1. Introduction
White spruce (
Picea glauca [Moench] Voss) spans the North American continent from Newfoundland to Alaska and is one of the most important boreal conifer species for the Canadian forest industry. White spruce wood is highly valued for pulp and especially for lumber due to its good machining and structural properties [
1]. In common with other boreal species, white spruce is very slow growing compared to more temperate species such as radiata pine (
Pinus radiata D. Don), loblolly pine (
Pinus taeda L.) or even Douglas-fir (
Pseudotsuga menziesii [Mirb.] Franco). Tree breeding programs have been established for white spruce since the 1960s in different parts of Canada and the United States, but these have mainly focused on the improvement of growth and adaptive traits, even though faster growth is generally negatively correlated with important wood quality traits such as density and mechanical properties [
2,
3]. There is therefore a need to include wood traits in tree selection programs to ensure future wood supplies have the appropriate mechanical properties for structural applications and other end uses.
Previous studies have examined the genetic parameters of wood density and its components in white spruce from eastern [
2] and western Canada [
4], where abundant hybridization occurs between white spruce and Engleman spruce (
Picea engelmannii Parry ex Engelm.). In more recent work, genetic variation of other wood traits, such as decay resistance [
5], fibre length [
6] or veneer quality [
7] has been studied, although genetic control of those traits was found to be rather low. Moderate to high genetic control of wood traits related to mechanical end uses, such as density and microfibril angle (MFA) [
3,
8] (in hybrid spruce), or modulus of elasticity (MOE) [
9] (in eastern white spruce) has also been reported. These studies concluded that breeding for certain traits should result in considerable genetic gain and improvement of the next generation’s wood quality.
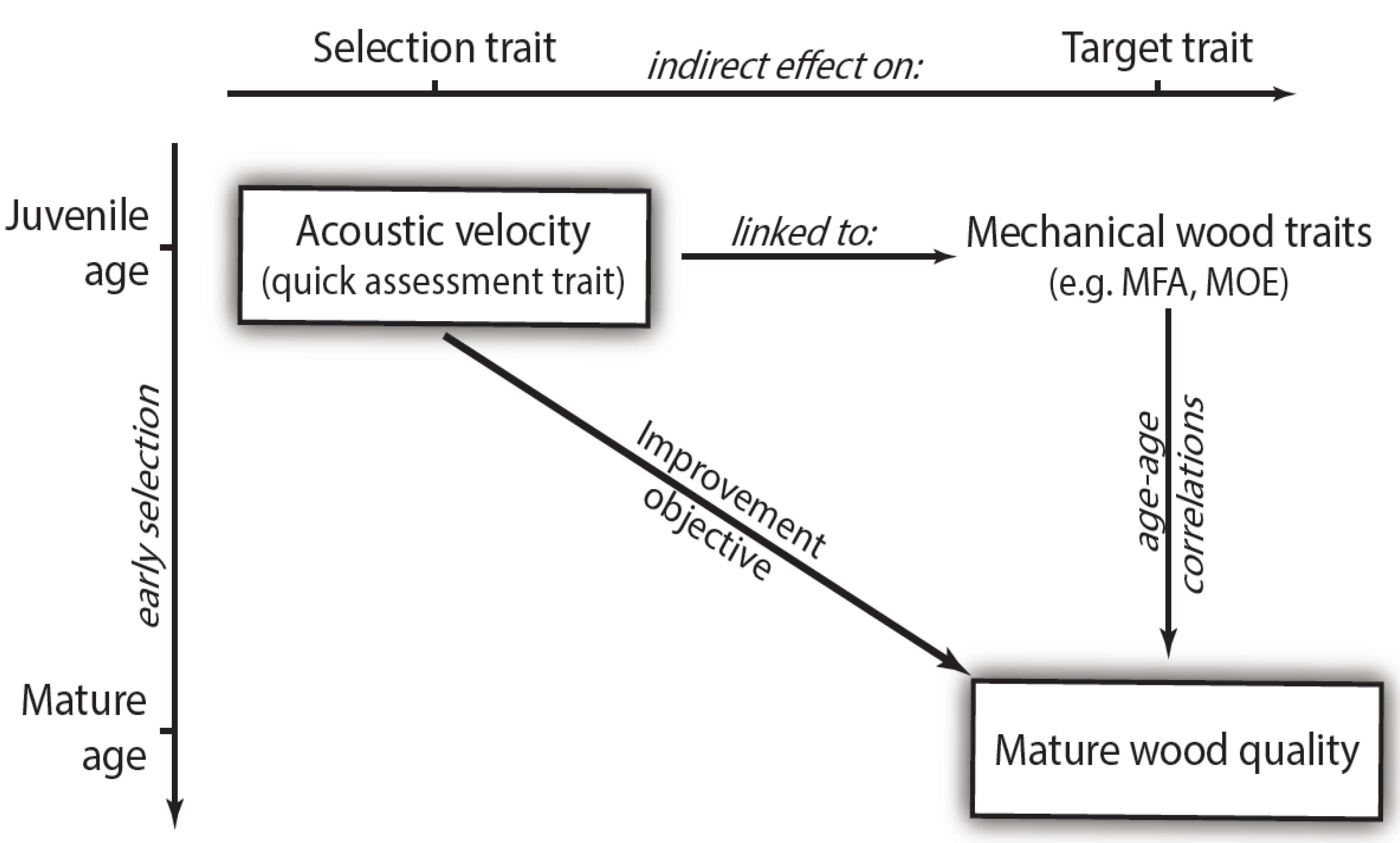
In advanced breeding generations, selection criteria are constantly adapted in order to improve additional wood quality traits. However, there are two major problems that hinder the application of cost-effective breeding strategies for the improvement of wood quality (
Figure 1). The first is the high cost of traditional wood trait evaluation methods, and the second is the length of time before mature wood can be evaluated, especially in slow growing boreal conifers. Since manufacturing investment cycles are far shorter than harvest rotation periods in such species, the development of non-destructive methods for evaluating young trees in order to select for adult wood quality is becoming increasingly important.
