
Effect of Tree Spacing on Tree Level Volume Growth, Morphology, and Wood Properties in a 25-Year-Old Pinus banksiana Plantation in the Boreal Forest of Quebec

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Growth Measurements
2.4. Wood Density
2.5. MOE and MOR Tests
2.6. Statistical Analyses
3. Results
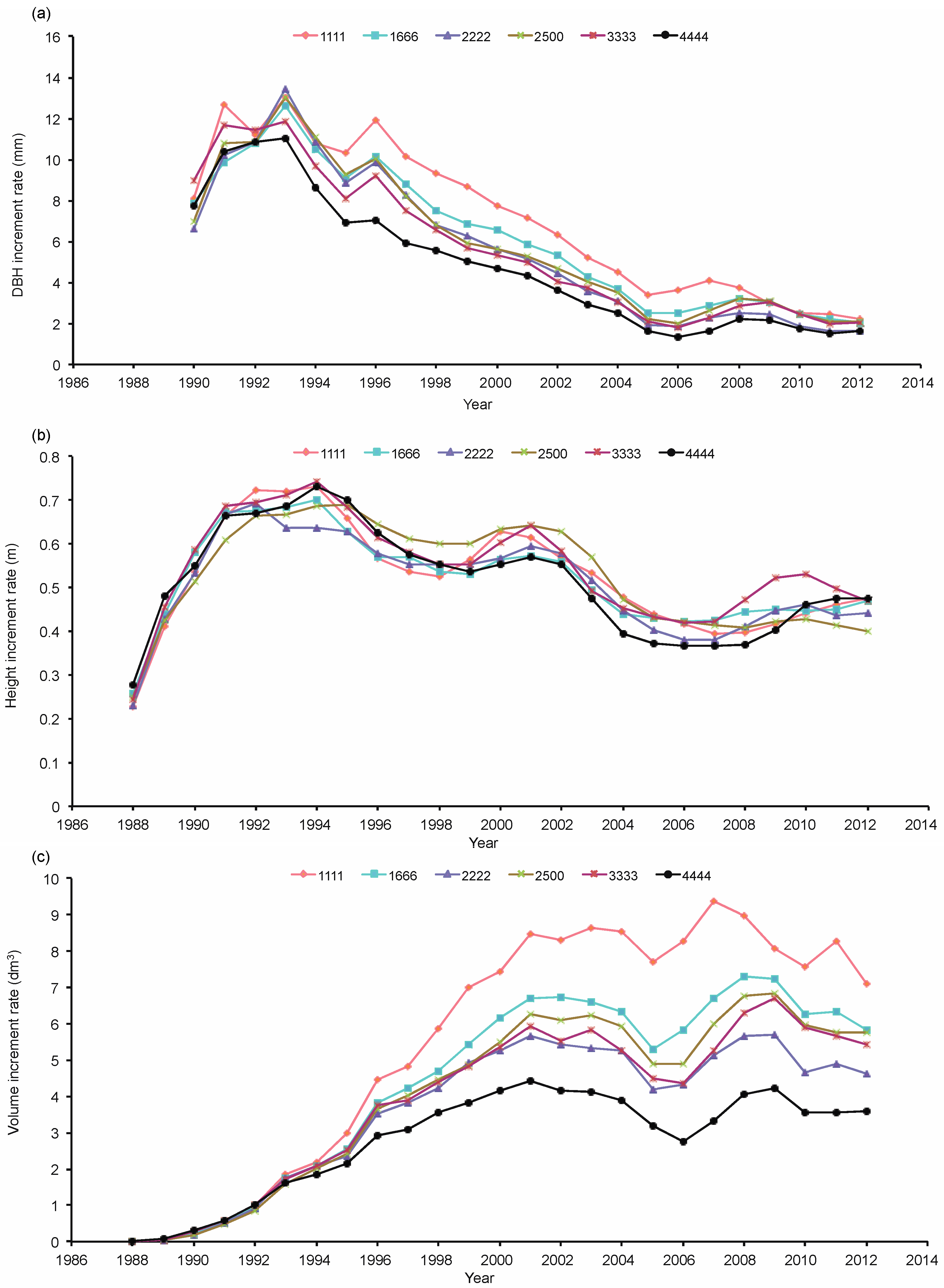
3.1. Growth Rate
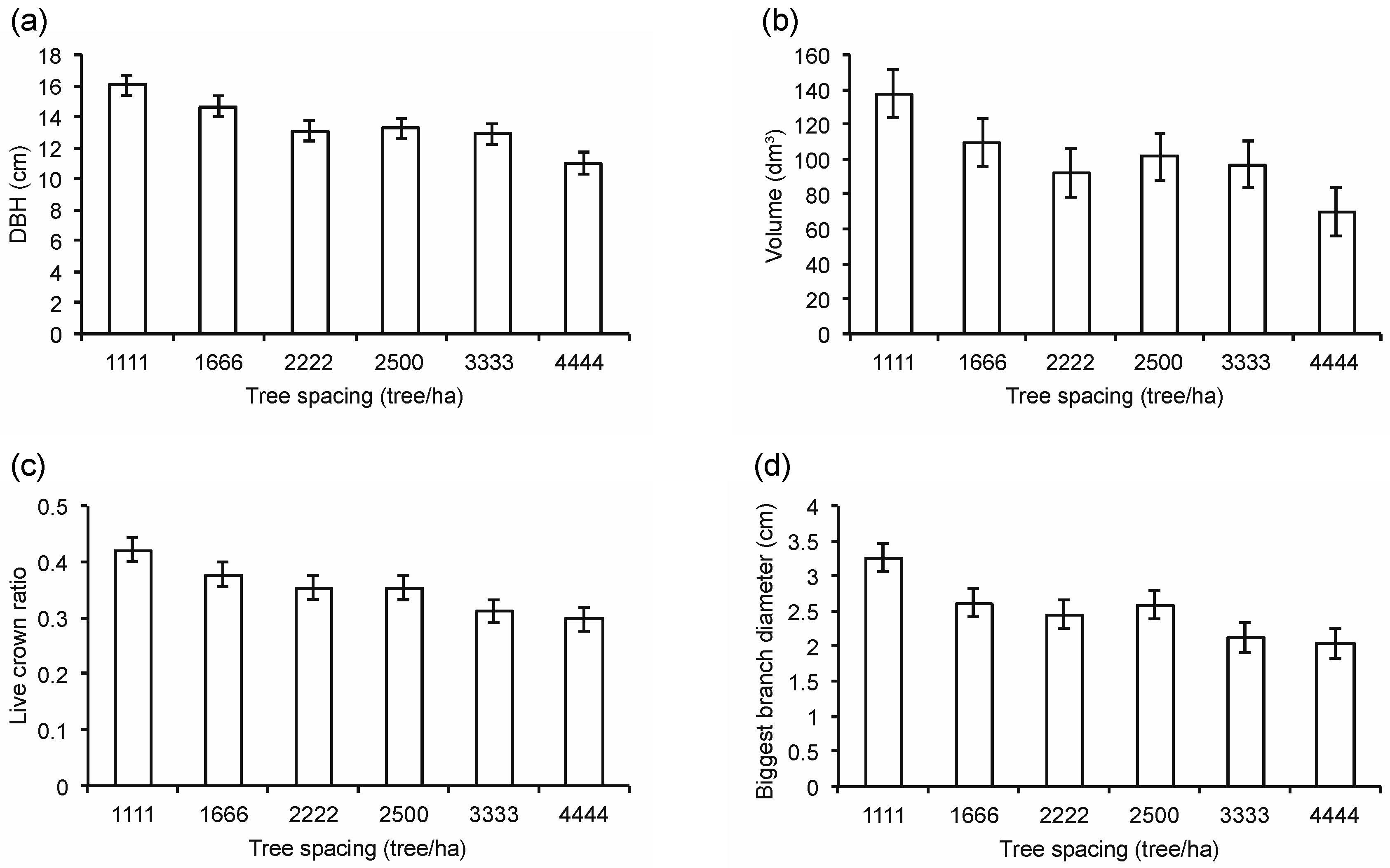
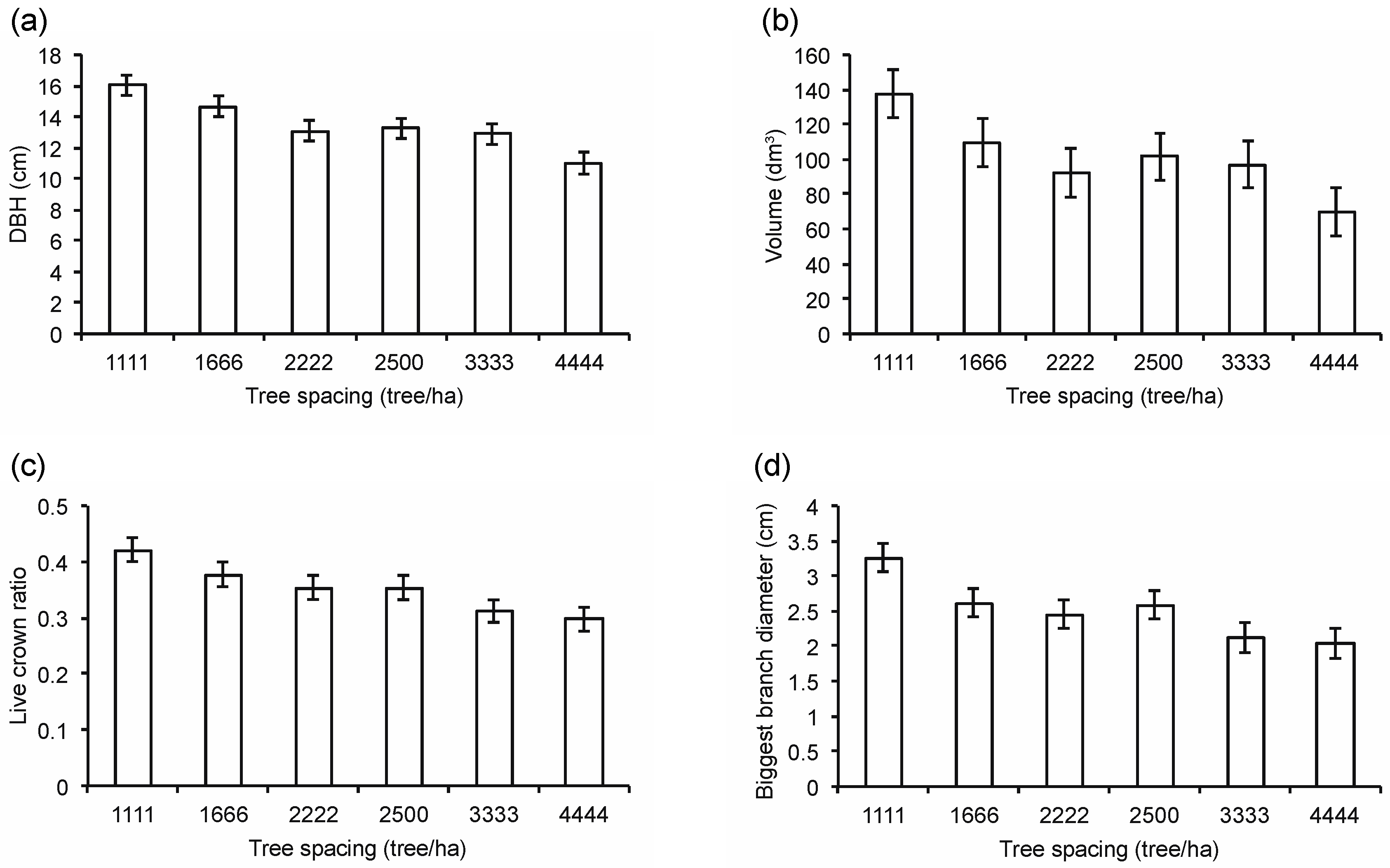
3.2. Tree Morphology
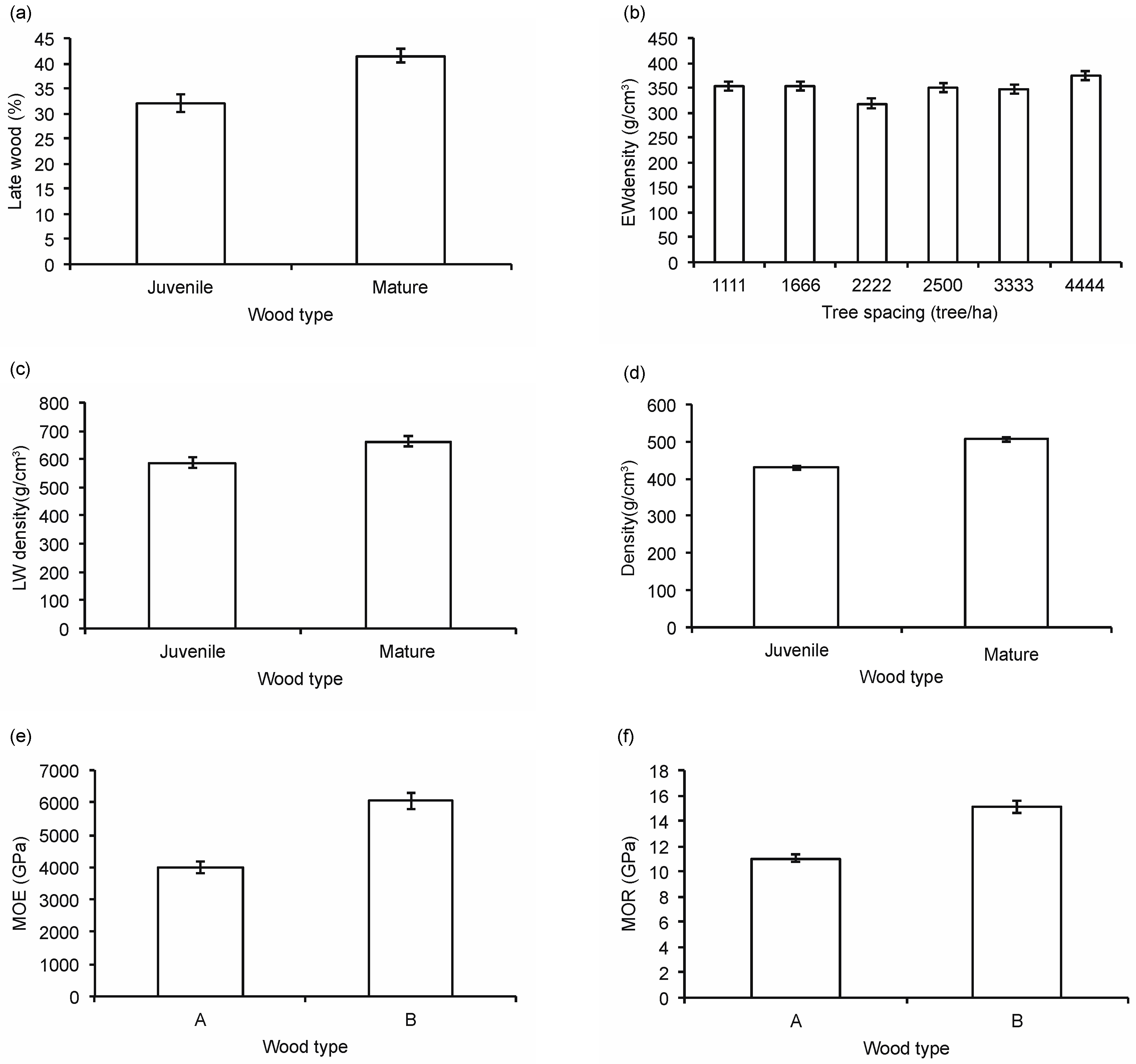
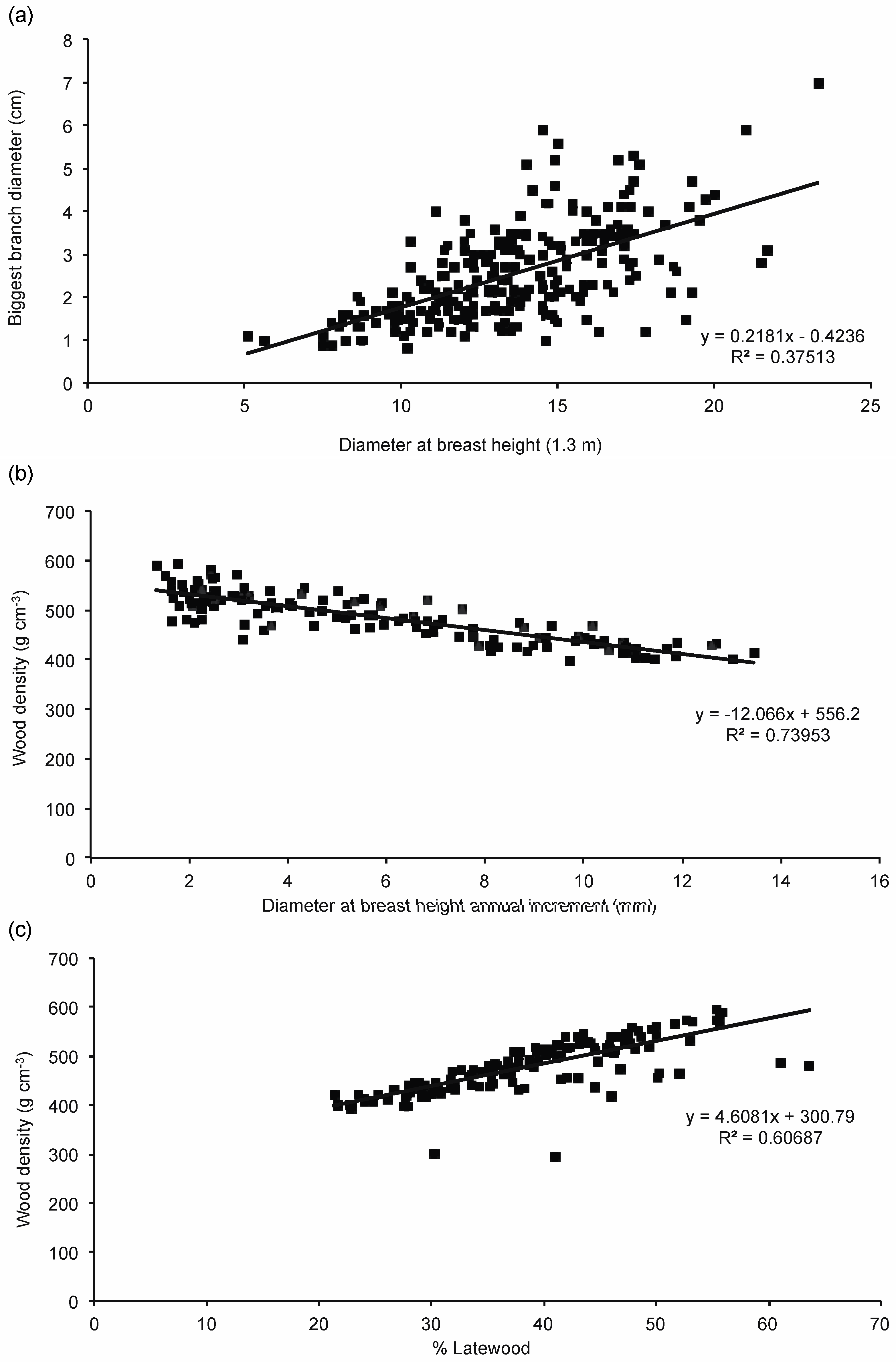
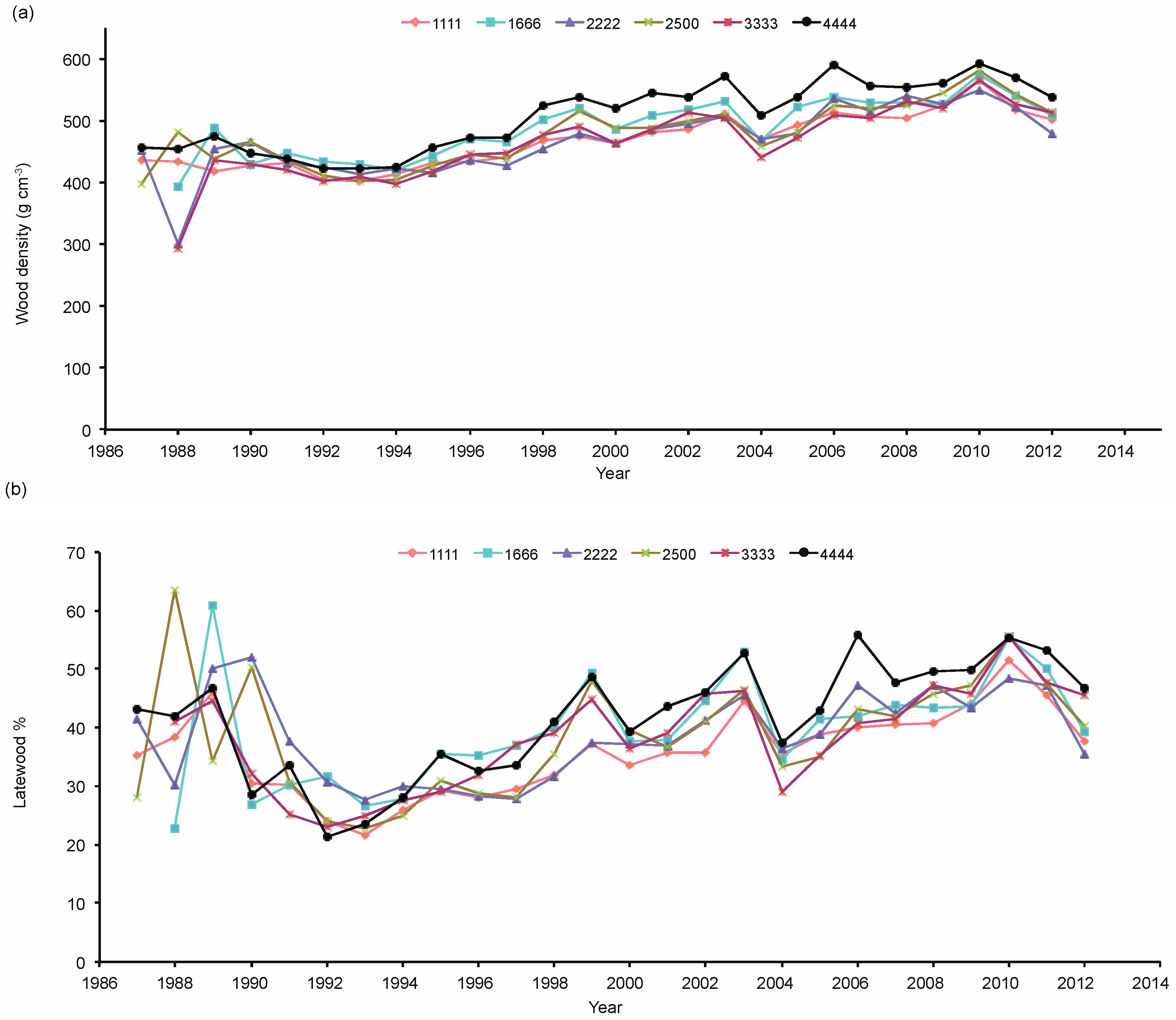
3.3. Wood Properties
4. Discussion
4.1. Spacing and Tree Growth
4.2. Spacing and Wood Quality Attributes
4.3. Wood Quality and Juvenile—Mature Wood
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thiffault, N.; Roy, V.; Prégent, G.; Cyr, G.; Jobidon, R. La sylviculture des plantations résineuses au Québec. Nat. Can. 2003, 127, 63–80. [Google Scholar]
- Baldwin, V.C.J.; Peterson, K.D.; Clark, A., III; Ferguson, R.B.; Strub, M.R.; Bower, D.R. The effects of spacing and thinning on stand and tree characteristics of 38-year-old loblolly pine. For. Ecol. Manag. 2000, 137, 91–102. [Google Scholar] [CrossRef]
