
Optimization of Traps Used in the Management of Monochamus galloprovincialis (Coleoptera: Cerambycidae), the Insect-Vector of Pinewood Nematode, to Reduce By-Catches of Non-Target Insects


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Assay Location and Time of Experiment
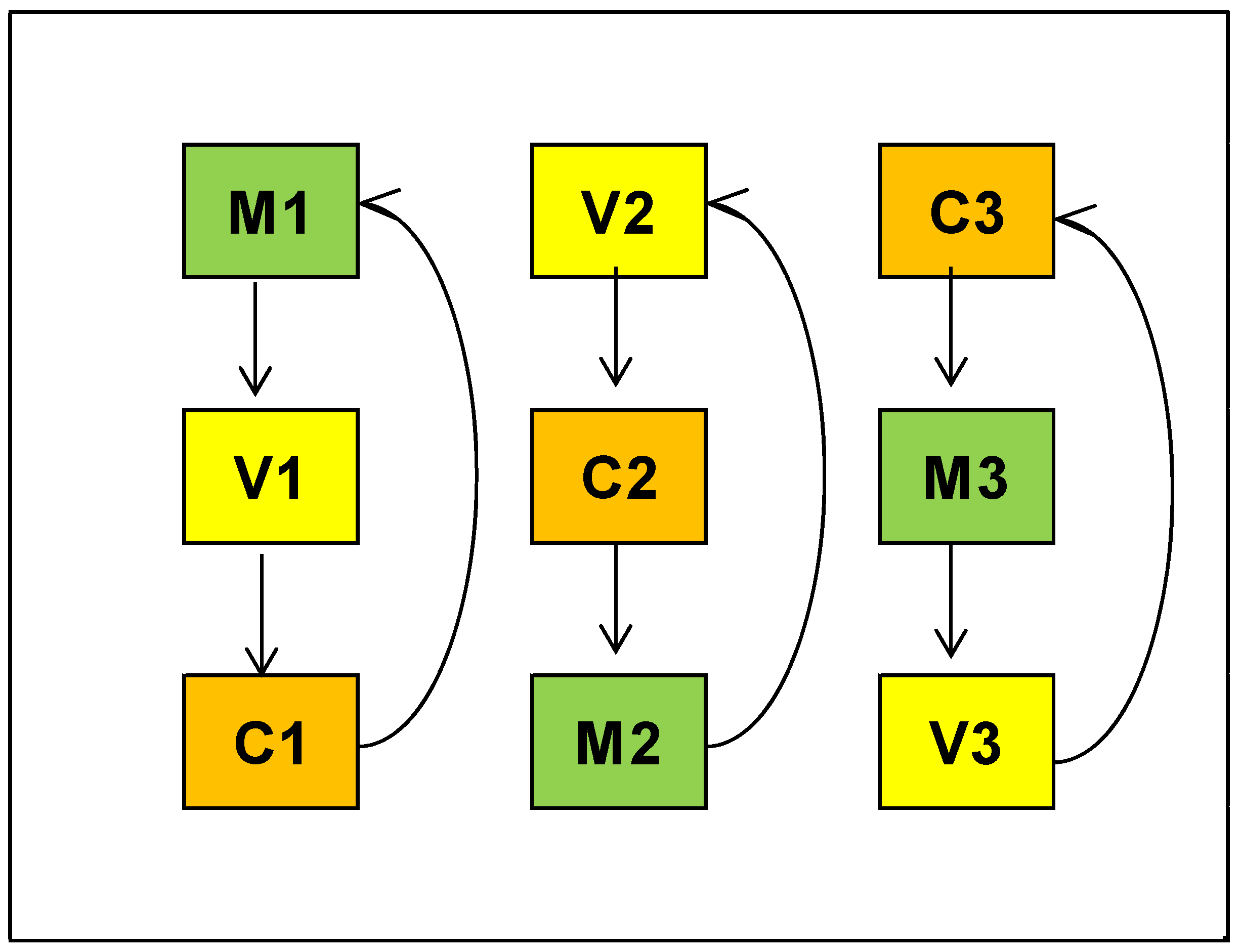
2.2. Experimental Design
2.3. Data Analysis
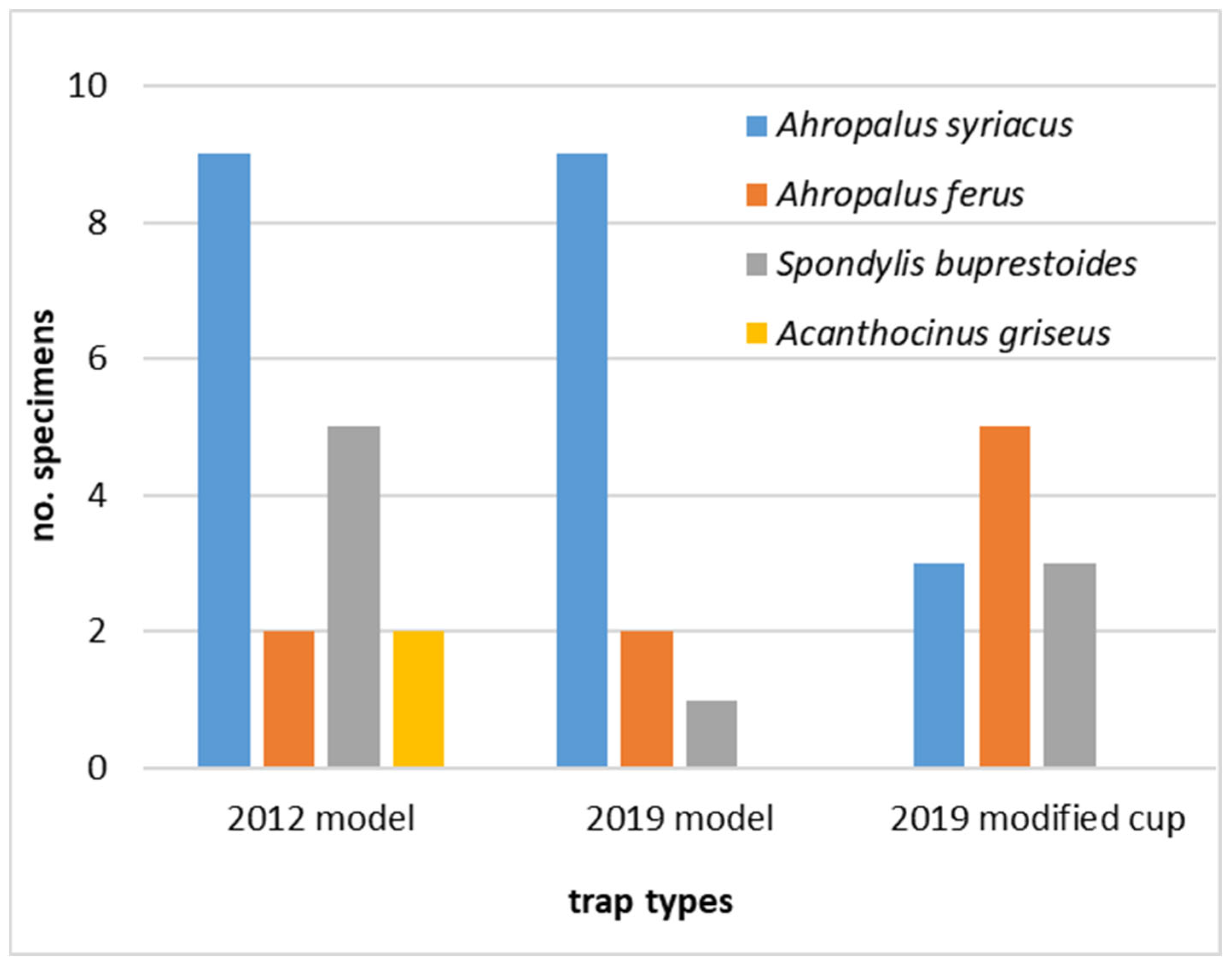
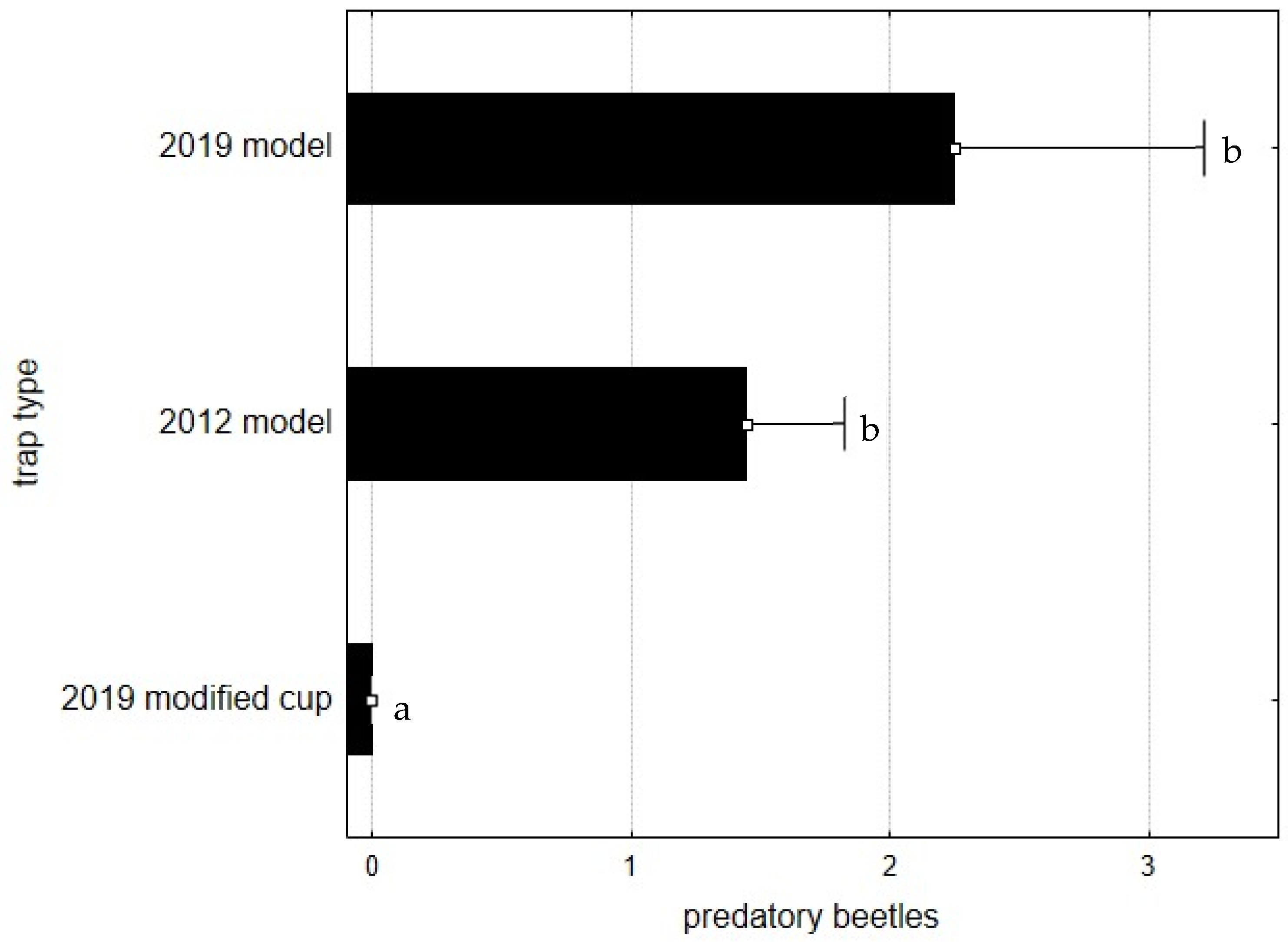
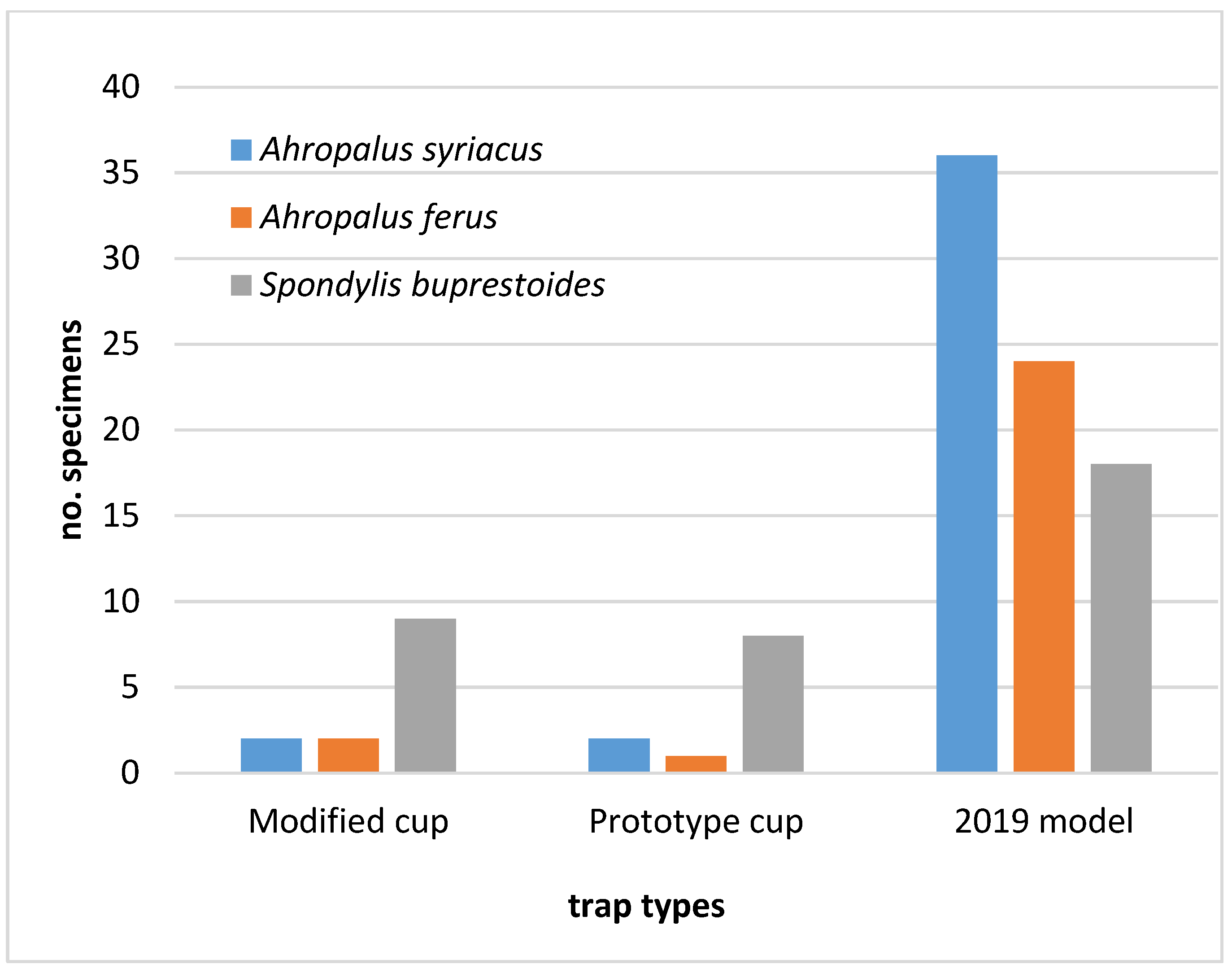
