
Effects of Nitrogen and Phosphorus Additions on the Stability of Soil Carbon Fractions in Subtropical Castanopsis sclerophylla Forests

Abstract
1. Introduction
2. Materials and Methods
2.1. Site Descriptions
2.2. Experimental Design and Treatments
2.2.1. Soil Sampling
2.2.2. Soil Physicochemical Properties and Carbon Fraction Measurements
2.3. Statistical Analysis
3. Results
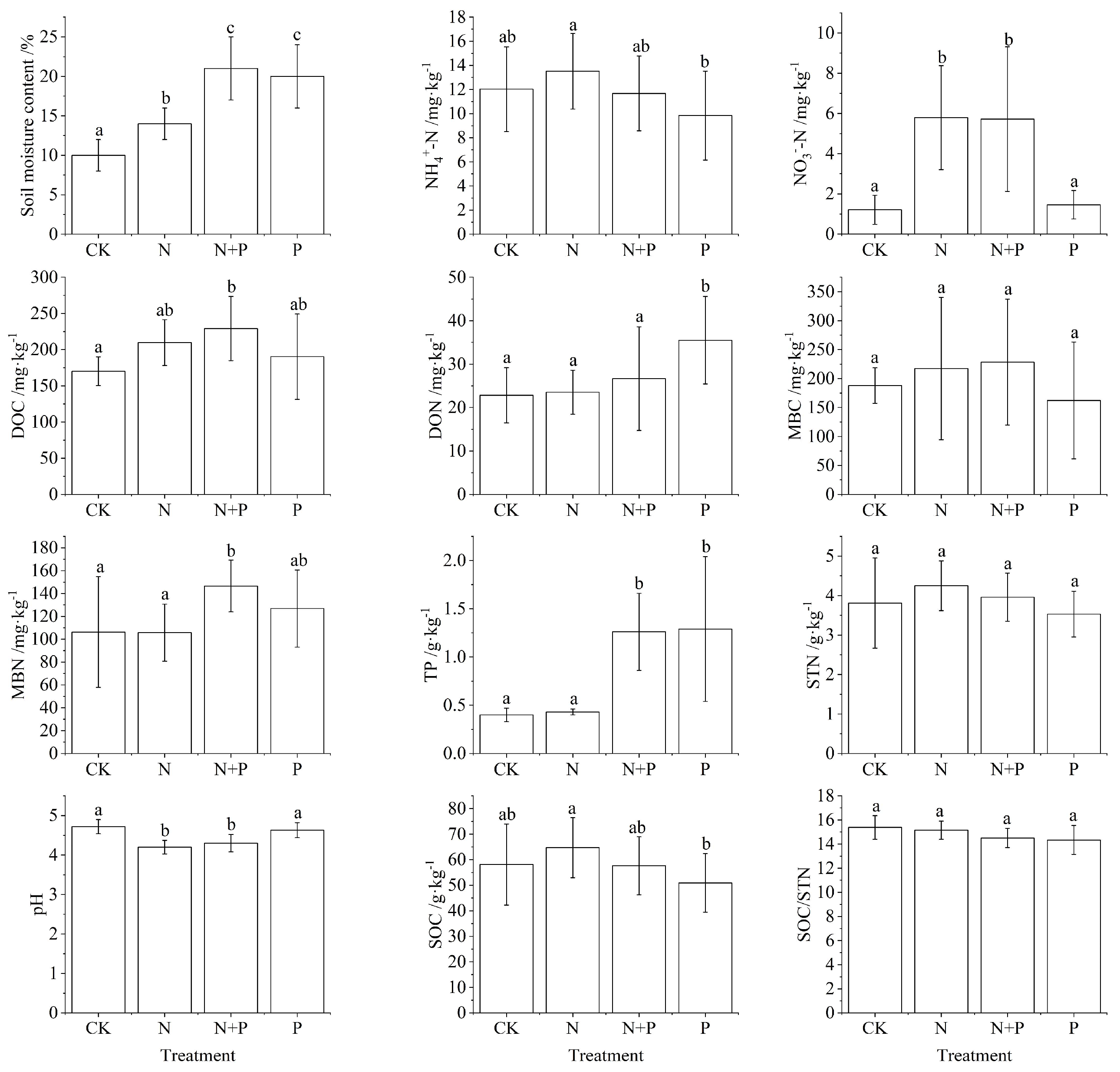
3.1. Changes in Soil Physical and Chemical Properties Under N and P Addition
3.2. Effect of N and P Addition on Different Active ROC
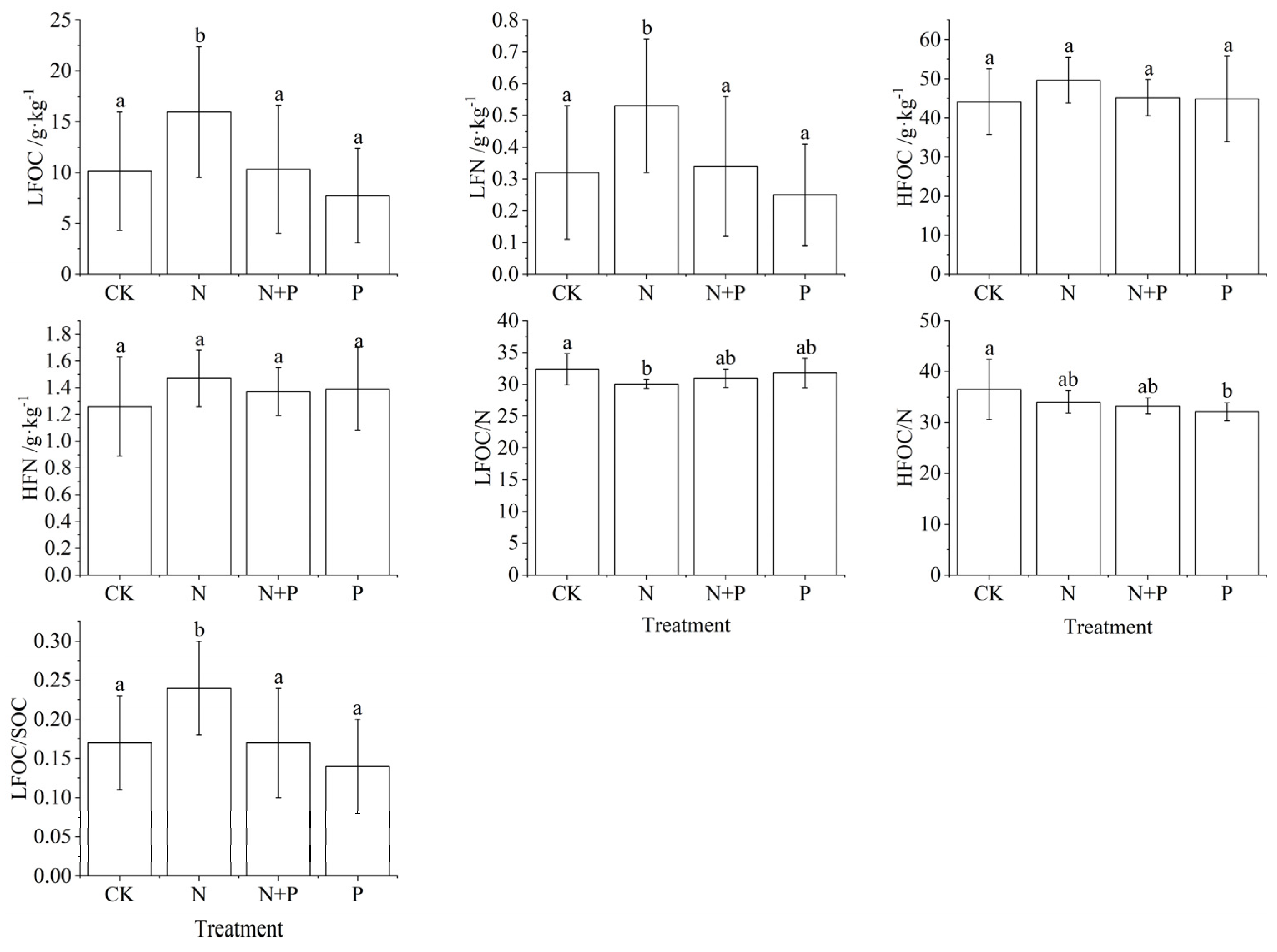
3.3. Effects of N and P Addition on Light and Heavy Fractions C and N
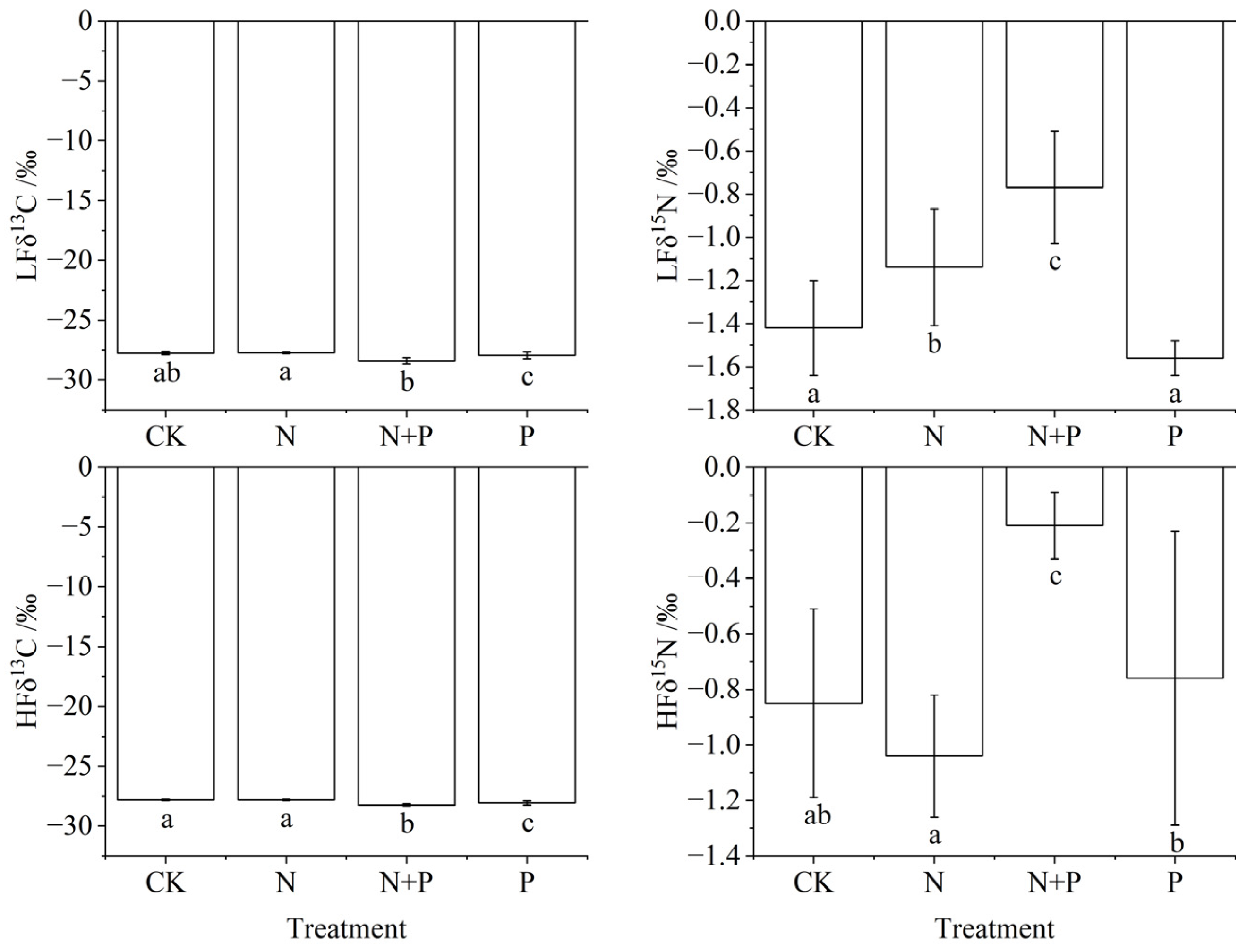
3.4. Effects of N and P Addition on C and N Stable Isotopes of Soil Density Fractions
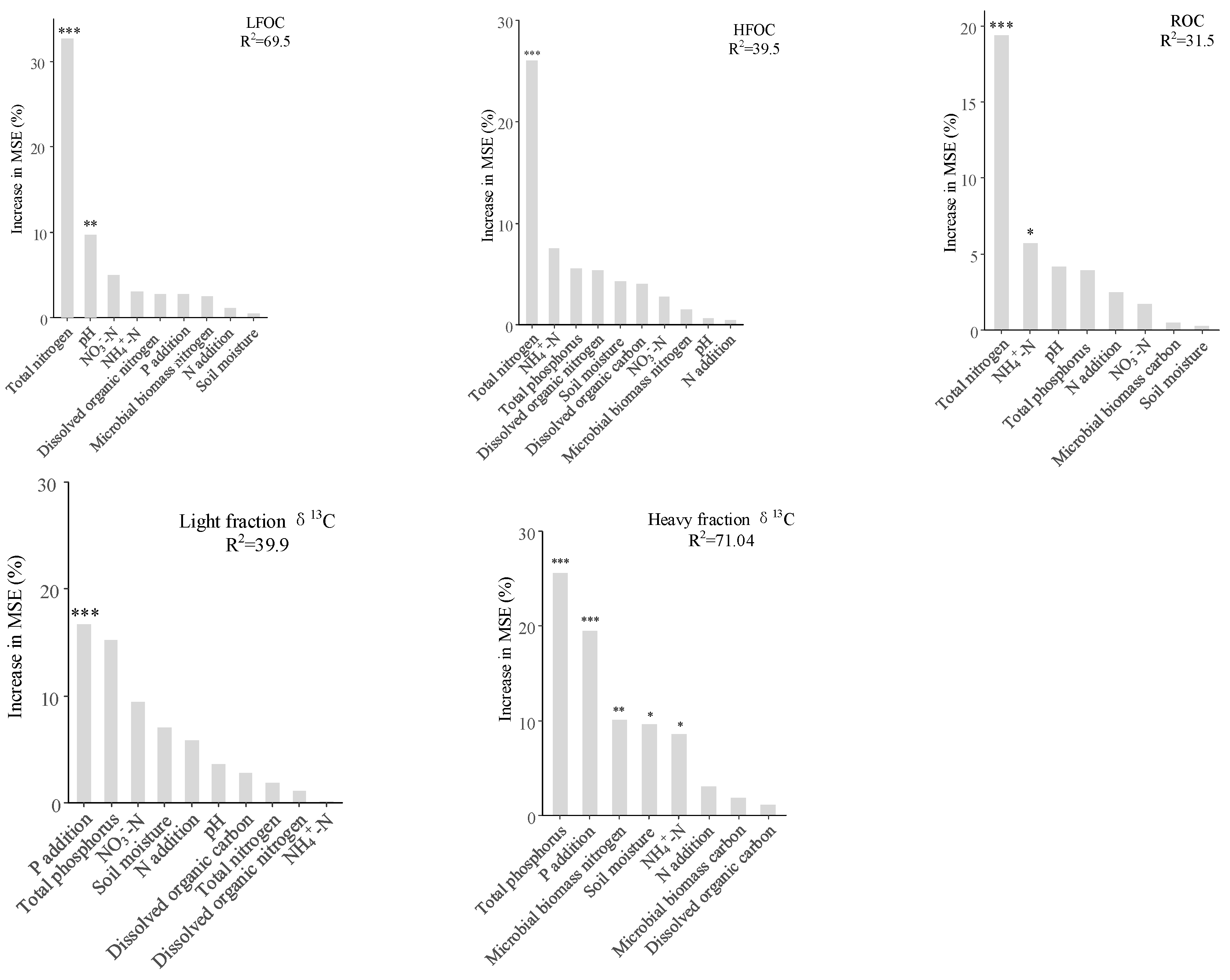
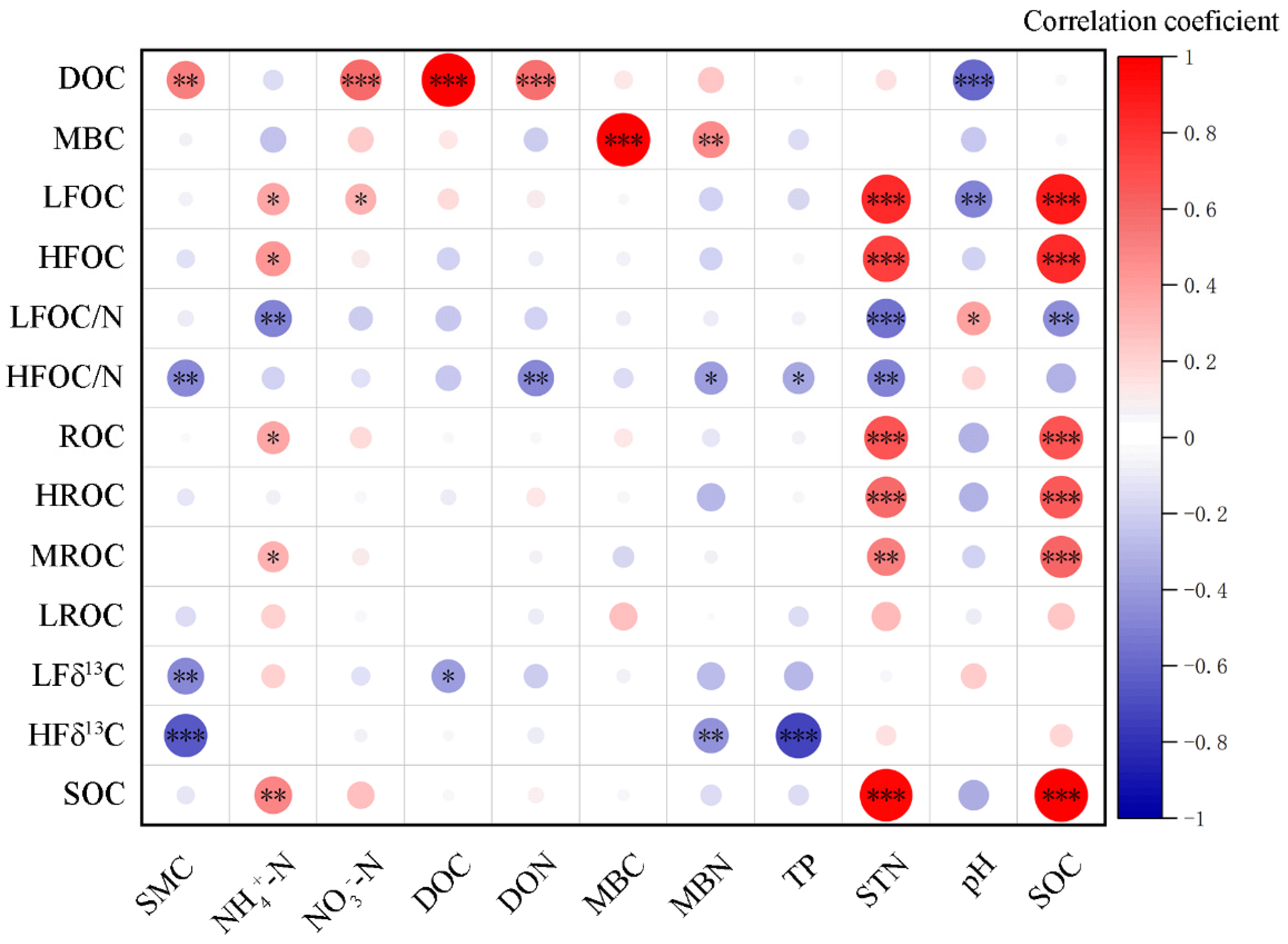
3.5. Correlation Between Environmental Factors and Soil C Fractions
4. Discussion
4.1. Response of SOC and Its Fractions to N and P Addition
4.2. Carbon and Nitrogen Isotope Response of Soil Light and Heavy Fractions to N and P Addition
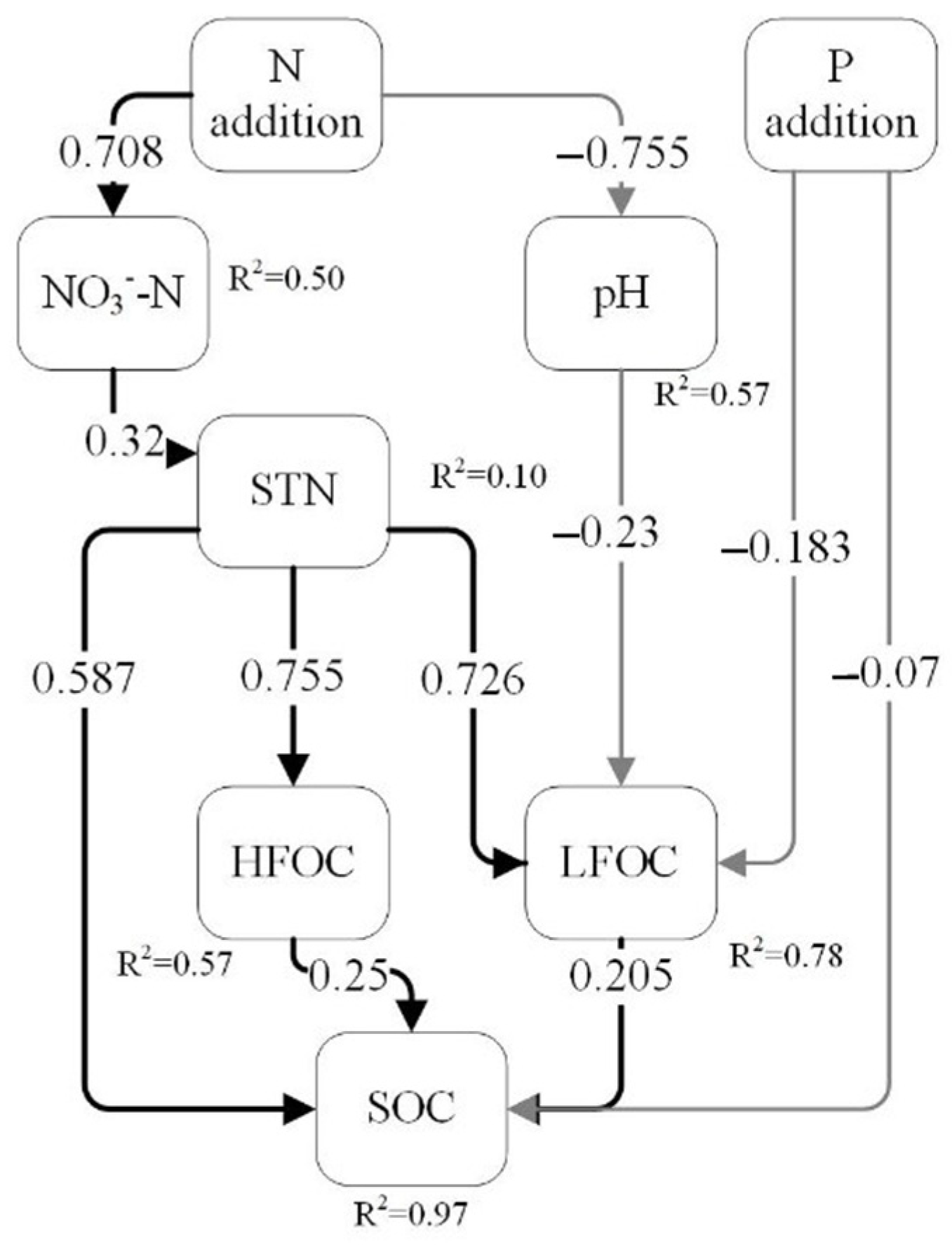
4.3. Factors Affecting Changes in SOC Fractions Under N and P Addition
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, X.L.; Zhao, X.; Bai, Y.F.; Tang, Z.Y.; Wang, W.T.; Zhao, Y.C.; Wan, H.W.; Xie, Z.Q.; Shi, X.Z.; Wu, B.F.; et al. Carbon pools in China’s terrestrial ecosystems: New estimates based on an intensive field survey. Proc. Natl. Acad. Sci. USA 2018, 115, 4021–4026. [Google Scholar] [CrossRef]
- Pan, B.X.; Deng, Z.Q.; Yu, Y.C.; Xu, X.N.; Bi, L.L.; Tao, X. Differential responses of soil respiration and its components to nitrogen application in urban forests. Forests 2022, 13, 2064. [Google Scholar] [CrossRef]