- Akers, M.K.; Kane, M.; Zhao, D.; Teskey, R.O.; Daniels, R.F. Effects of planting density and cultural intensity on stand and crown attributes of mid-rotation loblolly pine plantations. For. Ecol. Manag. 2013, 310, 468–475. [Google Scholar] [CrossRef]
- Cardoso, D.J.; Lacerda, A.E.B.; Rosot, M.A.D.; Garrastazú, M.C.; Lima, R.T. Influence of spacing regimes on the development of loblolly pine (Pinus taeda) in southern Brazil. For. Ecol. Manag. 2013, 310, 761–769. [Google Scholar] [CrossRef]
- Barron-Gafford, G.A.; Will, R.E.; Burkes, E.C.; Shiver, B.; Teskey, R.O. Nutrient concentrations and contents, and their relation to stem growth, of intensively managed Pinus teada and Pinus elliottii stands of different planting densities. For. Sci. 2003, 49, 291–300. [Google Scholar]
- Zhu, J.; Scott, C.T.; Scallon, K.L.; Myers, G.C. Effects of plantation density on wood density and anatomical properties of red pine (Pinus resinosa ait.). Wood Fiber Sci. 2007, 39, 502–512. [Google Scholar]
- Will, R.; Hennessey, T.; Lynch, T.; Holeman, R.; Heinemann, R. Effects of planting density and seed source on loblolly pine stands in southeastern Oklahoma. For. Sci. 2010, 56, 437–443. [Google Scholar]
- Cao, Q.V.; Burkhart, H.E.; Max, T.A. Evaluation of two methods for cubic-volume prediction of loblolly pine to any merchantable limit. For. Sci. 1980, 26, 71–80. [Google Scholar]