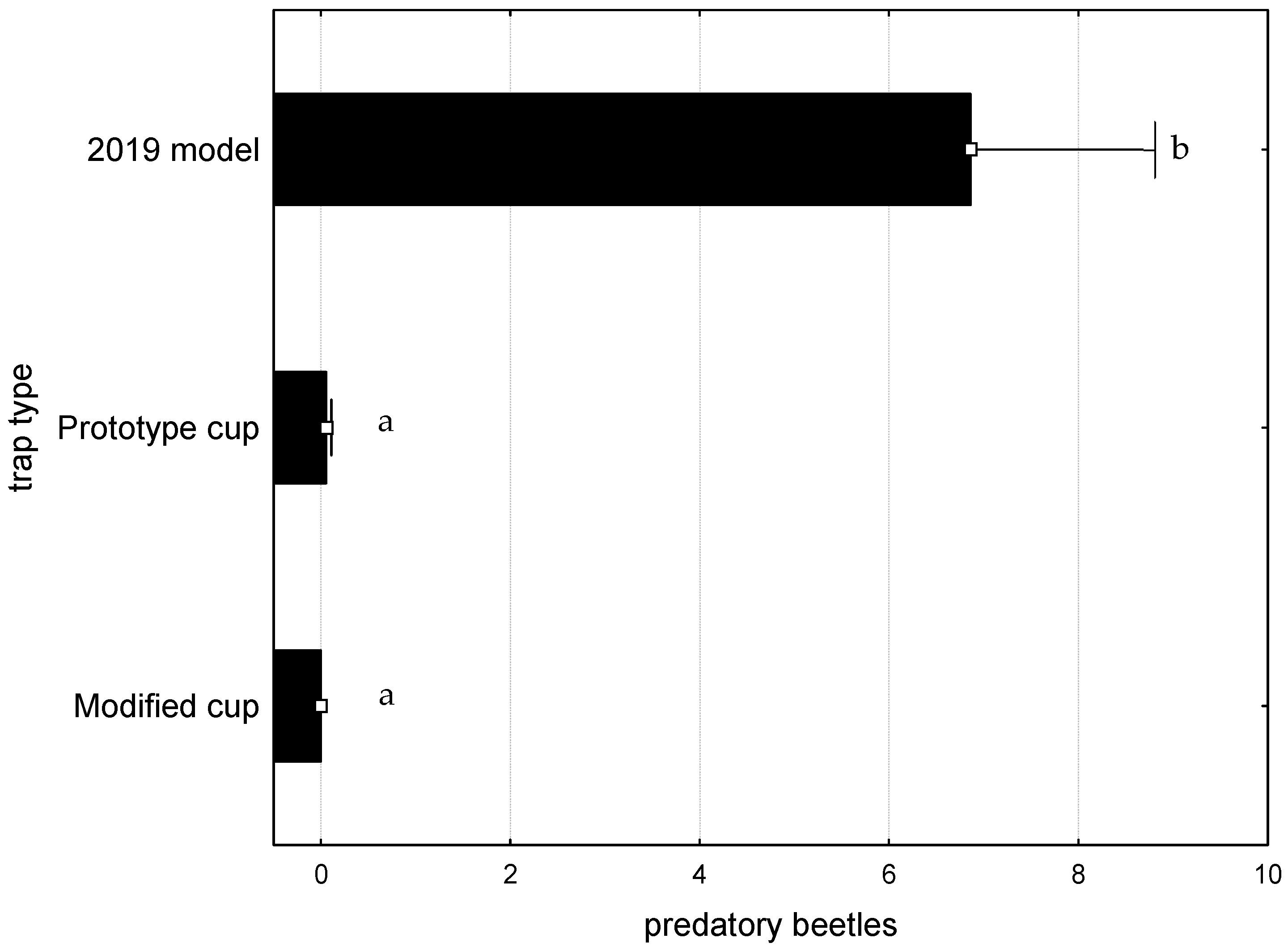
3. Results
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Futai, K. Pine Wilt in Japan: From First Incidence to the Present. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 5–12. [Google Scholar]
- Zhao, B.G. Pine wilt disease in China. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 18–25. [Google Scholar]
- Back, M.A.; Bonifácio, L.; Inácio, M.L.; Mota, M.; Boa, E. Pine wilt disease: A global threat to forestry. Plant Pathol. 2024, 73, 1026–1041. [Google Scholar] [CrossRef]
- EPPO. Bursaphelenchus xylophilus. EPPO Datasheets on Pests Recommended for Regulation. 2025. Available online: https://gd.eppo.int (accessed on 6 May 2025).
