due to the lack of fuel and associated decrease in flame intensities. Hummel and Agee [217] also found that coarse, woody fuel loads increased by 50% following a WSBW outbreak in the central Cascades, USA. Their plot data inputted into surface fire models predicted a significant increase in surface fire flame lengths. Site-specific fuel models are important to achieve better fire behavior predictions [218].
5.4. Dwarf Mistletoe, Fire, and Other Disturbances
Episodic natural fires help prevent Douglas-fir dwarf mistletoe from spreading by continually removing infected overstory trees from stands and killing infected and uninfected understory tree hosts [209]. Ingrowth and expansion of Douglas-fir due to the lack of fire have increased the abundance of susceptible hosts. Stand density increases have raised fuel accumulations and increased the spread of Douglas-fir mistletoe across the interior west [186].
Douglas-fir dwarf mistletoe impacts on infected trees primarily affect canopy fuels of infected trees. Koonce and Roth [219] reported 73% greater aerial fuels (live and dead witches’ brooms) in dwarf mistletoe–infested stands compared to non-infested stands. Witches’ brooms typically form in the lower portion of tree crowns. This growth formation of closely spaced small branches traps fallen needles, creating vertically oriented fine fuels and increasing stand flammability. The development of vertical ladder fuels provides a mechanism for surface fire to transition into crowns, increasing the wildfire hazard in interior Douglas-fir stands [176,209,220]. Dwarf mistletoe in Douglas-fir forests on the Bitterroot National Forest in western Montana likely contributed to the high fire intensity observed during the summer of 2000 [221].
The interaction of Douglas-fir dwarf mistletoe with agents of disturbance other than fire can also adversely affect forest health. The combined impacts of Douglas-fir dwarf mistletoe and WSBW, for example, can increase seedling and sapling mortality rates or elevate the susceptibility of mature trees to DFB. An increase in overall tree mortality can increase fuel accumulations contributing to poor stand health [222].
5.5. Western Spruce Budworm and Douglas-Fir Beetle
Previous insect disturbances in a stand can serve as initial stressors to tree vigor, which subsequently diminishes tree defenses and increases the likelihood of successful DFB colonization [147,165]. As an example, in 2011, DFB-caused tree mortality in southern Idaho, USA, Nevada, USA, and Utah, USA more than doubled [223]. This trend continued through 2013, the last year of reported data from the US Forest Service Forest Health Protection Program. This increase was partly attributed to stress induced by several years of WSBW defoliation [224]. In one study it was found that trees in Colorado, USA, infested by DFB had reduced growth attributed to a previous WSBW outbreak [225]. Negrón [132] (p. 82) mentioned, “In the Colorado Front Range, it seems that the primary disturbance agent, although not the only one that triggers DFB outbreaks, is WSBW defoliation”. In Logan Canyon, Utah, USA, Fredericks and Jenkins [145] found that trees defoliated by WSBW reduced host tree defenses against DFB. Their observation is consistent with other research [132,165,226]. It also has been found that other defoliators including the Douglas-fir tussock moth (Orgyia pseudotsugata McDunnough, Lepidoptera: Lymantriidae) decrease plant vigor, predisposing hosts to subsequent DFB attack [147,227].
5.6. Root Disease, Windthrow, and Douglas-Fir Beetle
The presence of root disease can predispose a stand to additional disturbance agents which can result in tree mortality and decreased stand health. Root pathogens such as Armillaria infect healthy trees, which decreases plant vigor and predisposes trees to attack by insects [189]. Armillaria primarily weakens root systems, leaving infected trees highly susceptible to windthrow [228]. Wind-felling frequently triggers bark beetle epidemics where they take advantage of abundant breeding material which becomes available following such events [205].
Endemic Douglas-fir beetle populations are often correlated with root diseases [122], which can lead to subsequent build-up of downed trees following wind events. The abundance of fresh slash material enables DFB populations to increase and mass attack surrounding standing live trees, facilitating the development of epidemic populations [229].
6. Anthropogenic Influences
6.1. Historic Human Mediated Disturbance
In addition to natural disturbances, anthropogenic influences cause changes within interior Douglas-fir forests. Human-mediated disturbances have also influenced the present structure and composition of interior Douglas-fir forests. Prior to European settlement, Native Americans indigenous to the central Rocky Mountains used fire as a tool for improving wildlife habitat and hunting grounds [230,231]. These activities likely influenced local fire patterns in interior Douglas-fir forests. Fire type (ground, surface, or canopy), frequency, and extent of Native American fire use and the subsequent effect on altering fire regimes have been debated [231]. Although the scale of their fire practices is likely smaller than originally thought, elucidating the degree to which fire was intentionally used by Native Americans to manipulate interior Douglas-fir forests has proven difficult to assess as conventional fire reconstruction methods are limited in their abilities to differentiate historic natural ignitions from human-caused ignitions [232].
Human-mediated disturbance within these forests increased with the arrival of European settlers. During the mid- to late-19th century, many settlers along the Wasatch Front, Utah, USA, utilized local timber from Douglas-fir forests to construct homesteads, towns, waterwheels, and other goods [233]. Unrestricted logging practices left many slopes devoid of trees, leading to the development of even-age stand structures that currently exist throughout this forest type within the Wasatch Mountains [234].
Throughout the mid-20th century, interior Douglas-fir has remained a viable commercial species [45]. The wood is extremely strong and used as structural timber, railroad ties, plywood, and for pulp [43,235]. The impacts of logging vary depending on the size, intensity, and type of harvesting practices employed (e.g., group tree selection versus clear-cutting) [9]. Logging, in some cases, has led to a more homogeneous stand structure, higher tree densities, lack of structural diversity, and loss of old, mature trees [99]. In other cases, previously logged stands have had a number of large trees left uncut, which resulted in a similar volume of large mature trees found in nearby unlogged stands [99].
6.2. Forest Restoration
Early forest management practices were conducted on the premise of a limitless supply of resources. This type of mindset contributed to forest degradation following land use practices including, logging, grazing, and fire suppression [71,236]. Throughout the 21st century, natural resource managers have been shifting management strategies from sole resource extraction to include restoration principles. Often, the goals of forest restoration are to improve the resiliency and ecosystem function of a stand, and return it to a state within the historic range of conditions prior to Euro-American influence [71,237]. Treatments are often considered in cases where anthropogenic activities have greatly altered stand structure, ecosystem function, and composition [236]. In interior Douglas-fir and other forests characterized as having a mixed-severity fire regime, the complex mosaic of forest structures present across the landscape make it difficult to implement effective restoration plans. Even with good intentions, some restoration procedures can create unintentional forest health issues.
6.2.1. Pre-Fire Restoration Treatments
Fire management practices, primarily fuel treatment prescriptions, have greatly influenced forest health. Often, the focus of fire management objectives is to reduce hazardous fuels by decreasing stand density and ladder fuels through thinning [65,238]. These methods are effective at reducing overall canopy bulk density and increasing canopy base height in a stand, which reduces the hazard of crown fire. Fuel reduction treatments, however, can result in unintended consequences including exacerbating the incidence and severity of root diseases [239]. Mechanical damage to tree boles during tree removal operations may also weaken trees, leaving them more susceptible to insect infestation and infection by decay fungi [240]. For example, increasing the proportion of large-diameter trees when stands are thinned from below can increase their susceptibility to DFB [204].
6.2.2. Fire Restoration Treatments
Prescribed fire is another fire management tool widely used to reduce dense accumulations of fuels, remove logging debris, improve wildlife habitat, and manage vegetation [59,241]. Before the era of aggressive fire suppression, periodic fire maintained vegetative diversity on the landscape which helped mitigate the effects of bark beetle attacks. Since then, the lack of fire has created more uniform stands capable of supporting the spread of bark beetles or enhancing the effects of defoliators [220,242]. The reintroduction of fire after years of suppression can have unintended forest health consequences. These fires can burn with greater intensity and severity, especially where tree mortality has increased due to insects and diseases. Widespread tree mortality also causes dead fuels to accumulate for decades, increasing the hazard of high-intensity fire over time [243]. This might particularly be true at mid-elevations (2400–2700 m) where stands that had a naturally mixed-severity fire regime are now altered by fire suppression and transition into higher elevation forests. This may increase the potential for high-intensity fires ignited at lower elevations to burn into higher elevation stands where hazardous fuel accumulations may result in unnatural levels of fire damage and more severe resource effects [240].
6.2.3. Post-Fire Restoration Treatments
A common practice following wildfire is salvage logging, which involves the removal of fire-killed trees to recover economic value before degradation by decay [244]. The impact of these activities can lead to increased sediment erosion [245], loss of snags that provide wildlife habitat, and shifts in wildlife composition toward bird and invertebrate species that prefer more open habitats [245]. In some instances, post-wildfire logging may actually increase future short-term fire risk through the rapid accumulation of coarse, woody debris associated with salvage logging activities. Furthermore, soil disturbance and logging slash can inhibit seedling growth and result in a net reduction in post-fire regeneration [246]. Sanitation harvesting is another technique used during post-fire remediation and involves the removal of both live and dead trees affected by fire. Live fire-damaged trees are often removed to decrease future insect infestations.
7. Future Research Needs
The complex and heterogeneous nature of interior Douglas-fir forests often makes assessing forest health difficult, and devising appropriate management strategies challenging. Maintaining ecosystem function, enhancing biodiversity, and reducing the risk of catastrophic fires in these forests necessitates that management plans consider a holistic, integrated, and adaptive approach. Implementing such an approach requires further research to better understand the effects of multiple disturbance agents on interior Douglas-fir forest communities [247]. We have identified several gaps where further research may increase our understanding of disturbance agents, their interacting roles, and influences on long-term forest health.
