

Multi-Annual Dendroclimatic Patterns for the Desert National Wildlife Refuge, Southern Nevada, USA



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Dendrochronological Analysis
2.3. Dendroclimatic Reconstruction
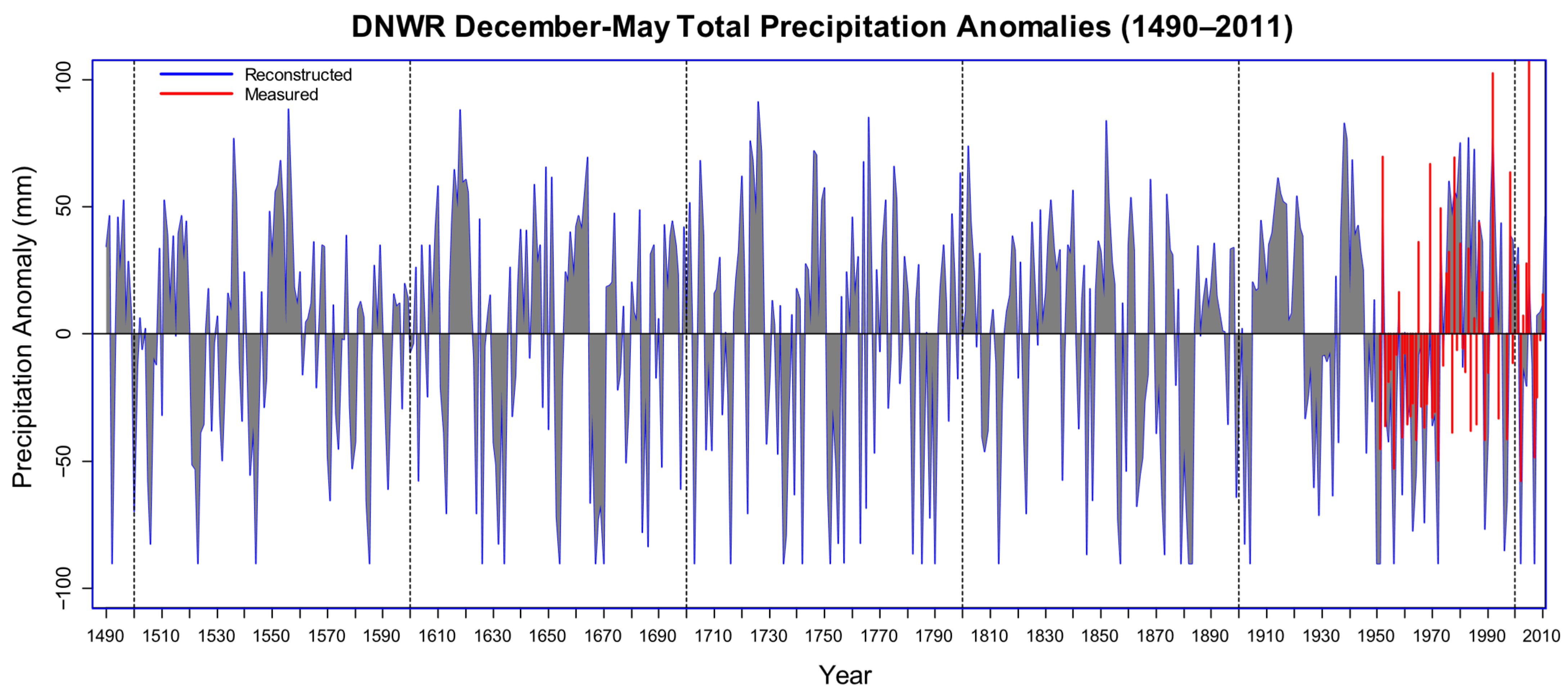
3. Results
3.1. Climate and Tree Rings
3.2. Long-Term Dendroclimatic Patterns
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grayson, D.K. The Great Basin: A Natural Prehistory, Revised and Expanded ed.; University of California Press: Berkeley, CA, USA, 2011. [Google Scholar]
- Osmond, C.B.; Pitelka, L.F.; Hidy, G.M. (Eds.) Plant Biology of the Basin and Range; Springer: Berlin/Heidelberg, Germany, 1990; Volume 80. [Google Scholar]
- Dominguez, F.; Villegas, J.C.; Breshears, D.D. Spatial extent of the North American Monsoon: Increased crossregional linkages via atmospheric pathways. Geophys. Res. Let. 2009, 36, L07401. [Google Scholar] [CrossRef]
- Warshall, P. The Madrean sky island archipelago: A planetary overview. In Biodiversity and Management of the Madrean Archipelago: The Sky Islands of the Southwestern United States and Northwestern Mexico; DeBano, L.F., Ffolliott, P.F., Ortega-Rubio, A., Gottfried, G.J., Hamre, R.H., Edminster, C.B., Eds.; US Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station: Fort Collins, CO, USA, 1995; pp. 6–18. [Google Scholar]
- Charlet, D.A. Distribution patterns of Great Basin conifers: Implications of extinction and immigration. Aliso 2007, 24, 31–61. [Google Scholar] [CrossRef]
- Potter, K.M.; Hipkins, V.D.; Mahalovich, M.F.; Means, R.E. Mitochondrial DNA haplotype distribution patterns in Pinus ponderosa (Pinaceae): Range-wide evolutionary history and implications for conservation. Am. J. Bot. 2013, 100, 1562–1579. [Google Scholar] [CrossRef] [PubMed]
- Willyard, A.; Gernandt, D.S.; Potter, K.; Hipkins, V.; Marquardt, P.; Mahalovich, M.F.; Langer, S.K.; Telewski, F.W.; Cooper, B.; Douglas, C.; et al. Pinus ponderosa: A checkered past obscured four species. Am. J. Bot. 2017, 104, 161–181. [Google Scholar] [CrossRef]
- Allen, C.D.; Savage, M.; Falk, D.A.; Suckling, K.F.; Swetnam, T.W.; Schulke, T.; Stacey, P.B.; Morgan, P.; Hoffman, M.; Klingel, J.T. Ecological restoration of southwestern ponderosa pine ecosystems: A broad perspective. Ecol. Appl. 2002, 12, 1418–1433. [Google Scholar] [CrossRef]
