
Early Stage Fine-Root Decomposition and Its Relationship with Root Order and Soil Depth in a Larix gmelinii Plantation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Decomposition Experiment
2.3. Chemical Analyses
2.4. Statistical Analysis
3. Results
3.1. Initial Substrate Chemistry
3.2. Effects of Root Class on Decomposition
3.3. Effects of Soil Depth on Decomposition
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R. Climate, leaf litter chemistry and leaf litter decomposition in terrestrial ecosystems: A triangular relationship. Oikos 1997, 79, 439–449. [Google Scholar] [CrossRef]
- Aerts, R. The freezer defrosting: Global warming and litter decomposition rates in cold biomes. J. Ecol. 2006, 94, 713–724. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.C.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mommer, L.; de Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Freschet, G.T.; Cornwell, W.K.; Wardle, D.A.; Elumeeva, T.G.; Liu, W.; Jackson, B.G.; Onipchenko, V.G.; Soudzilovskaia, N.A.; Tao, J.; Cornelissen, J.H. Linking litter decomposition of above-and below-ground organs to plant-soil feedbacks worldwide. J. Ecol. 2013, 101, 943–952. [Google Scholar] [CrossRef]
- Xia, M.; Talhelm, A.F.; Pregitzer, K.S. Fine roots are the dominant source of recalcitrant plant litter in sugar maple-dominated northern hardwood forests. New Phytol. 2015, 208, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Silver, W.L.; Miya, R.K. Global patterns in root decomposition: Comparisons of climate and litter quality effects. Oecologia 2001, 129, 407–419. [Google Scholar] [CrossRef]
- Hobbie, S.E.; Oleksyn, J.; Eissenstat, D.M.; Reich, P.B. Fine root decomposition rates do not mirror those of leaf litter among temperate tree species. Oecologia 2010, 162, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Goebel, M.; Hobbie, S.E.; Bulaj, B.; Zadworny, M.; Archibald, D.D.; Oleksyn, J.; Reich, P.B.; Eissenstat, D.M. Decomposition of the finest root branching orders: Linking belowground dynamics to fine-root function and structure. Ecol. Monogr. 2011, 81, 89–102. [Google Scholar] [CrossRef]
