

Structural Diversity and Biodiversity of Forest and Hedgerow in Areas Managed for Pheasant Shooting Across the UK
 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Systematic Evidence Evaluation
- What is the existing evidence on the impact of pheasant shooting management on the quality and quantity of woodland and hedgerow habitats in the UK?
- How do management practices, such as pheasant release pens, affect woodland ecology, including biodiversity and habitat composition?
- What woodland and hedgerow management activities can improve or enhance ecological impacts on areas used for pheasant shooting?
2.2. Spatial Analysis Approach
2.3. Selection of Sites Managed for Pheasant Shooting and Comparison Sites
2.4. Satellite Remote Sensing of Outcome Variables
2.5. Biodiversity Data and Rarefaction
- Administrative area = United Kingdom;
- Year = 2010–2024;
- Basis of record = human observation OR preserved specimen;
- Taxon rank = species;
- Scientific name = class Aves;
- Scientific name = kingdom Plantae;
- Scientific name = family Hesperiidae OR Papilionidae OR Pieridae OR Lycaenidae OR Riodinidae OR Nymphalidae.
- Plants (URL: https://doi.org/10.15468/dl.eh653t);
- Birds (URL: https://doi.org/10.15468/dl.px4x77);
- Butterflies (URL: https://doi.org/10.15468/dl.dq98b4).
2.6. Testing for Differences in Outcome Variables Between Pheasant and Comparison Sites
3. Results
3.1. Location of Studies in the Project
3.2. Systematic Review of Literature
3.2.1. Literature Assessed and Selected
3.2.2. Site Characteristics
3.2.3. Impact of Sites Managed for Pheasant Shooting on Biodiversity
3.2.4. Quality of the Evidence Base
3.3. Remote Sensing
3.3.1. Performance of the Matching Procedure
3.3.2. Habitat Quality
3.3.3. Biodiversity (Birds, Plants, and Butterflies)
3.3.4. Differences Between Nations Within Great Britain
3.3.5. Effect of Size of Polygon Area
4. Discussion
4.1. Extent and Composition of Forest, Margins, and Hedgerows
4.2. Knowledge Gaps and Future Research Directions
4.3. Limitations
4.3.1. Systematic Evidence Evaluation
4.3.2. Remote Sensing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AONB | Area of Outstanding Natural Beauty |
| BASC | British Association for Shooting and Conservation |
| BACI | Before–After Control Intervention |
| BTO | British Trust for Ornithology |
| EIA | Environmental Impact Assessment |
| EU | European Union |
| GBIF | Global Biodiversity Information Facility |
| IUCN | International Union for Conservation of Nature |
| JNCC | Joint Nature Conservation Committee |
| LD | Linear Dichroism |
| MPA | Marine-Protected Area |
| NGO | Non-Governmental Organisation |
| NVC | National Vegetation Classification |
| RSPB | Royal Society for the Protection of Birds |
| SAC | Special Area of Conservation |
| SPA | Special Protection Area |
| SSSI | Site of Special Scientific Interest |
| UK | United Kingdom |
| WWF | World Wide Fund for Nature |
Appendix A
- What is the existing evidence on the impact of pheasant shooting management on the quality and quantity of woodland and hedgerow habitats in the UK?
- How do management practices, such as pheasant release pens, affect woodland ecology, including biodiversity and habitat composition?
- What woodland and hedgerow management activities can improve or enhance ecological impacts on areas used for pheasant shooting?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Description |
|---|---|
| Population | Estates/land holdings with woodlands in the UK. |
| Intervention | Pheasant shooting on the estate. |
| Comparator | Non-pheasant shooting site. |
| Outcome | Ecological impacts on habitat quality (e.g., biodiversity, species richness, and habitat structure) and habitat quantity (e.g., extent of woodland and hedgerows). |
| Category | Inclusion | Exclusion |
|---|---|---|
| Population: Woodland and Hedgerow Habitats in the UK | Studies focussing on ecological characteristics (e.g., biodiversity, structural composition, and extent) of woodland and hedgerow habitats in the UK managed specifically for game shooting activities (specifically pheasant). | Studies focussing on non-UK habitats or areas where game shooting is not a central land management practice. Studies examining agricultural landscapes without substantial woodland or hedgerow components. |
| Intervention: Land Management Practices Associated with Game Shooting | Studies investigating specific land management practices for game shooting, such as pheasant release pens, woodland management for game species, or shooting-related interventions affecting woodland and hedgerow habitats. | Studies examining unrelated land management activities, such as purely agricultural practices or recreational land uses without a connection to shooting. |
| Comparator: Sites with Different Management Practices | Studies including comparisons with control sites where game shooting management is not conducted or where alternative land uses (e.g., conservation-only and agricultural without shooting) are implemented. | Studies without any form of comparator, including purely descriptive accounts of game shooting sites without comparison to non-shooting or differently managed sites. |
| Outcome: Ecological Impacts on Habitat Quality and Quantity | Studies assessing the impact of game shooting management on habitat quality (e.g., species richness, species abundance, and community composition) or habitat quantity (e.g., extent of woodland or hedgerows) in the UK. | Studies focussing exclusively on economic, social, or non-ecological outcomes. Studies lacking an evaluation of woodland or hedgerow impacts on habitat quality. |
| Study Type: Qualitative and Empirical Studies | Qualitative and empirical studies, including peer-reviewed articles, reports, and grey literature (e.g., government reports and NGO publications) evaluating the impact of game shooting on woodland and hedgerow habitats. | Modelling studies using third party data, opinion pieces, or anecdotal accounts without full methodological descriptions. |
Appendix B
| Article Number | Citation | Number of Studies |
|---|---|---|
| 1 | Robertson, P. A. and Woodburn, M. I. A. and Hill, D. A., 1988. The Effects Of Woodland Management For Pheasants On The Abundance Of Butterflies In Dorset, England. BIOLOGICAL CONSERVATION. 45(3), 159–167. 10.1016/0006-3207(88)90136-X | 1 |
| 2 | Sage, R. B. and Ludolf, C. and Robertson, P. A., 2005. The ground flora of ancient semi-natural woodlands in pheasant release pens in England. BIOLOGICAL CONSERVATION. 122(2), 243–252. 10.1016/j.biocon.2004.07.014 | 1 |
| 3 | Draycott, R. A. H. and Hoodless, A. N. and Sage, R. B., 2008. Effects of pheasant management on vegetation and birds in lowland woodlands. JOURNAL OF APPLIED ECOLOGY. 45(1), 334–341. 10.1111/j.1365-2664.2007.01379.x | 2 |
| 4 | Draycott, R. A. H. and Hoodless, A. N. and Cooke, M. and Sage, R. B., 2012. The influence of pheasant releasing and associated management on farmland hedgerows and birds in England. EUROPEAN JOURNAL OF WILDLIFE RESEARCH. 58(1), 227–234. 10.1007/s10344-011-0568-0 | 2 |