- Keenan, T.F.; Williams, C.A. The terrestrial carbon sink. Annu. Rev. Environ. Resour. 2018, 43, 219–243. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, C.C.; Deng, J.; Zhang, B.W.; Chen, F.S.; Chen, W.; Fang, X.M.; Li, J.J.; Zu, K.L.; Bu, W.S. Response of tree growth to nutrient addition is size dependent in a subtropical forest. Sci. Total Environ. 2024, 923, 171501. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.H.; Shi, Y.; Sun, W.J.; Chang, J.F.; Zhu, J.X.; Chen, L.Y.; Wang, X.; Guo, Y.P.; Zhang, H.T.; Yu, L.F.; et al. Terrestrial carbon sinks in China and around the world and their contribution to carbon neutrality. Sci. China Life Sci. 2022, 65, 861–895. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.R.; Wen, Z.Y.; Yin, R.Y.; Deng, P.F.; Gao, Y.; Xu, H.; Xu, X.N. Soil organic carbon turnover response to nitrogen and phosphorus additions in eastern China: Evidence from stable carbon isotopes. Forests 2023, 14, 1314. [Google Scholar] [CrossRef]
- Mendoza-Martinez, V.; Collins, S.L.; McLaren, J.R. Long-term fertilization increases soil but not plant or microbial N in a Chihuahuan Desert grassland. Biogeosciences 2024, 21, 2655–2667. [Google Scholar] [CrossRef]
- Penuelas, J.; Poulter, B.; Sardans, J.; Ciais, P.; van der Velde, M.; Bopp, L.; Boucher, O.; Godderis, Y.; Hinsinger, P.; Llusia, J.; et al. Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe. Nat. Commun. 2013, 4, 2934. [Google Scholar] [CrossRef]
- Gao, Y.; Jia, Y.L.; Yu, G.R.; He, N.P.; Zhang, L.; Zhu, B.; Wang, Y.F. Anthropogenic reactive nitrogen deposition and associated nutrient limitation effect on gross primary productivity in inland water of China. J. Clean. Prod. 2019, 208, 530–540. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013; p. 1535. [Google Scholar]
- Zhou, Z.H.; Ren, C.J.; Wang, C.K.; Delgado-Baquerizo, M.; Luo, Y.Q.; Luo, Z.K.; Du, Z.G.; Zhu, B.; Yang, Y.H.; Jiao, S.; et al. Global turnover of soil mineral-associated and particulate organic carbon. Nat. Commun. 2024, 15, 5329. [Google Scholar] [CrossRef]
- Riggs, C.E.; Hobbie, S.E.; Bach, E.M.; Hofmockel, K.S.; Kazanski, C.E. Nitrogen addition changes grassland soil organic matter decomposition. Biogeochemistry 2015, 125, 203–219. [Google Scholar] [CrossRef]
