Abstract
Climate change has intensified over recent decades, prompting shifts in forest management strategies, particularly in the Sudetes region of Poland, where native species like Norway spruce (Picea abies), European beech (Fagus sylvatica), and silver fir (Abies alba) have historically dominated. To address these changes, non-native species such as Douglas fir (Pseudotsuga menziesii) have been introduced as potential alternatives. This study, conducted in the Jugów and Świerki forest districts, compared the soil properties and water retention capacities of Douglas fir (Dg) and Norway spruce (Sw) stands (age classes from 8–127 years) in the Fresh Mountain Mixed Forest Site habitat. Field measurements included temperature, humidity, organic matter content, water capacity, and granulometric composition. Results indicate that, in comparison to Sw stands, Dg stands were consistently linked to soils that were naturally finer textured. The observed hydrological changes were mostly supported by these textural differences: In all investigated circumstances, Dg soils demonstrated greater water retention, displaying a water capacity that was around 5% higher. In addition to texture, Dg stands showed reduced soil water repellency and a substantially greater organic matter content (59.74% compared to 27.91% in Sw), which further enhanced soil structure and moisture retention. Conversely, with increasing climatic stress, Sw soils, with coarser textures and less organic matter, showed decreased water retention. The study highlights the importance of species selection in sustainable forest management, especially under climate change. Future research should explore long-term ecological impacts, including effects on microbial communities, nutrient cycling, and biodiversity, to optimize forest resilience and sustainability.
1. Introduction
Ecosystems throughout the world are facing an unprecedented challenge due to global climate change, with forests being especially vulnerable [1,2,3]. Significant changes in forest ecosystems have occurred in recent decades, resulting in changed evolutionary paths and ecological instability [4,5]. Particularly in areas that depend on winter snow cover for water availability, these changes have a substantial influence on soil water storage capacity, which is a crucial component of forest stability [6,7]. Climate-induced disruptions, including more intense heat waves and altered precipitation patterns, result in increased soil aridity, threatening not only agricultural livelihoods but also forest-dependent communities [8,9]. Furthermore, environmental stress promotes the replacement of native species by invasive alien species, with serious negative effects on human health and biodiversity [9]. As a result, native forest ecosystems are being deteriorated by pyrophytic species proliferation, increasing wildfire frequency, and decreasing forest production [4].
The 160 million hectares of forests in the European Union are seriously threatened by the faster pace of climate change, which is surpassing the inherent adaptation capability of forest ecosystems [5,10,11]. In these increasingly complex times, traditional forest management, which is frequently focused on equilibrium-based ecology, finds it difficult to balance the preservation of biodiversity with the production of lumber [8,12]. This draws attention to a crucial area of research: understanding how various forest species and their associated soil conditions respond to climate change is essential for developing effective management and mitigation strategies.
Understanding intraspecific genetic variation is critical for forecasting forest tree species’ reactions to climate change, especially over large geographic areas [11]. Documented alterations in species distribution in Poland, particularly in the topographically varied Sudetes region, demonstrate the local flora’s low adaptive flexibility to altered environmental conditions [13]. The historical dominance of Picea abies (Spruce) monocultures in the Sudetes has made them more vulnerable to biotic and abiotic perturbations, such as insect infestations and climate-mediated phytopathologies [14]. The increased frequency of anomalous precipitation and extended xeric periods is worsening Picea abies’ susceptibility to fungal infections, such as Heterobasidion annosum-induced root rot [6].
On the other hand, non-native Pseudotsuga menziesii (Douglas fir) is gaining ecological and silvicultural importance in Polish forests [15,16,17]. Its higher productivity and perceived resistance to climate-related stresses are driving market demand and suggesting a potential shift in regional forest composition [18,19]. Poland currently hosts the largest deployment of an allochthonous forest-forming tree species in Europe, with Douglas fir plantations spanning over 823,534 hectares [20,21]. While existing forest management trends largely restrict their introduction, the reported decline of native gymnosperms like Abies alba, Picea abies, and Pinus sylvestris necessitates the investigation of alternative methods for mitigating climate change impacts [21,22].
At this point, the properties shaped by various tree species during long-term stand growth, and the soil’s capacity to retain water, are particularly significant [23]. In basic terms, water retention refers to the soil’s ability to hold water, which is primarily determined by its texture (or distribution of grain sizes) [24,25]. Also, organic matter content enhances water holding capacity, particularly in finer-textured soils, and its accumulation can be influenced by forest vegetation over stand timescales [24,25]. Soil water repellency (SWR) also impacts the water retention capacity of forest soils [26,27]. According to recent research, mild SWR can improve the distribution and storage of water in the top soil layers, which may be advantageous in the face of shifting fire and climatic regimes [28,29,30].
Considering the relevance of soil water storage capacity in facing the threat of climate change, and the documented differences between spruce and Douglas fir forest stands, there is a clear need to compare these species within the same ecosystem. Previous research has emphasized the decline of native species and the potential for non-native replacements, yet a systematic assessment of their ecological consequences on critical soil qualities, particularly considering existing soil characteristics, remains unstudied.
Therefore, given the context of climate change and its implications for soil water storage capacity, and acknowledging the observed contrasting characteristics of spruce and Douglas fir forest stands and their associated inherent soil conditions, this research aimed to compare Douglas fir and spruce forest stands within the same habitat, examining soil property variables and tree characteristics. The main hypothesis is that established Douglas fir stands, occurring on inherently finer-textured soils, exhibit distinct and potentially advantageous soil water retention qualities and stand features compared to spruce stands in similar climatic circumstances.
This research involved measuring trees and conducting a field assessment of soil profiles in paired Douglas fir and spruce stands in the Sudetes of Poland. Soil texture, organic content, water retention curves, and water repellency indices were all evaluated by laboratory tests. The data’s statistical comparisons evaluated species differences in an effort to provide insight into each species’ unique contributions to the stability of forest ecosystems in the face of climate change.
2. Materials and Methods
2.1. Area Description
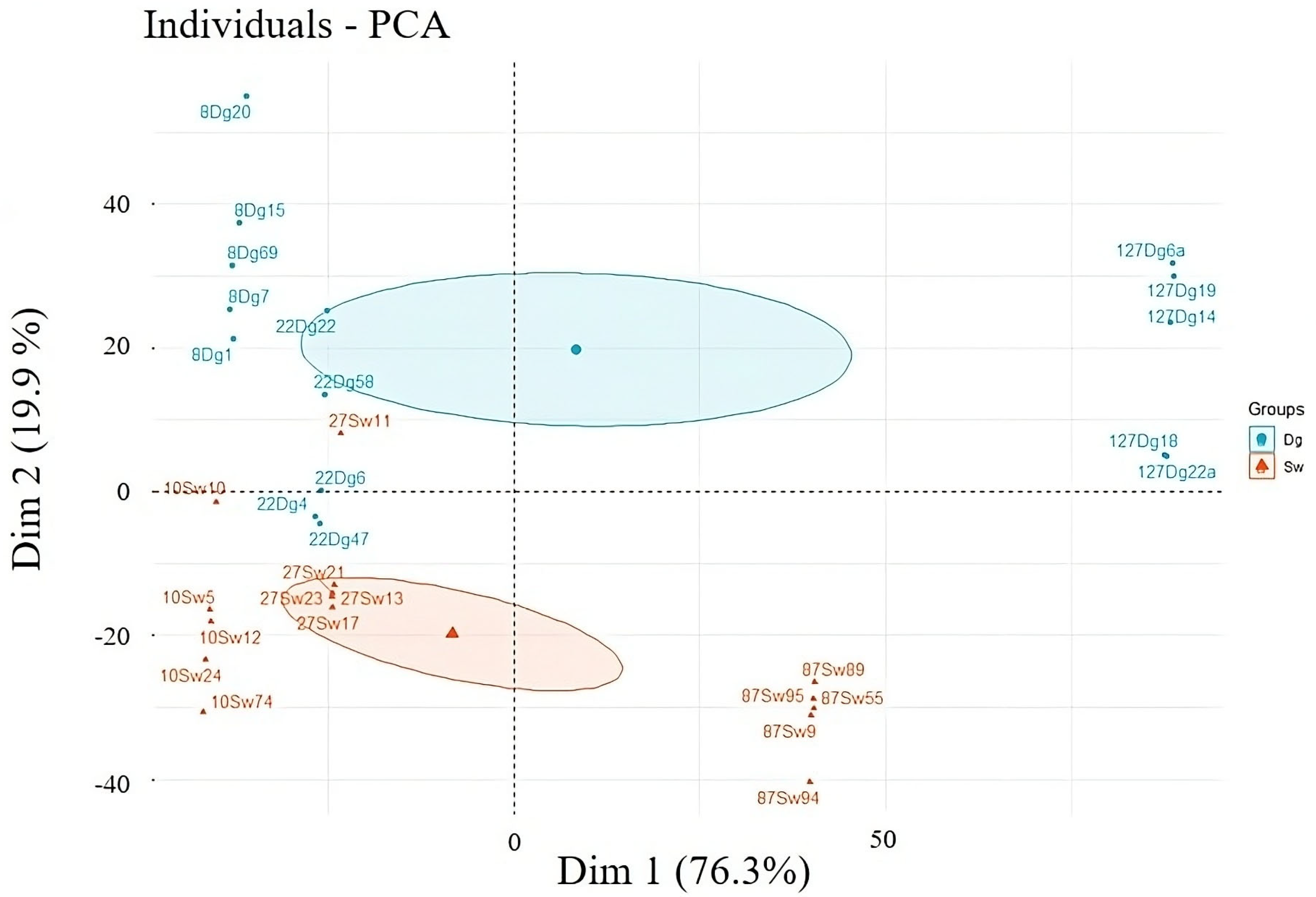
The research areas, due to the availability of suitable stands necessary for conducting the research, were located in the Jugów Forest and Świerki Forest districts. This area is located in the Włodzica River Valley. This region is characterized by numerous watercourses and streams, but most of them occur only seasonally. Due to the numerous slopes of the terrain, which accelerate surface runoff and, consequently, water outflow into areas located in the depressions of the terrain. The selection of research areas depended on the species composition of the stand, the size of the share of the studied species, and the age of the trees. The focus was on selecting stands with the highest possible share of the studied species. For both spruce and Douglas fir, three sample areas were selected in the Fresh Mountain Mixed Forest Site (LMGśw) habitat. The retention properties were studied in three different spruce stands, the age of which was 10, 22, and 87 years (I, II, and III age classes), while the age of the Douglas fir stands was 8, 22, and 127 years (I, II, III age classes). The source document used to obtain information on a given division was the Forest Management Plan of the selected forestry (Figure 1).