- (1)
- Forest conditions including “stand structure, fuel characteristics, and fire regimes” have been greatly altered in part due to management practices that started in the 20th century, where “forest structure, fuel characteristics, and fire regimes of the mixed-conifer forests in the western United States have been dramatically altered” [248,249] (p. 22). One important issue is understanding the historic role and extent of fire in these ecosystems. The present accumulations of live and dead fuels have resulted in a shift toward higher-severity fire behavior. Although high-severity crown fires are not outside the historical range of variability for this fire regime type, the frequency and overall size of these types of fires appear to be increasing [66]. Additional fire reconstruction studies across various geographic scales can help elucidate the natural fire regimes of interior montane forests [231,250,251,252].
- (2)
- The inherent variability of interior Douglas-fir stands occurring on sites ranging from xeric to mesic and from lower-montane to subalpine zones necessitates that forest managers have a better understanding of how geographic locales influence forest fuel conditions. As stand composition and structure have shifted, so too have changes to fuel complexes.
- (3)
- Research is needed to determine how forest insects, particularly bark beetles and defoliators, affect fuels and fire behavior across a wider range of geographic locales and whether bark beetle alterations of surface and canopy fuels can elevate the potential for fire to spread into upper elevation forests.
- (4)
- Little is known about the combined effects of DFB and WSBW on interior Douglas-fir forests and, in turn, on ecosystem values across various spatial scales.
- (5)
- Information regarding climate change influences on the occurrence, timing, frequency, extent, and duration of disturbances at various temporal and spatial scales for interior Douglas-fir forests is lacking.
- (6)
- A greater understanding of DFB population dynamics in interior Douglas-fir stands is needed.
- (7)
- Douglas-fir dwarf mistletoe and DFB interactions are poorly understood.
- (8)
- Research on how snow avalanches and other abiotic disturbances affect interior Douglas-fir forest health is lacking.
8. Conclusions
Interior Douglas-fir forests are a principal forest type in the central Rocky Mountains. The adaptability of Douglas-fir to a variety of site conditions across a broad latitudinal range results in unique and diverse plant communities that provide for numerous ecosystem and social values. Both natural disturbance processes, including fire, wind, insect outbreaks, pathogens, and human-mediated disturbances will continue to have an important role in shaping these forest communities.
Disturbance-induced alterations to the mixed-severity fire regime characteristic of this forest type will continue to exert a large influence over future stand development. Past forest management policies promoted the advancement of more insect and pathogen outbreaks, modifying the susceptibility to future disturbance events (e.g., windthrow, landslides, snow avalanches), and will continue to affect overall forest health. Current and future forest management decisions should weigh the costs and benefits of implementing certain practices (e.g., harvesting, prescribed fire) so that these activities do not adversely affect the health in certain, already vulnerable interior Douglas-fir forests.
The diversified nature of interior Douglas-fir forest communities discourages a “one size fits all” approach to management. Rather, the management of interior Douglas-fir forests in the future will necessitate developing more holistic, integrated, and adaptive management strategies to maintain forest and ecosystem health to meet multiple management objectives.
Acknowledgments
Funding for this project was provided by the Joint Fire Science Program (Project #11-1-4-16) and supported by the Ecology Center and the Utah Agricultural Experiment Station at Utah State University. We give special thanks to Michaela Teich for assistance with manuscript preparation and review, Wanda Lindquist for graphics assistance, and Laura Dunning for manuscript formatting. The comments from four anonymous reviewers have greatly improved the final manuscript.
Author Contributions
Andrew Giunta was the primary author of this paper and conducted the literature review. Elizabeth Hebertson contributed to the writing, editing, and assisted with literature review. Michael Jenkins provided edits and reviews. Allen Munson provided edits, contributed to the writing, and reviewed all manuscript drafts.
Conflicts of Interest
The authors declare no conflict of interest. The funding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Oliver, C.D. Forest development in North America following major disturbances. For. Ecol. Manag. 1981, 3, 153–168. [Google Scholar] [CrossRef]
- Delcourt, H.R.; Delcourt, P.A.; Webb, T., III. Dynamic plant ecology: The spectrum of vegetational change in space and time. Quat. Sci. Rev. 1983, 1, 153–175. [Google Scholar] [CrossRef]
- White, P.S.; Pickett, S.T.A. Natural disturbance and patch dynamics: An introduction. In The Ecology of Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 3–13. [Google Scholar]
- Parsons, D.J.; DeBenedetti, S.H. Impact of fire suppression on a mixed-conifer forest. For. Ecol. Manag. 1979, 2, 21–33. [Google Scholar] [CrossRef]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western U.S. forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar] [CrossRef] [PubMed]
- Fulé, P.Z.; Covington, W.W.; Moore, M.M. Determining reference conditions for ecosystem management of southwestern ponderosa pine forests. Ecol. Appl. 1997, 7, 894–908. [Google Scholar] [CrossRef]
- Allen, C.D.; Savage, M.; Falk, D.A.; Suckling, K.F.; Swetnam, T.W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T. Ecological restoration of southwestern ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Cocke, A.E.; Fulé, P.Z.; Crouse, J.E. Forest change on a steep mountain gradient after extended fire exclusion: San Francisco Peaks, Arizona, USA. J. Appl. Ecol. 2005, 42, 814–823. [Google Scholar] [CrossRef]
- Hejl, S.J.; Hutto, R.L.; Preston, C.R.; Finch, D.M. Effects of silvicultuer treatments in the Rocky Mountains. In Ecology and Management of Neotropical Migratory Birds: A Synthesis and Review of Critical Issues, 1st ed.; Martin, T.E., Finch, D.M., Eds.; Oxford University Press: New York, NY, USA, 1995; pp. 220–244. [Google Scholar]
- Bentz, B. Bark beetle outbreaks in western North America. Causes and consequences. In Proceedings of the Bark Beetle Symposium, Snowbird, UT, USA, 15–17 November 2005.
- Raffa, K.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Turner, M.G.; Romme, W.H. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: The dynamics of bark beetle eruptions. BioScience 2008, 58, 501–517. [Google Scholar] [CrossRef]
- Bentz, B.J.; Régnière, J.; Hansen, M.; Hayes, J.L.; Hicke, J.A.; Kelsey, R.G.; Negrón, J.F.; Seybold, S.J. Climate change and bark beetles of the western United States and Canada: Direct and indirect effects. BioScience 2010, 60, 602–613. [Google Scholar] [CrossRef]
- Shaw, J.D.; Steed, B.E.; DeBlander, L.T. Forest inventory and analysis (FIA) annual inventory answers the question: What is happening to pinyon-juniper woodlands? J. For. 2005, 103, 280–285. [Google Scholar]
- Kleinman, S.J.; DeGomez, T.E.; Snider, G.B.; Williams, K.E. Large-scale pinyon ips (Ips confusus) outbreak in the southwestern United States tied with elevation and land cover. J. For. 2012, 110, 194–200. [Google Scholar] [CrossRef]
- Wulder, M.A.; White, J.C.; Bentz, B.; Alvarez, M.F.; Coops, N.C. Estimating the probability of mountain pine beetle red-attack damage. Remote Sens. Environ. 2006, 101, 150–166. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G.; Rampley, G.J.; Neilson, E.T.; Carroll, A.L.; Ebata, T.; Safranyik, L. Mountain pine beetle and carbon forest feeback to climate change. Nature 2008, 452, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, M.J.; Hebertson, E.G.; Munson, A.S. Spruce beetle biology, ecology and management in the Rocky Mountains: An addendum to spruce beetle in the Rockies. Forests 2014, 5, 21–71. [Google Scholar] [CrossRef]
- Page, W.G.; Jenkins, M.J.; Runyon, J.B. Spruce beetle-induced changes to Engelmann spruce foliage flammability. For. Sci. 2014, 60, 691–702. [Google Scholar] [CrossRef]
- Schmitz, R.F.; Gibson, K.E. Douglas-Fir Beetle; Forest and Insect Disease Leaflet; USDA Forest Service: Washington, DC, USA, 1996; pp. 5–7.
- Negrón, J.F.; Anhold, J.A.; Munson, S.A. Within-stand spatial distribution of tree mortality caused by the Douglas-fir beetle (Coleoptera: Scolytidae). Environ. Entomol. 2001, 30, 215–224. [Google Scholar] [CrossRef]
- Kolb, T.E.; Wagner, M.R.; Covington, W.W. Forest health from different perspectives, forest health through silviculture. In Proceedings of the 1995 National Silviculture Workshop, Mescalaro, NM, USA, 8–11 May 1995; pp. 5–13.
- Healthy Forest Restoration Act. “HR 1904”. PL108-148; U.S. Department of Agriculture, 2003. Available online: http://www.fs.fed.us/emc/applit/includes/hfr2003.pdf (accessed on 2 March 2016).