| 5 | Neumann, J. L. and Holloway, G. J. and Sage, R. B. and Hoodless, A. N., 2015. Releasing of pheasants for shooting in the UK alters woodland invertebrate communities. BIOLOGICAL CONSERVATION. 191, 50–59. 10.1016/j.biocon.2015.06.022 | 2 |
| 6 | Capstick, L. A. and Sage, R. B. and Hoodless, A., 2019. Ground flora recovery in disused pheasant pens is limited and affected by pheasant release density. BIOLOGICAL CONSERVATION. 231, 181–188. 10.1016/j.biocon.2018.12.020 | 1 |
| 7 | Madden, J. R. and Buckley, R. and Ratcliffe, S., 2023. Large-scale correlations between gamebird release and management and animal biodiversity metrics in lowland Great Britain. ECOLOGY AND EVOLUTION. 13(5). 10.1002/ece3.10059 | 1 |
| 8 | Capstick, L., Draycott, R., Wheelwright, C., Ling, D., Sage, R. & Hoodless, A, 2019. The effect of game management on the conservation value of woodland rides. Forest Ecology and Management. | 1 |
| 9 | Greenall, T., 2007. Management of gamebird shooting in lowland Britain: Social attitudes, biodiversity benefits and willingness-to-pay. PhD Thesis, University of Kent., 296 pp. https://kar.kent.ac.uk/86403/ | 3 |
| 10 | Hall, A., Sage, R. A., & Madden, J. R., 2021. The effects of released pheasants on invertebrate populations in and around woodland release sites. Ecology and Evolution. 11, 13559–13569. | 1 |
| 11 | Hoodless, A. N. & Draycott, K., 2006. Effects of pheasant management at woodland edges. The Game Conservancy Trust Review. 37, 30–31 | 1 |
| 12 | Hoodless, A. N., Lewis, R.,& Palmer, J., 2006. Songbird use of pheasant woods in winter. The Game Conservancy Trust Review. 37, 28–29 | 1 |
| 13 | Hoodless, A., & Draycott, R., 2007. Pheasant releasing and woodland rides. The Game Conservancy Trust Review. 38, 16–17 | 1 |
| 14 | Stoate, C., 2002. Multifunctional use of a natural resource on farmland: wild pheasant (Phasianus colchicus) management and the conservation of farmland passerines. Biodiversity and Conservation. 11, 561–573 | 1 |
| 15 | Pressland, C.L., 2009. The impact of releasing pheasants for shooting on invertebrates in british woodlands | 1 |
| 16 | Sage, R.B., 2017. Impacts of pheasant releasing for shooting on habitats and wildlife on the south Exmoor estates. Report, Game & Wildlife Conservation Trust | 1 |
| 17 | Sage, R. B. and Putaala, A. and Pradell-Ruiz, V. and Greenall, T. L. and Woodburn, M. I. A. and Draycott, R. A. H., 2003. Incubation success of released hand-reared pheasants Phasianus colchicus compared with wild ones. Wildlife Biology. 9(3), 179–184 | 1 |
| 18 | Sage, R. B. and Woodburn, M. I. A. and Draycott, R. A. H. and Hoodless, A. N. and Clarke, S., 2009. The flora and structure of farmland hedges and hedgebanks near to pheasant release pens compared with other hedges. BIOLOGICAL CONSERVATION. 142(7), 1362–1369. 10.1016/j.biocon.2009.01.034 | 1 |
| 19 | Aebischer, N. J. and Bailey, C. M. and Gibbons, D. W. and Morris, A. J. and Peach, W. J. and Stoate, C., 2016. Twenty years of local farmland bird conservation: the effects of management on avian abundance at two UK demonstration sites. BIRD STUDY. 63(1), 10–30. 10.1080/00063657.2015.1090391 | 1 |
| 20 | Sánchez-García, C. and Buner, F. D. and Aebischer, N. J., 2015. Supplementary winter food for gamebirds through feeders: Which species actually benefit?. JOURNAL OF WILDLIFE MANAGEMENT. 79(5), 832–845. 10.1002/jwmg.889 | 4 |
| 21 | Sage, R. and Woodburn, M. and McCready, S. and Coomes, J., 2024. Winter game crop plots for gamebirds retain hedgerow breeding songbirds in an improved grassland landscape. WILDLIFE BIOLOGY. 2024(3). 10.1002/wlb3.01156 | 1 |
| 22 | Blake, D., 1996. What effects do releasing pheasants have on the ground flora of woodland rides?. WPA News. 50, 11–14 | 1 |
| 23 | Swan, G. J., Bearhop, S., Redpath, S. M., Silk, M. J., Padfield, D., Goodwin, C. E., & McDonald, R. A., 2022. Associations between abundances of free-roaming gamebirds and common buzzards Buteo buteo are not driven by consumption of gamebirds in the buzzard breeding season. Ecology and Evolution. 12, e8877. https://doi.org/10.1002/ece3.8877 | 1 |
| Species Studied | Common Name | Taxonomic Group | Status | Increased Presence | Decreased Presence | No Difference | Total | Article Number |
|---|---|---|---|---|---|---|---|---|
| Abax parallelepipedus | Parallel-Bordered Harp Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Acupalpus meridianus | Midday Harp Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Agonum marginatum | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Amara aenea | Common Sun Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Amara similata | Sun Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Asaphidion curtum | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Asaphidion flavipes | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Badister bipustulatus | Two-Spotted Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Bembidion harpaloides | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Bembidion lampros | Shiny Riverbank Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Bembidion obtusum | Paris Riverbank Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Calathus fuscipes | Dark-Footed Harp Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Calathus melanocephalus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Calathus rotundicollis | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Carabus arvensis | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Carabus monilis | Necklace Ground Beetle | Aboveground Invertebrates | Priority Species | 0 | 1 | 1 | 2 | 5 |
| Carabus nemoralis | European Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Carabus problematicus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Carabus violaceus | Violet Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Clivina fossor | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Cychrus caraboides | Snail Hunter Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Dromius agilis | Agile Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Harpalus latus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Harpalus rufipes | Strawberry Seed Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Leistus ferrugineus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Leistus fulvibarbis | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Leistus rufomarginatus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Leistus spinibarbis | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Leistus terminatus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Loricera pilicornis | Springtail Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Nebria brevicollis | Common Heart-Shield | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Nebria salina | Bare Footed Heart Shield | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Notiophilus biguttatus | Big-Eyed Bronze Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Patrobus atrorufus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pieris napi | Green-veined White | Aboveground Invertebrates | Non-Priority | 2 | 0 | 0 | 2 | 1, 9 |