- Chen, H.; Li, D.; Feng, W.; Niu, S.; Plante, A.F.; Luo, Y.; Wang, K. Different responses of soil organic carbon fractions to nitrogen additions. Eur. J. Soil Sci. 2018, 69, 1098–1104. [Google Scholar] [CrossRef]
- Tan, W.B.; Wang, G.A.; Huang, C.H.; Gao, R.T.; Xi, B.D.; Zhu, B.A. Physico-chemical protection, rather than biochemical composition, governs the responses of soil organic carbon decomposition to nitrogen addition in a temperate agroecosystem. Sci. Total Environ. 2017, 598, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.J.; Zhu, W.X.; Zhou, Y.B.; Yin, Y. Soil organic carbon chemical functional groups under different revegetation types are coupled with changes in the microbial community composition and the functional genes. Forests 2019, 10, 240. [Google Scholar] [CrossRef]
- Fan, F.L.; Li, Z.J.; Wakelin, S.A.; Yu, W.T.; Liang, Y.C. Mineral fertilizer alters cellulolytic community structure and suppresses soil cellobiohydrolase activity in a long-term fertilization experiment. Soil Biol. Biochem. 2012, 55, 70–77. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Elliott, E.T. Methods for physical separation and characterization of soil organic matter fractions. Geoderma 1993, 56, 449–457. [Google Scholar] [CrossRef]
- Christensen, B.T. Physical fractionation of soil and structural and functional complexity in organic matter turnover. Eur. J. Soil Sci. 2010, 52, 345–353. [Google Scholar] [CrossRef]
- von Luetzow, M.; Koegel-Knabner, I.; Ekschmitt, K.; Flessa, H.; Guggenberger, G.; Matzner, E.; Marschner, B. SOM fractionation methods: Relevance to functional pools and to stabilization mechanisms. Soil Biol. Biochem. 2007, 39, 2183–2207. [Google Scholar] [CrossRef]
- Song, B.; Niu, S.L.; Li, L.H.; Zhang, L.X.; Yu, G.R. Soil carbon fractions in grasslands respond differently to various levels of nitrogen enrichments. Plant Soil 2014, 384, 401–412. [Google Scholar] [CrossRef]
- Mori, T.; Lu, X.K.; Aoyagi, R.; Mo, J.M. Reconsidering the phosphorus limitation of soil microbial activity in tropical forests. Funct. Ecol. 2018, 32, 1145–1154. [Google Scholar] [CrossRef]