- Sun, T.; Mao, Z.; Dong, L.; Hou, Y.; Wang, X. Further evidence for slow decomposition of very fine roots using two methods: Litterbags and intact cores. Plant Soil 2013, 366, 633–646. [Google Scholar] [CrossRef]
- Sun, T.; Mao, Z.; Han, Y. Slow decomposition of very fine roots and some factors controlling the process: A 4-year experiment in four temperate tree species. Plant Soil 2013, 372, 445–458. [Google Scholar] [CrossRef]
- Xiong, Y.; Fan, P.; Fu, S.; Zeng, H.; Guo, D. Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees. Plant Soil 2013, 363, 19–31. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Wells, C.E.; Yanai, R.D.; Whitbeck, J. Building roots in a changing environment: Implications for root longevity. New Phytol. 2000, 147, 33–42. [Google Scholar] [CrossRef]
- Wells, C.E.; Eissenstat, D.M. Marked differences in survivorship among apple roots of different diameters. Ecology 2001, 82, 882–892. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- Guo, D.; Mitchell, R.J.; Withington, J.M.; Fan, P.P.; Hendricks, J.J. Endogenous and exogenous controls of root life span, mortality and nitrogen flux in a longleaf pine forest: Root branch order predominates. J. Ecol. 2008, 96, 737–745. [Google Scholar] [CrossRef]
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef] [PubMed]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Guo, D. Slow decomposition of lower order roots: A key mechanism of root carbon and nutrient retention in the soil. Oecologia 2010, 163, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.H.; Burke, I.C.; Lauenroth, W.K. Soil organic matter and nutrient availability responses to reduced plant inputs in shortgrass steppe. Ecology 1996, 77, 2516–2527. [Google Scholar] [CrossRef]
- Paul, E.A.; Follett, R.F.; Leavitt, S.W.; Halvorson, A.; Peterson, G.A. Radiocarbon dating for determination of soil organic matter pool sizes and dynamics. Soil Sci. Soc. Am. J. 1997, 61, 1058–1067. [Google Scholar] [CrossRef]
- Batjes, N.H. Total carbon and nitrogen in the soils of the world. Eur. J. Soil Sci. 1996, 47, 151–163. [Google Scholar] [CrossRef]
- Gill, R.A.; Burke, I.C. Influence of soil depth on the decomposition of Bouteloua gracilis roots in the shortgrass steppe. Plant Soil 2002, 241, 233–242. [Google Scholar] [CrossRef]