| Platynus assimilis | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus cupreus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus diligens | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus macer | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus madidus | Blackclock Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus melanarius | Common Black Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus minor | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus niger | Large Black Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus nigrita | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus oblongopunctatus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Pterostichus strenuus | Rough-Chested Blackclock | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Stomis pumicatus | Longjaw Ground-Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Synuchus vivalis | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Trechus obtusus | London Riverbank Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Trechus quadristriatus | Ground Beetle | Aboveground Invertebrates | Non-Priority | 0 | 1 | 1 | 2 | 5 |
| Bombus lapidaries | Red-tailed Bumblebee | Aboveground Invertebrates | Non-Priority | 0 | 0 | 1 | 1 | 9 |
| Bombus lucorum | White-tailed Bumblebee | Aboveground Invertebrates | Non-Priority | 0 | 0 | 1 | 1 | 9 |
| Bombus pascuorum | Common Carder Bumblebee | Aboveground Invertebrates | Non-Priority | 0 | 0 | 1 | 1 | 9 |
| Bombus terrestris | Buff-tailed Bumblebee | Aboveground Invertebrates | Non-Priority | 0 | 0 | 1 | 1 | 9 |
| Maniola jurtina | Meadow Brown | Aboveground Invertebrates | Non-Priority | 1 | 0 | 0 | 1 | 9 |
| Pieris brassicae | Large White | Aboveground Invertebrates | Non-Priority | 1 | 0 | 0 | 1 | 9 |
| Pieris rapae | Small White | Aboveground Invertebrates | Non-Priority | 1 | 0 | 0 | 1 | 9 |
| Pica pica | Eurasian Magpie | Birds | Least Concern | 0 | 4 | 2 | 6 | 4, 7, 14 |
| Columba palumbus | Common Wood Pigeon | Birds | Bird-Amber | 3 | 0 | 2 | 5 | 3, 4, 12 |
| Phylloscopus trochilus | Willow Warbler | Birds | Bird-Amber | 3 | 0 | 2 | 5 | 3, 4, 14 |
| Regulus regulus | Goldcrest | Birds | Least Concern | 3 | 2 | 0 | 5 | 3, 4, 12 |
| Sylvia atricapilla | Blackcap | Birds | Least Concern | 3 | 0 | 2 | 5 | 3, 4, 14 |
| Sylvia communis | Common Whitethroat | Birds | Least Concern | 3 | 0 | 2 | 5 | 3, 4, 14 |
| Erithacus rubecula | European Robin | Birds | Least Concern | 1 | 0 | 3 | 4 | 4, 12, 14 |
| Fringilla coelebs | Chaffinch | Birds | Least Concern | 1 | 0 | 3 | 4 | 4, 12, 14 |
| Prunella modularis | Dunnock | Birds | Bird-Amber | 2 | 0 | 2 | 4 | 4, 12, 14 |
| Troglodytes troglodytes | Eurasian Wren | Birds | Bird-Amber | 1 | 1 | 2 | 4 | 4, 12, 14 |
| Carduelis cannabina | Common Linnet | Birds | Least Concern | 1 | 0 | 2 | 3 | 4, 14 |
| Carduelis carduelis | European Goldfinch | Birds | Least Concern | 1 | 0 | 2 | 3 | 4, 14 |
| Carduelis chloris | European Greenfinch | Birds | Least Concern | 1 | 0 | 2 | 3 | 4, 14 |
| Corvus corone | Carrion Crow | Birds | Least Concern | 0 | 3 | 0 | 3 | 4, 7 |
| Parus caeruleus | Eurasian Blue Tit | Birds | Least Concern | 1 | 0 | 2 | 3 | 4, 14 |
| Parus major | Great Tit | Birds | Least Concern | 0 | 1 | 2 | 3 | 4, 14 |
| Sylvia borin | Garden Warbler | Birds | Least Concern | 2 | 1 | 0 | 3 | 3, 14 |
| Turdus merula | Common Blackbird | Birds | Least Concern | 1 | 0 | 2 | 3 | 4, 14 |
| Aegithalos caudatus | Long-tailed Tit | Birds | Least Concern | 0 | 0 | 2 | 2 | 4 |
| Anthus trivialis | Tree Pipit | Birds | Bird-Red | 2 | 0 | 0 | 2 | 4 |
| Athene noctua | Little Owl | Birds | Least Concern | 2 | 0 | 0 | 2 | 4 |
| Corvus frugilegus | Rook | Birds | Bird-Amber | 2 | 0 | 0 | 2 | 4 |
| Dendrocopos major | Great Spotted Woodpecker | Birds | Least Concern | 0 | 2 | 0 | 2 | 4 |
| Emberiza citronella | Yellowhammer | Birds | Least Concern | 0 | 0 | 2 | 2 | 4 |
| Falco tinnunculus | Common Kestrel | Birds | Bird-Amber | 0 | 2 | 0 | 2 | 4 |
| Garrulus glandarius | Eurasian Jay | Birds | Least Concern | 2 | 0 | 0 | 2 | 4 |
| Hirundo rustica | Barn Swallow | Birds | Least Concern | 0 | 2 | 0 | 2 | 4 |
| Motacilla flava | Western Yellow Wagtail | Birds | Bird-Red | 2 | 0 | 0 | 2 | 4 |
| Perdix perdix | Grey Partridge | Birds | Bird-Red | 2 | 0 | 0 | 2 | 4 |
| Phylloscopus collybita | Common Chiffchaff | Birds | Least Concern | 2 | 0 | 0 | 2 | 3 |
| Picus viridis | European Green Woodpecker | Birds | Least Concern | 0 | 0 | 2 | 2 | 4 |
| Poecile palustris | Marsh Tit | Birds | Bird-Red | 2 | 0 | 0 | 2 | 4 |
| Pyrrhula pyrrhula | Eurasian Bullfinch | Birds | Bird-Amber | 0 | 0 | 2 | 2 | 4 |
| Sylvia curruca | Lesser Whitethroat | Birds | Least Concern | 2 | 0 | 0 | 2 | 4 |
| Turdus philomelus | Song Thrush | Birds | Least Concern | 2 | 0 | 0 | 2 | 4 |
| Alauda arvensis | Skylark | Birds | Bird-Red | 0 | 1 | 0 | 1 | 14 |
| Buteo buteo | Common Buzzard | Birds | Least Concern | 0 | 1 | 0 | 1 | 7 |
| Corvus monedula | Western Jackdaw | Birds | Least Concern | 1 | 0 | 0 | 1 | 14 |
| Emberiza citrinella | Yellowhammer | Birds | Bird-Red | 0 | 0 | 1 | 1 | 14 |
| Regulus ignicapilla | Firecrest | Birds | Least Concern | 1 | 0 | 0 | 1 | 12 |
| Sitta europaea | Eurasian Nuthatch | Birds | Least Concern | 1 | 0 | 0 | 1 | 12 |
| Sciurus carolinensis | Eastern Grey Squirrel | Mammals | Non-Priority | 1 | 0 | 0 | 1 | 7 |
| Vulpes vulpes | Red Fox | Mammals | Non-Priority | 0 | 1 | 0 | 1 | 7 |
| Hedera helix | Common Ivy | Vascular Plants | Non-Priority | 0 | 1 | 0 | 1 | 2 |
| Hyacinthoides non-scripta | English Bluebell | Vascular Plants | Non-Priority | 0 | 1 | 0 | 1 | 6 |
| Lamium galeobdolon | Yellow Archangel | Vascular Plants | Non-Priority | 0 | 1 | 0 | 1 | 6 |
| Mercurialis perennis | Dog’s Mercury | Vascular Plants | Non-Priority | 0 | 1 | 0 | 1 | 2 |
| Rubus fruticosus | Blackberry | Vascular Plants | Non-Priority | 0 | 1 | 0 | 1 | 2 |
| Rumex obtusifolius | Broad-leaved Dock | Vascular Plants | Non-Priority | 1 | 0 | 0 | 1 | 13 |
| Tripleurospermum inodorum | Scentless Mayweed | Vascular Plants | Non-Priority | 1 | 0 | 0 | 1 | 13 |
| Urtica dioica | Stinging Nettle | Vascular Plants | Non-Priority | 1 | 0 | 0 | 1 | 13 |
Appendix C
| Variable | Data Format; Units | Spatial Resolution | Years | Source | Processing |
|---|---|---|---|---|---|
| Pheasant sites | polygon shapefile | NA | 2000–2024 | British Association for Shooting and Conservation (BASC) personal communication I Danby | Removed duplicates. |
| Cadastral polygons | geodatabase | NA | 2024 | Land registry of England and Wales https://use-land-property-data.service.gov.uk/datasets/inspire/download (accessed on 30 May 2025) Land Register of Scotland https://www.ros.gov.uk/our-registers/land-register-of-scotland (accessed on 30 May 2025) | Converted to shapefiles. Masking of urban areas. Zonal geometry. Zonal mean of matching covariates. Selection of comparison sites. |
| Built-up areas | polygon shapefile | NA | 2024 | Ordnance Survey https://www.ordnancesurvey.co.uk/products/os-open-built-up-areas (accessed on 30 May 2025). | Used to identify rural cadastral polygons. |