Figure 1.
Selection using an indirect method at juvenile tree age for improvement of mature wood quality. Illustration of the two axes: age and indirect selection. Repeated measurements of the quick-assessment traits are rarely available and age–age correlations mostly unknown. Evaluation of indirect early selection thus relies on the coincidental assessment of mechanical wood traits.
Figure 1.
Selection using an indirect method at juvenile tree age for improvement of mature wood quality. Illustration of the two axes: age and indirect selection. Repeated measurements of the quick-assessment traits are rarely available and age–age correlations mostly unknown. Evaluation of indirect early selection thus relies on the coincidental assessment of mechanical wood traits.
1.1. Indirect Assessment Methods
In recent decades, many indirect and non-destructive methods for wood quality evaluation on standing trees have been developed and most were reviewed by Wessels
et al. [
10]. Many of these techniques have also been evaluated for tree breeding. For example, spectroscopic analysis for the prediction of wood chemical composition (e.g., [
11]), or the assessment of mechanical wood traits (e.g., [
12,
13]) using increment cores. Such approaches, however, require the development of models and then further validation, which can be complicated and expensive depending on the trait of interest and its variation in the breeding populations. Other in-field assessment methods such as Pilodyn pin penetration or drill resistance measurements have also been used in genetic studies, since both are strongly correlated to wood density (e.g., [
14,
15]).
Other promising methods for the evaluation of wood mechanical properties are tools that can measure the transit time of sound waves in lumber or standing trees. These tools are easier to handle than traditional destructive mechanical testing. Acoustic velocity is directly related to dynamic wood stiffness, or modulus of elasticity (MOE) [
16], which is highly correlated with modulus of rupture (MOR). Since MOE and MOR are affected by intrinsic clear-wood properties such as microfibril angle and wood density, the latter are very important determinants of timber performance for structural end-uses [
17]. Both MOE and MOR have been noted to increase linearly with wood density [
18]. Alteyrac
et al. [
19], for example, reported that MFA explained 50% and more than 70% of variation in black spruce MOR and MOE, respectively. However, the mechanical properties of lumber pieces may be reduced by the occurrence of knots, depending on their size and location [
17,
18]. In contrast, knots have very little influence on acoustic velocity because the dilatational wave tends to take the fastest path of travel around local defects. In consequence, an acoustic velocity measurement gives a mean estimate of MOE over the entire stem section in which it has been measured, but it may overestimate local minima due to knots or other defects in pieces of solid wood [
20].
Numerous studies have investigated the links between acoustic measurements in standing trees, logs and the mechanical properties of wood samples from the same stem. A strong relationship between acoustic measurements on standing trees and felled logs was reported by Grabianowski
et al. [
21] in radiata pine. Using acoustic velocity in logs, Achim
et al. [
22] reported a strong correlation between dynamic and static MOE in trembling aspen (
Populus tremuloides Michx.), and good correlations were reported between standing tree acoustic velocity and MOR in Scots pine (
Pinus sylvestris L.) [
23].
Largely due to these encouraging results, the use of acoustic measurements for the evaluation of mechanical wood properties has promising applications in a tree breeding context [
24,
25,
26]. Already, acoustic devices are being deployed on an operational basis in various tree improvement programs, particularly for fast growing pine species, such as radiata pine [
27]. However, somewhat contrasting estimates of genetic parameters, such as heritability, have been reported for different species. For example, heritability values for acoustic velocity ranged from below 0.2 in Douglas-fir [
28] to almost 0.6 in radiata pine [
29]. To our knowledge, no genetic study has directly assessed the use of acoustics in white spruce breeding programs.