- Coulson, R.N.; Stephens, F.M. Impacts of insects in forest landscapes: Implications for forest health management. In Invasive Forest Insects, Introduced Forest Trees, and Altered Ecosystems: Ecological Pest Management in Global Forests of a Changing World; Payne, T.D., Ed.; Springer-Verlag: New York, NY, USA, 2006; pp. 101–125. [Google Scholar]
- Fettig, C.J.; Klepzig, K.D.; Billings, R.F.; Munson, S.A.; Nebeker, T.E.; Negrón, J.F.; Nowak, J.T. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. For. Ecol. Manag. 2007, 238, 24–53. [Google Scholar] [CrossRef]
- Turner, M.G. Disturbance and landscape dynamics in a changing world. Ecology 2003, 84, 362–371. [Google Scholar]
- Kulakowski, D.; Veblen, T.T.; Bebi, P. Effects of fire and spruce beetle outbreak legacies on the disturbance regime of a subalpine forest in Colorado. J. Biogeogr. 2003, 30, 1445–1456. [Google Scholar] [CrossRef]
- Kulakowski, D.; Veblen, T.T. Effects of prior disturbances on the extent and severity of wildfire in Colorado subalpine forests. Ecology 2007, 88, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Bigler, C.; Kulakowski, D.; Veblen, T.T. Multiple disturbance interactions and drought influence fire severity in Rocky Mountain subalpine forests. Ecology 2005, 86, 3018–3029. [Google Scholar] [CrossRef]
- Jorgensen, C.A.; Jenkins, M.J. Fuel complex alterations associated with spruce beetle-induced tree mortality in intermountain spruce-fir forests, USA. For. Sci. 2011, 57, 232–240. [Google Scholar]
- O’Connor, C.D.; Lynch, A.M.; Falk, D.A.; Swetnam, T.W. Post-fire forest dynamics and climate variability affect spatial and temporal properties of spruce beetle outbreaks on a Sky Island mountain range. For. Ecol. Manag. 2015, 336, 148–162. [Google Scholar] [CrossRef]
- Kulakowski, D.; Jarvis, D.; Veblen, T.T.; Smith, J. Stand replacing fires reduce susceptibility of lodgepole pine to mountain pine beetle outbreaks in Colorado. J. Biogeogr. 2012, 39, 2052–2060. [Google Scholar] [CrossRef]
- Meidinger, D.; Pojar, J. Ecosystems of British Columbia; British Columbia Ministry of Forests, 1991; p. 330. Available online: http://www.for.gov.bc.ca/hfd/pubs/Docs/Srs/SRseries.htm (accessed on 18 April 2015).
- Arno, S.F. Forest Regions of Montana; Research Paper INT-218; USDA Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1979; p. 39.
- Silen, R.R. Genetics of Douglas-Fir; Research Paper WO-35; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1978; p. 34.
- Simard, S.W. The foundational role of mycorrhizal networks in self-organization of interior Douglas-fir forests. For. Ecol. Manag. 2009, 258, 95–107. [Google Scholar] [CrossRef]
- Little, E.L., Jr. Atlas of United States Trees; Vol. 1, U.S. Department of Agriculture: Washington, DC, USA, 1971; pp. 1–9.
- Donato, D.C.; Harvey, B.J.; Romme, W.H.; Simard, M.; Turner, M.G. Bark beetle effects on fuel profiles across a range of stand structures in Douglas-fir forests of Greater Yellowstone. Ecol. Appl. 2013, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Fowells, H.A. Silvics of Forest Trees of the United States; No. 2171; USDA Forest Service Agriculture Handbook: Washington, DC, USA, 1965; pp. 546–556.
- Wright, J.W.; Kung, F.H.; Read, R.A.; Lemmien, W.A.; Bright, J.N. Genetic variation in Rocky Mountain Douglas-fir. Silv. Genet. 1971, 20, 54–60. [Google Scholar]
- Van Hooser, D.D.; Wadell, K.L.; Mills, J.R.; Tymcio, R.P. The interior Douglas-fir resource: Current status and projections to the year 2040. In Interior Douglas-Fir, the Species and Its Management, Proceedings of the Interior Douglas-fir: The Species and Its Management, Pullman, WA, USA, 27 February–1 March 1991; Baumgartner, D.M., Lotan, J.E., Eds.; Washington State University: Pullman, WA, USA, 1991; pp. 9–14. [Google Scholar]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina mountains Arizona. I. Ecological classification and distribution of species. J. Ariz. Acad. Sci. 1964, 3, 9–34. [Google Scholar] [CrossRef]
- Whittaker, R.H.; Niering, W.A. Vegetation of the Santa Catalina mountains, Arizona. V. biomass, production, and diversity along the elevation gradient. Ecology 1975, 56, 771–790. [Google Scholar] [CrossRef]
- Niering, W.A.; Lowe, C.H. Vegetation of the Santa Catalina Mountains, Arizona, USA: Community types and dynamics. Vegetatio 1984, 58, 3–28. [Google Scholar]
- Lavendar, D.P.; Hermann, R.K. Douglas-Fir: The Genus Pseudotsuga, 1st ed.; Oregon State University: Corvallis, OR, USA, 2014; pp. 19–24. [Google Scholar]
- Hermann, R.K.; Lavendar, D.P. Pseudotsuga menziesii (Mirb.) Franco Douglas-fir. In Silvics of North American Conifers; Burns, R.M., Honkala, B.H., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1990; Volume 1, pp. 527–540. [Google Scholar]
- Kilgore, B.M. Fire in ecosystem distribution and structure: Western forests and scrublands. In Fire Regimes and Ecosystem Properties; General Technical Report WO-GTR-26; Mooney, H.A., Bonnickson, T.M., Christensen, N.L., Lotan, J.E., Reiners, W.A., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1981; pp. 58–89. [Google Scholar]
- Bailey, R.G. Descriptions of the Ecoregions of the United States; Miscellaneous Publication 1391; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1980; p. 77.
- Eyre, F.H. Forest Cover Types of the United States and Canada, 1st ed.; Society of American Foresters: Washington, DC, USA, 1980; p. 148. [Google Scholar]
- Harlow, W.M.; Harrrar, E.S.; White, F.M. Textbook of Dendrology: Covering the Important Forest Trees of the United States and Canada, 6th ed.; McGraw-Hill: New York, NY, USA, 1979; p. 510. [Google Scholar]
- Habeck, J.R.; Mutch, R.W. Fire dependent forests in the northern Rocky Mountains. Quat. Res. 1973, 3, 408–424. [Google Scholar] [CrossRef]
- Mauk, R.L.; Henderson, J.A. Coniferous Forest Habitat Types of Northern Utah; General Technical Report INT-170; U.S. Department of Agriculture, Forest Service, Intermountain Range and Experiment Station: Ogden, UT, USA, 1984.
- Youngblood, A.P.; Mauk, R.L. Coniferous Forest Habitat Types of Central and Southern Utah; General Technical Report INT-187; Department of Agriculture, Forest Service, Intermountain Range and Experiment Station: Ogden, UT, USA, 1985.
- Keegan, C.E.; Swanson, L.D.; Wichman, D.P.; VanHooser, D.D. Montana’s Forest Products Industry: A Descriptive analysis 1969–1988; Bureau of Business and Economic Research: Missoula, MT, USA, 1990; p. 52. [Google Scholar]
- Lanner, R.M. Trees of the Great Basin—A Natural History, 1st ed.; University of Nevada Press: Reno, NV, USA, 1984; p. 215. [Google Scholar]
- Lilleholm, R.J.; Kessler, W.B.; Merrill, K. Stand density index applied to timber and goshawk habitat objectives in Douglas-fir. Environ. Manag. 1993, 17, 773–779. [Google Scholar] [CrossRef]
- McKenzie, D.; Gedalof, Z.; Peterson, D.L.; Mote, P. Climate change, wildfire, and conservation. Conserv. Biol. 2004, 18, 890–902. [Google Scholar] [CrossRef]
- Hicke, J.A.; Johnson, M.C.; Hayes, J.L.; Preisler, H.K. Effects of bark beetle-caused tree mortality on wildfire. For. Ecol. Manag. 2012, 271, 81–90. [Google Scholar] [CrossRef]
- Jenkins, M.J.; Page, W.G.; Hebertson, E.G.; Alexander, M.E. Fuels and fire behavior dynamics in bark beetle attacked forests in western North America and implications for fire management. For. Ecol. Manag. 2012, 275, 23–34. [Google Scholar] [CrossRef]
- Pyne, S.J.; Andrews, P.L.; Laven, R.D. Introduction to Wildland Fire, 2nd ed.; Wiley: New York, NY, USA, 1996; p. 769. [Google Scholar]
- Brown, J.K. Introduction and fire regimes. In Wildland Fire in Ecosystems: Effects on Fire on Flora; Brown, J.K., Smith, J.K., Eds.; General Technical Report RMRS-42; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000. [Google Scholar]
- Morgan, P.; Hardy, C.C.; Swetnam, T.W.; Rollins, M.G.; Long, D.G. Mapping fire regimes across time and space: Understanding coarse and fine-scale fire patterns. Int. J. Wildland Fire 2001, 10, 329–324. [Google Scholar] [CrossRef]
- Halofsky, J.E.; Donato, D.C.; Hibbs, D.E.; Cambell, J.L.; Cannon, M.D.; Fontaine, J.B.; Thompson, J.R.; Anthony, R.G.; Bormann, B.T.; Kayes, L.J.; et al. Mixed-severity fire regimes: Lessons and hypotheses from Klamath-Siskiyou Ecoregion. Ecosphere 2011, 2, 1–19. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire intensity, fire severity and burn severity: A brief review. Int. J. Wildland Fire 2009, 18, 116–126. [Google Scholar] [CrossRef]
- Van Wagner, C.E. Conditions for the start and spread of crown fire. Can. J. For. Res. 1977, 7, 23–34. [Google Scholar] [CrossRef]
- Agee, J.K.; Skinner, C.N. Basic principles of forest fuel reduction treatments. For. Ecol. Manag. 2005, 211, 83–96. [Google Scholar] [CrossRef]
- Schoennagel, T.; Veblen, T.T.; Romme, W. Interaction of fire, fuels, and climate across Rocky Mountain forests. BioScience 2004, 54, 661–676. [Google Scholar] [CrossRef]
- Chappell, C.B.; Agee, J.K. Fire severity and tree seedling establishment in Abies magnifica forests, southern Cascades, Oregon. Ecol. Appl. 1996, 6, 628–640. [Google Scholar] [CrossRef]
- Agee, J.K. The complex nature of mixed-severity fire regimes. In Mixed Severity Fire Regimes: Ecology and Management; Lagene, L., Zelnik, J., Cadwallader, S., Hughes, B., Eds.; Washington State University: Pullman, WA, USA; The Association for Fire Ecology: Spokane, WA, USA, 2004; pp. 1–10. [Google Scholar]
- Arno, S.F.; Parsons, D.J.; Keane, R.E. Mixed-Severity Fire Regimes in the Northern Rocky Mountains: Consequences of Fire Exclusion and Options for the Future; Research Paper RMRS-P-15-VOL-5; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2000; pp. 225–232.