| Regions | polygon shapefile | NA | 2023 | European Commission NUTS https://ec.europa.eu/eurostat/web/nuts (accessed on 30 May 2025) | Used to allocate pheasant sites and potential comparison sites to regions in UK. |
| Forest | raster (0/1) | 30 m | 2023 | Global Forest Watch https://storage.googleapis.com/earthenginepartners-hansen/GFC-2023-v1.11/download.html (accessed on 30 May 2025) | Zonal mean (proportion forest) in pheasant and potential comparison sites used as a covariate in matching. Zonal mean of ln area (ha) of contiguous forest cover in pheasant and comparison sites used as an outcome variable. |
| Elevation | raster (m) | 30 m | 2000 | Shuttle Radar Topography Mission (SRTM) https://www.earthdata.nasa.gov/data/instruments/srtm (accessed on 30 May 2025) | Used as a covariate in matching. |
| Spectral diversity | raster (reflectance) | 30 m | 2023 | Landsat surface reflectance https://www.usgs.gov/landsat-missions/landsat-collection-2-surface-reflectance (accessed on 30 May 2025) | Calculated spectral diversity using centre vs. neighbour differences in 3 × 3 kernel for 6 wavebands. Final spectral diversity is RMS across wavebands. Zonal mean within pheasant and comparison polygons. |
| Structural diversity | raster (dB) | 100 m | 2023 | Sentinel 1 C-band sigma 0 https://documentation.dataspace.copernicus.eu/Data/SentinelMissions/Sentinel1.html (accessed on 30 May 2025) | Calculated structural diversity using centre vs. neighbour differences in C-band sigma 0 in a 3 × 3 kernel. Zonal mean within pheasant and comparison polygons. |
| Hedgerow density | line shapefile | NA | 2016 | Woody linear features framework, Great Britain, v.1.0. NERC Environmental Information Data Centre https://doi.org/10.5285/d7da6cb9-104b-4dbc-b709-c1f7ba94fb16 (accessed on 30 May 2025) | Calculated length (m) of hedgerow in each pheasant and comparison polygon. Calculated hedgerow density (m/ha) using polygon area. |
| Biodiversity records | point records | NA | 2000–2023 | Global Biodiversity Information Facility (GBIF) Plants (URL: https://doi.org/10.15468/dl.eh653t) Birds (URL: https://doi.org/10.15468/dl.px4x77) Butterflies (URL: https://doi.org/10.15468/dl.dq98b4) | Counted number of distinct species in each taxonomic group in each pheasant and comparison site. Found area of each pheasant and comparison site. Calculated species density of each taxonomic group in each pheasant and comparison site. |
References
- Mason, L.R.; Bicknell, J.E.; Smart, J.; Peach, W.J. The Impacts of Non-Native Gamebird Release in the UK: An Updated Evidence Review; Report 66; RSPB Centre Conserv Sci: Sandy, UK, 2020; Available online: https://www.researchgate.net/publication/345717757_The_impacts_of_non-native_gamebird_release_in_the_UK_an_updated_evidence_review (accessed on 30 May 2025).
- Sage, R.B.; Hoodless, A.N.; Woodburn, M.I.; Draycott, R.A.; Madden, J.R.; Sotherton, N.W. Summary review and synthesis: Effects on habitats and wildlife of the release and management of pheasants and red-legged partridges on UK lowland shoots. Wildl. Biol. 2020, 2020, 1–12. [Google Scholar] [CrossRef]
- Mustin, K.; Arroyo, B.; Beja, P.; Newey, S.; Irivine, R.J.; Kestler, J.; Redpath, S.M. Consequences of game bird management for non-game species in Europe. J. Appl. Ecol. 2018, 55, 2285–2295. [Google Scholar] [CrossRef]
- Bicknell, J.; Smart, J.; Hoccom, D.; Amar, A.; Evans, A.; Walton, P.; Knott, J.; Lodge, T. Impacts of Non-Native Gamebird Release in the UK: A Review; Royal Society for the Protection of Birds: Bedfordshire, UK, 2010. [Google Scholar]
- Madden, J.R.; Sage, R.B. Ecological Consequences of Gamebird Releasing and Management on Lowland Shoots in England. Natural England Report (NEER016). 2020. Available online: https://publications.naturalengland.org.uk/publication/5078605686374400 (accessed on 30 May 2025).
- Madden, J.R.; Buckley, R.; Ratcliffe, S. Large-scale correlations between gamebird release and management and animal biodiversity metrics in lowland Great Britain. Ecol. Evol. 2023, 13, e10059. [Google Scholar] [CrossRef] [PubMed]
- Pullin, A.S.; Cheng, S.H.; Cooke, S.J.; Haddaway, N.R.; Macura, B.; Mckinnon, M.C.; Taylor, J.J. Informing conservation decisions through evidence synthesis and communication. In Conservation Research, Policy and Practice; Cambridge University Press: Cambridge, UK, 2020; pp. 114–128. [Google Scholar]
- Konno, K.; Pullin, A.S. Assessing the risk of bias in choice of search sources for environmental meta-analyses. Res. Synth. Methods 2020, 11, 698–713. [Google Scholar] [CrossRef] [PubMed]
- Haddaway, N.R.; Bernes, C.; Jonsson, B.G.; Hedlund, K. The benefits of systematic mapping to evidence-based environmental management. Ambio 2016, 45, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.; Coughlin, D.; Miller, J.; Kirk, S. The Production of Quick Scoping Reviews and Rapid Evidence Assessments: A How to Guide. 2015. Available online: https://nora.nerc.ac.uk/id/eprint/512448/ (accessed on 30 May 2025).
- Gribbons, B.; Herman, J. True and Quasi-Experimental Designs. Pract. Assess. Res. Eval. 1996, 5, 14. [Google Scholar] [CrossRef]
- Timmermans, J.; Kissling, W.D. Advancing terrestrial biodiversity monitoring with satellite remote sensing in the context of the Kunming-Montreal global biodiversity framework. Ecol. Indic. 2023, 154, 110773. [Google Scholar] [CrossRef]
- Farella, M.M.; Fisher, J.B.; Jiao, W.; Key, K.B.; Barnes, M.L. Thermal remote sensing for plant ecology from leaf to globe. J. Ecol. 2022, 110, 1996–2014. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Schnieder, F.D.; Santos, M.J.; Armstrong, A.; Carnaval, A.; Dahlin, K.M.; Fatoyinbo, L.; Hurt, G.C.; Schimel, D.; Townsend, P.A.; et al. Integrating remote sensing with ecology and evolution to advance biodiversity conservation. Nat. Ecol. Evol. 2022, 6, 506–519. [Google Scholar] [CrossRef] [PubMed]
- Schweiger, A.K.; Laliberte, E. Plant beta-diversity across biomes captured by imaging spectroscopy. Nat. Commun. 2022, 13, 2767. [Google Scholar] [CrossRef] [PubMed]
- Lines, E.R.; Fischer, F.J.; Foord Owen, H.J.; Jucker, T. The shape of trees: Reimagining forest ecology in three dimensions with remote sensing. J. Ecol. 2022, 110, 1730–1745. [Google Scholar] [CrossRef]
- Potapov, P.; Li, X.; Hernandez-Serna, A.; Tyukavina, A.; Hansen, M.; Kommareddy, A.; Pickens, A.; Turubanova, A.; Tang, H.; Silva, C.E.; et al. Mapping global forest canopy height through integration of GEDI and Landsat data. Remote Sens. Environ. 2020, 253, 112165. [Google Scholar] [CrossRef]
- BASC. Green Shoots Mapping. 2025. Available online: https://basc.org.uk/conservation-in-action/green-shoots-mapping/ (accessed on 30 May 2025).
- Office for National Statistics. Administrative Geographies. 2025. Available online: https://www.google.com/url?q=https://www.ons.gov.uk/methodology/geography/ukgeographies/administrativegeography&sa=D&source=docs&ust=1748598873027847&usg=AOvVaw3a_Ai2BSGPWAOWIpHGtv05 (accessed on 30 May 2025).