- Auty, D.; Achim, A.; Bédard, P.; Pothier, D. Statsaw: Modelling lumber product assortment using zero-inflated poisson regression. Can. J. For. Res. 2014, 44, 638–647. [Google Scholar] [CrossRef]
- Zhang, S.; Chauret, G.; Ren, H. Impact of initial spacing on plantation black spruce lumber grade yield, bending properties, and MSR yield. Wood Fiber Sci. 2002, 34, 460–475. [Google Scholar]
- Gardiner, B.; Moore, J. Creating the wood supply of the future. In Challenges and Opportunities for the World’s Forests in the 21st Century; Fenning, T., Ed.; Springer: Dordrecht, The Netherlands, 2013; Volume 81 of the Series Forest Science, pp. 677–704. [Google Scholar]
- Carino, H.F.; Biblis, E.J. Comparative analysis of the quality of sawlogs from 35-, 40-, and 50-year-old loblolly pine plantation stands. For. Prod. J. 2000, 50, 48–52. [Google Scholar]
- Carino, H.F.; Biblis, E.J. Impact of stand density on the quality and value of 35-year-old loblolly pine plantation sawtimber: A case study. For. Prod. J. 2009, 59, 62–66. [Google Scholar]
- Zhang, S.Y. Effect of age on the variation, correlations and inheritance of selected wood characteristics in black spruce (Picea mariana). Wood Sci. Technol. 1998, 32, 197–204. [Google Scholar]