- Brown, J.K. Fire regims and their relevance to ecosystem manaagment. In Proceedings of the Society of American Foresters National Convention, Anchorage, AK, USA, 18–22 September 1994; Society of American Foresters: Bethesda, MD, USA, 1995; pp. 171–178. [Google Scholar]
- Baker, W.L.; Veblen, T.T.; Sherriff, R.L. Fire, fuels, and restoration of ponderosa pine-Douglas-fir forests in the Rocky Mountains, USA. J. Biogeogr. 2007, 34, 251–269. [Google Scholar] [CrossRef]
- Jain, T.B.; Battaglia, M.A.; Han, H.S.; Graham, R.T.; Keyes, C.R.; Fried, J.S.; Sandquist, J.E. A Comprehensive Guide to Fuel Management Practices for Dry Mixed Conifer Forests in the Northwestern United States: Inventory and Model-Based Economic Analysis of Mechanical Fuel Treatments; Research Note RMRS-RN-64; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2014.
- Romme, W.H.; Floyd, M.L.; Hanna, D. Historical Range of Variability and Current Landscape Condition Analysis: South Central Highlands Section, Southwestern Colorado & Northwestern New Mexico; Colorado Restoration Institute, Colorado State University: Fort Collins, CO, USA; Region 2 U.S. Department of Agriculture, Forest Service: Golden, CO, USA, 2009; Available online: http://www.coloradoforestrestoration.org/CFRIpdfs/2009_HRVSouthCentral-Highlands.pdf (accessed on 1 March 2016).
- Wyant, J.G.; Omni, P.N.; Laven, R.D. Fire induced tree mortality in a Colorado ponderosa pine/Douglas-fir stand. For. Sci. 1986, 32, 49–59. [Google Scholar]
- Stephens, S.L.; Finney, M.A. Prescribed fire mortality of Sierra Nevada mixed conifer tree species: Effects of crown damage and forest floor combustion. For. Ecol. Manag. 2002, 162, 261–271. [Google Scholar] [CrossRef]
- Van Wagner, C.E. Height of crown scorch in forest fires. Can. J. For. Res. 1973, 3, 373–378. [Google Scholar] [CrossRef]
- Methven, I.R. Prescribed Fire, Crown Scorch and Mortality: Field and Laboratory Studies on Red and White Pine; Research Report PS-X-31; Canadian Forest Service, Petawawa Forest Experimental Station: Chalk River, ON, Canada, 1971; p. 10. [Google Scholar]
- Peterson, D.L. Crown scorch volume and scorch height: Estimates of post-fire tree condition. Can. J. For. Res. 1985, 15, 596–598. [Google Scholar] [CrossRef]
- Chambers, J.L.; Dougherty, P.M.; Hennessey, T.C. Fire: Its effects on growth and physiological processes in conifer forests. In Stress Physiology and Forest Productivity, 1st ed.; Hennessey, T.C., Dougherty, P.M., Kossuth, S.V., Johnson, J.D., Eds.; Martinaus Nijhoff: Hingham, MA, USA, 1986; pp. 171–191. [Google Scholar]
- Peterson, D.L.; Ryan, K.C. Modeling postfire conifer mortality for long-range planning. Environ. Manag. 1986, 10, 797–808. [Google Scholar] [CrossRef]
- Ryan, K.C.; Reinhardt, E.D. Predicting post fire severity in seven western conifers. Can. J. For. Res. 1988, 18, 1291–1297. [Google Scholar] [CrossRef]
- Peterson, D.L.; Arbaugh, M.J. Postfire survival in Douglas-fir and lodgepole pine: Comparing the effects of crown and bole damage. Can. J. For. Res. 1986, 16, 1175–1179. [Google Scholar] [CrossRef]
- Costa, J.J.; Oliveria, L.A.; Viegas, D.X.; Neto, L.P. On the temperature distribution inside a tree under fire conditions. Int. J. Wildland Fire 1991, 1, 87–96. [Google Scholar] [CrossRef]
- Hood, S.; Bentz, B. Predicting postfire Douglas-fir beetle attacks and tree mortality in the northern Rocky Mountains. Can. J. For. Res. 2007, 37, 1058–1069. [Google Scholar] [CrossRef]
- Hungerford, R.D.; Harrington, M.G.; Frandsen, W.H.; Ryan, K.C.; Niehoff, G.J. Influence of fire on factors that affect site productivity. In Proceedings of the Management and Productivity of Western-Montane Forest Soils, Boise, ID, USA, 10–12 April 1990; Harvey, A.E., Neuenschwander, L.F., Eds.; General Technical Report INT-280. U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1991; pp. 32–50. [Google Scholar]
- Neary, D.G.; Klopatek, C.C.; DeBano, L.F.; Ffolliott, P.F. Fire effects on below ground sustainability: A review and synthesis. For. Ecol. Manag. 1999, 122, 51–71. [Google Scholar] [CrossRef]
- Egan, D. Protecting Old Trees from Prescribed Burning; Working Paper No. 24; Ecological Restoration Institute: Flagstaff, AZ, USA, 2011; p. 5. [Google Scholar]
- Swezy, D.M.; Agee, J.K. Prescribed fire effects on fine-root and tree mortality in old-growth ponderosa pine. Can. J. For. Res. 1991, 21, 626–634. [Google Scholar] [CrossRef]
- Stendell, E.; Horton, T.R.; Bruns, T.D. Early effects of prescribed fire on the structure of the fungal ectomycorrhizal community in a Sierra Nevada ponderosa pine forest. Mycol. Res. 1999, 103, 1353–1359. [Google Scholar] [CrossRef]
- Hart, S.C.; Classen, A.T.; Wright, R.J. Long-term interval burning alters fine root and mycorrhizal dynamics in a ponderosa pine forest. J. Appl. Ecol. 2005, 42, 752–761. [Google Scholar] [CrossRef]
- Rogstad, A. Recovering From Wildfire a Guide to Arizona’s Forest Owners. 2002. Available online: http://ag.arizona.edu/pubs/natresources/az1294/ (accessed on 1 March 2016).
- Gordon, W.S.; Jackson, R.B. Nutrient concentrations in fine roots. Ecology 2000, 81, 275–280. [Google Scholar] [CrossRef]
- Reubens, B.; Poesen, J.; Danjon, F.; Geudens, G.; Muys, B. The role of fine and coarse roots in shallow slope stability and soil erosion control with a focus on root system architecture: A review. Trees 2007, 21, 385–402. [Google Scholar] [CrossRef]
- Gyssels, G.; Poesen, J.; Bochet, E.; Li, Y. Impact of plant roots on the resistance of soils to erosion by water: A review. Prog. Phys. Geogr. 2005, 29, 189–217. [Google Scholar] [CrossRef]
- Lertzman, K.; Riccius, P.; Fall, J. Does fire isolate elements of the landscape mosaic? Bull. Ecol. Soc. Am. 1997, 78, 92. [Google Scholar]
- Brown, J.K. The unnatural fuel build up issue. In Proceedings of the Symposium and Workshop on Wilderness Fire, Missoula, MT, USA, 15–18 November 1983; Lotan, J.E., Kilgore, B.M., Fischer, W.C., Mutch, R.W., Eds.; General Technical Report INT-182. U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1983; pp. 127–128. [Google Scholar]
- Turner, M.G.; Romme, W.H. Landscape dynamics in crown fire ecosystems. Landsc. Ecol. 1994, 9, 59–77. [Google Scholar] [CrossRef]
- Agee, J.K. The landscape ecology of western forest fire regimes. Northwest Sci. 1998, 72, 24–34. [Google Scholar]
- Kaufmann, M.R.; Regan, C.M.; Brown, P.M. Heterogeneity in ponderosa pine/Douglas-fir forests: Age and size structure in unlogged and logged landscapes of central Colorado. Can. J. For. Res. 2000, 30, 698–711. [Google Scholar] [CrossRef]
- Covington, W.W.; Moore, M.M. Post settlement changes in natural fire regimes and forest structure: Ecological restoration of old-growth ponderosa pine forests. J. Sustain. For. 1994, 2, 153–181. [Google Scholar] [CrossRef]
- Heyerdahl, E.K.; Miller, R.F.; Parson, R.A. History of fire and Douglas-fir establishment in a savanna and sagebrush-grassland mosaic, southwestern Montana, USA. For. Ecol. Manag. 2006, 230, 107–118. [Google Scholar] [CrossRef]
- Daniels, L.D. Climate and fire: A case study of the Cariboo forest, British Columbia. In Proceedings of the Symposium on the Ecology and Management of Mixed Severity Fire Regimes Conference, Spokane, WA, USA, 17–19 November 2004; Taylor, L., Zelnik, J., Cadwallader, S., Hughes, B., Eds.; pp. 235–246.