- HM Land Registry. Index Polygons Spatial Data (INSPIRE). 2021. Available online: https://use-land-property-data.service.gov.uk/datasets/inspire/download (accessed on 30 May 2025).
- Registers of Scotland. Land Register of Scotland. 2025. Available online: https://www.ros.gov.uk/our-registers/land-register-of-scotland (accessed on 30 May 2025).
- Ordnance Survey. OS Open Built up Areas. 2025. Available online: https://www.ordnancesurvey.co.uk/products/os-open-built-up-areas (accessed on 30 May 2025).
- Hansen, M.C.; Potapov, P.V.; Moore, R.; Hancher, M.; Turubanova, S.A.; Tyukavina, A.; Thau, D.; Stehman, S.V.; Goetz, S.J.; Loveland, T.R.; et al. High-Resolution Global Maps of 21st-Century Forest Cover Change. Science 2013, 342, 850–853. [Google Scholar] [CrossRef] [PubMed]
- Farr, T.G.; Rosen, P.A.; Caro, E.; Crippen, R.; Duren, R.; Hensley, S.; Kobrick, M.; Paller, M.; Rodriguez, E.; Roth, L.; et al. The Shuttle Radar Topography Mission. Rev. Geophys. 2007, 45, RG2004. [Google Scholar] [CrossRef]
- Ho, D.; Imai, K.; King, G.; Stuart, E.A. MatchIt: Nonparametric preprocessing for parametric causal inference. J. Stat. Softw. 2011, 42, 1–28. [Google Scholar] [CrossRef]
- Santoro, M.; Beer, C.; Cartus, O.; Schmullius, C.; Shvidenko, A.; McCallum, I.; Wegmüller, U.; Wiesmann, A. Retrieval of growing stock volume in boreal forest using hyper-temporal series of Envisat ASAR ScanSAR backscatter measurements. Remote Sens. Environ. 2011, 115, 490–507. [Google Scholar] [CrossRef]
- Scholefield, P.A.; Morton, R.D.; Rowland, C.S.; Henrys, P.A.; Howard, D.C.; Norton, L.R. Woody Linear Features Framework; Great Britain v. 1.0; NERC Environmental Information Data Centre (Dataset): Polaris House, UK, 2016. [Google Scholar]
- Oksanen, J.; Simpson, G.L.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Solymos, P.; Stevens, M.H.H.; Szoecs, E.; et al. Vegan: Community Ecology Package; Version 2.6-4; CRAN: Stamford, CT, USA, 2022. [Google Scholar]
- Macarthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967; Available online: http://www.jstor.org/stable/j.ctt19cc1t2 (accessed on 30 May 2025).
- Rosenzweig, M.L. Species Diversity in Space and Time; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Long, P.R.; Benz, D.; Macias-Fauria, M.; Seddon, A.W.R.; Holland, P.W.A.; Martin, A.C.; Hagemann, R.; Frost, T.K.; Simpson, A.C.; Power, D.J.; et al. LEFT—A web-based tool for the remote measurement and estimation of ecological value across global landscapes. Methods Ecol. Evol. 2017, 9, 571–579. [Google Scholar] [CrossRef]
- Clifford, P.; Richardson, S.; Hemon, D. Assessing the significance of the correlation between two spatial processes. Biometrics 1989, 45, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Dutilleul, P. Modifying the t test for assessing the correlation between two spatial processes. Biometrics 1993, 49, 305–314. [Google Scholar] [CrossRef]
- Vallejos, R.; Osorio, F.; Bevilacqua, M. Spatial Relationships Between Two Georeferenced Variables: With Applications in R; Springer: Berlin/Heidelberg, Germany, 2020. [Google Scholar]
- Draycott, R.A.H.; Hoodless, A.N.; Cooke, M.; Sage, R.B. The influence of pheasant releasing and associated management on farmland hedgerows and birds in England. Eur. J. Wildl. Res. 2012, 58, 227–234. [Google Scholar] [CrossRef]
- Hoodless, A.; Draycott, R. Pheasant releasing and woodland rides. Game Conserv. Trust. Rev. 2007, 38, 16–17. [Google Scholar]
- Stoate, C. Multifunctional use of a natural resource on farmland: Wild pheasant (Phasianus colchicus) management and the conservation of farmland passerines. Biodivers. Conserv. 2002, 11, 561–573. [Google Scholar] [CrossRef]
- Sage, R.B.S. Impacts of Pheasant Releasing for Shooting on Habitats and Wildlife on the South Exmoor Estates; Game & Wildlife Conservation Trust Report; Game & Wildlife Conservation Trust: Hampshire, UK, 2018. [Google Scholar]
- Robertson, P.A.; Woodburn, M.I.A.; Hill, D.A. The Effects of Woodland Management for Pheasants on the Abundance of Butterflies in Dorset, England. Biol. Conserv. 1988, 45, 159–167. [Google Scholar] [CrossRef]
- Draycott, R.A.H.; Hoodless, A.N.; Sage, R.B. Effects of pheasant management on vegetation and birds in lowland woodlands. J. Appl. Ecol. 2008, 45, 334–341. [Google Scholar] [CrossRef]
- Hoodless, A.N.; Lewis, R.; Palmer, J. Songbird use of pheasant woods in winter. Game Conserv. Trust. Rev. 2006, 37, 28–29. [Google Scholar]
- Hoodless, A.N.; Draycott, K. Effects of pheasant management at woodland edges. Game Conserv. Trust. Rev. 2006, 37, 30–31. [Google Scholar]
- Greenall, T. Management of Gamebird Shooting in Lowland Britain: Social Attitudes, Biodiversity Benefits and Willingness-to-Pay. Ph.D. Thesis, University of Kent, Canterbury, UK, 2007. Available online: https://kar.kent.ac.uk/86403/ (accessed on 30 May 2025).