1.2. Early Selection
Besides the cost-effective evaluation of traits, early selection efficiency
i.e., the possibility to select individuals at a juvenile age that will have superior rotation-age properties plays an important role in tree breeding. Most wood traits in conifers vary systematically from juvenility to maturity [
30]. These patterns are essentially dependent on cambial age but may also be influenced by environmental variables and other traits, such as annual ring-width [
31,
32]. In order to evaluate if adult traits are linked to juvenile traits and whether these relationships can be used as a basis for early selection, a common practice in genetics is to analyze age–age correlations. In white spruce, mechanical wood traits, such as MOE and MOR, along with microstructural properties, such as MFA, show high age–age correlation and thus have strong potential for effective early selection [
33]. For such purposes, non-destructive techniques such as the acoustic evaluation of standing trees to indirectly measure stiffness carry a clear advantage over destructive methods. Indeed, Li
et al. [
34] found velocity instruments to be excellent tools for the evaluation of wood stiffness in 8-year-old slash pine. Similar results were reported by Eckard
et al. [
25] in a study on 8-year-old loblolly pine clones.
At the start of the current study, it was not known how a combination of slow growth, small diameter stems and a high incidence of live branches along the whole length of the stem would influence the accuracy of the velocity measurements and, consequently, the suitability of acoustic tools for use in breeding programmes for slow-growing boreal conifers. The overall objective of this study was to assess whether early selection based on acoustic velocity can improve white spruce mechanical wood properties. Specific objectives were (1) to evaluate the selection efficiency of acoustic tools in young trees of this species, and (2) investigate the relationship between acoustic velocity, tree growth-related variables and mechanical wood traits in young and mature white spruce trees.
3. Results
Table 1 gives an overview of wood trait means and coefficients of variation (CV) at the three sites analyzed in this study. As expected, acoustic velocity was higher, and MFA lower, in the mature trees. Earlywood density showed similar dispersion across all three sites. Average ring density as well as latewood density and latewood width were higher in the mature trees. The CVs were generally higher in young trees for all traits expect for earlywood density and MFA. Of the young trees, those from the Cabano plantation were established in a more heterogeneous environment, which was reflected in slower height and diameter growth, lower acoustic velocity, higher MFA and slightly higher overall variation compared to the trees from Saint-Casimir.
Table 1.
Means and coefficient of variation (CV, in parentheses) of wood traits at each site.
Table 1.
Means and coefficient of variation (CV, in parentheses) of wood traits at each site.
| | Mature trees | Young trees |
|---|
| | Mirabel | Cabano | Saint-Casimir |
| Sample Trees | 102 | 883 | 912 |
| Ring Number | 22.09 (6.75) | 7.40 (11.18) | 7.59 (11.74) |
| Velocity [km s−1] | 4.17 (10.34) | 2.70 (13.69) | 2.87 (12.24) |
| MFA [°] | 19.51 (21.94) | 36.25 (9.24) | 33.09 (8.53) |
| MOESS [GPa] | 8.80 (20.97) | – | – |
| Vel² × Dens [GPa] | 7.30 (23.47) | 2.84 (28.29) | 3.33 (27.03) |
| Density [kg m−³] | 413.91 (7.49) | 386.90 (9.73) | 396.13 (7.31) |
| EW density [kg m−³] | 361.19 (7.01) | 361.77 (7.32) | 368.84 (6.24) |
| LW density [kg m−³] | 713.55 (6.59) | 661.11 (3.74) | 691.64 (3.7) |
| Ring width [mm] | 3.74 (16.46) | 4.3 (20.03) | 4.21 (19.72) |
| EW width [mm] | 3.27 (17.66) | 3.87 (23.77) | 3.82 (21.31) |
| LW width [mm] | 0.62 (24.3) | 0.43 (48.5) | 0.39 (34.5) |
| EW percentage | 0.83 (4.03) | 0.89 (7.16) | 0.90 (4.04) |
| DBH [cm] | 18.89 (15.89) | 7.48 (21.79) | 7.58 (22.24) |
| Height [m] | 12.62 (12.64) | 4.91 (16.09) | 5.64 (17.58) |
3.1. Genetic Parameters for Young Trees
Parameter estimates were obtained from trees of 59 controlled pollinated crosses that are currently being evaluated in the Quebec white spruce breeding program.
Table 2 presents individual heritability estimates as well as genetic and phenotypic correlations for the individual tree level.
Table 3 contains family heritability and estimates for genetic gain based on family means. Associated errors of the estimates were rather low and varied between 0 and 0.19 (
Table 2,
Table 3). Moderate individual heritability estimates were found for tree height and for most wood traits (
Table 2); the highest heritability estimates were for acoustic velocity (
h² = 0.38) and earlywood density (
h² = 0.41). The heritability of MFA (
h² = 0.22) was rather low and of the same range as that of growth increment-related traits, such as DBH, average ring width and earlywood width. Latewood density and width were also under weak genetic control.