- Schaetzl, R.F.; Burns, S.F.; Johnson, D.L.; Small, T.W. Tree uprotting: Review of impacts on forest ecology. Vegetatio 1989, 79, 65–176. [Google Scholar]
- Patten, R.S.; Knight, D. Snow avalanches and vegetation pattern in Cascade Canyon, Grand Teton National Park. Arct. Alp. Res. 1994, 26, 35–41. [Google Scholar] [CrossRef]
- Mitchel, S.J. A diagnostic framework for windthrow risk estimation. For. Chron. 1998, 74, 100–105. [Google Scholar] [CrossRef]
- Ulanova, N.G. The effects of windthrow on forests at different spatial scales. A review. For. Ecol. Manag. 2000, 135, 155–167. [Google Scholar] [CrossRef]
- Bouget, C.; Duelli, P. The effects of windthrow on forest insect communities: A literature review. Biol. Conserv. 2004, 118, 281–299. [Google Scholar] [CrossRef]
- Everham, E.M., III; Browkaw, N.V.L. Forest damage and recovery from catastrophic wind. Bot. Rev. 1996, 62, 114–175. [Google Scholar] [CrossRef]
- Mitchel, S.J. The windthrow triangle: A relative windthrow hazard assessment procedure for forest managers. For. Chron. 1995, 71, 446–450. [Google Scholar] [CrossRef]
- Beaty, S.W.; Stone, E.L. The variety of microsites created by tree falls. Can. J. For. Res. 1986, 16, 539–548. [Google Scholar] [CrossRef]
- Teich, M.; Bartlet, P.; Gret-Regamey, A.; Bebi, P. Snow avalanches in forested terrain: Influence of forest parameters, topography, and avalanche characteristics on runout distance. Arct. Antarct. Alp. Res. 2012, 44, 509–519. [Google Scholar] [CrossRef]
- Luckman, B.H. The geomorphic activity of snow avalanches. Geografiska Annaler. Series A. Phys. Geogr. 1977, 59, 31–48. [Google Scholar]
- McClung, D.; Schaerer, P. The Avalanche Handbook, 3rd ed.; The Mountainers Books: Seattle, WA, USA, 2006; p. 338. [Google Scholar]
- Burrows, C.L.; Burrows, V.L. Procedures for the study of snow avalanche chronology using growth layers of woody plants. Inst. Arct. Alp. Res. 1976, 23, 54. [Google Scholar]
- Hebertson, E.G. Snow Avalanche Disturbance in Intermountain Spruce-Fir Forests and Implications for the Spruce Bark Beetle (Coleoptera: Scolytidae). Ph.D. Thesis, Utah State University, Logan, UT, USA, 2004. [Google Scholar]
- Malanson, G.P.; Butler, D.R. Transverse pattern of vegetation on avalanche paths in the northern Rocky Mountains, Montana. Gt. Basin Nat. 1984, 44, 453–458. [Google Scholar]
- Oliver, C.D.; Adams, A.B.; Zasoski, R.J. Disturbance patterns and forest development in a recently deglaciated valley in northwestern Cascade range of Washington USA. Can. J. For. Res. 1984, 15, 221–232. [Google Scholar] [CrossRef]
- Butler, D.R.; Malanson, G.P. Non-equilibrium geomorphic processes and patterns on avalanche paths in the northern Rocky Mountains, U.S.A. Z. Geomorphol. 1990, 34, 257–270. [Google Scholar]
- Walsh, S.J.; Butler, D.R.; Allen, T.R.; Malanson, G.P. Influence of snow patterns and snow avalanches on the alpine treeline ecotone. J. Veg. Sci. 1994, 5, 657–672. [Google Scholar] [CrossRef]
- Veblen, T.T.; Kitzberger, T.; Donnegan, J. Climatic and human influences on fire regimes in ponderosa pine forests in the Colorado Front Range. Ecol. Appl. 2000, 10, 1178–1195. [Google Scholar] [CrossRef]
- Bebi, P.; Kulakowski, D.; Rixen, C. Snow avalanche disturbances in forest ecosystems-State of research and implications for management. For. Ecol Manag. 2009, 257, 1883–1892. [Google Scholar] [CrossRef]
- McGregor, M.D.; Furniss, M.M.; Oaks, R.D.; Gibson, K.E.; Meyer, H.E. MCH Pheromone for preventing Douglas-fir beetle infestation in windthrown trees. J. For. 1984, 82, 613–616. [Google Scholar]
- Blackford, D.C. Aspen Grove Trailhead Area Pleasant Grove RD, Uinta, NF; Report No. OFO-TR-05-15; U.S. Department of Agriculture, Forest Service Forest Health Protection: Ogden, UT, USA, 2005.
- Blackford, D.C.; Guyon, J. Recreation Sites Insect and Disease Activity Duchesne/Roosevelt RD, Ashley NF, Functional Assistance Trip Report; Report No. OFO-TR-06-05; U.S. Department of Agriculture, Forest Service, Forest Health Protection: Ogden, UT, USA, 2005.
- Hebertson, E.G. BLM Richfield Office-Henry Mountains Field Station Functional Assistance Visit; Report No. OFO-TR-06-03; U.S. Department of Agriculture, Forest Service, Forest Health Protection: Ogden, UT, USA, 2005.
- Raffa, K.F.; Berryman, A.A. Interacting selective pressure in conifer-bark beetle systems: A basis for reciprocal adaptations? Am. Nat. 1987, 129, 234–262. [Google Scholar] [CrossRef]
- Samman, S.; Logan, J. Assessment and Response to Bark Beetle Outbreaks in the Rocky Mountain Area; Report to Congress from Forest Health Protection, Washington Office, Forest Service, U.S. Department of Agriculture; General Technical Report RMRS-GTR-62; U.S. Department of Agricutlure Forest Service Rocky Mountain Research Station: Fort Collins, CO, USA, 2000; p. 46.
- Hardwood, W.G.; Rudinsky, J.A. The Flight and Olfactory Behavior of Checkered Beetles (Coleoptera: Cleridae) Predatory on the Douglas-Fir Beetle; Technical Bulletin 95; Agriculture Experiment Station, Oregon State University: Corvallis, OR, USA, 1966; p. 36. [Google Scholar]
- Jenkins, M.J.; Hebertson, E.; Page, W.G.; Jorgensen, C.A. Bark beetles, fuels, fires and implications for forest management in the Intermountain West. For. Ecol. Manag. 2008, 254, 16–34. [Google Scholar] [CrossRef]
- Anderson, M.N. Mechanisms of odor coding in coniferous bark beetles: From neuron to behavior and application. Psyche J. Entomol. 2012. [Google Scholar] [CrossRef]
- Furniss, M.M.; McGregor, M.D.; Foiles, M.W.; Patridge, M.D. Chronology and Characteristics of a Douglas-Fir Beetle [Dendroctonus pseudotsugae] Outbreak in Northern Idaho [Pseudotsuga menziesii]; General Technical Report, INT-59; U.S. Department of Agriculture, Forest Service, Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1979. Available online: http://agris.fao.org/agris-search/search.do?recordID=US7935406 (accessed on 1 March 2016).
- Negrón, J.F. Probability of infestation and extent of mortality associated with the Dogulas-fir beetle in Colorado Front Range. For. Ecol. Manag. 1998, 107, 71–85. [Google Scholar] [CrossRef]
- Rudinsky, J.A. Scolytid beetles associated with Douglas-fir: Response to terpenes. Science 1966, 152, 218–219. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.; Daterman, G. Using pheromone-baited traps to control the amount and distribution of tree mortality during outbreaks of the Douglas-fir beetle. For. Sci. 1997, 43, 65–70. [Google Scholar]
- Furniss, M.M. The Douglas-fir beetle in western forests a historical perspective—Part 1. Am. Entomol. 2014, 60, 84–96. [Google Scholar] [CrossRef]
- Lejeune, R.; McMullen, L.; Atkins, M. Influence of logging on Douglas fir beetle populations. For. Chron. 1961, 37, 308–314. [Google Scholar] [CrossRef] [Green Version]
- Furniss, M.M. Susceptibility of fire-injured Douglas-fir to bark beetle attack in southern Idaho. J. For. 1965, 63, 8–11. [Google Scholar]
- Lewisohn, E.; Gijzen, M.; Croteau, R. Defense mechanisms of conifers. Differences in constitutive and wound-induced monoterepene biosynthesis among species. Plant Physiol. 1991, 96, 44–49. [Google Scholar] [CrossRef]
- Franceschi, V.R.; Krokene, P.; Christiansen, E.; Krekling, T. Anatomical and chemical defenses of conifer bark against bark beetles and other pests. New Phytol. 2005, 167, 353–376. [Google Scholar] [CrossRef] [PubMed]
- Cates, R.G.; Alexander, H. Host resistance and susceptibility. In Bark Beetles in North American Conifers, A System for the Study of Evolutionary Biology, 1st ed.; Mitton, J.B., Sturgeon, K.B., Eds.; University of Texas Press: Austin, TX, USA, 1982; pp. 212–263. [Google Scholar]
- Goheen, D.J.; Hansen, E. Effects of pathogens and bark beetles on forests. In Beetle-Pathogen Interactions in Conifer Forests, 1st ed.; Schowalter, T.D., Filip, G.M., Eds.; Academic Press: London, UK, 1993; pp. 175–196. [Google Scholar]
- Mattson, W.; Haack, R. Role of drought in outbreaks of plant-eating insects. Bioscience 1987, 37, 110–118. [Google Scholar] [CrossRef]
- Cunningham, C.; Jenkins, M.; Roberts, D. Attack and brood production by the Douglas-fir beetle Dendroctonus pseudotsugae (Coleoptera: Scolytidae) in Douglas-fir, Pseudotsuga menziesii var. glauca (Pinaceae), following a wildfire. West. N. Am. Nat. 2005, 65, 70–79. [Google Scholar]
- McGregor, M.; Furniss, M.M.; Bousfield, W.E.; Almas, D.P.; Gravelle, P.J.; Oakes, R.D. Evaluation of the Douglas-Fir Beetle Infestation North Fork Clearwater River Drainage, Northern Idaho, 1970–73; Report No. 74-7; U.S. Department of Agriculture, Forest Service, Northern Region State and Private Forestry, Forest Pest Management: Missoula, MT, USA, 1974; p. 17.