- Hall, A.; Sage, R.A.; Madden, J.R. The effects of released pheasants on invertebrate populations in and around woodland release sites. Ecol. Evol. 2021, 11, 13559–13569. [Google Scholar] [CrossRef] [PubMed]
- Sage, R.B.; Ludolf, C.; Robertson, P.A. The ground flora of ancient semi-natural woodlands in pheasant release pens in England. Biol. Conserv. 2005, 122, 243–252. [Google Scholar] [CrossRef]
- Neumann, J.L.; Holloway, G.J.; Sage, R.B.; Hoodless, A.N. Releasing of pheasants for shooting in the UK alters woodland invertebrate communities. Biol. Conserv. 2015, 191, 50–59. [Google Scholar] [CrossRef]
- Capstick, L.A.; Sage, R.B.; Hoodless, A. Ground flora recovery in disused pheasant pens is limited and affected by pheasant release density. Biol. Conserv. 2019, 231, 181–188. [Google Scholar] [CrossRef]
- Capstick, L.; Draycott, R.; Wheelwright, C.; Ling, D.; Sage, R.; Hoodless, A. The effect of game management on the conservation value of woodland rides. For. Ecol. Manag. 2019, 454, 117242. [Google Scholar] [CrossRef]
- Pressland, C.L. The Impact of Releasing Pheasants for Shooting on Invertebrates in British Woodlands. Ph.D. Thesis, University of Bristol, Bristol, UK, 2009. [Google Scholar]
- Woodburn, M.I.A.; Sage, R.B. Effect of pheasant releasing on edge habitats. Game Conserv. Trust. Rev. 2004, 36, 36–37. [Google Scholar]
- Livoreil, B.; Glanville, J.; Haddaway, N.R.; Bayliss, H.; Bethel, A.; de Lachapelle, F.F.; Robalino, S.; Savilaakso, S.; Zhou, W.; Petrokofsky, G.; et al. Systematic searching for environmental evidence using multiple tools and sources. Environ. Evid. 2017, 6, 23. [Google Scholar] [CrossRef]
- Päivinen, R.; Petrokofsky, G.; Harvey, W.J.; Petrokofsky, L.; Puttonen, P.; Kangas, J.; Mikkola, E.; Byholm, L.; Käär, L. State of forest research in 2010s—A bibliographic study with special reference to Finland, Sweden and Austria. Scand. J. For. Res. 2023, 38, 23–38. [Google Scholar] [CrossRef]
- Altman, D.G. Mathematics for kappa. Pract. Stat. Med. Res. 1991, 1991, 406–407. [Google Scholar]
- Frampton, G.K.; Livoreil, B.; Petrokofsky, G. Eligibility screening in evidence synthesis of environmental management topics. Environ. Evid. 2017, 6, 27. [Google Scholar] [CrossRef]