- Wang, L.; Payette, S.; Begin, Y. Relationships between anatomical and densitometric characteristics of black spruce and summer temperature at tree line in northern Quebec. Can. J. For. Res. 2002, 32, 477–486. [Google Scholar] [CrossRef]
- Makinen, H.; Saranpää, P.; Linder, S. Effect of growth rate on fibre characteristics in Norway spruce (Picea abies (L.) Karst.). Holzforschung 2002, 56, 449–460. [Google Scholar] [CrossRef]
- Spicer, R.; Gartner, B.L.; Darbyshire, R.L. Sinuous stem growth in a Douglas-fir (Pseudotsuga menziesii) plantation: Growth patterns and wood-quality effects. Can. J. For. Res. 2000, 30, 761–768. [Google Scholar] [CrossRef]
- Krause, C.; Déry Bouchard, C.-A.; Plourde, P.-Y.; Mailly, D. Compression wood and stem horizontal displacement in black spruce and jack pine plantations in the boreal forest. For. Ecol. Manag. 2013, 302, 154–162. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Chui, Y.H. Selecting dry fiber weight for higher and better quality jack pine fiber production. Wood Fiber Sci. 1996, 28, 146–152. [Google Scholar]
- Watt, M.S.; Moore, J.R.; Façon, J.-P.; Downes, G.M.; Clinton, P.W.; Coker, G.; Davis, M.R.; Simcock, R.; Parfitt, R.L.; Dando, J.; et al. Modelling the influence of stand structural, edaphic and climatic influences on juvenile Pinus radiata dynamic modulus of elasticity. For. Ecol. Manag. 2006, 229, 136–144. [Google Scholar] [CrossRef]
- Cameron, A.D.; Gardiner, B.A.; Ramsay, J.; Drewett, T.A. Effect of early release from intense competition within high density natural regeneration on the properties of juvenile and mature wood of 40-year-old sitka spruce (Picea sitchensis (Bong.) Carr.). Ann. For. Sci. 2014, 72, 99–107. [Google Scholar] [CrossRef]
- Karlsson, L.; Mörling, T.; Bergsten, U. Influence of silvicultural regimes on the volume and proportion of juvenile and mature wood in boreal Scots pine. Silva Fenn. 2013, 47, 1–17. [Google Scholar] [CrossRef]
- Larson, P.R. Stem form of young Larix as influenced by wind and pruning. For. Sci. 1965, 11, 412–424. [Google Scholar]
- Vincent, M.; Duchesne, I. Modeling flexural properties in white spruce (Picea glauca) and jack pine (Pinus banksiana) plantation trees. Can. J. For. Res. 2014, 44, 82–91. [Google Scholar] [CrossRef]
- Parent, G.; Boulay, E.; Fortin, C. Ressources et Industries Forestières, Portrait Statistique Édition 2012 edMinistère des Ressources Naturelles et de la Faune du: Québec, QC, Canada, 2012; pp. 1–73.