Acoustic velocity gave moderate phenotypic correlation (
rP = −0.49) and strong genetic correlation (
rG = −0.84) with MFA (
Table 2) indicating that both traits may be controlled by largely the same set of genes. However, the relationship between velocity and wood density as well as other growth-related traits was rather weak. While the correlation between velocity and radial increment (ring width and DBH) was very low in young trees, results indicate a positive but weak correlation between height growth and acoustic velocity (
rP = 0.12,
rG = 0.17). At the genetic level, faster growth was associated with low MFA, indicating a tendency for higher mechanical properties in faster-growing genotypes. Moderate negative correlations were found for MFA with DBH (
rG = −0.38), height (
rG = −0.44), and ring width (
rG = −0.37), as well as with earlywood width (
rG = −0.34) and earlywood percentage (
rG = −0.57); however, the equivalent phenotypic correlations were quite weak.
Table 2.
Genetic parameter estimates for the combined site analyses of young trees. Genetic correlations are above and phenotypic correlations are below the diagonal. Individual heritability estimates are in bold on the diagonal. Errors associated to correlations and heritability estimates are in parentheses.
Table 2.
Genetic parameter estimates for the combined site analyses of young trees. Genetic correlations are above and phenotypic correlations are below the diagonal. Individual heritability estimates are in bold on the diagonal. Errors associated to correlations and heritability estimates are in parentheses.
| | Velocity | MFA | Vel² × Dens | Density | EW density | LW density | Ring width | EW width | LW width | EW percentage | DBH | Height |
|---|
| Velocity | 0.38 (0.07) | −0.84 (0.07) | 0.96 (0.01) | 0.13 (0.03) | 0.20 (0.02) | 0.10 (0.18) | 0.03 (0.17) | 0.06 (0.17) | −0.18 (0.17) | 0.19 (0.18) | 0.01 (0.02) | 0.17 (0.02) |
| MFA | −0.49 (0.02) | 0.22 (0.05) | −0.72 (0.09) | 0.23 (0.16) | 0.13 (0.16) | 0.05 (0.19) | −0.37 (0.16) | −0.44 (0.16) | 0.38 (0.16) | −0.57 (0.15) | −0.38 (0.16) | −0.44 (0.05) |
| Vel² × Dens | 0.95 (0.00) | −0.45 (0.03) | 0.36 (0.07) | 0.38 (0.14) | 0.44 (0.13) | 0.19 (0.17) | −0.09 (0.17) | −0.10 (0.17) | 0.02 (0.17) | −0.10 (0.02) | −0.06 (0.17) | 0.16 (0.16) |
| Density | 0.12 (0.01) | 0.04 (0.03) | 0.41 (0.03) | 0.32 (0.06) | 0.99 (0.01) | 0.36 (0.16) | −0.39 (0.14) | −0.51 (0.13) | 0.70 (0.10) | −0.86 (0.06) | −0.41 (0.14) | −0.28 (0.15) |
| EW Density | 0.14 (0.01) | 0.04 (0.03) | 0.40 (0.03) | 0.94 (0.01) | 0.41 (0.07) | 0.31 (0.16) | −0.36 (0.05) | −0.48 (0.13) | 0.63 (0.11) | −0.78 (0.08) | −0.40 (0.14) | −0.28 (0.15) |
| LW Density | 0.22 (0.03) | −0.17 (0.03) | 0.30 (0.03) | 0.35 (0.03) | 0.22 (0.03) | 0.14 (0.04) | −0.14 (0.19) | −0.21 (0.19) | 0.40 (0.17) | −0.38 (0.18) | −0.04 (0.19) | 0.10 (0.18) |
| Ring width | −0.11 (0.03) | −0.09 (0.03) | −0.23 (0.03) | −0.50 (0.02) | −0.45 (0.02) | −0.20 (0.03) | 0.23 (0.05) | 0.99 (0.01) | 0.15 (0.18) | 0.42 (0.16) | 0.96 (0.01) | 0.83 (0.06) |
| EW width | −0.10 (0.03) | −0.10 (0.03) | −0.26 (0.03) | −0.60 (0.02) | −0.54 (0.02) | −0.23 (0.03) | 0.98 (0.00) | 0.22 (0.05) | −0.01 (0.19) | 0.56 (0.13) | 0.95 (0.03) | 0.79 (0.03) |
| LW width | −0.01 (0.03) | 0.07 (0.03) | 0.17 (0.03) | 0.63 (0.02) | 0.50 (0.02) | 0.19 (0.03) | −0.04 (0.03) | −0.22 (0.03) | 0.16 (0.04) | −0.83 (0.06) | 0.15 (0.18) | 0.19 (0.17) |
| EW percentage | −0.01 (0.03) | −0.10 (0.03) | −0.21 (0.03) | −0.78 (0.01) | −0.62 (0.02) | −0.25 (0.03) | 0.46 (0.02) | 0.62 (0.02) | −0.87 (0.01) | 0.12 (0.04) | 0.42 (0.16) | 0.28 (0.03) |
| DBH | −0.05 (0.00) | −0.12 (0.03) | −0.15 (0.03) | −0.40 (0.03) | −0.40 (0.03) | −0.06 (0.03) | 0.80 (0.01) | 0.78 (0.01) | −0.01 (0.03) | 0.35 (0.02) | 0.23 (0.05) | 0.93 (0.03) |
| Height | 0.12 (0.01) | −0.19 (0.01) | 0.06 (0.04) | −0.31 (0.03) | −0.31 (0.03) | 0.02 (0.03) | 0.66 (0.02) | 0.63 (0.03) | 0.04 (0.03) | 0.27 (0.02) | 0.82 (0.01) | 0.33 (0.07) |
Similarly, there was a weak to moderate genetic link between MFA and density (rG = 0.23) and between MFA and latewood width (rG = 0.38). Their phenotypic correlations were again close to zero. Genetic parameters of Vel² × Dens were close to the estimates for its individual components, especially velocity. However, it is interesting to note that while phenotypic correlations between Vel² × Dens and radial growth parameters (i.e., ring width, earlywood percentage and DBH) were weak and negative, virtually no correlation was found on the genetic level.