- Margolis, E.Q.; Guiterman, C.H.; Chavardès, R.D.; Coop, J.D.; Copes-Gerbitz, K.; Dawe, D.A.; Falk, D.A.; Johnston, J.D.; Larson, E.; Li, H.; et al. The North American tree-ring fire-scar network. Ecosphere 2022, 13, e4159. [Google Scholar] [CrossRef]
- Parks, S.A.; Guiterman, C.H.; Margolis, E.Q.; Lonergan, M.; Whitman, E.; Abatzoglou, J.T.; Falk, D.A.; Johnston, J.D.; Daniels, L.D.; Lafon, C.W.; et al. A fire deficit persists across diverse North American forests despite recent increases in area burned. Nat. Commun. 2025, 16, 1493. [Google Scholar] [CrossRef]
- Kilpatrick, M.; Biondi, F.; Strachan, S.; Sibold, J.S. Fire history of mixed conifer ecosystems in the Great Basin/Mojave Deserts transition zone, Nevada, USA. Trees 2013, 27, 1789–1803. [Google Scholar] [CrossRef]
- Biondi, F.; Jamieson, L.P.; Strachan, S.; Sibold, J. Dendroecological testing of the pyroclimatic hypothesis in the central Great Basin, Nevada, USA. Ecosphere 2011, 2, 1–20. [Google Scholar] [CrossRef]
- Kilpatrick, M.; Roberts, J.; Biondi, F. Fire history of an old-growth ponderosa pine stand in the Sheep Range, Desert National Wildlife Refuge, Nevada, USA. Tree Ring Res. 2019, 75, 127–138. [Google Scholar] [CrossRef]
- Walker, R.B.; Coop, J.D.; Parks, S.A.; Trader, L. Fire regimes approaching historic norms reduce wildfire-facilitated conversion from forest to non-forest. Ecosphere 2018, 9, e02182. [Google Scholar] [CrossRef]
- Stephens, S.L.; Lydersen, J.M.; Collins, B.M.; Fry, D.L.; Meyer, M.D. Historical and current landscape-scale ponderosa pine and mixed conifer forest structure in the Southern Sierra Nevada. Ecosphere 2015, 6, 1–63. [Google Scholar] [CrossRef]
- McEvoy, D.J.; Mejia, J.F.; Huntington, J.L. Use of an observation network in the Great Basin to evaluate gridded climate data. J. Hydrometeorol. 2014, 15, 1913–1931. [Google Scholar] [CrossRef]
- Vose, R.S.; Applequist, S.; Squires, M.; Durre, I.; Menne, M.J.; Williams, C.N.; Fenimore, C.; Gleason, K.; Arndt, D. Improved historical temperature and precipitation time series for U.S. Climate Divisions. J. Appl. Meteorol. Clim. 2014, 53, 1232–1251. [Google Scholar] [CrossRef]
- US Western Regional Climate Center. Climate Data for DESERT NATL WL RANGE, NEVADA (262243). Available online: https://wrcc.dri.edu/cgi-bin/cliMAIN.pl?nv2243 (accessed on 14 April 2025).