- Waring, R.H.; Thies, W.G.; Muscato, D. Stem growth per unit of leaf area: A measure of tree vigor. For. Sci. 1980, 26, 112–117. [Google Scholar]
- Campoe, O.C.; Stape, J.L.; Nouvellon, Y.; Laclau, J.-P.; Bauerle, W.L.; Binkley, D.; Le Maire, G. Stem production, light absorption and light use efficiency between dominant and non-dominant trees of Eucalyptus grandis across a productivity gradient in Brazil. For. Ecol. Manag. 2013, 288, 14–20. [Google Scholar] [CrossRef]
- Dell, T.R.; Feduccia, D.P.; Campbell, T.E.; Mann, W.F., Jr.; Palmer, B.H. Yields of Unthinned Slash Pine Plantations on Cutover Sites in the West Gulf Region; Paper SO-147; U.S. Department of Agriculture Forest Service Research: New Orleans, LA, USA, 1979.
- Chmura, D.J.; Rahman, M.S.; Tjoelker, M.G. Crown structure and biomass allocation patterns modulate aboveground productivity in young loblolly pine and slash pine. For. Ecol. Manag. 2007, 243, 219–230. [Google Scholar] [CrossRef]
- Saucier, J.-P.; Robitaille, A.; Grondin, P. Cadre bioclimatique du Québec. In Manuel de Foresterie, 2nd ed.; Ordre des Ingénieurs Forestiers du Québec: Québec, QC, Canada, 2009; pp. 186–205. [Google Scholar]
- Environment Canada. Climatic Normals and Means, Shipshaw, Year 1971–2000, Atmospheric Environment Service; Environment Canada: Shipshaw, QC, Canada, 2014.
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: London, UK, 1968; pp. 1–73. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Guay, R.; Gagnon, R.; Morin, H. A new automatic and interactive tree ring measurement system based on a line scan camera. For. Chron. 1992, 68, 138–142. [Google Scholar] [CrossRef]
- Carmean, W.H. Site index curves for upland oaks in the central states. For. Sci. 1972, 18, 109–120. [Google Scholar]
- Xiang, W.; Leitch, M.; Auty, D.; Duchateau, E.; Achim, A. Radial trends in black spruce wood density can show an age- and growth-related decline. Ann. For. Sci. 2014, 71, 603–615. [Google Scholar] [CrossRef]
- Koubaa, A.; Zhang, S.Y.T.; Makni, S. Defining the transition from earlywood to latewood in black spruce based on intra-ring wood density profiles from X-ray densitometry. Ann. For. Sci. 2002, 59, 511–518. [Google Scholar] [CrossRef]
- Sauter, U.H.; Mutz, R.; Munro, B.D. Determining juvenile-mature wood transition in scots pine using latewood density. Wood Fiber Sci. 1999, 31, 416–425. [Google Scholar]
- Alteyrac, J.; Cloutier, A.; Zhang, S.Y. Characterization of juvenile wood to mature wood transition age in black spruce (Picea mariana (Mill.) B.S.P.) at different stand densities and sampling heights. Wood Sci. Technol. 2005, 40, 124–138. [Google Scholar] [CrossRef]
- ASTM. Standard Test Methods for Small Clear Specimens of Timber; D143-94; ASTM International: Philadelphia, PA, USA, 2007; pp. 20–52. [Google Scholar]
- Littel, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabernerger, O. SAS for Mixed Models, 2nd ed.; SAS Institute, Inc.: Cary, NC, USA, 2006; pp. 1–814. [Google Scholar]
- Kenward, M.G.; Roger, J.H. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef] [PubMed]
- Ung, C.H.; Végiard, S. Problèmes d'inférence statistique reliés à la transformation logarithmique en régression. Can. J. For. Res. 1988, 18, 733–738. [Google Scholar] [CrossRef]
- Larson, P.R.; Kretschmann, D.E.; Clark, A.; Isebrands, J.G. Formation and Properties of Juvenile Wood in Southern Pines: A Synopsis; USDA Forest Service General Technical Report FPL-GTR 129 General Technical Report; USDA Forest Service: Madison, WI, USA, 2001; pp. 1–42.