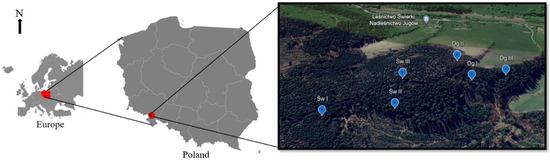
Figure 1.
Study area with Douglas fir (Dg) and spruce (Sw) forest stand in Jugów Forest District in the Włodzica River Valley, Poland.
The study area experiences a transitional climate between oceanic and continental influences typical of southwestern Poland. The mean annual temperature is around 8 °C, with daily highs reaching approximately 21–24 °C in July and lows dropping to –6 °C in January. Annual precipitation averages between 750–800 mm, with the highest monthly totals in June and July (~90 mm) and the lowest in February (~40 mm). The research was conducted on plots located on slopes with southern, south-eastern, and south-western exposures, at elevations ranging from 657 to 673 m above sea level. The slope gradients were gentle, ranging from 2° to 5°. Canopy closure varied across the plots—from full closure in younger plantations and pole-stage stands, through moderate closure in stands that had undergone initial shelterwood cuts.
The Włodzica River Valley occupies a unique geotectonic position at the junction of three distinct rock massifs: the Stone Mountains, Sowie Mountains, and Table Mountains. The valley is underlain by dark, weakly calcareous clays, previously classified as Lower Cretaceous in age. However, recent findings have revealed the presence of olistoliths—large, light-colored calcareous rock blocks—embedded within these clays. These olistoliths contain microfaunal assemblages characteristic of the Cenomanian and Maastrichtian stages, indicating complex stratigraphic relationships and suggesting post-depositional transport or tectonic emplacement. Additionally, the valley exhibits features indicative of redeposition processes, with loess deposits that may have originated from the Sudeten region. The geomorphological context is further emphasized by the position of Sarny Castle, which rests on the slope of the Stone Mountains, with the Sowie Mountains extending behind it and the Table Mountains beginning across the Ścinawka River, delineating a distinct boundary among these geological units [31].
The Włodzica River Valley in Poland is predominantly characterized by alluvial soils, locally referred to as mady. These soils are typically nutrient-rich and form in fluvial environments, making them highly suitable for agricultural use. Their physical and chemical properties, as well as their dynamic transformation processes, have been the subject of ongoing pedological and environmental research. The distribution and composition of these alluvial soils vary across the valley, with finer, heavier textures commonly found in proximity to the river channel and lighter, sandier or silty variants occurring at greater distances from the river. This spatial variability reflects the influence of hydrological regimes and sediment deposition patterns associated with periodic flooding and overbank flow [32].
2.2. Field Work
For both Douglas fir and Norway spruce, trees ranging from 8 to more than 100 years old were selected for each site, distributed in each age category in Dg and Sw (I, II, II) (Figure 1).
Following the completion of initial site preparations, environmental parameters, including ambient air temperature and relative humidity, were measured using a calibrated thermohygrometer. These measurements were taken directly at the study site to accurately characterize the prevailing environmental conditions during the period of data collection, which may influence soil physical properties and infiltration behavior.
A rectangular plot measuring 1.0 m × 1.5 m was subsequently delineated for soil sampling and infiltration measurements. To ensure direct and uniform access to the soil surface, all overgrown vegetation, as well as the upper layer of undecomposed organic matter (e.g., leaf litter and surface debris), was meticulously removed using appropriate manual tools such as hand rakes and spades. This careful preparation minimized surface obstructions and ensured consistency across sampling points.
Soil infiltration capacity was assessed using a Mini Disk Infiltrometer, which delivered a controlled volume of 2 cm3 of water to the prepared soil surface. Prior to each infiltration test, the instrument was calibrated and positioned according to the manufacturer’s operational protocol to ensure measurement accuracy and repeatability. The time required for the water to fully infiltrate the soil was recorded, providing data on the infiltration rate and surface hydraulic conductivity under the specific field conditions present at the time of measurement.
For the purpose of soil physical property analysis, a total of 15 undisturbed soil core samples were collected randomly from each plot selected using standard Kopecki cylinders with an internal volume of 100 cm3, and a diameter of 5 cm. Sampling locations were systematically distributed, with cores extracted from the four corners and the central point of the plot. All samples were taken at a uniform depth of 15 cm to ensure comparability across the plot and to reflect conditions in the topsoil layer, which is most responsive to surface treatments and environmental variations.
2.3. Tree Height Measurement Using Suunto Altimeter
Tree height was measured using a Suunto altimeter (Finland), based on the principles of trigonometric height calculation.
The observer positioned themselves at a known horizontal distance (typically 15 or 20 m) from the base of the tree on level ground. The distance was measured using a measuring tape or laser rangefinder for accuracy. Using the Suunto altimeter, the observer sighted the top of the tree and recorded the angle of elevation. If the base of the tree was above or below eye level, the angle to the base was also measured. The height of the tree was calculated using basic trigonometric functions. When both top and base angles were measured, the total height (H) was derived using the Formula (1):
where:
D = horizontal distance to the tree,
= angle to the top of the tree,
= angle to the base of the tree (use negative values for downward angles).
All measurements were conducted under stable atmospheric conditions to minimize optical distortion and human error. Each tree was measured from at least two directions, and the average value was used as the final tree height.
2.4. Work Performed in the Laboratory
The first step of the laboratory work was to weigh the mass (M) samples as soon as possible after collecting them from the field, like mass fresh (Mf). The weights of individual samples were measured using a laboratory balance, and the results were measured to the nearest 0.1 g.
In order to simulate short-term precipitation and gain information about changes in capacity during soaking, the collected mass samples were immersed in water for 4 and 24 h. After this time, they were taken out of it and weighed again on a laboratory balance (M4 and M24). The important information we are able to obtain during this type of measurement is to know how much water the soil can hold during steady and uniform rainfall.
To obtain information on what amount of water was in each sample, they were placed in a laboratory dryer for 24 h at 105 °C, and then their weight was measured as mass dried (Md).
2.5. Calculation of Water Capacities of Collected Samples
Each step of measuring the masses of the collected samples was aimed at calculating what mass of water the soil could take up and retain at different degrees of moisture content, so from the differences in masses was calculated, current water capacity (Sa), using the Formula (2), after 4 h in water (S4), using the Formula (3) and after 24 h in water (S24), using the Formula (4).
where:
Sa—current water capacity;
Mf—fresh mass, i.e., the mass of water in the samples immediately after collection from the site;
Md—dry mass, the mass of the soil sample after 24 h in a drying oven at 105 °C.
where:
S4—water capacity after 4 h in water;
M4—mass of soil samples after 4 h immersion in water;
Md—dry mass.
where:
S24—water capacity of the sample, after 24 h in water;
M24—mass of soil samples after 24 h of immersion in water;
Md—dry mass.
Overlaying soil samples with ethanol solutions (MT) and distilled water (WDPT). The hydrophobicity of various substances is that the greater the hydrophobicity, the less capacity a material has to infiltrate and store water. To assess this property, two tests were performed. The water drop penetration time (WDPT) test measured the time required for distilled water drops to infiltrate fresh soil samples, providing an indicator of surface hydrophobicity under field-moist conditions. In contrast, the molarity of ethanol droplet (MT) test was conducted on samples previously dried at 105 °C (for dry mass determination). Using a medical dropper, five droplets of ethanol solutions with increasing concentrations (from 0% to 30%, in 2.5% increments) were applied to the soil surface to determine the lowest concentration that infiltrated within 5 s. Both tests helped characterize the strength and persistence of soil water repellency under different moisture conditions [33,34], as presented in Table 1 for results.

Table 1.
Description information of the variables by species.
2.6. Removal of Organic Matter from Collected Samples
Organic matter significantly influences soil hydrophilicity and water retention capacity. To evaluate which tree species—green Douglas fir (Pseudotsuga menziesii) or common spruce (Picea abies)—contributes more biomass to the substrate, collected soil samples were subjected to combustion at 700 °C in a muffle furnace. This process eliminated all organic phytomass. By comparing the oven-dry weights of the samples before and after combustion, the organic matter content was calculated gravimetrically.