- Rovira, P.; Vallejo, V.R. Organic carbon and nitrogen mineralization under Mediterranean climatic conditions: The effects of incubation depth. Soil Biol. Biochem. 1997, 29, 1509–1520. [Google Scholar] [CrossRef]
- Rovira, P.; Vallejo, V.R. Mineralization of carbon and nitrogen from plant debris, as affected by debris size and depth of burial. Soil Biol. Biochem. 2002, 34, 327–339. [Google Scholar] [CrossRef]
- Sun, T.; Dong, L.; Wang, Z.; Lü, X.; Mao, Z. Effects of long-term nitrogen deposition on fine root decomposition and its extracellular enzyme activities in temperate forests. Soil Biol. Biochem. 2016, 93, 50–59. [Google Scholar] [CrossRef]
- Sun, T.; Dong, L.; Mao, Z.J. Simulated Atmospheric Nitrogen Deposition Alters Decomposition of Ephemeral Roots. Ecosystems 2015, 18, 1240–1252. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, D.; Wang, X.; Gu, J.; Mei, L. Fine root architecture, morphology, and biomass of different branch orders of two Chinese temperate tree species. Plant Soil 2006, 288, 155–171. [Google Scholar] [CrossRef]
- Eissenstat, D.M.; Volder, A. The efficiency of nutrient acquisition over the life of a root. In Nutrient Acquisition by Plants: An Ecological Perspective; BarririRad, H., Ed.; Ecological Studies 191; Springer: New York, NY, USA, 2004; pp. 185–220. [Google Scholar]
- Li, A.; Fahey, T.J.; Pawlowska, T.E.; Fisk, M.C.; Burtis, J. Fine root decomposition, nutrient mobilization and fungal communities in a pine forest ecosystem. Soil Biol. Biochem. 2015, 83, 76–83. [Google Scholar] [CrossRef]
- Koide, R.T.; Fernandez, C.W.; Peoples, M.S. Can ectomycorrhizal colonization of Pinus resinosa roots affect their decomposition? New Phytol. 2011, 191, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.; Coq, S.; Barantal, S.; Handa, I.T. Leaf traits and decomposition in tropical rainforests: Revisiting some commonly held views and towards a new hypothesis. New Phytol. 2011, 189, 950–965. [Google Scholar] [CrossRef] [PubMed]
- Hättenschwiler, S.; Gasser, P. Soil animals alter plant litter diversity effects on decomposition. Proc. Natl. Acad. Sci. USA 2005, 102, 1519–1524. [Google Scholar] [CrossRef] [PubMed]