- McEvoy, T.J. Positive Impact Forestry: A Sustainable Approach to Managing Woodlands; Island Press: Washington, DC, USA, 2004; pp. 1–268. [Google Scholar]
- Ulvcrona, K.A.; Ulvcrona, T.; Nilsson, U.; Lundmark, T. Stand density and fertilization effects on aboveground allocation patterns and stem form of Pinus sylvestris in young stands. Scand. J. For. Res. 2014, 29, 197–209. [Google Scholar]
- Zhang, J.; Ritchie, M.W.; Maguire, D.A.; Oliver, W.W. Thinning ponderosa pine (Pinus ponderosa) stands reduces mortality while maintaining stand productivity. Can. J. For. Res. 2013, 43, 311–320. [Google Scholar] [CrossRef]
- Sharma, M.; Parton, J. Modeling stand density effects on taper for jack pine and black spruce plantations using dimensional analysis. For. Sci. 2009, 55, 268–282. [Google Scholar]
- Feduccia, D.P.; Dell, T.R.; Mann, W.F.; Campbell, T.E.; Palmer, B.H. Yields of Unthinned Loblolly Pine Plantations on Cutover Sites in the West Gulf Region; Paper SO-148; Agriculture Forest Service Research: New Orleans, LA, USA, 1979; pp. 1–88.
- Baldwin, V.C.J.; Polmer, B.H. Taper functions for unthinned longleaf pine plantation on cutover West Gulf sites. In General Technical Reports SO34, Proceedings of the 1st Biennial Southern Silviculture Research Conference, Atlanta, GA, USA, 6–7 November 1981; U.S. Department of Agriculture Forest Service Research: Washington, DC, USA, 1981; pp. 156–163. [Google Scholar]
- Valenti, M.A.; Cao, Q.V. Use of crown ratio to improve loblolly pine taper equations. Can. J. For. Res. 1986, 5, 1141–1145. [Google Scholar] [CrossRef]
- Fowells, H.A. Silvics of Forest Trees of the United States; Agriculture Handbook 271; U.S. Department of Agriculture: Washington, DC, USA, 1965; pp. 1–762.
- Bassow, S.L.; Bazzaz, F.A. Intra- and inter-specific variation in canopy photosynthesis in a mixed deciduous forest. Oecologia 1997, 109, 507–515. [Google Scholar] [CrossRef]
- Newton, P.F.; Jolliffe, P.A. Aboveground modular component responses to intraspecific competition within density-stressed black spruce stands. Can. J. For. Res. 1998, 28, 1587–1610. [Google Scholar] [CrossRef]
- Beaulieu, E.; Schneider, R.; Berninger, F.; Ung, C.-H.; Swift, E.D. Modeling jack pine branch characteristics in eastern Canada. For. Ecol. Manag. 2011, 262, 1748–1757. [Google Scholar] [CrossRef]
- Mäkinen, H. Effect of stand density on radial growth of branches of scots pine in southern and central Finland. Can. J. For. Res. 1999, 29, 1216–1224. [Google Scholar] [CrossRef]
- Sala, A.; Woodruff, D.R.; Meinzer, F.C. Carbon dynamics in trees: Feast of famine? Tree Physiol. 2012, 32, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Barbour, R.J.; Fayle, D.C.F.; Chauret, G.; Cook, J.; Karsh, M.B.; Ran, S. Breast-height relative density and radial growth in mature jack pine (Pinus banksiana) for 38 years after thinning. Can. J. For. Res. 1994, 24, 2439–2447. [Google Scholar] [CrossRef]
- Fengel, D.; Wegener, G. Wood Chemical Ultrastructure Reactions; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 1989; pp. 1–613. [Google Scholar]
- Wang, S.-Y.; Ko, C.-Y. Dynamic modulus of elasticity and bending properties of large beams of Taiwan-grown Japanese cedar from different plantation spacing sites. J. Wood Sci. 1998, 44, 62–68. [Google Scholar] [CrossRef]
- Chuang, S.-T.; Wang, S.-Y. Evaluation of standing tree quality of Japanese cedar grown with different spacing using stress-wave and ultrasonic-wave methods. J. Wood Sci. 2001, 47, 245–253. [Google Scholar] [CrossRef]
- Wang, X.; Ross, R.J.; McClellan, M.; Barbour, R.J.; Erickson, J.R.; Forsman, J.W.; McGinnis, G.D. Nondestructive evaluation of standing trees with a stress wave method. Wood Fiber Sci. 2001, 33, 522–533. [Google Scholar]
- Lasserre, J.P.; Mason, E.G.; Watt, M.S. The influence of initial spacing on corewood modulus of elasticity in a clonal experiment of 11-year-old Pinus radiata D. Don. N. Z. J. For. 2004, 49, 18–23. [Google Scholar]
- Waghorn, M.J.; Mason, E.G.; Watt, M.S. Influence of initial stand density and genotype on longitudinal variation in modulus of elasticity for 17-year-old Pinus radiata. For. Ecol. Manag. 2007, 252, 67–72. [Google Scholar] [CrossRef]
- Madsen, B. Structural Behaviour of Timber; Timber Engineering Ltd.: North Vancouver, BC, Canada, 1992; pp. 1–437. [Google Scholar]
- Schneider, R.; Zhang, S.Y.; Swift, D.E.; Bégin, J.; Lussier, J.-M. Predicting selected wood properties of jack pine following commercial thinning. Can. J. For. Res. 2008, 38, 2030–2043. [Google Scholar] [CrossRef]