- Neff, J.C.; Townsend, A.R.; Gleixner, G.; Lehman, S.J.; Turnbull, J.; Bowman, W.D. Variable effects of nitrogen additions on the stability and turnover of soil carbon. Nature 2002, 419, 915–917. [Google Scholar] [CrossRef]
- Liu, L.; Zhang, T.; Gilliam, F.S.; Gundersen, P.; Zhang, W.; Chen, H.; Mo, J.M. Interactive effects of nitrogen and phosphorus on soil microbial communities in a tropical forest. PLoS ONE 2013, 8, e61188. [Google Scholar] [CrossRef] [PubMed]
- Argiroff, W.A.; Zak, D.R.; Upchurch, R.A.; Salley, S.O.; Grandy, A.S. Anthropogenic N deposition alters soil organic matter biochemistry and microbial communities on decaying fine roots. Global Change Biol. 2019, 25, 4369–4382. [Google Scholar] [CrossRef]
- Wu, J.P.; Liu, W.F.; Zhang, W.X.; Shao, Y.H.; Duan, H.L.; Chen, B.D.; Wei, X.H.; Fan, H.B. Long-term nitrogen addition changes soil microbial community and litter decomposition rate in a subtropical forest. Appl. Soil Ecol. 2019, 142, 43–51. [Google Scholar] [CrossRef]
- Feng, J.G.; Zhang, Q.F.; Yuan, X.; Zhu, B. Effects of nitrogen and phosphorus addition on soil organic carbon: Review and prospects. Chin. J. Plant Ecol. 2022, 46, 855–870. [Google Scholar] [CrossRef]
- Wang, R.Z.; Penuelas, J.; Li, T.; Liu, H.Y.; Wu, H.; Zhang, Y.G.; Sardans, J.; Jiang, Y. Natural abundance of 13C and 15N provides evidence for plant-soil carbon and nitrogen dynamics in a N-fertilized meadow. Ecology 2021, 102, e03348. [Google Scholar] [CrossRef]
- Chen, Z.J.; Geng, S.C.; Zhang, J.H.; Setala, H.; Gu, Y.; Wang, F.; Zhang, X.; Wang, X.X.; Han, S.J. Addition of nitrogen enhances stability of soil organic matter in a temperate forest. Eur. J. Soil Sci. 2017, 68, 189–199. [Google Scholar] [CrossRef]
- Wu, J.; Joergensen, R.G.; Pommerening, B.; Chaussod, R.; Brookes, P.C. Measurement of soil microbial biomass C by fumigation-extraction—An automated procedure. Soil Biol. Biochem. 1990, 22, 1167–1169. [Google Scholar] [CrossRef]