- Stanbury, A.; Eaton, M.; Aebischer, N.; Balmer, D.; Brown, A.; Douse, A.; Lindley, P.; McCulloch, N.; Noble, D.; Win, I. The status of our bird populations: The fifth Birds of Conservation Concern in the United Kingdom, Channel Islands and Isle of Man and second IUCN Red List assessment of extinction risk for Great Britain. Br. Birds 2021, 114, 723–747. [Google Scholar]
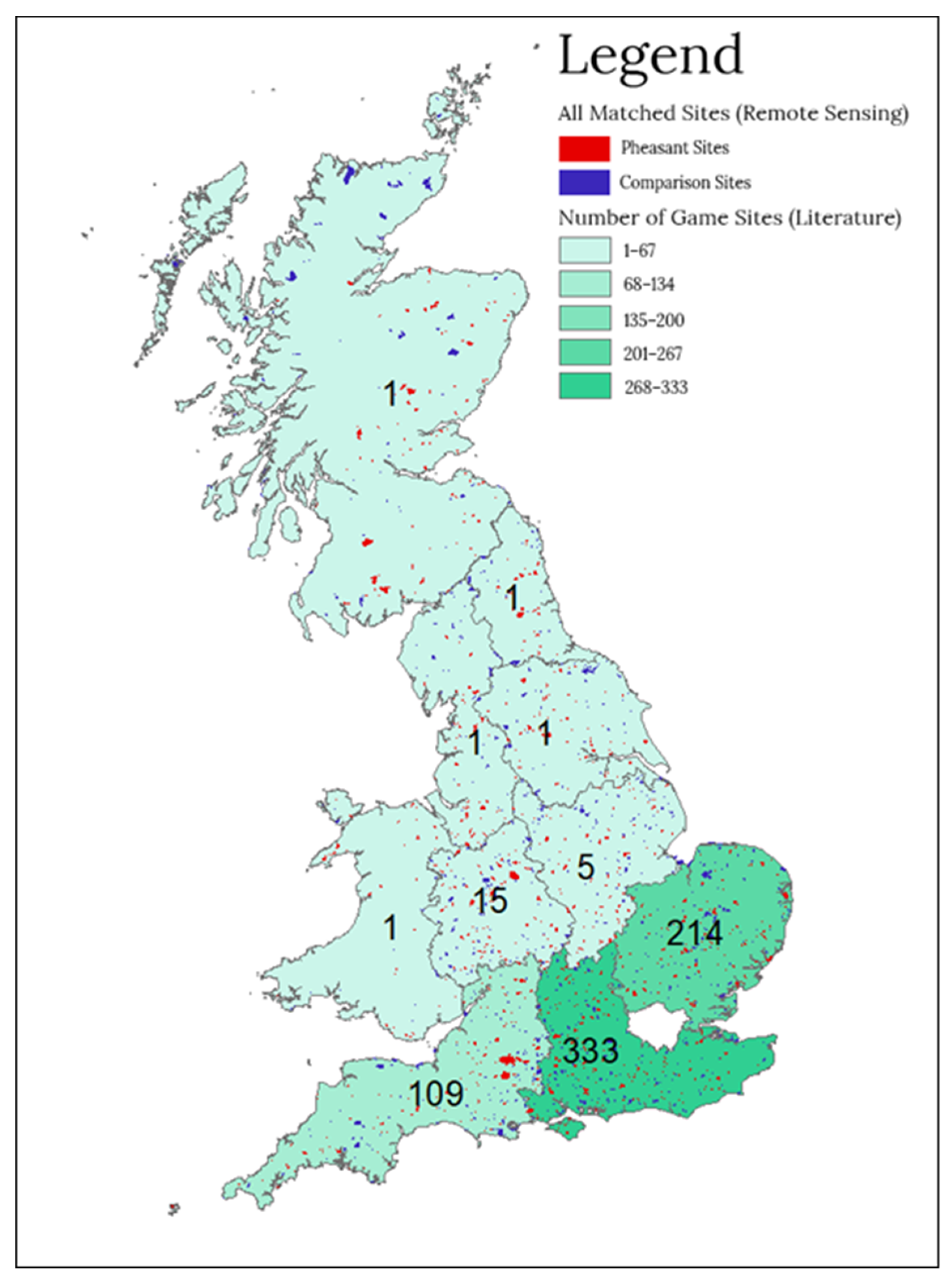
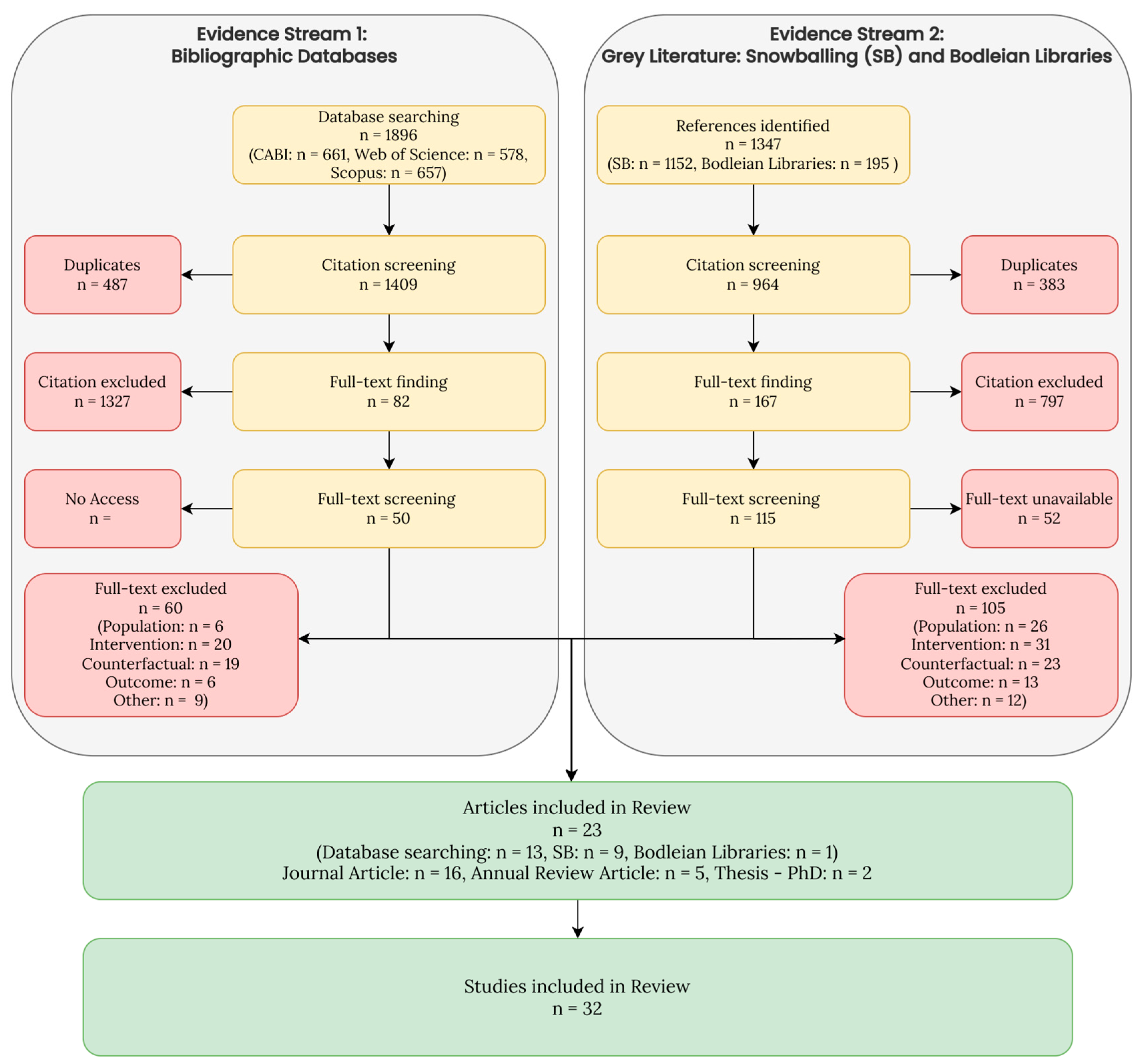
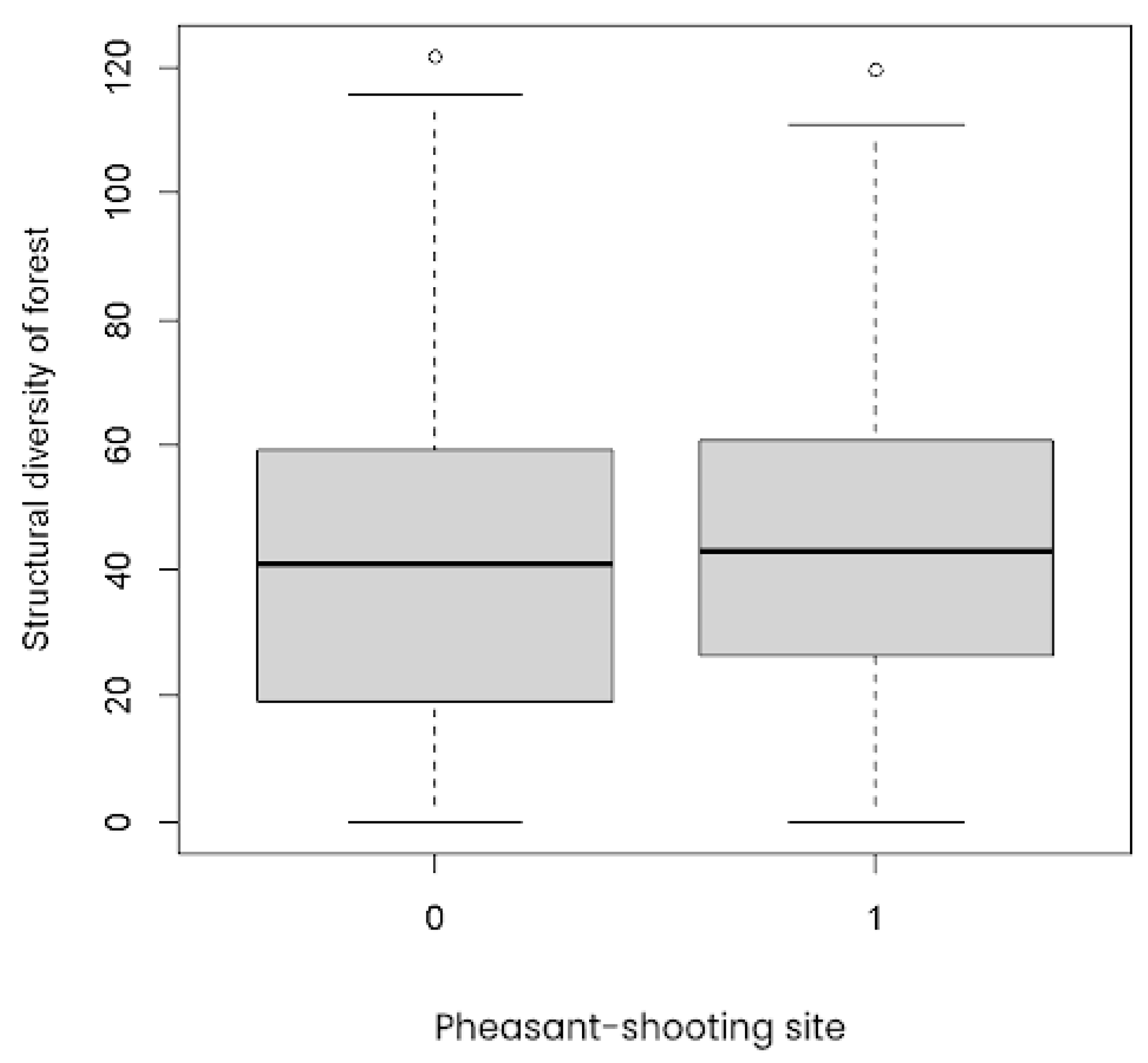
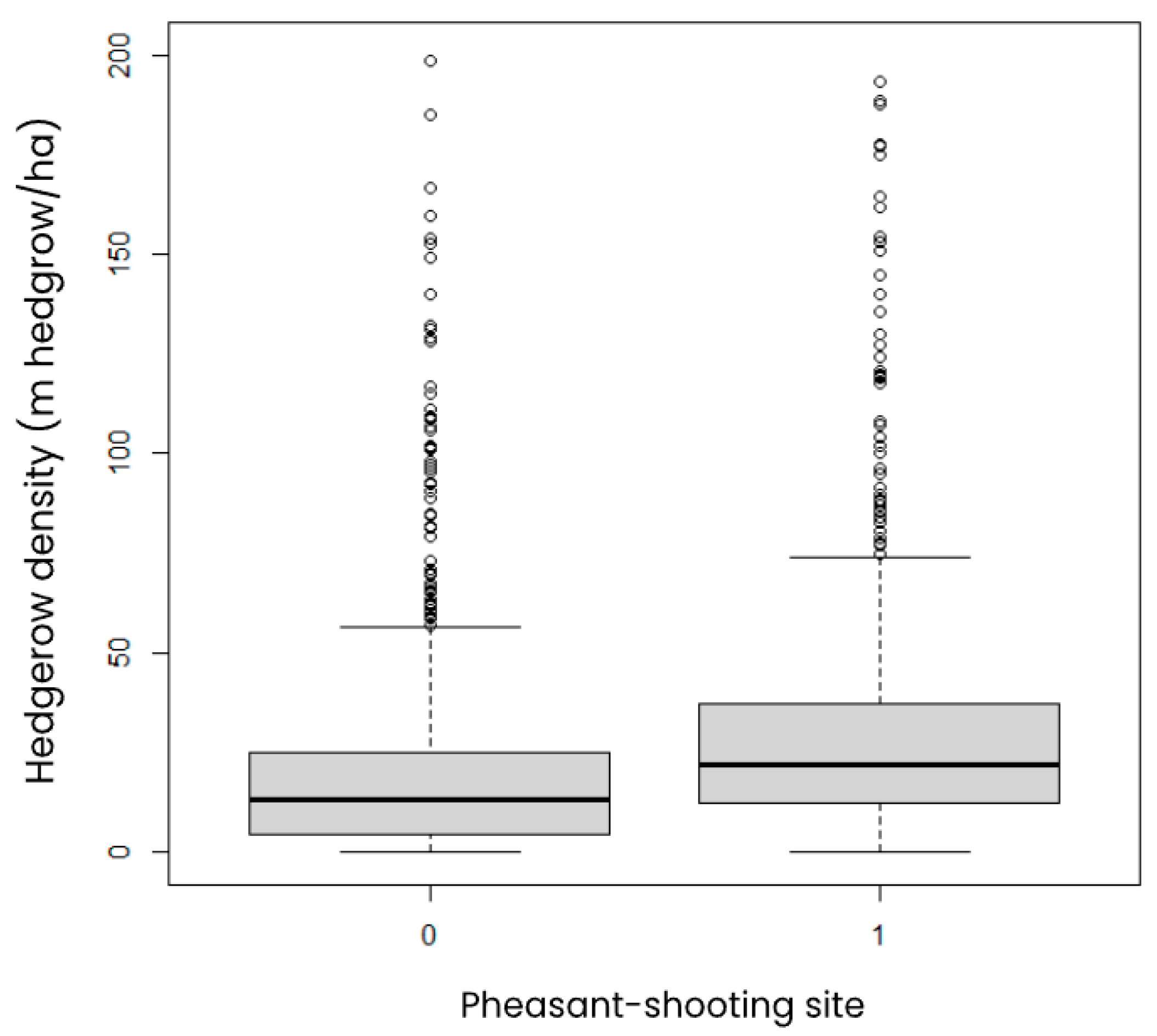
| Region | No. Sites Managed for Shooting—Remote Sensing | No. Comparison Sites—Remote Sensing | No. Sites Managed for Shooting—Literature Review | No. Comparison Sites—Literature Review |
|---|---|---|---|---|
| East Midlands, England | 117 | 117 | 5 | 5 |
| East of England | 152 | 152 | 214 | 119 |
| Northeast England | 42 | 42 | 1 | 1 |
| Northwest England | 79 | 79 | 1 | 1 |
| Scotland | 98 | 98 | 1 | 1 |
| Southeast England | 213 | 213 | 333 | 299 |
| Southwest England | 183 | 183 | 109 | 80 |
| Wales | 51 | 51 | 1 | 1 |
| West Midlands, England | 118 | 118 | 15 | 15 |
| Yorkshire Humber, England | 78 | 78 | 1 | 1 |
| Total | 1131 | 1131 | 681 | 523 |
| Birds | Aboveground Invertebrates | Vascular Plants | Mammals | Soil Invertebrates | Non-Vascular Plants | Fungi | Herptiles | Total | |
|---|---|---|---|---|---|---|---|---|---|
| Birds | 4 | 3 | 6 | 0 | 1 | 0 | 0 | 21 | |
| Aboveground invertebrates | 4 | 0 | 1 | 3 | 0 | 0 | 0 | 10 | |
| Vascular plants | 3 | 0 | 0 | 0 | 1 | 1 | 1 | 8 | |
| Mammals | 6 | 1 | 0 | 0 | 0 | 0 | 0 | 6 | |
| Soil invertebrates | 0 | 3 | 0 | 0 | 0 | 0 | 0 | 3 | |
| Non-vascular plants | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 1 | |
| Fungi | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Herptiles | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Variable | Region | Mean Pheasant Sites | Mean Comparison Sites | t | p | n (Each Group) |
|---|---|---|---|---|---|---|
| Area (ha) | East Midlands, England | 242 | 236 | −0.185 | 0.8526 | 117 |
| East of England | 254 | 254 | 0.0061 | 0.9951 | 152 | |
| Northeast England | 320 | 227 | −1.0544 | 0.2952 | 42 | |
| Northwest England | 295 | 146 | −2.6965 | 0.0077 | 79 | |
| Scotland | 495 | 335 | −1.5405 | 0.1251 | 98 | |
| Southeast England | 228 | 231 | 0.13319 | 0.8941 | 213 | |
| Southwest England | 277 | 207 | −1.1246 | 0.2618 | 183 | |
| Wales | 47 | 230 | −4.984 | 0.0001 * | 51 | |
| West Midlands, England | 258 | 234 | −0.5002 | 0.6176 | 118 | |
| Yorkshire Humber, England | 270 | 216 | −0.9042 | 0.3673 | 78 | |