- Fredericks, S.E.; Jenkins, M.J. Douglas-fir beetle {Dendroctonus pseudotsugae Hopkins, Coleoptera: Scolytidae} brood production on Douglas-fir defoliated by western spruce budworm (Choristoneura occidentalis Freeman, Lepidoptera: Tortricidae) in Logan Canyon, UT. Gt. Basin Nat. 1988, 48, 348–351. [Google Scholar]
- McMullen, L.H.; Atkins, M.D. On the flight and host selection of the Douglas-fir beetle, Dendroctonus pseudotsugae Hopk. (Coleoptera: Scolytidae). Can. Entomol. 1961, 94, 1309–1325. [Google Scholar] [CrossRef]
- Wright, L.C.; Berryman, A.A.; Wickman, B.E. Abundance of the fir engraver, Scolytus ventralis, and the Douglas-fir beetle, Dendroctonus pseudotsugae, following tree defoliation by the Douglas-fir tussock moth, Orgyia seudotsugata. Can. Entomol. 1984, 116, 293–305. [Google Scholar] [CrossRef]
- Edmonds, R.L.; Eglitis, A. The role of Douglas-fir and wood borers in the decomposition of and nutrient release from Douglas-fir logs. Can. J. For. Res. 1989, 19, 853–859. [Google Scholar] [CrossRef]
- Kegley, S. Douglas-Fir Beetle Management; Forest Health Protection and State Forestry Organizations. Chapter 4; U.S. Department of Agriculture, Forest Service. Availabel online: http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5187396.pdf (archived on 16 May 2015).
- Coulson, R.N. Population dynamics of bark beetles. Annu. Rev. Entomol. 1979, 24, 417–447. [Google Scholar] [CrossRef]
- Negrón, J.F.; Schaupp, W.C.; Gibson, K.E.; Anhold, J.; Hansen, D.; Their, R.; Mocettini, P. Estimating extent of mortality associated with the Douglas-fir beetle in the central and northern Rockies. West. J. Appl. For. 1999, 14, 121–127. [Google Scholar]
- Atkins, M.D. Behavioral variation among scolytids in relation to their habitat. Can. Entomol. 1966, 98, 285–288. [Google Scholar] [CrossRef]
- Lawrence, R. Early Detection of Douglas-fir beetle infestation with subcanopy resolution hyperspectral imagery. West. J. Appl. For. 2003, 18, 202–206. [Google Scholar]
- Weatherby, J.C.; Thier, R.W. A preliminary Validation of Douglas-Fir Beetle Hazard Rating System. Mountain Home Ranger District, Boise National Forest, 1992; Forest Pest Management Report No. R4-93-05; U.S. Department of Agriculture, Forest Service: Boise, ID, USA, 1993; p. 7.
- Kulman, H.M. Effects of insect defoliation on growth and mortality of trees. Annu. Rev. Entomol. 1971, 16, 289–324. [Google Scholar] [CrossRef]
- Brookes, M.H.; Colbert, J.J.; Mitchell, R.G.; Stark, R.W. Western Spruce Budworm; Technical Bulletin 1694; U.S. Department of Agriculture, Forest Service, Cooperative State Research Service: Washington, DC, USA, 1987; p. 108.
- Fellin, D.G.; Dewey, J.E. Western Spruce Budworm; Forest Insect and Disease Leaflet 53; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1982; p. 10.
- Swetnam, T.W.; Lynch, A.M. Multicentury, regional scale patterns of western spruce budworm outbreaks. Ecol. Monogr. 1993, 63, 399–424. [Google Scholar] [CrossRef]
- Myers, J.H. Synchrony in outbreaks of forest Lepidoptera: A possible example of the moran effect. Ecology 1998, 79, 1111–1117. [Google Scholar] [CrossRef]
- Ryerson, D.E.; Swetnam, T.W.; Lynch, A.M. A tree-ring reconstruction of western spruce budworm outbreaks in the San Juan Mountains, Colorado, U.S.A. Can. J. For. Res. 2003, 33, 1010–1028. [Google Scholar] [CrossRef]
- Murdock, T.Q.; Taylor, S.W.; Flower, A.; Mehlenbacher, A.; Montenegro, A.; Zwiers, F.W.; Alfaro, R.; Spittlehouse, D.L. Pest outbreak distribution and forest management impacts in a changing climate in British Columbia. Environ. Sci. Policy 2013, 6, 75–89. [Google Scholar] [CrossRef]
- Cates, R.G.; Redak, R.; Henderson, C.B. Natural product defensive chemistry of Douglas-fir, western spruce budworm success and forest management practices. J. Appl. Entomol. 1983, 96, 173–182. [Google Scholar]
- Wulf, N.; Cates, R. Site and stand characteristics. In Western Spruce Budworm; Technical Bulletin 1694; Brookes, M.H., Campbell, R.W., Colbert, J.J., Mitchell, R.G., Stark, R.W., Eds.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1987; pp. 90–115. [Google Scholar]
- Carlson, C.E.; Fellin, D.G.; Schmidt, W.C.; O’Laughlin, J.; Pfister, R.D. The western spruce budworm in northern Rocky Mountain forests: A review of ecology, insecticidal treatments and silvicultural practices. In Proceedings of the Symposium Management of Second-Growth Forests: The State of Knowledge and Research Needs, Missoula, MT, USA, 14 May 1982; O’Laughlin, J., Pfister, R.D., Eds.; pp. 76–103.
- Hadley, K.; Veblen, T. Stand response to western spruce budworm and Douglas-fir bark beetle outbreaks, Colorado Front Range. Can. J. For. Res. 1993, 23, 479–491. [Google Scholar] [CrossRef]
- Dewey, J.E. Damage to Douglas-fir cones by Choristoneura occidentalis. J. Econ. Entomol. 1970, 63, 1804–1806. [Google Scholar] [CrossRef]
- Frank, C.J.; Jenkins, M.J. Impact of the western spruce budworm on buds, developing cones and seeds of Douglas-fir in west-central Idaho. Environ. Entomol. 1987, 16, 304–308. [Google Scholar] [CrossRef]
- Owens, J.N. The Reproductive Cycle of Douglas-Fir; Research Paper BC-P-8; Canadian Forest Service Pacific Forest Research Center: Victoria, BC, Canada, 1976; p. 23. [Google Scholar]
- Lowry, W. Apparent meteorological requirements for abundant cone crops in Douglas-fir. For. Sci. 1966, 12, 185–192. [Google Scholar]
- Fellin, D.G.; Shearer, R.C.; Carlson, C.E. Western spruce budworm in the northern Rocky Mountains. West. Wildlands Nat. Res. J. 1983, 9, 2–7. [Google Scholar]
- Swetnam, T.W.; Thompson, M.A.; Sutherland, E.K. Using Dendrochronology to Measure Radial Growth of Defoliated Trees; Handbook No. 639; U.S. Department of Agriculture, Forest Service, Cooperative State Research Service: Washington, DC, USA, 1988; p. 39.
- Swetnam, T.W.; Lynch, A.M. A tree-ring reconstruction of western spruce budworm history in the southern Rocky Mountains. For. Sci. 1989, 35, 962–986. [Google Scholar]
- Alfaro, R.I.; Van Sickle, G.A.; Thomson, A.J.; Wegwitz, E. Tree mortality and radial growth losses caused by the western spruce budworm in a Douglas-fir stand in British Columbia. Can. J. For. Res. 1982, 12, 780–787. [Google Scholar] [CrossRef]
- Hawksworth, F.G.; Wiens, D. Dwarf Mistletoes: Biology, Pathology, and Systematics; Agricultural Handbook 709; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1996; p. 410.
- Watson, D.M. Mistletoe—A keystone resource in forests and woodlands worldwide. Annu. Rev. Ecol. Syst. 2001, 32, 219–249. [Google Scholar] [CrossRef]
- Hoffman, J.T. Management Guide for Dwarf Mistletoe Arceuthobium spp.; U.S. Department of Agriculture, Forest Service, 2010. Available online: http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5187427.pdf (accessed on 28 May 2015).
- Smith, R.H. Xylem Resin in the Resistance of the Pinaceae to Bark Beetles; General Technical Report PSW-1; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkeley, CA, USA, 1972; p. 7.
- Pate, J.S. Mineral relationships of parasites and their hosts. In Parasitic Plants, 1st ed.; Press, M.C., Graves, J., Eds.; Chapman & Hall: London, UK, 1995; pp. 80–102. [Google Scholar]
- Shaw, D.C.; Watson, D.M.; Mathiasen, R.L. Comparison of dwarf mistletoes (Arceuthobium spp., Viscaceae) in the western United States with mistletoes (Amyema spp., Loranhaceae) in Australia-ecological analogs and reciprocal models for ecosystem management. Aust. J. Bot. 2004, 52, 481–498. [Google Scholar] [CrossRef]
- Lamont, B. Mineral nutrition of mistletoes. In Biology of Mistletoes, 1st ed.; Calder, D.M., Bernhardt, P., Eds.; The Academic Press: Sydney, Australia, 1983; p. 348. [Google Scholar]
- Tinnin, R.O.; Hawksworth, F.G.; Knutson, D.M. Witches’ broom formation in conifers infected by Arceuthobium spp.: An example of parasitic impact upon community dynamics. Am. Midl. Nat. 1982, 107, 351–359. [Google Scholar] [CrossRef]
- Kuijt, J. The Biology of Parasitic Flowering Plants, 1st ed.; University of California Press: Berkeley, CA, USA, 1969; p. 246. [Google Scholar]
- Hawksworth, F.G.; Wiens, D. Biology and Classification of Dwarf Mistletoes (Arceuthobium); Agriculture Handbook Book Number 401; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1972; p. 234. Available online: http://naldc.nal.usda.gov/naldc/download.xhtml?id=CAT87208731 (accessed on 23 February 2016).