- Blair, G.J.; Lefroy, R.D.B.; Lisle, L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Aust. J. Agr. Res. 1995, 46, 1459–1466. [Google Scholar] [CrossRef]
- Janzen, H.H.; Campbell, C.A.; Brandt, S.A.; Lafond, G.P.; Townleysmith, L. Light-fraction organic matter in soils from long-term crop rotations. Soil Sci. Soc. Am. J. 1992, 56, 1799–1806. [Google Scholar] [CrossRef]
- Grace, J. Structural Equation Modeling and Natural Systems; Academic Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Xu, C.H.; Xu, X.; Ju, C.H.; Chen, H.Y.H.; Wilsey, B.J.; Luo, Y.Q.; Fan, W. Long-term, amplified responses of soil organic carbon to nitrogen addition worldwide. Global Change Biol. 2021, 27, 1170–1180. [Google Scholar] [CrossRef]
- Ye, H.; Zhao, Y.; He, S.; Wu, Z.D.; Yue, M.; Hong, M. Metagenomics reveals the response of desert steppe microbial communities and carbon-nitrogen cycling functional genes to nitrogen deposition. Front. Microbiol. 2024, 15, 1369196. [Google Scholar] [CrossRef]
- Liu, W.X.; Qiao, C.L.; Yang, S.; Bai, W.M.; Liu, L.L. Microbial carbon use efficiency and priming effect regulate soil carbon storage under nitrogen deposition by slowing soil organic matter decomposition. Geoderma 2018, 332, 37–44. [Google Scholar] [CrossRef]
- Qiu, T.; Clark, J.S.; Kovach, K.R.; Townsend, P.A.; Swenson, J.J. Remotely sensed crown nutrient concentrations modulate forest reproduction across the contiguous United States. Ecology 2024, 105, e4366. [Google Scholar] [CrossRef] [PubMed]
- FOG, K. The effect of added nitrogen on the rate of decomposition of organic matter. Biol. Rev. 1988, 63, 433–462. [Google Scholar] [CrossRef]
- Janssens, I.A.; Dieleman, W.; Luyssaert, S.; Subke, J.A.; Reichstein, M.; Ceulemans, R.; Ciais, P.; Dolman, A.J.; Grace, J.; Matteucci, G.; et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat. Geosci. 2010, 3, 315–322. [Google Scholar] [CrossRef]
- Spohn, M.; Schleuss, P.M. Addition of inorganic phosphorus to soil leads to desorption of organic compounds and thus to increased soil respiration. Soil Biol. Biochem. 2019, 130, 220–226. [Google Scholar] [CrossRef]