2.7. Granulometric Composition Tests
Another key factor determining the soil’s ability to retain water is its granulometric composition. To assess the distribution of particle size fractions like clay < 0.002; silt 0.002–0.05; fine silt 0.002–0.0063; coarse silt 0.0063–0.02, and sand 0.05–2.00 (mm). A laser diffraction method was used. Soil samples were analyzed with an Analysette 22 laser particle size analyzer (Fritsch GmbH), and the results were expressed as percentages [%] of the total mass for each granulometric fraction.
2.8. Statistical Data Analysis
A Pearson correlation matrix was performed between the variables. ANOVA was used to determine which of the variables had the greatest influence, and a principal component analysis (PCA) was used to determine which factors were grouped by species. The analyses were performed in R-Studio version 4.4.2 [35].
3. Results
The results obtained in this study directly address the research aim by highlighting significant differences in soil water retention and stand characteristics between Douglas fir and spruce forests. These findings provide valuable insight into the ecological suitability of Douglas fir as a potential alternative in habitats currently dominated by spruce, particularly in the context of increasing climate-related water stress.
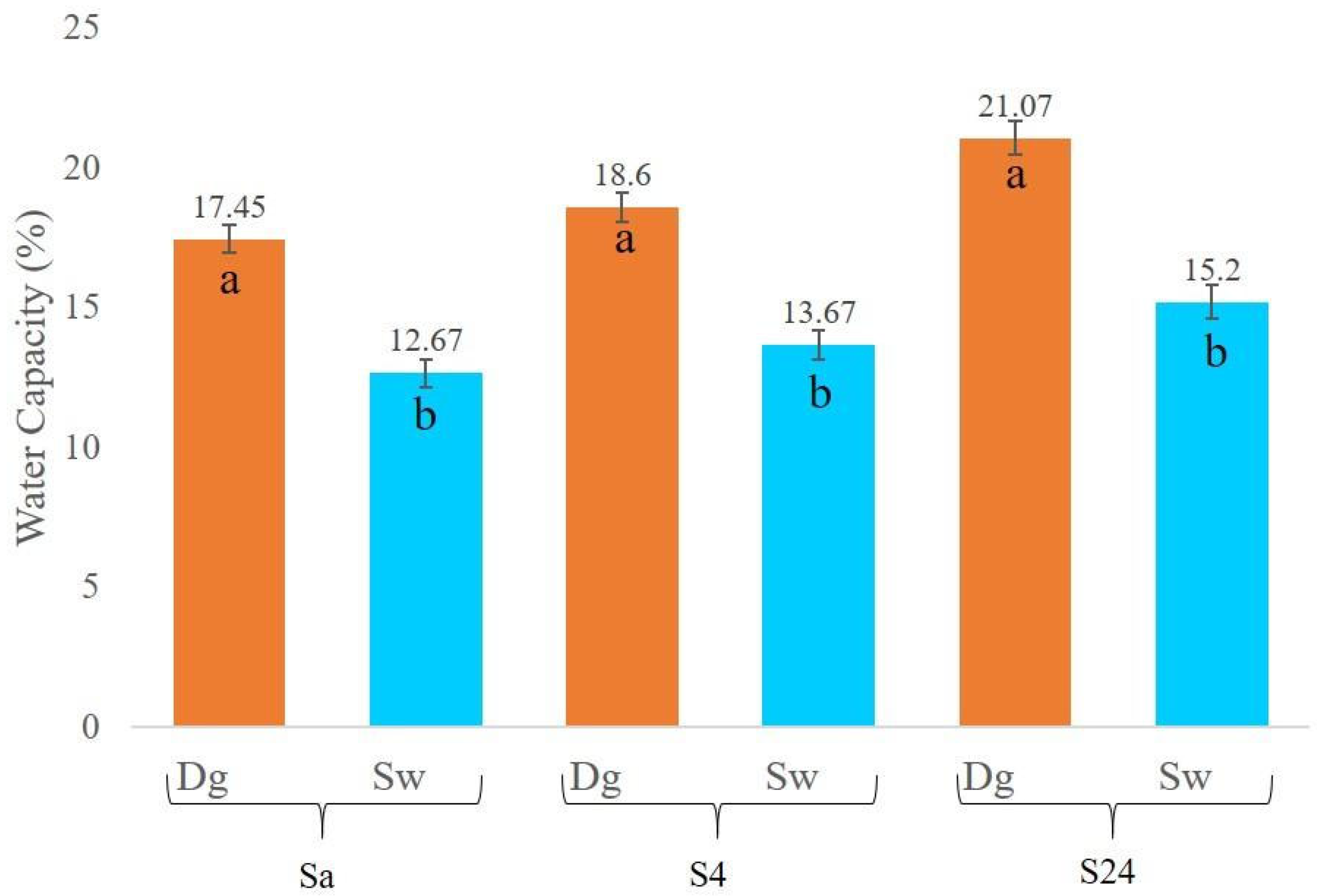
A comparison of soil water retention capacity in Douglas fir (Dg) and spruce (Sw) stands was conducted at three time intervals: immediately after sampling (Sa), after four hours (S4), and after 24 h (S24). As shown in Figure 2, soils under Douglas fir consistently exhibited higher water retention capacity across all measurement times. The observed differences were statistically significant (p < 0.0001), with values approximately 5% higher than those recorded for spruce stands.
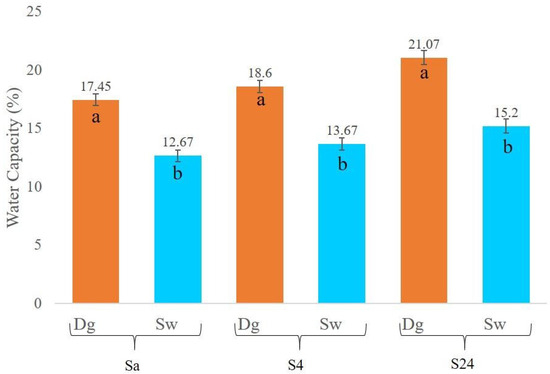
Figure 2.
Water capacity in (%) current (Sa), after 4 h (S4), and after 24 h (S24) by species. Douglas stand (Dg); spruce stands (Sw). Means with a common letter are not significantly different (p > 0.05).
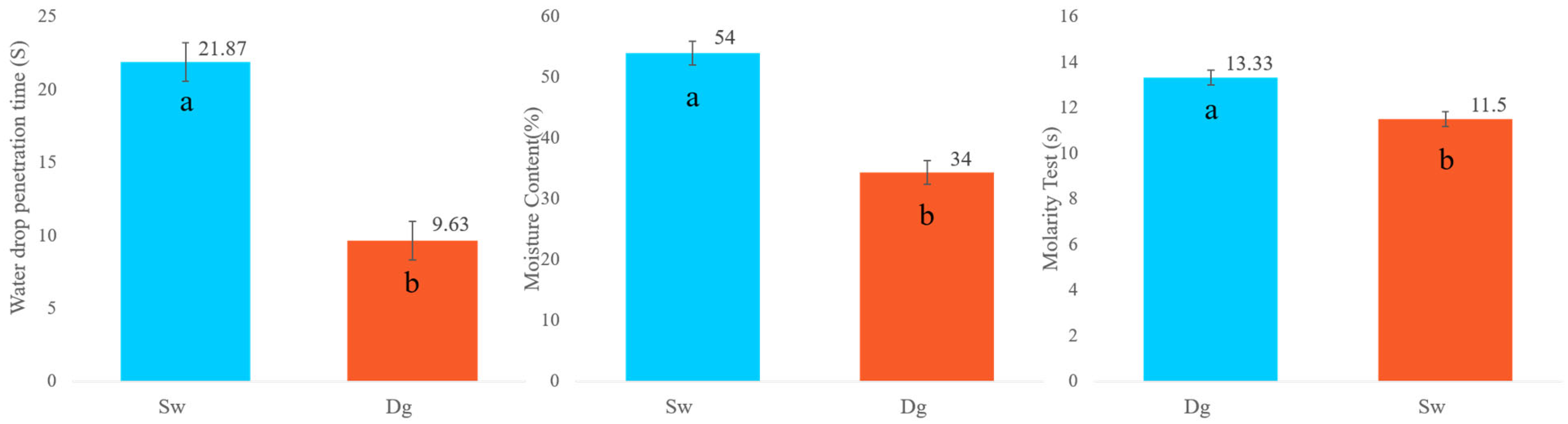
For the water drop penetration time (WDPT) test, both Douglas fir (Dg) and spruce (Sw) soils were classified as exhibiting low water repellency (SWR), with values ranging from 5 to 60 s, indicating that “treatment will improve” water infiltration. However, as shown in Figure 3, Douglas fir soils exhibited a significantly shorter average WDPT (9.63 s) compared to spruce soils (21.87 s), suggesting faster water infiltration in Dg stands.
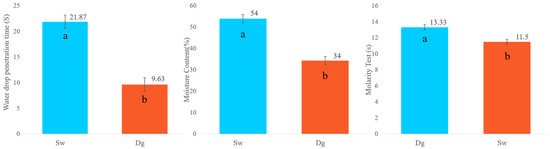
Figure 3.