Genetic and phenotypic correlations were strongly positive between density and latewood width (rP = 0.35, rG = 0.70) and moderately negative for density and traits related to growth, which was expected. Additionally, earlywood and latewood width often showed opposite correlations with the same traits especially on the genetic level.
Table 3.
Estimates of family heritability and genetic gain based on family means in young trees. A selection intensity of α = 0.05 was used for the calculation of gain.
Table 3.
Estimates of family heritability and genetic gain based on family means in young trees. A selection intensity of α = 0.05 was used for the calculation of gain.
| | Heritability | Gain |
|---|
| Velocity | 0.78 (0.04) | 9.66 |
| MFA | 0.69 (0.06) | 4.96 |
| Vel² × Dens | 0.77 (0.04) | 20.17 |
| Density | 0.78 (0.04) | 5.89 |
| EW density | 0.82 (0.04) | 5.44 |
| LW density | 0.60 (0.08) | 1.44 |
| Ring width | 0.66 (0.06) | 10.71 |
| EW width | 0.64 (0.06) | 10.46 |
| LW width | 0.67 (0.06) | 18.64 |
| EW percentage | 0.58 (0.08) | 2.04 |
| DBH | 0.69 (0.06) | 12.15 |
| Height | 0.77 (0.04) | 11.99 |
Based on the heritability estimates and the phenotypic variance components of wood traits, we assessed the expected genetic gain under a standardized selection scenario (
i = 2.06 in Equation 6, which means that about 5% of individuals (
Table 4) or families (
Table 3) are retained during selection. Despite the moderate individual heritability estimates of most traits analyzed in this study, the expected genetic gain for a specific trait over its mean value varied from about 1% to almost 20% (for selection on the individual tree level and the family level). For MFA, the expected gain was almost 5%, while a gain of more than 10% could be expected when selection is based on growth-related traits. The expected gain for density was approximately 5.5%. Selection for acoustic velocity could improve this trait by almost 10%, which is encouraging for its practical application in a tree breeding program. The highest gain was expected for Vel² × Dens, which could be improved by 20% in this selection scenario (
Table 4).
Heritability estimates based on family means were moderate to high for all traits (
Table 3) and more than twice as big as their individual counterparts. The relative differences in genetic control between traits did not change. The estimated genetic gain was of the same magnitude as the genetic response expected on the individual tree level. According to our results, a higher gain could be expected based on selection of entire families for DBH (
GInd = 10.20%,
GFam = 12.15%) and for MFA (
GInd = 4.02%,
GFam = 4.96%).
Table 4 also shows the selection efficiency of different traits that could be obtained by indirect selection of another trait. Selection based on acoustic velocity had a small effect on both ring width and DBH, and only 14% of the total possible gain in wood density was predicted when selection was based on velocity alone. At the same time, 18% of the possible gain in tree height was estimated. However, the use of acoustic velocity had rather little effect on other growth- and density-related traits. In contrast, the indirect selection efficiency for MFA was theoretically greater than 100%, indicating that genetic gains for MFA were higher through the indirect selection for acoustic velocity than through direct selection for MFA itself. This was related to the important difference in genetic control of both MFA and velocity. Selecting for lower MFA values at the same time indirectly improved the gain from DBH and height by about a third in each case (
Eind = −0.37 and
Eind = −0.35, respectively). A selection strategy for improving tree height or DBH could at the same time produce gains of earlywood ring width (respectively
Eind = 0.96,
Eind = 1.00) and overall ring width (respectively
Eind = 0.98,
Eind = 0.99). This strategy also resulted in a loss in wood density which could counterbalance a third of its possible gain and had a small impact on velocity (height selection
Eind = 0.16, DBH,
Eind = 0.00).
Table 4.
Efficiency of indirect selection among different wood traits. Estimates for genetic gain (in percent of the population mean) are in bold on the diagonal.
Table 4.