- van Buuren, S.; Groothuis-Oudshoorn, K. Mice: Multivariate Imputation by Chained Equations in R. J. Stat. Softw. 2011, 45, 1–67. [Google Scholar] [CrossRef]
- Daly, C.; Halbleib, M.; Smith, J.I.; Gibson, W.P.; Doggett, M.K.; Taylor, G.H.; Curtis, J.; Pasteris, P.P. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int. J. Climatol. 2008, 28, 2031–2064. [Google Scholar] [CrossRef]
- Diaz, H.F.; Wahl, E.R. Recent California water year precipitation deficits: A 440-year perspective. J. Clim. 2015, 28, 4637–4652. [Google Scholar] [CrossRef]
- Hipel, K.W.; McLeod, A.I. Time Series Modelling of Water Resources and Environmental Systems; Elsevier Science: Amsterdam, The Netherlands, 1994. [Google Scholar]
- McLeod, A.I. R Foundation for Statistical Computing: CRAN, Package ‘Kendall’. Available online: https://cran.r-project.org/web/packages/Kendall/index.html (accessed on 8 July 2025).
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating, Reprint of the 1968 ed.; University of Arizona Press: Tucson, AZ, USA, 1996. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree Ring Res. 2001, 57, 205–221. [Google Scholar]
- Bunn, A.G. Statistical and visual crossdating in R using the dplR library. Dendrochronologia 2010, 28, 251–258. [Google Scholar] [CrossRef]
- Speer, J.H. Fundamentals of Tree-Ring Research; University of Arizona Press: Tuscon, AZ, USA, 2010. [Google Scholar]
- Cropper, J.P.; Ferguson, C.W. Sawmill Canyon—ABCO—ITRDB NV506; NOAA/WDS Paleoclimatology; NOAA National Centers for Environmental Information: Silver Spring, MD, USA, 1978. [CrossRef]
- Cook, E.R.; Peters, K. The smoothing spline: A new approach to standardizing forest interior tree-ring width series for dendroclimatic studies. Tree Ring Bull. 1981, 41, 45–53. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Clim. Appl. Meteorol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Biondi, F.; Qeadan, F. Inequality in paleorecords. Ecology 2008, 89, 1056–1067. [Google Scholar] [CrossRef] [PubMed]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Kim, S. Ppcor: An R package for a fast calculation to semi-partial correlation coefficients. Commun. Stat. Appl. Methods 2015, 22, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Zang, C.; Biondi, F. treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Helsel, D.R.; Hirsch, R.M.; Ryberg, K.R.; Archfield, S.A.; Gilroy, E.J. Statistical Methods in Water Resources; USGS: Reston, VA, USA, 2020; p. 484.