- Spohn, M.; Diakova, K.; Aburto, F.; Doetterl, S.; Borovec, J. Sorption and desorption of organic matter in soils as affected by phosphate. Geoderma 2022, 405, 115377. [Google Scholar] [CrossRef]
- Johnston, E.R.; Kim, M.; Hatt, J.K.; Phillips, J.R.; Yao, Q.M.; Song, Y.; Hazen, T.C.; Mayes, M.A.; Konstantinidis, K.T. Phosphate addition increases tropical forest soil respiration primarily by deconstraining microbial population growth. Soil Biol. Biochem. 2019, 130, 43–54. [Google Scholar] [CrossRef]
- Hui, D.F.; Porter, W.; Phillips, J.R.; Aidar, M.P.M.; Lebreux, S.J.; Schadt, C.W.; Mayes, M.A. Phosphorus rather than nitrogen enhances CO2 emissions in tropical forest soils: Evidence from a laboratory incubation study. Eur. J. Soil Sci. 2020, 71, 495–510. [Google Scholar] [CrossRef]
- Wang, Z.; Chlus, A.; Geygan, R.; Ye, Z.; Zheng, T.; Singh, A.; Couture, J.J.; Cavender-Bares, J.; Kruger, E.L.; Townsend, P.A. Foliar functional traits from imaging spectroscopy across biomes in eastern North America. New Phytol. 2020, 228, 494–511. [Google Scholar] [CrossRef] [PubMed]
- Grandy, A.S.; Neff, J.C. Molecular C dynamics downstream: The biochemical decomposition sequence and its impact on soil organic matter structure and function. Sci. Total Environ. 2008, 404, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.G.; Xiao, W.; Zheng, C.Y.; Zhu, B. Nitrogen addition has contrasting effects on particulate and mineral-associated soil organic carbon in a subtropical forest. Soil Biol. Biochem. 2020, 142, 107708. [Google Scholar] [CrossRef]
- Jian, J.N.; Liu, W.C.; Zhu, Y.F.; Li, J.X.; Wen, Y.H.; Liu, F.H.; Ren, C.J.; Han, X.H. Effects of Short-term Nitrogen Addition on Soil Organic Carbon Components in Robinia pseudoacacia L. Plantation. Huanjing Kexue 2023, 44, 2767–2774. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, C.; Peng, B.; Xia, Z.W.; Jiang, P.; Bai, E. Effect of nitrogen addition on the variations in the natural abundance of nitrogen isotopes of plant and soil components. Plant Soil 2017, 412, 453–464. [Google Scholar] [CrossRef]