Water drop penetration time (WDPT), moisture content, and molarity test (MT) by species. Douglas stand (Dg); spruce stands (Sw). Means with a common letter are not significantly different (p > 0.05).
In contrast, soil moisture content was significantly higher under spruce, with approximately 20% more water retained compared to Douglas fir.
Results from the molarity of ethanol droplet (MED) test further confirmed these differences: spruce soils showed water repellency at a lower ethanol concentration (11.5%) compared to Douglas fir soils (13.33%), with the difference being statistically significant (Figure 3).
A lower threshold of hydrophobicity indicates that soils under spruce become water-repellent at a lower ethanol concentration, meaning they are more hydrophobic than soils under Douglas fir.
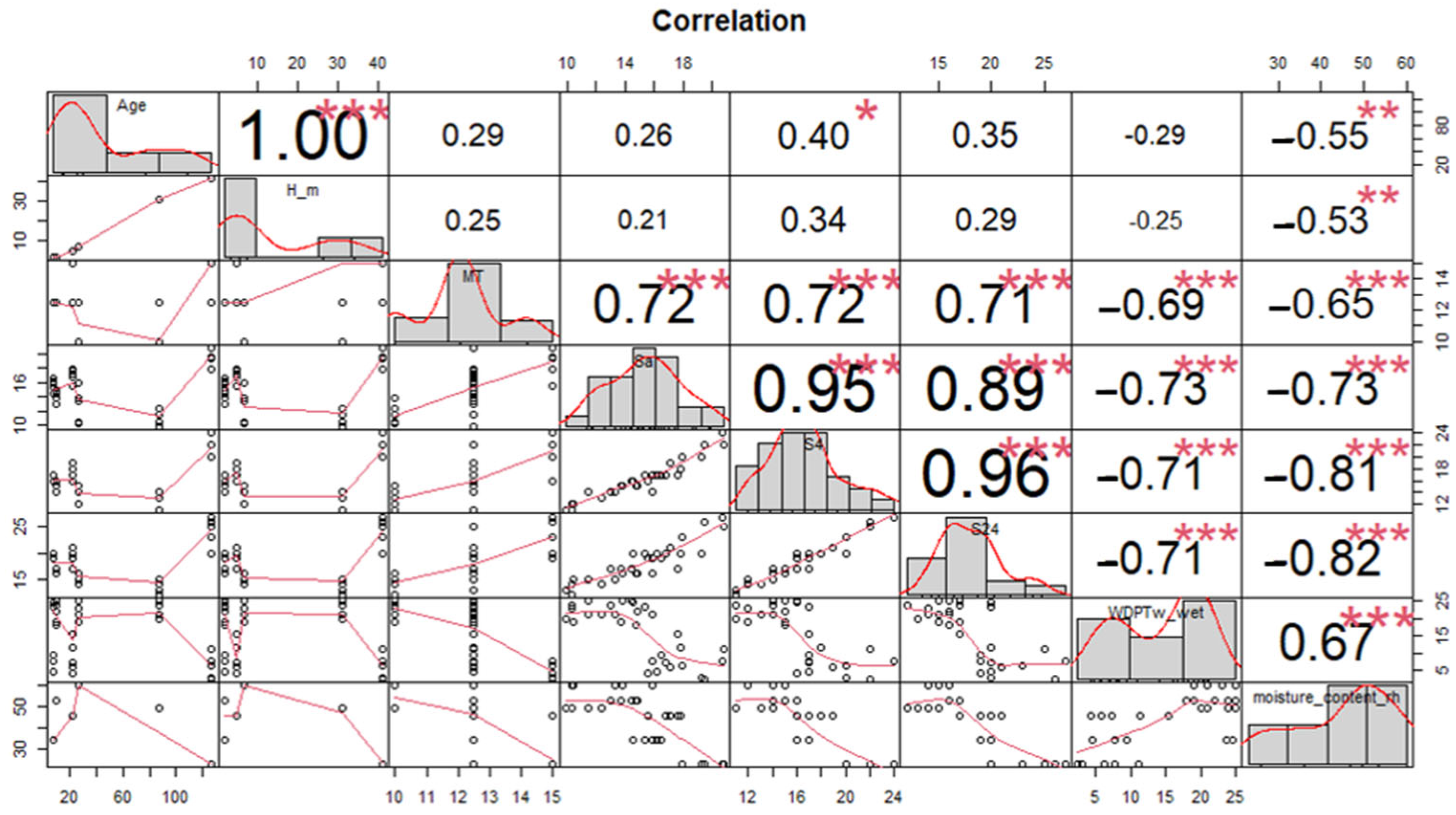
A parallel result of a Pearson correlation of the variables analyzed without the Species category is presented in Figure 4, where two correlated groups can be seen. One group (MT, Sa, S4, S24) is in a positive correlation. On the other hand, the group (WDPT, Moisture Content, Age, and Height of the Tree) is in a strongly negative correlation.
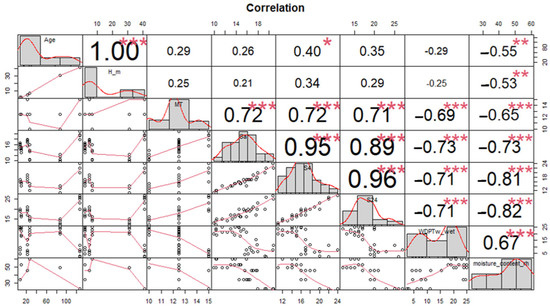
Figure 4.
Pearson correlation matrix between variables. Age, tree height (H_m), molarity test of ethanol droplet (MT), water capacity current (Sa), after 4 h (S4) and after 24 h (S24), water drop penetration time (WDPT), and moisture content. no significant differences (ns) p > 0.05; * p ≤ 0.05; ** p ≤ 0.01 and *** p ≤ 0.001.
3.1. Granulometric Fraction Composition and Organic Matter
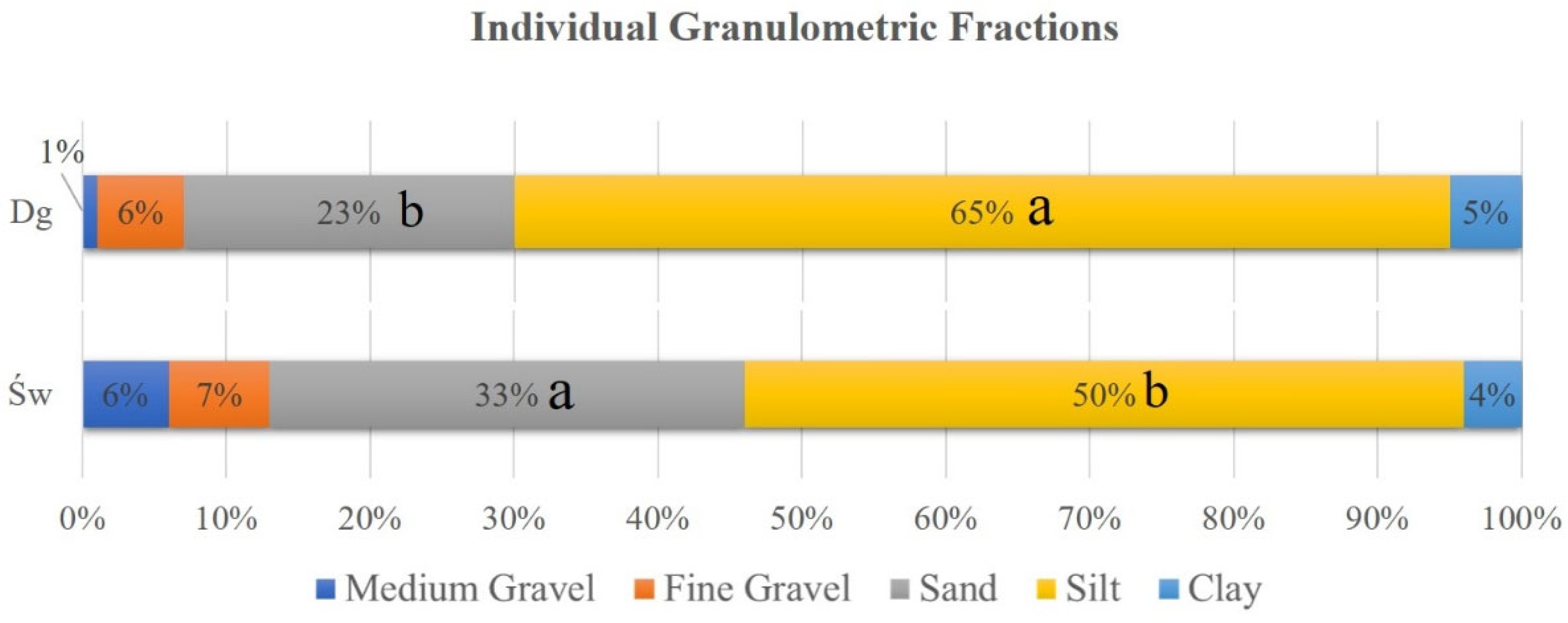
The granulometric composition of the soil at a depth of approximately 15 cm revealed significant differences between the study areas. The Douglas fir (Dg) stands exhibited a higher proportion of the silt (dust) fraction (65%) compared to the spruce (Sw) stands (50%). Conversely, the sand fraction was significantly more abundant in spruce soils (33%) than in Douglas fir soils (23%) (Figure 5). No significant differences were observed in the remaining soil fractions.