- Salas, J.D.; Raynal, J.A.; Tarawneh, Z.S.; Lee, T.S.; Frevert, D.K.; Fulp, T.J. Extending Short Records of Hydrologic Data. In Hydrology and Hydraulics; Singh, V.P., Ed.; Water Resources Publications, LLC.: Highlands Ranch, CO, USA, 2008; pp. 717–760. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Krause, P.; Boyle, D.P.; Bäse, F. Comparison of different efficiency criteria for hydrological model assessment. Adv. Geosci. 2005, 5, 89–97. [Google Scholar] [CrossRef]
- Nash, J.E.; Sutcliffe, J.V. River flow forecasting through conceptual models, Part I—A discussion of principles. J. Hydrol. 1970, 10, 282–290. [Google Scholar] [CrossRef]
- Draper, N.R.; Smith, H. Applied Regression Analysis, 3rd ed.; Wiley: New York, NJ, USA, 1998. [Google Scholar]
- Salas, J.D. Analysis and modeling of hydrologic time series. In Handbook of Hydrology; Maidment, D.R., Ed.; McGraw-Hill: New York, NY, USA, 1992; pp. 19.01–19.72. [Google Scholar]
- Biondi, F.; Kozubowski, T.J.; Panorska, A.K.; Saito, L. A new stochastic model of episode peak and duration for eco-hydro-climatic applications. Ecol. Model. 2008, 211, 383–395. [Google Scholar] [CrossRef]
- Kozubowski, T.J.; Panorska, A.K. A mixed bivariate distribution connected with geometric maxima of exponential variables. Commun. Stat. Theory Methods 2008, 37, 2903–2923. [Google Scholar] [CrossRef]
- Kozubowski, T.J.; Panorska, A.K. A mixed bivariate distribution with exponential and geometric marginals. J. Stat. Plan. Inference 2005, 134, 501–520. [Google Scholar] [CrossRef]
- Guttman, N.B.; Quayle, R.G. A historical perspective of US climate divisions. Bull. Am. Meteorol. Soc. 1996, 77, 293–303. [Google Scholar] [CrossRef]
- Cook, B.I.; Seager, R.; Smerdon, J.E. The worst North American drought year of the last millennium: 1934. Geophys. Res. Lett. 2014, 41, 7298–7305. [Google Scholar] [CrossRef]
- Diaz, H.F. Drought in the United States: Some aspects of major dry and wet periods in the contiguous united states, 1895–1981. J. Clim. Appl. Meteorol. 1983, 22, 3–16. [Google Scholar] [CrossRef]
- Cook, B.I.; Seager, R.; Miller, R.L. On the causes and dynamics of the early twentieth-century North American pluvial. J. Clim. 2011, 24, 5043–5060. [Google Scholar] [CrossRef]
- Woodhouse, C.A.; Kunkel, K.E.; Easterling, D.R.; Cook, E.R. The twentieth-century pluvial in the western United States. Geophys. Res. Let. 2005, 32, L07701. [Google Scholar] [CrossRef]
- Biondi, F.; Meko, D.M. Long-term hydroclimatic patterns in the Truckee-Carson Basin of the eastern Sierra Nevada, USA. Water Resour. Res. 2019, 55, 5559–5574. [Google Scholar] [CrossRef]
- Gottfried, G.J.; Gebow, B.S.; Eskew, L.G.; Edminster, C.B. (Eds.) Connecting Mountain Islands and Desert Seas: Biodiversity and Management of the Madrean Archipelago II; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005.
- Ellison, A.M.; Bank, M.S.; Clinton, B.D.; Colburn, E.A.; Elliott, K.; Ford, C.R.; Foster, D.R.; Kloeppel, B.D.; Knoepp, J.D.; Lovett, G.M.; et al. Loss of foundation species: Consequences for the structure and dynamics of forested ecosystems. Front. Ecol. Environ. 2005, 3, 479–486. [Google Scholar] [CrossRef]
- Morino, K.; Minor, R.L.; Barron-Gafford, G.A.; Brown, P.M.; Hughes, M.K. Bimodal cambial activity and false-ring formation in conifers under a monsoon climate. Tree Physiol. 2021, 41, 1893–1905. [Google Scholar] [CrossRef]
- Griffin, R.D.; Woodhouse, C.A.; Meko, D.M.; Stahle, D.W.; Faulstich, H.L.; Carrillo, C.; Touchan, R.; Castro, C.L.; Leavitt, S.W. North American monsoon precipitation reconstructed from tree-ring latewood. Geophys. Res. Lett. 2013, 40, 954–958. [Google Scholar] [CrossRef]
- Buras, A. A comment on the expressed population signal. Dendrochronologia 2017, 44, 130–132. [Google Scholar] [CrossRef]