Efficiency of indirect selection among different wood traits. Estimates for genetic gain (in percent of the population mean) are in bold on the diagonal.
| | | Indirect effect on: | Density | EW density | LW density | Ring width | EW width | LW width | EW percentage | DBH | Height |
|---|
| Velocity | MFA | Vel² × Dens |
|---|
| Selection trait: | Velocity | 9.87 | −1.10 | 0.99 | 0.14 | 0.20 | 0.17 | 0.03 | 0.07 | −0.27 | 0.33 | 0.00 | 0.18 |
| MFA | −0.64 | 4.02 | −0.57 | 0.19 | 0.09 | 0.06 | −0.36 | −0.44 | 0.44 | −0.75 | −0.37 | −0.35 |
| Vel² × Dens | 0.93 | −0.92 | 19.98 | 0.40 | 0.42 | 0.31 | −0.11 | −0.12 | 0.03 | −0.17 | −0.07 | 0.17 |
| Density | 0.12 | 0.28 | 0.36 | 5.48 | 0.88 | 0.55 | −0.46 | −0.62 | 0.97 | −1.37 | −0.48 | −0.27 |
| EW density | 0.21 | 0.17 | 0.47 | 1.12 | 5.52 | 0.53 | −0.48 | −0.65 | 0.99 | −1.41 | −0.53 | −0.31 |
| LW density | 0.06 | 0.04 | 0.12 | 0.23 | 0.18 | 1.05 | −0.11 | −0.17 | 0.37 | −0.40 | −0.03 | 0.06 |
| Ring width | 0.02 | −0.38 | −0.07 | −0.33 | −0.27 | −0.18 | 9.37 | 1.02 | 0.18 | 0.58 | 0.97 | 0.68 |
| EW width | 0.04 | −0.43 | −0.08 | −0.42 | −0.35 | −0.26 | 0.95 | 9.87 | −0.01 | 0.74 | 0.92 | 0.63 |
| LW width | −0.12 | 0.32 | 0.02 | 0.50 | 0.40 | 0.44 | 0.13 | −0.01 | 13.43 | −0.95 | 0.13 | 0.13 |
| EW percentage | 0.11 | −0.43 | −0.06 | −0.53 | −0.43 | −0.36 | 0.31 | 0.43 | −0.72 | 1.40 | 0.31 | 0.17 |
| DBH | 0.00 | −0.38 | −0.04 | −0.35 | −0.30 | −0.05 | 0.96 | 0.98 | 0.18 | 0.56 | 10.20 | 0.76 |
| Height | 0.16 | −0.55 | 0.16 | −0.29 | −0.26 | 0.15 | 1.00 | 0.99 | 0.27 | 0.47 | 1.13 | 11.54 |
3.2. Phenotypic Modeling of Mechanical Wood Traits
In the previous section, genetic and phenotypic correlations between acoustic velocity and MFA have been shown to differ in magnitude. To better understand why phenotypic correlations between both traits were considerably weaker than their genotypic counterpart, we investigated which traits—besides velocity—may influence mechanical wood quality on the observational level.
3.2.1. Models for Predicting MFA in Young Trees
There was a weak to moderate relationship between acoustic velocity and MFA in young trees (
Table 5). The best model for predicting MFA from velocity (model 4) contained both velocity and HD-ratio in the fixed effects, which was slightly better in terms of AIC and fit indices than the model with velocity and DBH as predictors (model 3). The average density of the last rings was not a significant predictor (model 5). Overall, the amount of variation explained by the fixed effects was between 24.1 and 25.3%, with only a small increase if fit indices included the family random effect (
Table 3).
Table 5.
Models for predicting MFA in young trees showing parameter estimates, AIC and fit indices calculated from the fixed, and fixed plus random effects.
Table 5.
Models for predicting MFA in young trees showing parameter estimates, AIC and fit indices calculated from the fixed, and fixed plus random effects.
| Model | Fixed effects parameters | AIC | Fit indices (
R²) |
|---|
| Intercept | Vel | Vel2 | DBH | HD | Density | Fixed | Family |
|---|
| 1 | 47.16 ** | −4.52 ** | – | – | – | – | 8388.83 | 0.241 | 0.276 |
| 2 | 40.86 ** | – | −0.80 ** | – | – | – | 8389.99 | 0.240 | 0.276 |
| 3 | 48.39 ** | −4.53 ** | – | −0.02 * | – | – | 8381.12 | 0.246 | 0.279 |
| 4 | 49.34 ** | −4.13 ** | – | – | −0.46 ** | – | 8359.31 | 0.253 | 0.293 |
| 5 | 48.05 ** | −4.50 ** | – | – | – | −0.002 ns | 8389.76 | 0.240 | 0.278 |
3.2.2. Models for Predicting MFA and MOESS in Mature Trees
The relationship between MFA and acoustic velocity was moderate to strong in mature trees (
Table 6). Fit indices of fixed effects ranged from 52.5% to 54.6%, considerably higher than in the models for juvenile trees. Similar to juvenile trees, the HD-ratio was significant in the mature trees; however, the best model contained acoustic velocity as the only independent variable. The models describing MOE were slightly better than the MFA models and their fit indices of fixed effects varied between 55.2% and 62.5%. Besides acoustic velocity, DBH was found to significantly contribute to explaining MOE. The best model included the HD-ratio and the fixed effects explained more than 62% of the variation of MOE. Including the family random effect increased the fit indices by around 10% for MFA models or by 15% for MOE models. In any case, using the square of acoustic velocity for modelling MFA or MOE did not significantly improve the model fit in either the young or the mature trees.