- Amundson, R.; Austin, A.T.; Schuur, E.A.G.; Yoo, K.; Matzek, V.; Kendall, C.; Uebersax, A.; Brenner, D.; Baisden, W.T. Global patterns of the isotopic composition of soil and plant nitrogen. Glob. Biogeochem. Cycles 2003, 17, 1031. [Google Scholar] [CrossRef]
- Brookshire, E.N.J.; Hedin, L.O.; Newbold, J.D.; Sigman, D.M.; Jackson, J.K. Sustained losses of bioavailable nitrogen from montane tropical forests. Nat. Geosci. 2012, 5, 123–126. [Google Scholar] [CrossRef]
- Wang, C.; Wang, X.B.; Liu, D.W.; Wu, H.H.; Lu, X.T.; Fang, Y.T.; Cheng, W.X.; Luo, W.T.; Jiang, P.; Shi, J.S.; et al. Aridity threshold in controlling ecosystem nitrogen cycling in arid and semi-arid grasslands. Nat. Commun. 2014, 5, 4799. [Google Scholar] [CrossRef]
- Wang, R.Z.; Dijkstra, F.A.; Liu, H.Y.; Yin, J.F.; Wang, X.; Feng, X.; Xu, Z.W.; Jiang, Y. Response of soil carbon to nitrogen and water addition differs between labile and recalcitrant fractions: Evidence from multi-year data and different soil depths in a semi-arid steppe. Catena 2019, 172, 857–865. [Google Scholar] [CrossRef]
- Jilling, A.; Keiluweit, M.; Gutknecht, J.L.M.; Grandy, A.S. Priming mechanisms providing plants and microbes access to mineral-associated organic matter. Soil Biol. Biochem. 2021, 158, 108265. [Google Scholar] [CrossRef]
- Keiluweit, M.; Bougoure, J.J.; Nico, P.S.; Pett-Ridge, J.; Weber, P.K.; Kleber, M. Mineral protection of soil carbon counteracted by root exudates. Nat. Clim. Change 2015, 5, 588–595. [Google Scholar] [CrossRef]
- Zeng, Z.Q.; Wang, S.L.; Zhang, C.M.; Tang, H.; Li, X.Q.; Wu, Z.J.; Luo, J. Soil microbial activity and nutrients of evergreen broad-leaf forests in mid-subtropical region of China. J. Forestry Res. 2015, 26, 673–678. [Google Scholar] [CrossRef]
- Niu, S.; Yang, H.; Zhang, Z.; Wu, M.; Lu, Q.; Li, L.; Han, X.; Wan, S. Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe. Ecosystems 2009, 12, 915–926. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | N Addition | P Addition | N + P | |||