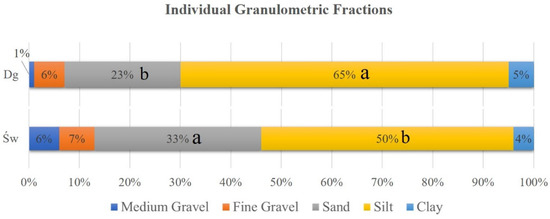
Figure 5.
Percentage of granulometric fractions in samples by species. Douglas stand (Dg); spruce stands (Sw). Means with a common letter are not significantly different (p > 0.05).
In terms of soil organic matter content, Douglas fir soils contained a significantly higher percentage (59.74%) compared to those under spruce (27.91%).
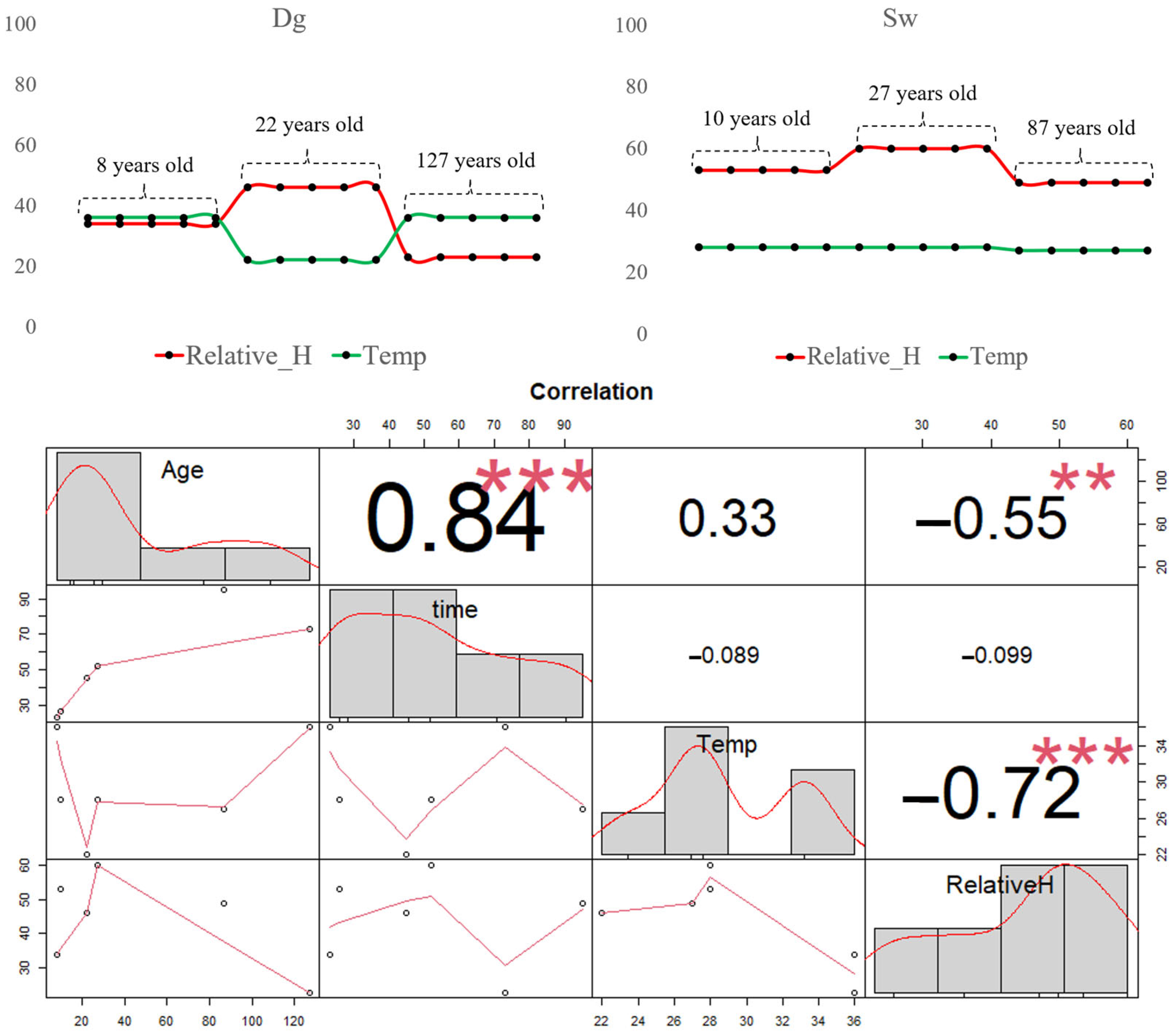
Analysis of the Pearson correlation matrix revealed a strong positive correlation (r = 0.84 ***) between stand age and infiltration time (measured in seconds using the Mini Disk Infiltrometer). This indicates that older stands required more time for the same volume of water to infiltrate into the soil. This trend was consistent across both Douglas fir (Dg) and spruce (Sw) stands, although Sw exhibited longer maximum infiltration times (up to 95 s) compared to Dg (maximum 73 s).
The soil temperature (green line) showed no significant correlation with infiltration time but had a weak positive correlation with stand age (r = 0.33), suggesting a slight tendency for older trees to be associated with higher soil temperatures. However, when analyzed separately by species (Figure 6), this pattern was only observed in Dg stands, where soil temperature increased with tree age. In Sw stands, temperature remained relatively stable regardless of age.
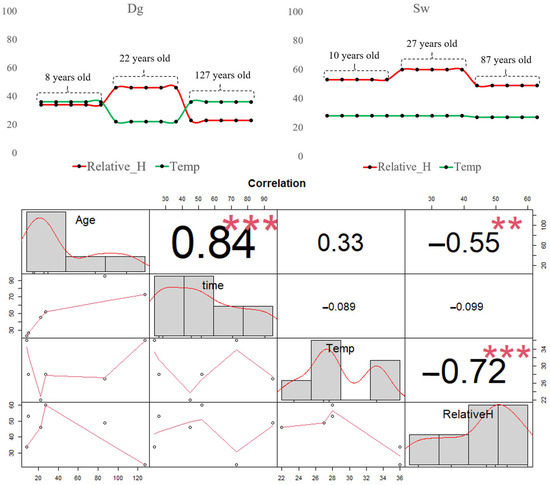
Figure 6.
Line of time with three variables: temperature (green line) and relative humidity (red line). Below is the Pearson correlation matrix. No significant differences (ns) p > 0.05; ** p ≤ 0.01 and *** p ≤ 0.001.
Relative soil moisture (red line) was negatively correlated with soil temperature (r = −0.72 ***), indicating that higher temperatures are associated with lower moisture levels. This relationship was particularly evident in Douglas fir stands. Additionally, a moderate negative correlation (r = −0.55 **) was found between relative soil moisture and stand age, suggesting that older trees are associated with drier soil conditions. This trend was observed in both species.
3.2. The Principal Component Analysis (PCA)
Principal component analysis (PCA) results (Figure 7, left) indicate that the variables Age, Organic Matter (OM), and Tree Height (H_m) had the highest contributions, collectively explaining most of the variance among the analyzed variables. In Figure 7 (right), additional variables such as Soil Moisture Content also contributed meaningfully to the model, alongside Age, OM, and Tree Height, while the remaining variables each contributed less than 20% to the explained variance.
In contrast, the spruce (Sw) plots were more closely associated with vectors such as Soil Moisture Content, WDPT, Sand, Medium Gravel, and Fine Gravel. This distribution may indicate that Sw is more likely to occur in, or tolerate, more degraded or coarse-textured soils. Alternatively, it is also possible that the long-term presence of spruce contributes to gradual soil degradation.
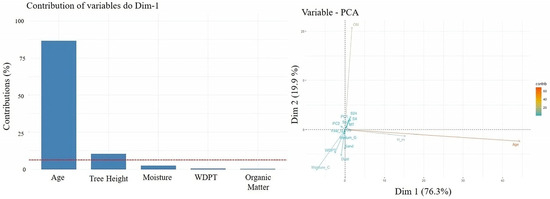
Figure 7.
Contribution of variables in % individual (Left Top 5) and variable in PCA (Right); Age, tree height (H_m), molarity test of ethanol droplet (MT), water capacity current (Sa), after 4 h (S4) and after 24 h (S24), water drop penetration time (WDPT), and moisture content. When a categorical variable (forest species) was introduced into the PCA (Figure 8), a clear separation emerged. The Douglas fir (Dg) plots clustered along the principal vectors with the greatest influence—namely OM, Age, and Tree Height—variables that are often associated with improved soil quality. This suggests that Dg may be better adapted to the study ecosystem, or that its presence supports the development of more favorable soil conditions.