- Geils, B.W.; Mathiasen, R.L. Intensification of dwarf mistletoe on Southwestern Douglas-fir. For. Sci. 1990, 36, 955–969. [Google Scholar]
- Sala, A.; Carey, E.V.; Callawy, R.M. Dwarf mistletoe affects whole-tree water relations of Douglas-fir and western larch primarily through changes in leaf to sapwood ratios. Oecologia 2001, 126, 42–52. [Google Scholar] [CrossRef]
- Hadfield, J.S.; Mathiasen, R.L.; Hawksworth, F.G. Douglas-Fir Dwarf Mistletoe; Forest Insect and Disease Leaflet 54; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2000; p. 9.
- Castello, J.D.; Leopold, D.J.; Smallidge, P.J. Pathogens, patterns, and processes in forest ecosystems. BioScience 1995, 45, 16–24. [Google Scholar] [CrossRef]
- Ayres, M.P.; Lombardero, M.J. Assessing the consequences of global change for forest disturbance from herbivores and pathogens. Sci. Total Environ. 2000, 262, 263–286. [Google Scholar] [CrossRef]
- Williams, R.E.; Shaw, C.G., III; Wargo, P.M.; Sites, W.H. Armillaria Root Disease; Forest Insect and Disease Leaflet 78; U.S. Department of Agriculture, Forest Service, Northern Area State and Private Forestry, 1986. Available online: http://www.na.fs.fed.us/spfo/pubs/fidls/armillaria/armillaria.htm?) (accessed on 23 February 2016)Forest Insect and Disease Leaflet 78.
- Wargo, P.M.; Shaw, C.G., III. Armillaria root rot: The puzzle is being solved. Plant Dis. 1985, 69, 826–832. [Google Scholar] [CrossRef]
- McDonald, G.I.; Martin, N.E.; Harvey, A.E. Armillaria in the Northern Rockies: Pathogenicity and Host Susceptibility on Pristine and Disturbed Sites; Research Note INT-371; U.S. Department of Agriculture, Forest Service, Intermountain Research Station: Ogden, UT, USA, 1987. Available online: https://archive.org/details/armillariainnort371mcdo (accessed on 23 February 2016).
- Shaw, C.G., III; Kile, G.A. (Eds.) Armillaria Root Disease; Agricultural Handbook No. 691; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1991.
- Wickman, B.E. Tree Mortality and Top-Kill Related to Defoliation by the Douglass-Fir Tussock Moth in the Blue Ridge Mountains Outbreak; Forest Research Paper PNW-223; U.S. Department of Agriculture, Forest Service Pacific Northwest Research Station: Portland, OR, USA, 1978; p. 39.
- Johnson, D.W. Forest Pest Training Manual; U.S. Department of Agriculture, Forest Service, Forest Pest Management, Rocky Mountain Region: Lakewood, CO, USA, 1985.
- Haggle, S.K. Managmement Guide for Armaliaria Root Disease; U.S. Department of Agriculture, Forest Service, Forest Health Protection and State Forestry Organizations, 2008. Available online: http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5187208.pdf (accessed on 24 February 2016).
- Guillaumin, J.J.; Legrand, P. Armillaria root rots. In Infectious Forest Diseases, 1st ed.; Gonthier, P., Nicolotti, G., Eds.; CABI: Boston, MA, USA, 2013; pp. 159–178. [Google Scholar]
- Worral, J.J.; Sullivan, K.F.; Harrington, T.C.; Steimel, J.P. Incidence, host relations and population structure of Armillaria ostoyae in Colorado campgrounds. For. Ecol. Manag. 2004, 192, 191–206. [Google Scholar] [CrossRef]
- Amman, G.D. Bark beetle-fire associations in the Greater Yellowstone area. In Proceedings of the Fire and the Environment Symposium, Knoxville, TN, USA, 20–24 March 1991; Nordvin, S.C., Waldrop, T.A., Eds.; pp. 313–320.
- Amman, G.D.; Ryan, K.C. Insect Infestation of Fire-Injured Trees in the Greater Yellowstone Area; Research Note INT-398; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1991; p. 9.
- Ryan, K.C.; Amman, G.D. Interactions between fire-injured trees and insects in the greater Yellowstone area. In Plants and Their Environment, Proceedings of the First Biennial Scientific Conference on the Greater Yellowstone Ecosystem, Yellowstone National Park, WY, USA, 16–17 September 1991; Despain, D.G., Ed.; National Park Service: Denver, CO, USA, 1995; pp. 259–271. [Google Scholar]
- Rasmussen, L.; Amman, G.; Vandygriff, J.; Oakes, R.; Munson, S.; Gibson, K. Bark Beetle and Wood Borer Infestation in the Greater Yellowstone Area during Four Postfire Years; Research Paper INT-RP-487; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1996; p. 9.
- Dickinson, M.B.; Johnson, E.A. Fire effets on trees. In Forest Fires Behavior and Ecological Effects, 1st ed.; Johnson, E.A., Miyanishi, K., Eds.; Academic Press: New York, NY, USA, 2001; pp. 477–521. [Google Scholar]
- Stevens-Rumman, C.P.; Morgan, C.P.; Hoffman, C. Bark beetles and wildfires: How does forest recovery change with repeated disturbances in mixed conifer forests? Ecosphere 2015, 6, 1–17. [Google Scholar] [CrossRef]
- Furniss, M.M.; Livingston, R.L.; McGregor, M.D. Development of a stand susceptibility classification for Douglas-fir beetle. In Hazard-Rating Systems in Forest Insect Pest Management: Symposium Proceedings, Athens, GA, USA, 31 July–1 August 1980; Hedden, R.L., Barras, S., Coster, J.E., Eds.; U.S. Department of Agriculture, Forest Service: Washington, D.C., USA, 1981; pp. 115–128. [Google Scholar]
- Christiansen, E.; Waring, R.H.; Berryman, A.A. Resistance of conifers to bark beetle attack: Searching for general relationships. For. Ecol. Manag. 1987, 22, 89–106. [Google Scholar] [CrossRef]
- Klutsch, J.G.; Negrón, J.F.; Costello, S.L.; Rhoades, C.C.; West, D.R.; Popp, J.; Caissie, R. Stand characteristics and downed woody debris accumulations associated with a mountain pine beetle (Dendroctonus ponderosae Hopkins) outbreak in Colorado. For. Ecol. Mang. 2009, 258, 641–649. [Google Scholar] [CrossRef]
- Collins, B.J.; Rhoades, C.C.; Battaglia, M.A.; Hubbard, R.M. The effects of bark beetle outbreaks on forest development, fuel loads and potential fire behavior in salvage logged and untreated lodgepole pine forests. For. Ecol. Manag. 2012, 284, 260–268. [Google Scholar] [CrossRef]
- DeRose, R.J.; Long, J.N. Wildfire and spruce beetle outbreak: Simulation of interacting disturbances in the central Rocky Mountains. Ecoscience 2009, 16, 28–38. [Google Scholar] [CrossRef]
- Parker, T.J.; Clancy, K.M.; Mathiasen, R.L. Interactions among fire, insects and pathogens in coniferous forests of the interior western United States and Canada. Agric. For. Entomol. 2006, 8, 167–189. [Google Scholar] [CrossRef]
- Giunta, A. Douglas-Fir beetle mediated changes to fuel complexes, foliar moisture content, and terpenes in interior Douglas-fir forests of the central Rocky Mountains. Master’s Thesis, Utah State University, Logan, UT, USA, 2016. [Google Scholar]
- McMillin, J.D.; Allen., K.K. Impacts of Douglas-Fir Beetles on Overstory and Understory Conditions of Douglas-Fir Stands; General Technical Report R2-64; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Golden, CO, USA, 2000; p. 17.
- Von Arx, J.A. Ueber die Ascomycetengattungen Ceratostomella Sacc., Ophiostoma Syd. Und Rostrella Zimmerman. Antonie Leeuwenhoek 1952, 18, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Gruell, G.E. Fire and Vegetative Trends in Northern Rockies: Interpretations from 1871–1982 Photographs; General Technical Report INT-130; U.S. Department of Agriculture, Forest Service Intermountain Research Station: Missoula, MT, USA, 1983; p. 117.
- Gavin, D.; Flower, A.; Cohn, G.; Heyerdahl, E.; Parsons, R. Interactions of Insects, Fire and Climate on Fuel Loads and Fire Behavior in Mixed Conifer Forest; Final Report for JFSP Project 09-1-06-5; 2013. Available online: http://www.firescience.gov/projects/09-1-06-5/project/09-1-06-5_final_report.pdf (accessed on 27 May 2015).