- Handa, I.T.; Aerts, R.; Berendse, F.; Berg, M.P.; Bruder, A.; Butenschoen, O.; Chauvet, E.; Gessner, M.O.; Jabiol, J.; Makkonen, M. Consequences of biodiversity loss for litter decomposition across biomes. Nature 2014, 509, 218–221. [Google Scholar] [CrossRef] [PubMed]
- Hobbie, S.E.; Reich, P.B.; Oleksyn, J.; Ogdahl, M.; Zytkowiak, R.; Hale, C.; Karolewski, P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology 2006, 87, 2288–2297. [Google Scholar] [CrossRef]
- Ryan, M.G.; Melillo, J.M.; Ricca, A. A comparison of methods for determining proximate carbon fractions of forest litter. Can. J. For. Res. 1990, 20, 166–171. [Google Scholar] [CrossRef]
- Seifter, S.; Dayton, S.; Novic, B.; Muntwyler, E. The estimation of glycogen with the anthrone reagent. Arch. Biochem. 1950, 25, 191–200. [Google Scholar] [PubMed]
- Valenzuela-Estrada, L.R.; Vera-Caraballo, V.; Ruth, L.E.; Eissenstat, D.M. Root anatomy, morphology, and longevity among root orders in Vaccinium corymbosum (Ericaceae). Am. J. Bot. 2008, 95, 1506–1514. [Google Scholar] [CrossRef] [PubMed]
- McClaugherty, C.A.; Aber, J.D.; Mellilo, J.M. Decomposition dynamics of fine roots in forested ecosystems. Oikos 1984, 42, 378–386. [Google Scholar] [CrossRef]
- Lõhmus, K.; Ivask, M. Decomposition and nitrogen dynamics of fine roots of Norway spruce (Picea abies (L.) Karst.) at different sites. Plant Soil 1995, 168, 89–94. [Google Scholar] [CrossRef]
- Berg, B. Decomposition of root litter and some factors regulating the process: Long-term root litter decomposition in a Scots pine forest. Soil Biol. Biochem. 1984, 16, 609–617. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant Litter. Decomposition, Humus Formation, Carbon Sequestration, 3rd ed.; Springer: Heidelberg/Berlin, Germany, 2014; pp. 171–187. [Google Scholar]
- Tamura, M.; Tharayil, N. Plant litter chemistry and microbial priming regulate the accrual, composition and stability of soil carbon in invaded ecosystems. New Phytol. 2014, 203, 110–124. [Google Scholar] [CrossRef] [PubMed]
- Pregitzer, K.S.; Laskowski, M.J.; Burton, A.J.; Lessard, V.C.; Zak, D.R. Variation in sugar maple root respiration with root diameter and soil depth. Tree Physiol. 1998, 18, 665–670. [Google Scholar] [CrossRef] [PubMed]