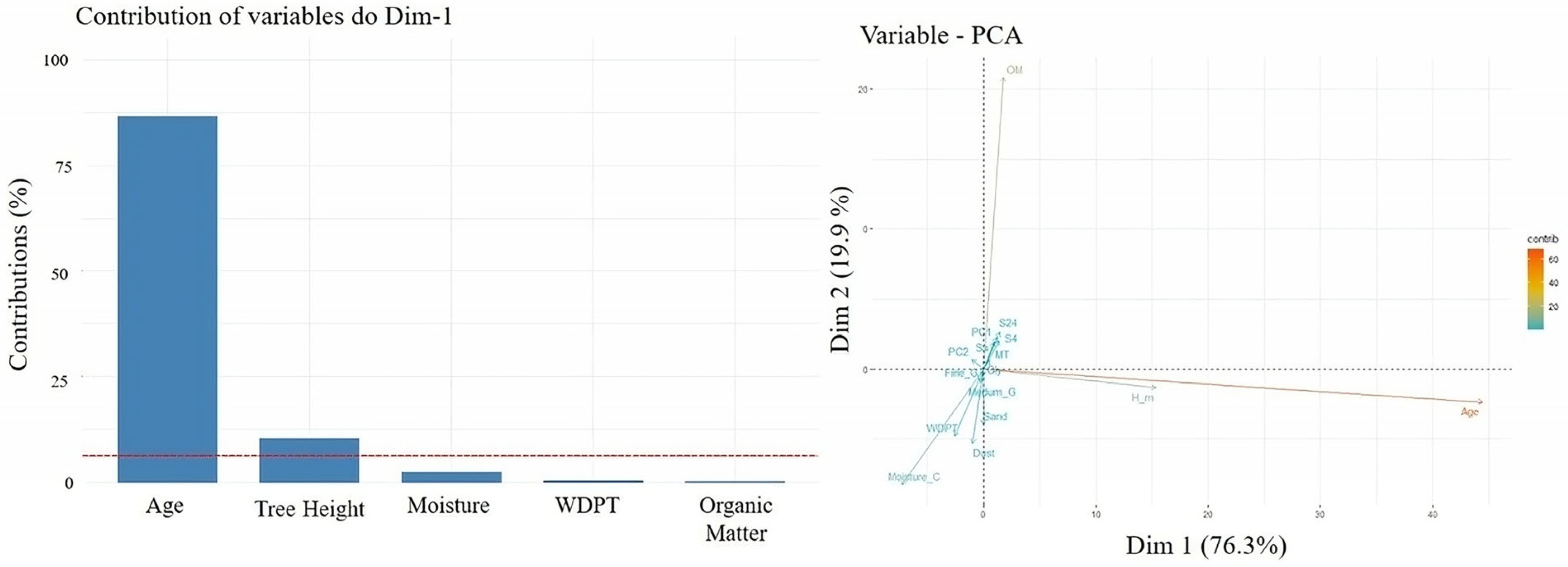
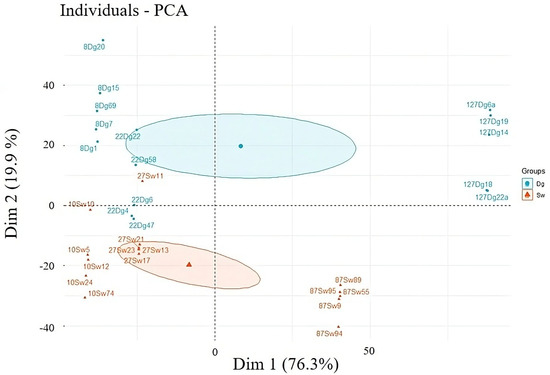
Figure 8.
Individuals—PCA by the first number is the age, followed by the species and then by the sample code: Douglas stand (Dg); spruce stands (Sw).
4. Discussion
This study aimed to compare soil hydrological properties and tree characteristics between established Douglas fir (Pseudotsuga menziesii) and spruce (Picea abies) stands growing under similar climatic and site conditions in the Sudetes of Poland. The results highlight ecological differences that can inform decisions regarding species selection and forest adaptation strategies in the face of climate change.
4.1. Tree Growth and Stand Characteristics
Remarkably, when we compared overall data without distinguishing between species, we discovered no significant variations in tree height or age between Douglas fir and spruce populations. Despite species-specific features, this convergence in development patterns might be due to severe competition for common resources, such as water and light, within the same habitat, where environmental circumstances are almost the same for both species. A study [36] on the impacts of animal density in Douglas fir and spruce forests found no significant variations in development patterns, supporting the idea that species-specific differences might be reduced under uniform environmental strain. This competitive dynamic, particularly for light and water under similar site circumstances, might result in convergence in tree growth. Additional research [37] demonstrates that species-specific features, such as rooting patterns, have a major impact on competitive dynamics. This study, which shows how species such as beech maintain growth during droughts due to fine root biomass distribution while Douglas fir growth slows, emphasizes the importance of complementary root strategies for improving resource utilization and stand productivity in mixed versus monospecific stands. whereas this stand-level data indicated broad shared characteristics, further detailed examination per species revealed slight but significant differences in soil properties.
4.2. Soil Texture and Water Retention
The two stand types’ soil particle size distributions differed significantly and statistically significantly, according to the granulometric study. The proportion of fine particles in soils associated with Douglas fir stands was much greater (65% silt, 23% sand) than in soils associated with spruce (50% silt, 33% sand). Since soil texture is mostly an intrinsic characteristic that has been developed over long geological timeframes, and is typically barely altered by plants within a single stand cycle. The idea that Douglas fir prefers to establish on naturally finer-textured soils, while spruce may tolerate or even prefer coarser textures in this area, is supported by the fact that these textural differences are most likely pre-existing site characteristics that may affect the distribution and success of these tree species.
The observed differences in soil water-holding capacity are directly caused by this natural textural variation. Across all studied time intervals (immediately after saturation, after 4 h, and after 24 h), samples from Douglas fir stands consistently held onto more water, showing a roughly 5% better water-holding capacity than spruce soils. This is in line with well-established soil science theories that state that finer-textured soils are inherently better at retaining water because of their increased surface area and smaller pore diameters [24,25].
4.3. Species-Specific Contributions to Dynamic Soil Properties
The much greater organic matter concentration under Douglas fir (59.74%) compared to spruce (27.91%) suggests species-specific impacts on litter input, breakdown, and total soil organic matter cycling, along with the main influence of intrinsic roughness. Given the favorable microclimatic and microbiological conditions that this species may create, Douglas fir likely supplies more biomass from litterfall and root exudates, which can break down more effectively [38]. For example, German studies show that on similar locations, Douglas fir is substantially more productive than native species such as pine, spruce, beech, and oak [39,40].
Increased organic matter input into the soil is an inevitable result of these greater production levels. On the other hand, slower rates of decomposition or lesser-quality litter may be the cause of the reduced organic matter beneath spruce, which would contribute less to the stable soil organic pool [41]. Moreover, [38] has demonstrated that Douglas fir stands support more dynamic nitrogen transformations and greater microbial activity than spruce, resulting in more favorable pedochemical characteristics and active soil layers.
A further significant aspect is Douglas fir’s deeper root system [42]. Deeper roots can increase water penetration, decrease surface runoff, and encourage bioturbation and soil mixing, all of which indirectly support better soil structure and water retention without directly changing the texture of bulk soil [15,43]. The shorter water drop penetration time (WDPT) seen in Douglas fir stands, which suggests greater wettability and quicker infiltration rates, may be partially explained by this rooting approach.
4.4. Soil Water Repellency and Hydrological Implications
Regarding soil water repellency (SWR) properties, both species’ soils fell into the low repellency category, however Douglas fir soils were much more wettable (WDPT ≈ 9.63 s) than spruce (WDPT ≈ 21.87 s). The molarity of ethanol droplet (MED) test corroborated these differences, with Douglas fir soils requiring a bit more ethanol (13.33%) than spruce (11.5%) to induce repellency. Spruce soils’ considerably stronger repellency may restrict infiltration during dry seasons, needing special soil management measures to counteract this impact. The observed variations in repellency and moisture retention call for additional inquiry, notably into the effect of species-specific root exudates, litter composition, and microbial community structure. Long-term monitoring would also be important in determining how these traits change in the context of species identification and climatic trends.
The different hydrological tendencies and water retention properties between spruce and Douglas fir emphasize how crucial species selection is to forest management, especially when it comes to overcoming climate change. According to [42], Douglas fir trees have deeper roots, which improves their access to subsurface water and increases their resistance to drought [44,45]. Spruce, on the other hand, tends to use surface moisture more quickly due to its shorter roots, which can accelerate depletion and exacerbate drought stress. Given their hydrological traits and vulnerability to deterioration under shifting environmental conditions, historical data [46,47] showing a decrease in species diversity as a result of pollution and an increase in spruce dominance raise questions regarding the long-term suitability of widespread spruce monocultures. Douglas fir soils’ greater wettability supports their capacity to improve hydrological conditions, which is essential for reducing drought [48].
4.5. Ecological and Management Considerations
Although established Douglas fir stands appear to offer advantages in terms of soil health and hydrology, the introduction of this non-native plant doesn’t come without ecological issues. Due mainly to shading effects, Douglas fir’s wide ecological amplitude and strong competitive capacity may cause it to outcompete native species and decrease understory diversity [38,49,50]. Because of its size and quick development, it can suppress herbaceous and shrub-layer vegetation as well as other tree species, which might simplify the structure of the forest floor [38]. A moderate introduction of Douglas fir, for instance, keeping it at about 30% in mixed stands, may help preserve the productivity of dwindling spruce forests while still being socially and environmentally acceptable, according to some research [51,52]. According to this standpoint, if its use is properly thought out and matched to habitat-compatible compositions, Douglas fir might be a useful element in mixed-species stands intended for resilience, productivity, and climate change adaptation [42,53,54].