- Oyler, J.W.; Dobrowski, S.Z.; Ballantyne, A.P.; Klene, A.E.; Running, S.W. Artificial amplification of warming trends across the mountains of the western United States. Geophys. Res. Lett. 2015, 42, 153–161. [Google Scholar] [CrossRef]
- Garfin, G.; Jardine, A.; Merideth, R.; Black, M.; LeRoy, S. (Eds.) Assessment of Climate Change in the Southwest United States: A Report Prepared for the National Climate Assessment; Island Press: Washington, DC, USA, 2013. [Google Scholar]
- Pederson, N.; Bell, A.R.; Knight, T.A.; Leland, C.; Malcomb, N.; Anchukaitis, K.J.; Tackett, K.; Scheff, J.; Brice, A.; Catron, B.; et al. A long-term perspective on a modern drought in the American Southeast. Environ. Res. Lett. 2012, 7, 014034. [Google Scholar] [CrossRef]
- Pielke, R.A., Sr.; Wilby, R.L. Regional climate downscaling: What’s the point? Eos Trans. Am. Geophys. Union 2012, 93, 52–53. [Google Scholar] [CrossRef]
- Hirsch, R.M. A comparison of four streamflow record extension techniques. Water Resour. Res. 1982, 18, 1081–1088. [Google Scholar] [CrossRef]
- Robeson, S.M.; Maxwell, J.T.; Ficklin, D.L. Bias correction of paleoclimatic reconstructions: A new look at 1,200+ years of Upper Colorado River Flow. Geophys. Res. Lett. 2020, 47, e2019GL086689. [Google Scholar] [CrossRef]
- Biondi, F. Dendroclimatic reconstruction at km-scale grid points: A case study from the Great Basin of north America. J. Hydrometeorol. 2014, 15, 891–906. [Google Scholar] [CrossRef]
- Kupfer, J.A.; Balmat, J.; Smith, J.L. Shifts in the potential distribution of sky island plant communities in response to climate change. In Connecting Mountain Islands and Desert Seas: Biodiversity and Management of the Madrean Archipelago II; Gottfried, G.J., Gebow, B.S., Eskew, L.G., Edminster, C.B., Eds.; US Department of Agriculture, Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2005; pp. 485–490. [Google Scholar]
- Thomas, K.A.; Stauffer, B.A.; Jarchow, C.J. Decoupling of species and plant communities of the U.S. Southwest: A CCSM4 climate scenario example. Ecosphere 2023, 14, e4414. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Lenoir, J.; Chen, I.-C. Limited evidence for range shift–driven extinction in mountain biota. Science 2025, 388, 741–747. [Google Scholar] [CrossRef]
- Shriver, R.K.; Pletcher, E.; Biondi, F.; Urza, A.K.; Weisberg, P.J. Long-term tree population growth can predict woody encroachment patterns. Proc. Nat. Acad. Sci. USA 2025, 122, e2424096122. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Site | First Year | Last Year | Dated Trees/ Cores | MSL (Years) | Dated LAR/ Total | Period with N ≥ 3 | Length (Years) | MISC | G | St. Dev. | A1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| NevCAN | 1629 | 2013 | 12/20 | 276 | 354/5512 | 1652–2013 | 362 | 0.87 | 0.31 | 0.52 | 0.29 |
| Pine Nut # | 1206 | 2011 | 18/31 | 285 | 593/8828 | 1490–2011 | 522 | 0.85 | 0.28 | 0.47 | 0.29 |
| Mt. Irish ## | 1363 | 2003 | 11/21 | 457 | 155/9596 | 1396–2003 | 608 | 0.87 | 0.17 | 0.30 | 0.27 |
| Clover Mtns. ### | 1470 | 2007 | 27/47 | 314 | 213/14,743 | 1485–2007 | 523 | 0.83 | 0.19 | 0.33 | 0.35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Biondi, F.; Roberts, J. Multi-Annual Dendroclimatic Patterns for the Desert National Wildlife Refuge, Southern Nevada, USA. Forests 2025, 16, 1142. https://doi.org/10.3390/f16071142
Biondi F, Roberts J. Multi-Annual Dendroclimatic Patterns for the Desert National Wildlife Refuge, Southern Nevada, USA. Forests. 2025; 16(7):1142. https://doi.org/10.3390/f16071142
Chicago/Turabian StyleBiondi, Franco, and James Roberts. 2025. "Multi-Annual Dendroclimatic Patterns for the Desert National Wildlife Refuge, Southern Nevada, USA" Forests 16, no. 7: 1142. https://doi.org/10.3390/f16071142
APA StyleBiondi, F., & Roberts, J. (2025). Multi-Annual Dendroclimatic Patterns for the Desert National Wildlife Refuge, Southern Nevada, USA. Forests, 16(7), 1142. https://doi.org/10.3390/f16071142