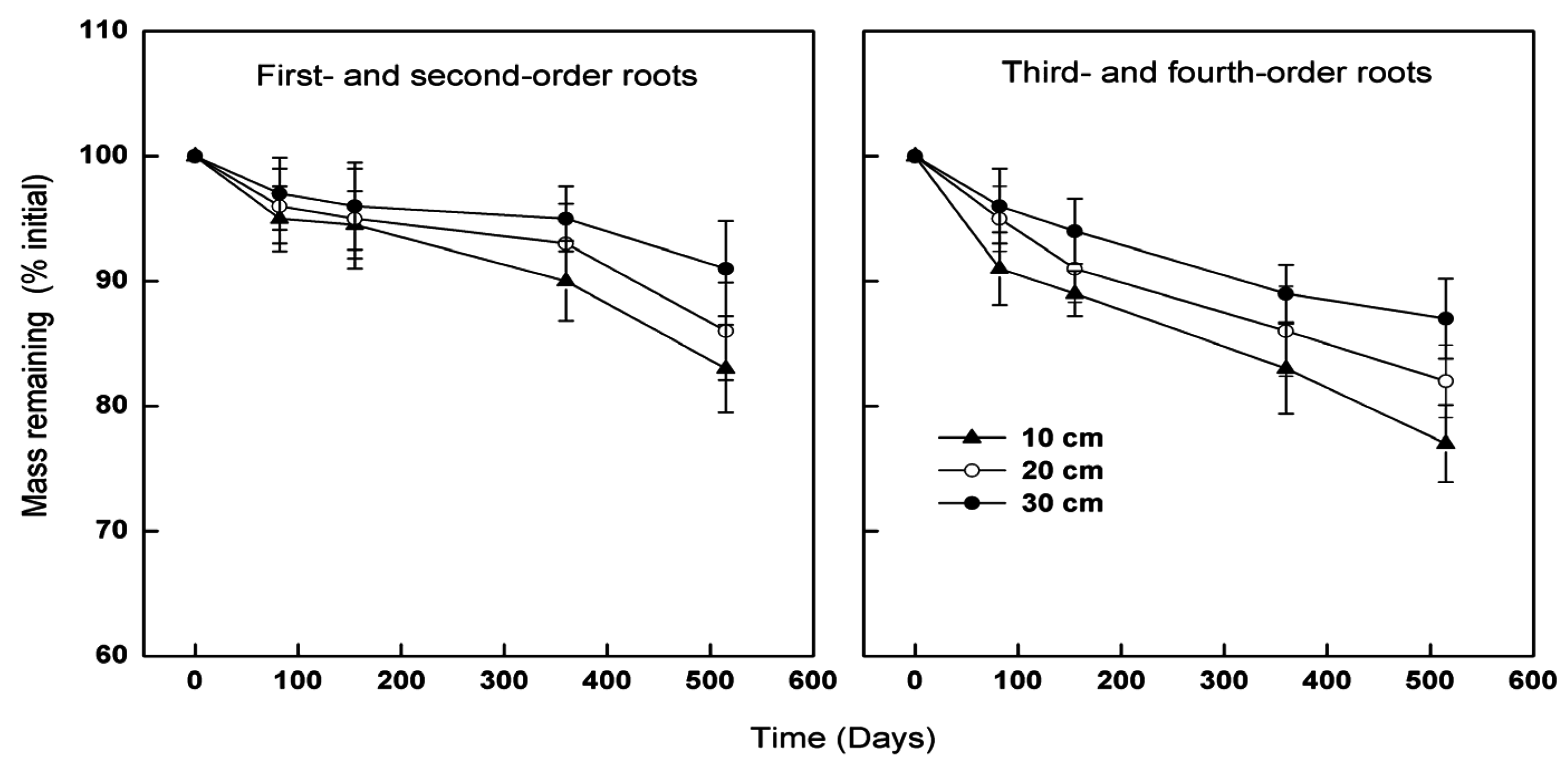

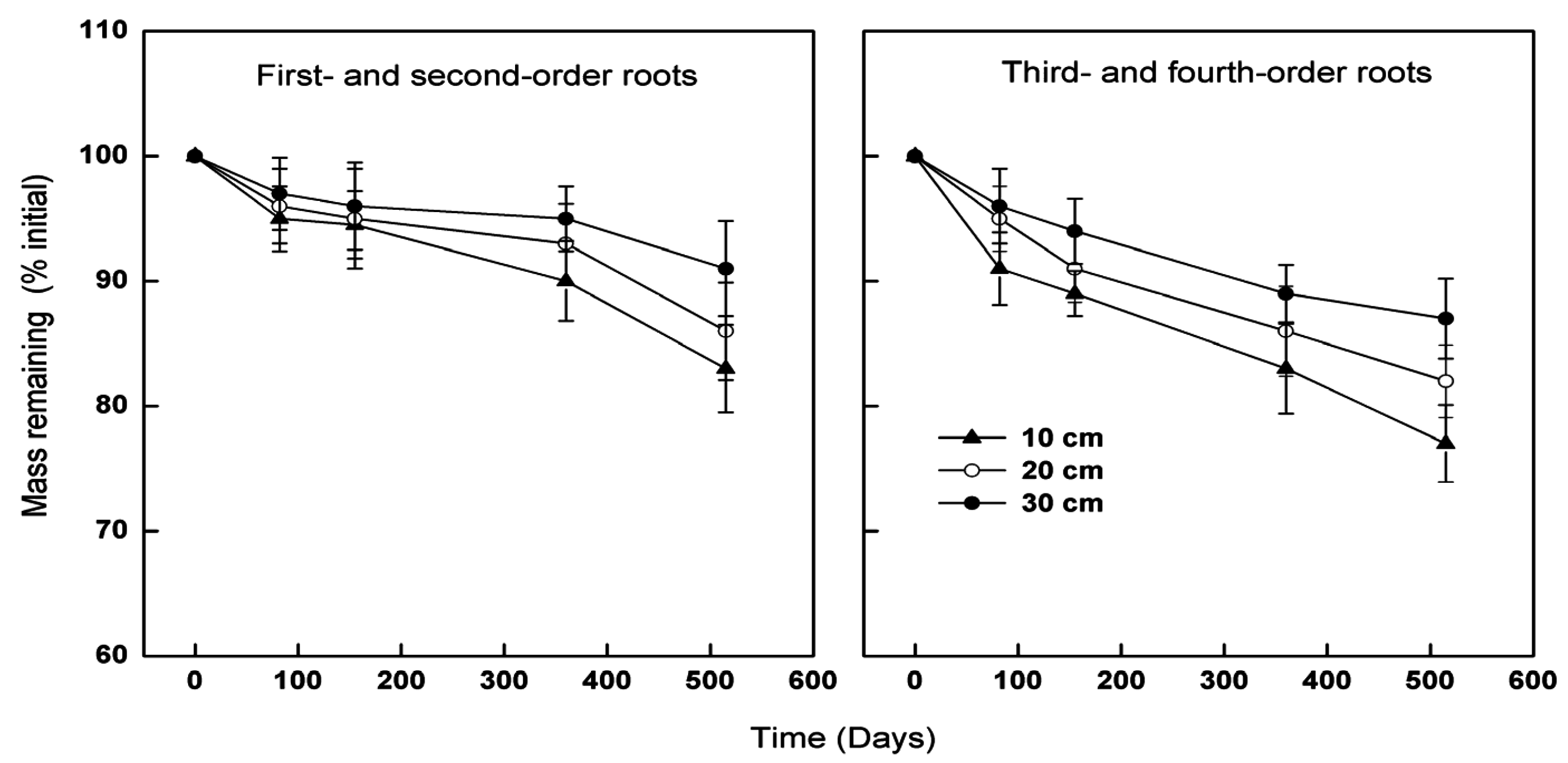{kind=link}
| Initial Root Nutrients (mg Element·g−1 Root) | Initial C-Fraction (mg Compound·g−1) | TNC (mg·g−1) | C:N | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Root Order | N | P | K | Ca | Mg | Extractives | AHF | AUF | ||
| order 1–2 | 26.1 (2.0) | 2.9 (0.2) | 6.1 (0.2) | 6.8 (1.5) | 3.8 (0.2) | 184.3 (15.1) | 280.6 (25.6) | 536.2 (45.1) | 106.8 (10.5) | 22.8 (4.5) |
| order 3–4 | 14.5 (1.6) | 1.2 (0.2) | 5.3 (0.4) | 4.3 (0.2) | 3.0 (0.3) | 215.9 (12.3) | 375.0 (23.2) | 409.7 (30.9) | 146.1 (16.8) | 42.1 (2.1) |
| Soil Depth | Root Class | k (Year−1) | r2 | SE | p |
|---|---|---|---|---|---|
| 10 cm | order 1–2 | 0.11 | 0.95 | 0.01 | <0.001 |
| order 3–4 | 0.17 | 0.87 | 0.03 | <0.001 | |
| 20 cm | order 1–2 | 0.09 | 0.90 | 0.01 | <0.001 |
| order 3–4 | 0.14 | 0.92 | 0.02 | <0.001 | |
| 30 cm | order 1–2 | 0.06 | 0.86 | 0.00 | <0.001 |
| order 3–4 | 0.10 | 0.88 | 0.01 | <0.001 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, T.; Dong, L.; Zhang, L.; Wu, Z.; Wang, Q.; Li, Y.; Zhang, H.; Wang, Z. Early Stage Fine-Root Decomposition and Its Relationship with Root Order and Soil Depth in a Larix gmelinii Plantation. Forests 2016, 7, 234. https://doi.org/10.3390/f7100234
Sun T, Dong L, Zhang L, Wu Z, Wang Q, Li Y, Zhang H, Wang Z. Early Stage Fine-Root Decomposition and Its Relationship with Root Order and Soil Depth in a Larix gmelinii Plantation. Forests. 2016; 7(10):234. https://doi.org/10.3390/f7100234
Chicago/Turabian StyleSun, Tao, Lili Dong, Lili Zhang, Zhijie Wu, Qingkui Wang, Yuanyuan Li, Hongguang Zhang, and Zhengwen Wang. 2016. "Early Stage Fine-Root Decomposition and Its Relationship with Root Order and Soil Depth in a Larix gmelinii Plantation" Forests 7, no. 10: 234. https://doi.org/10.3390/f7100234
APA StyleSun, T., Dong, L., Zhang, L., Wu, Z., Wang, Q., Li, Y., Zhang, H., & Wang, Z. (2016). Early Stage Fine-Root Decomposition and Its Relationship with Root Order and Soil Depth in a Larix gmelinii Plantation. Forests, 7(10), 234. https://doi.org/10.3390/f7100234