- Pestorper, M.; Johansson, M.; Kliger, R. Distortion of Norway spruce timber Part 1. Variation of relevant wood properties. Holz Roh Werkst. 2001, 59, 94–103. [Google Scholar] [CrossRef]
- Panshin, A.J.; de Zeeuw, C. Textbook of Wood Technology; McGraw-Hill Book Co.: New York, NY, USA, 1980; pp. 1–772. [Google Scholar]
- Yang, K.C.; Hazenberg, G. Impact of spacing on tracheid length, relative density, and growth rate of juvenile wood and mature wood in Picea mariana. Can. J. For. Res. 1994, 24, 996–1007. [Google Scholar] [CrossRef]
- Yang, K.-C. Impact of spacing on width and basal area of juvenile and mature wood in Picea mariana and Picea glauca. Wood Fiber Sci. 1994, 26, 479–488. [Google Scholar]
- Kucera, B. A hypothesis relating current annual height increment to juvenile wood formation in Norway spruce. Wood Fiber Sci. 1994, 26, 152–167. [Google Scholar]
- Clark, A., III; Jordan, L.; Schimleck, L.; Daniels, R.F. Effect of initial planting spacing on wood properties of unthinned loblolly pine at age 21. For. Prod. J. 2008, 58, 78–83. [Google Scholar]
- Zhang, J.; Oliver, W.W.; Busse, M.D. Growth and development of ponderosa pine on sites of contrasting productivities: Relative importance of stand density and shrub competition effects. Can. J. For. Res. 2006, 36, 2426–2438. [Google Scholar] [CrossRef]
- Amateis, R.L.; Burkhart, H.E. Relating quantity, quality, and value of lumber to planting density for loblolly pine plantations. South. J. Appl. For. 2013, 37, 97–101. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Density (Tree/ha) | Spacing (m) |
|---|---|
| 1111 | 3 × 3 |
| 1666 | 2 × 3 |
| 2222 | 1.5 × 3 |
| 2500 | 2 × 2 |
| 3333 | 1.5 × 2 |
| 4444 | 1.5 × 1.5 |
| Effect (Fixed) | Height | DBH | Volume | Stem Taper * | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ndf | ddf | F | Pr > F | ddf | F | Pr > F | ddf | F | Pr > F | ddf | F | Pr > F | |
| Spacing | 5 | 10 | 0.82 | 0.565 | 9.6 | 9.58 | 0.002 | 10 | 3.70 | 0.037 | 251 | 0.65 | 0.659 |
| Live crown ratio | Biggest branch diameter | Longest branch * | |||||||||||
| ndf | ddf | F | Pr > F | ddf | F | Pr > F | ddf | F | Pr > F | ||||
| Spacing | 5 | 12.4 | 4.43 | 0.015 | 11.9 | 4.48 | 0.016 | 78 | 4.40 | 0.001 | |||
| Density (Tree/ha) | Number of Juvenile Tree Rings |
|---|---|
| 1111 | 9.1 (2.2) |
| 1666 | 7.7 (2.0) |
| 2222 | 8.3 (0.5) |
| 2500 | 8.1 (1.4) |
| 3333 | 8.1 (1.1) |
| 4444 | 7.1 (1.4) |
| Effect (Fixed) | % Latewood | Earlywood Density | Latewood Density | Wood Density | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ndf | ddf | F | Pr > F | ddf | F | Pr > F | ddf | F | Pr > F | ddf | F | Pr > F | |
| Spacing (S) | 5 | 93.5 | 1.54 | 0.185 | 9.96 | 1.94 | 0.175 | 12.2 | 0.32 | 0.892 | 10.4 | 1.33 | 0.323 |
| Wood type (WT) | 1 | 1996 | 184.96 | <0.001 | 1977 | 19.20 | <0.001 | 1977 | 1110.73 | <0.001 | 1976 | 777.32 | <0.001 |
| S × WT | 5 | 1996 | 2.91 | 0.013 | 1977 | 6.07 | <0.001 | 1977 | 5.45 | <0.001 | 1976 | 1.50 | 0.187 |
| Effect (Fixed) | MOE | MOR | |||||
|---|---|---|---|---|---|---|---|
| ndf | ddf | F | Pr > F | ddf | F | Pr > F | |
| Spacing (S) | 5 | 12.1 | 1.37 | 0.301 | 10.2 | 0.62 | 0.691 |
| Sample location (SP) | 1 | 19.3 | 51.74 | <0.001 | 95.5 | 52.88 | <0.001 |
| S × SP | 5 | 12.6 | 1.11 | 0.404 | 90.5 | 0.88 | 0.500 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hébert, F.; Krause, C.; Plourde, P.-Y.; Achim, A.; Prégent, G.; Ménétrier, J. Effect of Tree Spacing on Tree Level Volume Growth, Morphology, and Wood Properties in a 25-Year-Old Pinus banksiana Plantation in the Boreal Forest of Quebec. Forests 2016, 7, 276. https://doi.org/10.3390/f7110276
Hébert F, Krause C, Plourde P-Y, Achim A, Prégent G, Ménétrier J. Effect of Tree Spacing on Tree Level Volume Growth, Morphology, and Wood Properties in a 25-Year-Old Pinus banksiana Plantation in the Boreal Forest of Quebec. Forests. 2016; 7(11):276. https://doi.org/10.3390/f7110276
Chicago/Turabian StyleHébert, François, Cornelia Krause, Pierre-Yves Plourde, Alexis Achim, Guy Prégent, and Jean Ménétrier. 2016. "Effect of Tree Spacing on Tree Level Volume Growth, Morphology, and Wood Properties in a 25-Year-Old Pinus banksiana Plantation in the Boreal Forest of Quebec" Forests 7, no. 11: 276. https://doi.org/10.3390/f7110276
APA StyleHébert, F., Krause, C., Plourde, P.-Y., Achim, A., Prégent, G., & Ménétrier, J. (2016). Effect of Tree Spacing on Tree Level Volume Growth, Morphology, and Wood Properties in a 25-Year-Old Pinus banksiana Plantation in the Boreal Forest of Quebec. Forests, 7(11), 276. https://doi.org/10.3390/f7110276