Table 6.
Models for predicting area-weighted core mean microfibril angle (MFA) and modulus of elasticity (MOE) in mature trees showing parameter estimates, AIC and fit indices calculated from the fixed, and fixed plus random effects.
Table 6.
Models for predicting area-weighted core mean microfibril angle (MFA) and modulus of elasticity (MOE) in mature trees showing parameter estimates, AIC and fit indices calculated from the fixed, and fixed plus random effects.
| Trait | Model | Fixed effects parameters | AIC | Fit indices (
R²) |
|---|
| Intercept | Vel | Vel2 | DBH | HD | Density | Fixed | Family |
|---|
| MFA | 1 | 48.55 ** | −6.74 ** | – | – | – | – | 512.81 | 0.538 | 0.647 |
| | 2 | 34.23 ** | – | −0.78 ** | – | – | – | 515.64 | 0.525 | 0.635 |
| | 3 | 46.23 ** | −6.68 ** | – | 0.11 ns | – | – | 513.51 | 0.546 | 0.635 |
| | 4 | 49.56 ** | −6.52 ** | – | – | −2.84 ** | – | 514.16 | 0.543 | 0.632 |
| | 5 | 45.78 ** | −6.88 ** | – | – | – | −0.008 ns | 514.13 | 0.539 | 0.660 |
| MOE | 6 | −3.76 ** | 2.92 ** | – | – | – | – | 328.72 | 0.558 | 0.771 |
| | 7 | 2.40 ** | – | 0.34 ** | – | – | – | 330.57 | 0.552 | 0.762 |
| | 8 | −1.19 ns | 2.86 ** | – | −0.12 * | – | – | 320.88 | 0.615 | 0.759 |
| | 9 | −5.41 ** | 2.55 ** | – | – | 4.78 ** | – | 318.79 | 0.625 | 0.760 |
4. Discussion
Genetic parameter estimates and phenotypic prediction models from 15-year-old and 32-year-old white spruce progeny tests were used to evaluate acoustic velocity as a selection trait for wood quality. The moderate heritability of acoustic velocity, the promising genetic gains and the strong relationship with MFA and MOE make it possible to use it as a surrogate for mechanical properties in white spruce tree improvement strategies. Heritability of acoustic velocity in young white spruce (
h² = 0.38) was found to be of the same magnitude as estimates for other wood traits [
9]. A comparable degree of genetic control of acoustic velocity was reported by Li
et al. [
34] from a study in young slash pine (
h² = 0.42), although somewhat higher heritability was estimated in young radiata pine (
h² = 0.47) by Kumar
et al. [
45].
The moderate genetic control of velocity led to an expected gain that was of the same magnitude as the expected improvement of the growth traits DBH and height. Combining acoustic velocity and density to an approximate measure of dynamic MOE (Vel² × Dens) increased the variation that is accessible for selection and led to the highest expected gain in the panel of analyzed traits. However, the use of density as an early selection trait requires caution, since its selection efficiency at a very early age has been found to be low in a previous work on white spruce [
33]. The rather small correlation between acoustic velocity and wood density in this study is in contrast to earlier results on Douglas-fir [
28]. This is probably linked to differences in wood structure, since white spruce has lower density, a more gradual earlywood–latewood transition and a lower latewood percentage than Douglas-fir. In addition, wood density in spruce species typically follows the “Type II” pattern of a rapid decline from a maximum value near the pith followed by a gradual increase to an intermediate value [
17]. The high density wood close to the pith also has the characteristically high MFA values associated with compression wood [
46]. Thus, early selection for density alone could be counterproductive because of the potential to select families that are prone to producing compression wood. The positive relationship between density and MFA on the genetic level casts further doubt on the utility of density for early selection of mechanical properties. Hence, the combined measurement Vel² × Dens may present a way to simultaneously select for high stiffness and high density, to identify individuals with superior mechanical properties but screening out high density, low stiffness individuals.
Direct selection based on MFA presents another possible strategy for wood quality improvement. Low MFA—and therefore higher stiffness—is moderately related to height and DBH, especially on the genotypic level. At first glance, this may appear counterintuitive because faster growth is usually associated with lower strength and stiffness. But in this study on young white spruce, more vigorous genotypes, especially those with a superior height growth, are thought to form less compression wood because they are already better positioned in the competition for light [
46]. They probably also started to produce wood with a less juvenile character at breast height compared to slower growing individuals. Overall, MFA has good early selection efficiency [
33]. Selection for MFA could therefore be easily combined with growth improvement strategies. Nevertheless, these gains are offset by a comparably small expected genetic gain for MFA on the individual and family mean levels in this dataset. This is most likely due to the rather weak heritability and low variation of this trait, especially in the young trees where only the MFA of the last complete ring was measured.