- Black, S.H.; Kulakowski, D.; Noon, B.R.; DellaSalla, D.A. Do bark beetle outbreaks increase wildfire risks in the central U.S. Rocky Mountains? Implications from recent research. Nat. Areas J. 2013, 33, 59–65. [Google Scholar] [CrossRef]
- Flower, A.; Gavin, D.G.; Heyerdahl, E.K.; Parson, R.A.; Cohn, G.M. Western spruce budworm outbreaks did not increase fire risk over the last three centuries: A dendrochronological analysis of inter-disturbance synergism. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed]
- Hummel, S.; Agee, J.K. Western spruce budworm defoliation effects on forest structure and potential fire behavior. Northwest Sci. 2003, 77, 159–169. [Google Scholar]
- Burgan, R.E.; Rothermel, R.C. BEHAVE: Fire Behavior Prediction and Fuel Modeling System FUEL Subsystem; General Technical Report INT-167; U.S. Department of Agriculture, Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1984; p. 126.
- Koonce, A.L.; Roth, L.F. The effects of dwarf mistletoe on fuel in precommercial ponderosa pine stands. In Proceedings of the Eighth Conference on Fire and Forest Meteorology: Weather—The Drive Train Connecting the Solar Engine to Forest Ecosystems, Detroit, MI, USA, 29 April–2 May 1985; Donoghue, L.R., Martin, R.E., Eds.; Society of American Foresters: Bethesda, MD, USA, 1985; pp. 66–72. [Google Scholar]
- Dickman, A.; Cook, S. Fire and fungus in a mountain hemlock forest. Can. J. Bot. 1989, 67, 2005–2016. [Google Scholar] [CrossRef]
- Jain, T.B.; Battaglia, M.A.; Han, H.S.; Graham, R.T.; Keyes, C.R.; Fried, J.S.; Sandquist, J.E. A Comprehensive Guide to Fuel Management Practices for Dry Mixed Conifer Forests in the Northwestern United States; General Technical Report RMRS-GTR-292; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Golden, CO, USA, 2012; p. 331.
- Hagle, S.K.; Gibson, K.E.; Tunnock, S. Field Guide to Diseases and Insect Pests of Northern and Central Rocky Mountain Conifers; General Technical Report R1-03-08; U.S. Department of Agriculture, Forest Service, State and Private Forestry, Northern and Intermountain Regions: Missoula, MT, USA; Ogden, UT, USA, 2003; p. 197.
- Man, G. Major Forest and Insect Disease Conditions in the United States: 2011 Update; Forest Service Report FS-1000; U.S. Department of Agriculture: Washington, DC, USA, 2012; pp. 14–16.
- Jenkins, M.L. Major Forest and Insect Disease Conditions in the United States: 2013; Forest Service Report FS-1054; U.S. Department of Agriculture, Forest Service, Forest Health Protection: Washington, DC, USA, 2015; pp. 18–20.
- Lessard, E.D.; Schmid, J.M. Emergence, attack densities, and host relationships for the Douglas-fir beetle, (Dendroctonus pseudotsugae Hopkins) in northern Colorado. Gt. Basin Nat. 1990, 50, 333–338. [Google Scholar]
- Schmid, J.M.; Mata, S.A. Natural Variability of Specific Forest Insect Populations and Their Associated Effects in Colorado; General Technical Report RM-GTR-275; U.S Department of Agriculture, Forest Service: Washington, DC, USA, 1996.
- Berryman, A.A.; Wright, L.C. Defoliation, tree condition, and bark beetles. In The Douglas-Fir Tussock Moth a Synthesis; Stark, M.H., Campell, R.W., Eds.; Technical Bulletin 1585; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1978; pp. 81–87. [Google Scholar]
- Singh, P. Research and management strategies for major tree diseases in Canada: Synthesis part 1. For. Chron. 1993, 69, 151–162. [Google Scholar] [CrossRef]
- Huber, D.P.W.; Borden, J.H. Angiosperm bark volatiles disrupt response of Douglas-fir beetle, Dendroctonus pseudotsugae, to attractant-baited traps. J. Chem. Ecol. 2001, 27, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Kay, C.E. Aboriginal overkill and native burning: Implications for modem ecosystem management. West. J. Appl. For. 1995, 10, 121–126. [Google Scholar]
- Keane, R.E.; Ryan, K.C.; Veblen, T.T.; Allen, C.D.; Logan, J.A.; Hawkes, B. The cascading effects of fire exclusion in Rocky Mountain ecosystems. In Rocky Mountain Futures: An ecological Perspective, 1st ed.; Baron, J.S., Ed.; Island Press: Washington, DC, USA, 2002; pp. 133–152. [Google Scholar]
- Baker, W.L. Indians and fire in the Rocky Mountains: The wilderness hypothesis reviewed. In Fire Native Peoples, and the Natural Landscape, 1st ed.; Vale, T.R., Ed.; Island Press: Washington, DC, USA, 2002; pp. 41–77. [Google Scholar]
- Peterson, C.S.; Speth, L.E. A History of the Wasatch-Cache National Forest. Utah State University, 1980. Available online: http://www.fs.usda.gov/Internet/FSE_DOCUMENTS/stelprdb5053310.pdf (accessed on 1 March 2016). [Google Scholar]
- Alexander, T.G.; Fish, R.J. The Forest Service in Utah. Available online: http://www.uen.org/utah_history_encyclopedia/f/FOREST_SERVICE.html (accessed on 1 March 2016).
- Owston, P.W.; Stein, W.I. Pseudotsuga Carr., Douglas-fir. In Seeds of Woody Plants in the United States; Agriculture Handbook 450; Schopmeyer, C.S., Ed.; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 1974; pp. 674–683. [Google Scholar]
- Noss, R.F.; Franklin, J.F.; Baker, W.L.; Schoennagel, T.; Moyle, P.B. Managing fire-prone forests in the western United States. Front. Ecol. Environ. 2006, 4, 481–487. [Google Scholar] [CrossRef]
- Landres, P.; Morgan, P.; Swanson, F. Overview of the use of natural variability concepts in managing ecological systems. Ecol. Appl. 1999, 9, 1179–1188. [Google Scholar]
- Graham, R.T.; Harvey, A.E.; Jain, T.B.; Tonn, J.R. Effects of Thinning and Similar Stand Treatments on Fire Behavior in Western Forests; General Technical Report PNW-GTR-463; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1999.
- Rippy, R.C.; Stewart, J.E.; Zambino, P.J.; Klopfenstein, N.B.; Tirocke, J.M.; Kim, M.S.; Thies, W.G. Root Diseases in Coniferous Forests of the Inland West: Potential Implications of Fuels Treatments; General Technical Report, RMRS-GTR-141; U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005.
- Edmonds, R.L.; Agee, J.K.; Gara, R.I. Forest Health and Protection, 1st ed.; Waveland Press, Inc.: Long Grove, IL, USA, 2000; pp. 359–397. [Google Scholar]
- Fernandes, P.M.; Hermínio, S.B. A review of prescribed burning effectiveness in fire hazard reduction. Int. J. Wildland Fire 2003, 12, 117–128. [Google Scholar] [CrossRef]
- Hessburg, P.F.; Mitchell, R.G.; Filip, G.M. Historical and Current Roles of Insects and Pathogens in Eastern Oregon and Washington Forested Landscapes; General Technical Report, PNW-GTR-327; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 1994.
- Arno, S.F. Forest fire history in the northern Rockies. J. For. 1980, 78, 460–465. [Google Scholar]
- McIver, J.D.; Starr, L. A literature review on the environmental effects of postfire logging. West. J. Appl. For. 2001, 16, 159–168. [Google Scholar]
- Potts, D.F.; Peterson, D.L.; Zuuring, H.R. Watershed Modeling for Fire Management Planning in the Northern Rocky Mountains; Research Paper PSW-177; U.S. Department of Agriculture, Forest Service, Pacific Southwest Forest and Range Experiment Station: Berkely, CA, USA, 1985.
- Donato, D.C.; Fontaine, J.B.; Campbell, J.L.; Robinson, W.D.; Kauffman, J.B.; Law, B.E. Post-wildfire logging hinders regeneration and increases fire risk. Science 2006. [Google Scholar] [CrossRef] [PubMed]
- Seidl, R.; Spies, T.A.; Peterson, D.L.; Stephens, S.L.; Hicke, J.A. Searching for resilience: Addressing the impacts of changing disturbance regimes on forest ecosystem services. J. Appl. Ecol. 2016, 53, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.T.; McCaffrey, S.; Jain, T.B. Science Basis for Changing Forest Structure to Modify Wildfire Behavior and Severity; General Technical Report RMRS-120; U.S. Department of Agriculture, Forest Service: Fort Collins, CO, USA, 2004.
- Stephens, S.L.; Moghaddas, J.J. Experimental fuel treatment impacts on forest structure, potential fire behavior, and predicted tree mortality in a California mixed conifer forest. For. Ecol. Manag. 2005, 215, 21–36. [Google Scholar] [CrossRef]
- Allen, C.D.; Betancourt, J.L.; Swetnam, T.W. Landscape changes in the southwestern United States: Techniques, long-term data sets, and trends. In Perspectives on the Land-Use History of North America: A Context for Understanding Our Changing Environment; Sisk, T.D., Ed.; U.S. Geological Survey: Reston, VA, USA; pp. 71–84.
- Caprio, A.C.; Graber, D.M. Returning fire to the mountains: Can we successfully restore the ecological role of pre-Euroamerican fire regimes to the Sierra Nevada. In Proceedings of the Wilderness Science in a Time of Change Conference, Missoula, MT, USA, 23–27 May 1999; Cole, D.N., McCool, S.F., Borrie, W.T., O’Laughlin, J., Eds.; U.S. Department of Agriculture, Forest Service: Proceedings RMRS-P-15-VOL-5. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station: Ogden, UT, USA, 2000; pp. 223–241. [Google Scholar]
- Heyerdahl, E.K.; Lertzman, K.; Wong, C.M. Mixed-severity fire regimes in dry forests of southern interior British Columbia, Canada. Can. J. For. Res. 2012, 42, 88–98. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).