From the perspective of pragmatic forestry, the thoughtful incorporation of Douglas fir into current or future stand compositions may provide quantifiable advantages for long-term forest production, soil quality, and water control. However, adaptive silvicultural techniques, like as species mixing, long-term monitoring, and site-specific planning, are crucial to maintaining ecological balance and preventing the dominance of a single, non-native species.
5. Conclusions
Dominant tree species have a high correlation with particular soil characteristics that affect water retention and other important ecological processes. Douglas fir stands, which are generally located on soils with a naturally finer texture, continuously showed greater levels of organic matter and better water retention. Which, along with Douglas fir’s deeper root system and better soil water repellency, point to the tree’s great potential for improving soil resilience and health, especially in areas that are prone to drought. On the other hand, spruce stands, often found in soils with a rougher texture, exhibited less water retention, which would suggest that they are less sustainable over the long term under rising climate stress.
Whereas Douglas fir has many benefits for soil health and hydrology, particularly when established on naturally more hospitable soil types, its introduction must be carefully weighed against the ecological effects on native species and ecosystem dynamics. Long-term monitoring of species-specific effects on dynamic soil characteristics such microbial communities, nitrogen cycling, and carbon sequestration should be a top priority for future studies. In the face of continuous climate change, these studies, along with thorough evaluations of the implications on biodiversity, will be essential for creating adaptive and sustainable forest management plans that capitalize on species-specific advantages while preserving ecological integrity.
It is important to clarify that this study does not claim that Douglas fir alters soil texture within a single stand generation. Rather, the observed differences in water retention and organic matter content are interpreted in the context of pre-existing edaphic conditions, where Douglas fir tends to occur on naturally finer-textured soils compared to Spruce. This distinction ensures that species-related differences are not conflated with long-term pedogenic processes.
Author Contributions
Conceptualization, A.K.-I. and P.B.; methodology, A.K.-I.; software, L.R.-B.; validation, L.R.-B.; formal analysis, L.R.-B.; investigation, A.K.-I.; data curation, P.B. and E.S.-O.; writing—original draft preparation, P.B.; writing—review and editing, L.R.-B.; visualization, M.I.D.-M. and L.R.-B.; supervision, A.K.-I.; project administration, A.K.-I.; funding acquisition, A.K.-I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financed by the Ministry of Science and Higher Education of the Republic of Poland, Department of Ecological Engineering and Forest Hydrology, and also Rector’s Grant A446.
Data Availability Statement
The data is available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| Dg | Douglas Fir |
| Sw | Norway Spruce |
| WDPT | Water Drop Penetration Time |
| H_m | Tree Height |
| Sa | Water Capacity |
| S4 | Water Capacity After 4 Hours |
| S24 | Water Capacity After 24 Hours |
| MT | Molarity Test with Ethanol Solution |
| PCA | Principal Component Analysis |
| SWR | Soil Water Repellent |
| OM | Organic Matter |
References
- Hartmann, H.; Bastos, A.; Das, A.J.; Esquivel-Muelbert, A.; Hammond, W.M.; Martínez-Vilalta, J.; McDowell, N.G.; Powers, J.S.; Pugh, T.A.M.; Ruthrof, K.X.; et al. Climate Change Risks to Global Forest Health: Emergence of Unexpected Events of Elevated Tree Mortality Worldwide. Annu. Rev. Plant Biol. 2022, 73, 673–702. [Google Scholar] [CrossRef]
- Mansoor, S.; Farooq, I.; Kachroo, M.M.; Mahmoud, A.E.D.; Fawzy, M.; Popescu, S.M.; Alyemeni, M.N.; Sonne, C.; Rinklebe, J.; Ahmad, P. Elevation in Wildfire Frequencies with Respect to the Climate Change. J. Environ. Manag. 2022, 301, 113769. [Google Scholar] [CrossRef]
- Forzieri, G.; Dakos, V.; McDowell, N.G.; Ramdane, A.; Cescatti, A. Emerging Signals of Declining Forest Resilience under Climate Change. Nature 2022, 608, 534–539. [Google Scholar] [CrossRef]
- Nunes, L.J.R.; Meireles, C.I.R.; Gomes, C.J.P.; Ribeiro, N.M.C.A. The Impact of Climate Change on Forest Development: A Sustainable Approach to Management Models Applied to Mediterranean-Type Climate Regions. Plants 2021, 11, 69. [Google Scholar] [CrossRef]
- Prichard, S.J.; Hessburg, P.F.; Hagmann, R.K.; Povak, N.A.; Dobrowski, S.Z.; Hurteau, M.D.; Kane, V.R.; Keane, R.E.; Kobziar, L.N.; Kolden, C.A.; et al. Adapting Western North American Forests to Climate Change and Wildfires: 10 Common Questions. Ecol. Appl. 2021, 31, e02433. [Google Scholar] [CrossRef]
- Leifsson, C.; Buras, A.; Rammig, A.; Zang, C. Changing Climate Sensitivity of Secondary Growth Following Extreme Drought Events in Forest Ecosystems: A Global Analysis. Environ. Res. Lett. 2023, 18, 014021. [Google Scholar] [CrossRef]
- Jarzyna, K. Climatic Hazards for Native Tree Species in Poland with Special Regards to Silver Fir (Abies alba Mill.) and European Beech (Fagus sylvatica L.). Theor. Appl. Clim. 2021, 144, 581–591. [Google Scholar] [CrossRef]
- Hylander, K.; Greiser, C.; Christiansen, D.M.; Koelemeijer, I.A. Climate Adaptation of Biodiversity Conservation in Managed Forest Landscapes. Conserv. Biol. 2022, 36, e13847. [Google Scholar] [CrossRef]
- Tekin, O.; Cetin, M.; Varol, T.; Ozel, H.B.; Sevik, H.; Zeren Cetin, I. Altitudinal Migration of Species of Fir (Abies Spp.) in Adaptation to Climate Change. Water Air Soil. Pollut. 2022, 233, 385. [Google Scholar] [CrossRef]
- European Environment Agency Forestry. Available online: https://climate-adapt.eea.europa.eu/en/eu-adaptation-policy/sector-policies/forestry (accessed on 15 January 2025).
- Leites, L.; Benito Garzón, M. Forest Tree Species Adaptation to Climate across Biomes: Building on the Legacy of Ecological Genetics to Anticipate Responses to Climate Change. Glob. Change Biol. 2023, 29, 4711–4730. [Google Scholar] [CrossRef]
- Palik, B.J.; Clark, P.W.; D′Amato, A.W.; Swanston, C.; Nagel, L. Operationalizing Forest-assisted Migration in the Context of Climate Change Adaptation: Examples from the Eastern USA. Ecosphere 2022, 13, e4260. [Google Scholar] [CrossRef]
- Miszuk, B. Climate Water Balance in the Warm Half-Year and Its Circulation Conditions in the Sudetes Mountains and Their Foreland (Poland and Czechia). Water 2023, 15, 795. [Google Scholar] [CrossRef]
- Bruchwald, A.; Dmyterko, E.; Chojnacka Ożga, L.; Sułkowska, M.; Wrzesiński, P. Characteristics of the Process of Tree Mortality Occurring in the Polish Sudetes Mts. Folia For. Pol. 2024, 66, 347–358. [Google Scholar] [CrossRef]
- Nicolescu, V.-N.; Mason, W.L.; Bastien, J.-C.; Vor, T.; Petkova, K.; Podrázský, V.; Đodan, M.; Perić, S.; La Porta, N.; Brus, R.; et al. Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) in Europe: An Overview of Management Practices. J. For. Res. 2023, 34, 871–888. [Google Scholar] [CrossRef]
- Guziejko, A.; Przybylski, P.; Jastrzębowski, S.; Mohytych, V.; Klisz, M. Genetic Variability of Pseudotsuga menziesii (Mirb.) Franco in Northern Poland. Sylwan 2024, 168, 274–286. [Google Scholar] [CrossRef]
- Damszel, M.; Szmidla, H.; Sikora, K.; Młodzińska, A.; Piętka, S.; Sierota, Z. Mycobiota of Fine Roots of Pseudotsuga menziesii Introduced to the Native Forest Environment. Forests 2021, 12, 1766. [Google Scholar] [CrossRef]
- Bijak, S.; Zastocki, D. Prices of Alien Tree Species Timber against Its Harvest in Poland in Years 2013–2018. Sylwan 2021, 165, 101–108. [Google Scholar] [CrossRef]