The relationships between wood density and growth-related traits, such as DBH, height, ring width, as well as earlywood width and its proportion, are similar to earlier reports [
2,
8]. More vigorous trees tend to allocate their resources into increased radial increment and height growth, and produce wood of lower density [
47].
The negative correlation between growth rate and wood density is problematic in a tree breeding context. Density, being related to the mechanical traits MOE and MOR, has traditionally been used as an easily measurable trait in order to improve wood quality [
47]. Different authors have attempted to find ways around this problem, for example by considering individual density components or traits that influence wood density. Nevertheless, Ivkovich
et al. [
3] found no particular benefits of using wood anatomy to solve the problem.
Recent advances in clonal forestry mean that multiple copies of individuals or lines that do not follow the general correlation trend can be produced. Park
et al. [
48] identified several fast-growing white spruce clones that maintained above-average density. However, clonal methods are relatively costly, so may only be economically profitable for intensely managed plantations and less suitable for mass reforestation on extensively managed sites. Acoustic velocity presents an interesting alternative to wood density measurements as its correlations with diameter increment were weak. At the same time the relationship between height growth and velocity was positive. Although it was weak in this study, a positive trend between tree height and velocity has been already observed in Douglas-fir [
28] and slash pine [
34]. Future selection strategies may profit from acoustic velocity determination because it is more closely related to mechanical traits than wood density alone [
29]. This is also shown by insignificance of density in the phenotypic prediction models in the juvenile and the mature trees.
The low to moderate phenotypic relationship between acoustic velocity and MFA is in contrast to the results on the genetic level, which indicate that MFA could be successfully improved by selection for velocity. Environmental influences appear to have attenuated the genetic relationship between both traits. The low
R2 in the MFA models for young trees is probably also related to the fact that MFA measurements were made on a single growth ring, compared to the large variation in MFA and MOE
SS in the mature trees. The addition of other easily measurable traits could not considerably ameliorate model fit for juvenile MFA. In MFA models that included both velocity and diameter as predictors, diameter was the only significant variable for juvenile trees. A significant influence of DBH on adult tree MOE has been reported in previous studies on black spruce [
49,
50]. Slenderness (HD-ratio) was an important predictor for mechanical wood traits in this study. More slender stems tend to be stiffer and thus have lower MFA in order to support the momentum resulting from the crown. Alternatively, this could be related to the properties of the acoustic wave as it propagates within the stem, since, for a given stiffness, acoustic velocity can be mediated by the ratio between tree diameter and the distance between the probes [
50]. The significance of the HD-ratio indicates that this variable is complementary to acoustic velocity for description of mechanical wood quality. This also agrees with the low correlation between growth and velocity and further encourages the use of acoustic velocity besides growth traits in tree improvement.
In contrast to what may be expected from the theoretical relationship of acoustic velocity and mechanical wood traits (Equation 1) the square of velocity did not improve MFA and MOE models in this study. But this may be different when more heterogeneous samples from different ages are considered and the variation in both traits is consequently higher. Additionally, the difference in model fit between juvenile trees and adult trees suggest an evolution of the link between MFA and velocity on the phenotypic level with tree age. Age–age correlations should therefore be monitored more closely in future studies.
Earlier reports on the use of acoustic techniques on young trees using the same or similar tools were based on fast growing pine species with larger stem diameters [
34,
45,
51] than diameters of the young trees in this study. We thus needed to evaluate if acoustic velocity could be measured with a sufficient precision in order to make selection for wood quality in more slowly growing trees. We initially expected high degree of variation of acoustic velocity in young white spruce due to the presence of large numbers of branches. Defects or knots may prevent an optimal propagation of the sound wave especially in small diameter stems. Lindström
et al. [
51] mentioned that density variations may weaken velocity measurements and hinder the detection of good quality stems. But the results of this study indicate that acoustic velocity is a useful tool for genetic improvement of wood quality also in slow growing boreal species. Velocity measurements were apparently of good precision also in small trees which is in accordance to the results by Zhang
et al. [
52] who noted a more uniform propagation of sound in smaller stems of red pine.
The occurrence of compression wood in juvenile stems is often made responsible for weaker genetic parameters in the juvenile wood than in adult wood for example. Apiolaza
et al. [
53] separately analyzed compression wood and opposite wood from systematically leaning radiata pine trees. The authors concluded that their approach reduces the noise of random occurrence of compression and opposite wood in juvenile stems. Although this approach may be too costly for many tree improvement programs, it may help to more precisely separate environmental effects and facilitate the identification of superior or inferior trees in future generations. This could possibly improve phenotypic prediction models at the juvenile age. Nevertheless, several strong genetic relationships were presented in this study which indicates a good power of detection of genetic effects in our experimental layout. Additionally, low errors were associated to heritability and correlation estimates. The precision is probably due to the high number of trees per family chosen for this study following the findings by Perron
et al. [
54].










{kind=link}