|---|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | |
| DOC | 10.17 | <0.01 ** | 0.94 | 0.35 | 13.22 | <0.01 ** |
| MBC | 0.49 | 0.50 | 0.53 | 0.48 | 1.16 | 0.30 |
| LFOC | 4.64 | 0.04 * | 4.28 | 0.04 * | 0.69 | 0.41 |
| LFN | 4.93 | 0.03 * | 4.05 | 0.05 | 0.76 | 0.39 |
| HFOC | 1.26 | 0.27 | 0.5 | 0.49 | 0.1 | 0.32 |
| HFN | 0.91 | 0.35 | 0.04 | 0.85 | 1.61 | 0.21 |
| LFOC/N | 6.44 | 0.02 * | 0.05 | 0.83 | 1.33 | 0.26 |
| HFOC/N | 0.34 | 0.56 | 5.3 | 0.03 * | 2.53 | 0.12 |
| LFOC/SOC | 6.42 | 0.02 * | 0.67 | 0.43 | 0.01 | 0.96 |
| ROC | 5.09 | 0.03 * | 0 | 0.98 | 0 | 0.99 |
| HROC | 0.07 | 0.79 | 1.42 | 0.24 | 4.69 | 0.04 * |
| MROC | 1.43 | 0.24 | 0.05 | 0.82 | 0.19 | 0.67 |
| LROC | 2.39 | 0.13 | 0 | 0.99 | 1.21 | 0.28 |
| ROC/SOC | 0.34 | 0.57 | 1.63 | 0.22 | 4.46 | 0.04 * |
| LFδ13C | 8.91 | <0.01 ** | 38.22 | <0.01 ** | 12.96 | <0.01 ** |
| LFδ15N | 51.27 | <0.01 ** | 2.66 | 0.11 | 11.77 | <0.01 ** |
| HFδ13C | 5.62 | 0.02 * | 80.37 | <0.01 ** | 5.99 | 0.02 * |
| HFδ15N | 5.14 | 0.03 * | 33.04 | <0.01 ** | 22.09 | <0.01 ** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, Y.; Xu, X.; Cuong, L. Effects of Nitrogen and Phosphorus Additions on the Stability of Soil Carbon Fractions in Subtropical Castanopsis sclerophylla Forests. Forests 2025, 16, 1264. https://doi.org/10.3390/f16081264
Dai Y, Xu X, Cuong L. Effects of Nitrogen and Phosphorus Additions on the Stability of Soil Carbon Fractions in Subtropical Castanopsis sclerophylla Forests. Forests. 2025; 16(8):1264. https://doi.org/10.3390/f16081264
Chicago/Turabian StyleDai, Yunze, Xiaoniu Xu, and LeVan Cuong. 2025. "Effects of Nitrogen and Phosphorus Additions on the Stability of Soil Carbon Fractions in Subtropical Castanopsis sclerophylla Forests" Forests 16, no. 8: 1264. https://doi.org/10.3390/f16081264
APA StyleDai, Y., Xu, X., & Cuong, L. (2025). Effects of Nitrogen and Phosphorus Additions on the Stability of Soil Carbon Fractions in Subtropical Castanopsis sclerophylla Forests. Forests, 16(8), 1264. https://doi.org/10.3390/f16081264