- Stangler, D.F.; Miller, T.W.; Honer, H.; Larysch, E.; Puhlmann, H.; Seifert, T.; Kahle, H.-P. Multivariate Drought Stress Response of Norway Spruce, Silver Fir and Douglas Fir along Elevational Gradients in Southwestern Germany. Front. Ecol. Evol. 2022, 10, 907492. [Google Scholar] [CrossRef]
- Chakraborty, D.; Schueler, S.; Lexer, M.J.; Wang, T. Genetic Trials Improve the Transfer of Douglas-fir Distribution Models across Continents. Ecography 2019, 42, 88–101. [Google Scholar] [CrossRef]
- Niemczyk, M.; Chmura, D.J.; Socha, J.; Wojda, T.; Mroczek, P.; Gil, W.; Thomas, B.R. How Geographic and Climatic Factors Affect the Adaptation of Douglas-Fir Provenances to the Temperate Continental Climate Zone in Europe. Eur. J. For. Res. 2021, 140, 1341–1361. [Google Scholar] [CrossRef]
- Jastrzębowski, S.; Ukalska, J.; Guziejko, A.; Puchałka, R. Current and Predicted Future Winter Warm Spells Would Affect Douglas Fir (Pseudotsuga menziesii (Mirb.), Franco) Seeds in the Early Stage of Germination More Than in the Late Stage of Germination. Forests 2021, 12, 796. [Google Scholar] [CrossRef]
- Ng, C.W.W.; Guo, H.; Ni, J.; Zhang, Q.; Chen, Z. Effects of Soil–Plant-Biochar Interactions on Water Retention and Slope Stability under Various Rainfall Patterns. Landslides 2022, 19, 1379–1390. [Google Scholar] [CrossRef]
- Panagea, I.S.; Berti, A.; Čermak, P.; Diels, J.; Elsen, A.; Kusá, H.; Piccoli, I.; Poesen, J.; Stoate, C.; Tits, M.; et al. Soil Water Retention as Affected by Management Induced Changes of Soil Organic Carbon: Analysis of Long-Term Experiments in Europe. Land 2021, 10, 1362. [Google Scholar] [CrossRef]
- Kang, M.W.; Yibeltal, M.; Kim, Y.H.; Oh, S.J.; Lee, J.C.; Kwon, E.E.; Lee, S.S. Enhancement of Soil Physical Properties and Soil Water Retention with Biochar-Based Soil Amendments. Sci. Total Environ. 2022, 836, 155746. [Google Scholar] [CrossRef] [PubMed]
- Hewelke, E.; Zaniewski, P.T.; Pędziwiatr, A.; Gozdowski, D.; Górska, E.B. The Relations between Soil Hydrophobicity and Vegetation in Abandoned Arable Fields on Sandy Soil. Biologia 2024, 80, 1115–1123. [Google Scholar] [CrossRef]
- Leelamanie, D.A.L.; Karube, J.; Yoshida, A. Characterizing Water Repellency Indices: Contact Angle and Water Drop Penetration Time of Hydrophobized Sand. Soil Sci. Plant Nutr. 2008, 54, 179–187. [Google Scholar] [CrossRef]
- Smettem, K.R.J.; Rye, C.; Henry, D.J.; Sochacki, S.J.; Harper, R.J. Soil Water Repellency and the Five Spheres of Influence: A Review of Mechanisms, Measurement and Ecological Implications. Sci. Total Environ. 2021, 787, 147429. [Google Scholar] [CrossRef]
- Popović, Z.; Cerdà, A. Soil Water Repellency and Plant Cover: A State-of-Knowledge Review. CATENA 2023, 229, 107213. [Google Scholar] [CrossRef]
- Sándor, R.; Iovino, M.; Lichner, L.; Alagna, V.; Forster, D.; Fraser, M.; Kollár, J.; Šurda, P.; Nagy, V.; Szabó, A.; et al. Impact of Climate, Soil Properties and Grassland Cover on Soil Water Repellency. Geoderma 2021, 383, 114780. [Google Scholar] [CrossRef]
- Falkowski, T. The Polish lowland rivers valleys geological structure and its relationship with channel crossing conditions. J. KONBiN 2022, 52, 175–186. [Google Scholar] [CrossRef]
- Kawałko, D.; Kaszubkiewicz, J.; Jezierski, P. Morphology and Selected Properties of Alluvial Soils in the Odra River Valley, SW Poland. Soil Sci. Annu. 2022, 73, 156062. [Google Scholar] [CrossRef]
- Doerr, S.H.; Shakesby, R.A.; Walsh, R.P.D. Soil Water Repellency: Its Causes, Characteristics and Hydro-Geomorphological Significance. Earth-Sci. Rev. 2000, 51, 33–65. [Google Scholar] [CrossRef]
- Dekker, L.W.; Ritsema, C.J. How Water Moves in a Waterrepellent Sandy Soil: 1. Potential and Actual Water Repellency. Water Resour. Res. 1994, 30, 2507–2517. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2024. [Google Scholar]
- Appleby, S.M.; Balkenhol, N. Douglas Fir and Norway Spruce Have Similar Effects on Small Mammal Density, but Not Survival, in Central European Managed Forests. Mamm. Biol. 2024, 104, 25–39. [Google Scholar] [CrossRef]
- Reyer, C.; Lasch, P.; Mohren, G.M.J.; Sterck, F.J. Inter-Specific Competition in Mixed Forests of Douglas-Fir (Pseudotsuga menziesii) and Common Beech (Fagus sylvatica) under Climate Change—A Model-Based Analysis. Ann. For. Sci. 2010, 67, 805. [Google Scholar] [CrossRef]
- Podrázský, V.; Kupka, I.; Prknová, H. Substitution of Norway Spruce for Douglas-Fir: Changes of Soil Microbial Activities as Climate Change Induced Shift in Species Composition—A Case Study. Cent. Eur. For. J. 2020, 66, 71–77. [Google Scholar] [CrossRef]
- Prietzel, J.; Weis, W.; Bachmann, S.; Kern, M. Effects of Douglas Fir Cultivation in German Forests on Soil Seepage Water Quantity and Quality. For. Ecol. Manag. 2023, 546, 121379. [Google Scholar] [CrossRef]
- Eberhard, B.R.; Eckhart, T.; Hasenauer, H. Evaluating Strategies for the Management of Douglas-Fir in Central Europe. Forests 2021, 12, 1040. [Google Scholar] [CrossRef]
- Berg, B.; Lönn, M.; Ni, X.; Sun, T.; Dong, L.; Gaitnieks, T.; Virzo De Santo, A.; Johansson, M.-B. Decomposition Rates in Late Stages of Scots Pine and Norway Spruce Needle Litter: Influence of Nutrients and Substrate Properties over a Climate Gradient. For. Ecol. Manag. 2022, 522, 120452. [Google Scholar] [CrossRef]
- Levanič, T.; Štraus, H. Effects of Climate on Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) Growth Southeast of the European Alps. Plants 2022, 11, 1571. [Google Scholar] [CrossRef]
- Staszkiewicz, S. Lasy Daglezji (Pseudotsuga menziesii) w Północno-Zachodnich Stanach USA. Sylwan 1968, 12. [Google Scholar]
- Leuschner, C.; Meinzer, F.C. Drought Resistance and Drought Adaptation of Douglas-Fir (Pseudotsuga menziesii)—A Review. Perspect. Plant Ecol. Evol. Syst. 2024, 65, 125829. [Google Scholar] [CrossRef]
- Domec, J.-C.; Warren, J.M.; Meinzer, F.C.; Brooks, J.R.; Coulombe, R. Native Root Xylem Embolism and Stomatal Closure in Stands of Douglas-Fir and Ponderosa Pine: Mitigation by Hydraulic Redistribution. Oecologia 2004, 141, 7–16. [Google Scholar] [CrossRef]
- Gorzelak, A. Odnawianie Lasu w Reglu Górnym Sudetów Zachodnich. Sylwan 1993, 137, 5–16. [Google Scholar]
- Walendzik, R. Degradacja Gleb Leśnych w Sudetach Zachodnich i Próby Jej Ograniczenia. Sylwan 1993, 137, 29–38. [Google Scholar]
- Thomas, F.M.; Rzepecki, A.; Werner, W. Non-Native Douglas Fir (Pseudotsuga menziesii) in Central Europe: Ecology, Performance and Nature Conservation. For. Ecol. Manag. 2022, 506, 119956. [Google Scholar] [CrossRef]
- Schmid, M.; Pautasso, M.; Holdenrieder, O. Ecological Consequences of Douglas Fir (Pseudotsuga menziesii) Cultivation in Europe. Eur. J. For. Res. 2014, 133, 13–29. [Google Scholar] [CrossRef]
- Mihai, G.; Curtu, A.-L.; Alexandru, A.-M.; Nita, I.-A.; Ciocîrlan, E.; Birsan, M.-V. Growth and Adaptive Capacity of Douglas Fir Genetic Resources from Western Romania under Climate Change. Forests 2022, 13, 805. [Google Scholar] [CrossRef]
- Fuchs, J.M.; Hittenbeck, A.; Brandl, S.; Schmidt, M.; Paul, C. Adaptation Strategies for Spruce Forests—Economic Potential of Bark Beetle Management and Douglas Fir Cultivation in Future Tree Species Portfolios. For. Int. J. For. Res. 2022, 95, 229–246. [Google Scholar] [CrossRef]
- Jactel, H.; Gritti, E.S.; Drossler, L.; Forrester, D.I.; Mason, W.L.; Morin, X.; Pretzsch, H.; Castagneyrol, B. Positive Biodiversity–Productivity Relationships in Forests: Climate Matters. Biol. Lett. 2018, 14, 20170747. [Google Scholar] [CrossRef]
- Heinrichs, S.; Dölle, M.; Vor, T.; Balcar, P.; Schmidt, W. Understory Vegetation Dynamics in Non-Native Douglas Fir Forests after Management Abandonment—A Case Study in Two Strict Forest Reserves in Southwest Germany. Diversity 2022, 14, 795. [Google Scholar] [CrossRef]
- Lasota, J.; Błońska, E.; Wanic, T.; Klamerus-Iwan, A.; Więcek, Z. Charakterystyka warunków glebowych gatunków introdukowanych na przykładzie Nadleśnictwa Prószków. Stud. I Mater. Cent. Edukac. Przyr. Leśnej 2012, 14, 121–129. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).