| Elevation (m) | East Midlands, England | 90 | 106 | 1.2901 | 0.1987 | 117 |
| East of England | 50 | 50 | 0.09639 | 0.9233 | 152 | |
| Northeast England | 142 | 143 | 0.037499 | 0.9702 | 42 | |
| Northwest England | 101 | 115 | 0.89397 | 0.3727 | 79 | |
| Scotland | 161 | 156 | −0.25507 | 0.799 | 98 | |
| Southeast England | 82 | 88 | 1.3539 | 0.1765 | 213 | |
| Southwest England | 115 | 120 | 0.59739 | 0.5507 | 183 | |
| Wales | 105 | 93 | −0.81658 | 0.4161 | 51 | |
| West Midlands England | 113 | 127 | 1.4657 | 0.1444 | 118 | |
| Yorkshire Humber, England | 107 | 124 | 0.95971 | 0.3387 | 78 | |
| Forest 2010 (proportion) | East Midlands, England | 0.08 | 0.13 | 2.0952 | 0.03732 | 117 |
| East of England | 0.08 | 0.08 | 0.09091 | 0.9276 | 152 | |
| Northeast England | 0.19 | 0.23 | 0.74704 | 0.4572 | 42 | |
| Northwest England | 0.14 | 0.27 | 3.7205 | 0.0002 * | 79 | |
| Scotland | 0.18 | 0.14 | −1.9579 | 0.051 | 98 | |
| Southeast England | 0.11 | 0.19 | 2.7658 | 0.00661 | 213 | |
| Southwest England | 0.15 | 0.25 | 3.7327 | 0.0002 * | 183 | |
| Wales | 0.15 | 0.3 | 3.3726 | 0.0011 * | 51 | |
| West Midlands, England | 0.11 | 0.13 | 0.8982 | 0.37 | 118 | |
| Yorkshire Humber, England | 0.11 | 0.19 | 2.7658 | 0.006 | 78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, P.R.; Petrokofsky, L.; Harvey, W.J.; Orsi, P.; Jordon, M.W.; Petrokofsky, G. Structural Diversity and Biodiversity of Forest and Hedgerow in Areas Managed for Pheasant Shooting Across the UK. Forests 2025, 16, 1249. https://doi.org/10.3390/f16081249
Long PR, Petrokofsky L, Harvey WJ, Orsi P, Jordon MW, Petrokofsky G. Structural Diversity and Biodiversity of Forest and Hedgerow in Areas Managed for Pheasant Shooting Across the UK. Forests. 2025; 16(8):1249. https://doi.org/10.3390/f16081249
Chicago/Turabian StyleLong, Peter R., Leo Petrokofsky, William J. Harvey, Paul Orsi, Matthew W. Jordon, and Gillian Petrokofsky. 2025. "Structural Diversity and Biodiversity of Forest and Hedgerow in Areas Managed for Pheasant Shooting Across the UK" Forests 16, no. 8: 1249. https://doi.org/10.3390/f16081249
APA StyleLong, P. R., Petrokofsky, L., Harvey, W. J., Orsi, P., Jordon, M. W., & Petrokofsky, G. (2025). Structural Diversity and Biodiversity of Forest and Hedgerow in Areas Managed for Pheasant Shooting Across the UK. Forests, 16(8), 1249. https://doi.org/10.3390/f16081249