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus xylophilus, Causal Agent of Pine Wilt Disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef]
- Alonso, M.; Menéndez, M.; Diaz, R. Constitutive chemical compounds in different tissues of seven pine species and their relationship with susceptibility to pinewood nematode (Bursaphelenchus xylophilus). Environ. Sci. Proc. 2021, 3, 68. [Google Scholar] [CrossRef]
- Fonseca, L.; Cardoso, J.M.S.; Lopes, A.; Pestana, M.; Abreu, F.; Nunes, N.; Mota, M.; Abrantes, I. The pinewood nematode, Bursaphelenchus xylophilus, in Madeira Island. Helminthologia 2012, 49, 96–103. [Google Scholar] [CrossRef]
- Kwon, T.S.; Shin, J.H.; Lim, J.H.; Kim, Y.K.; Lee, E.J. Management of pine wilt disease in Korea through preventative silvicultural control. For. Ecol. Manag. 2011, 261, 562–569. [Google Scholar] [CrossRef]
- Rodrigues, J.M.; Sousa, E.; Abrantes, I. Pine wilt disease historical review. In Pine Wilt Disease in Europe: Biological Interactions and Integrated Management; FNAPF-Federação Nacional das Associações de Proprietários Florestais: Lisbon, Portugal, 2015; pp. 13–32. [Google Scholar]
- Li, M.; Dai, Y.; Wang, Y.; Wang, L.; Sun, S.; Chen, F. New insights into the life history of Monochamus saltuarius (Cerambycidae: Coleoptera) can enhance surveillance strategies for pine wilt disease. J. For. Res. 2021, 32, 2699–2707. [Google Scholar] [CrossRef]
- Ikeda, T.; Oda, K.; Yamane, A.; Enda, N. Volatiles from pine logs as the attractants for the Japanese Pine Sawyer Monochamus alternatus Hope (Coleoptera: Cerambycidae). J. Jap. For. Soc. 1980, 62, 150–152. [Google Scholar]
- Ikeda, T.; Yamane, A.; Enda, N.; Oda, K.; Makihara, H.; Ito, K.; Ôkochi, I. Attractiveness of each volatile components of felled pine trees for Monochamus alternatus (Coleoptera: Cerambycidae). J. Jap. For. Soc. 1986, 68, 15–19. [Google Scholar]
- Sousa, E.; Bonifácio, L.; Naves, P.; Vieira, M. Pine wilt disease management. In Pine Wilt Disease in Europe: Biological Interactions and Integrated Management; FNAPF-Federação Nacional das Associações de Proprietários Florestais: Lisbon, Portugal, 2015; pp. 221–295. [Google Scholar]
- Shin, S.C. Pine wilt disease in Korea. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 26–32. [Google Scholar]
- Lee, H.R.; Lee, S.C.; Lee, D.H.; Jung, M.; Kwon, M.J.; Huh, M.J.; Kim, D.S.; Lee, J.E.; Park, I.K. Identification of aggregation-sex pheromone of the Korean Monochamus alternatus (Coleoptera: Cerambycidae) population, the main vector of Pine Wood Nematode. J. Econ. Entomol. 2018, 111, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Bonifácio, L.; Praias, F.; Sousa, E. Trapping Monochamus galloprovincialis (Coleoptera: Cerambycidae), vector of the pine wood nematode, with pine allelochemicals in Portugal. Silva Lusit. 2012, 20, 39–53. [Google Scholar]
- Holmes, S.B.; MacQuarrie, C.J.K. Chemical control in forest pest management. Can. Entomol. 2016, 148, S270–S295. [Google Scholar] [CrossRef]
- Alvarez, G.; Etxebeste, I.; Gallego, D.; David, G.; Bonifacio, L.; Jactel, H.; Sousa, E.; Pajares, J.A. Optimization of traps for live trapping of Pine Wood Nematode vector Monochamus galloprovincialis. J. Appl. Entomol. 2014, 139, 618–626. [Google Scholar] [CrossRef]
- Jactel, H.; Bonifácio, L.; van Halder, I.; Vétillard, F.; Robinet, C.; David, G. A novel, easy method for estimating pheromone trap attraction range: Application to the pine sawyer beetle Monochamus galloprovincialis. Agric. For. Entomol. 2019, 21, 8–14. [Google Scholar] [CrossRef]
- Pajares, J.A.; Alvarez, G.; Ibeas, F.; Gallego, D.; Hall, D.; Farman, D. Identification and field activity of a male-produced aggregation pheromone in the pine sawyer beetle, Monochamus galloprovincialis. J. Chem. Ecol. 2010, 36, 570–583. [Google Scholar] [CrossRef] [PubMed]
- Pajares, J.A.; Álvarez, G.; Hall, D.R.; Ibarra, N.; Hoch, G.; Halbig, P.; Schroeder, M. Attractants for management of the pine sawyer beetle Monochamus sutor, a potential vector of Bursaphelenchus xylophilus. J. Appl. Entomol. 2016, 141, 97–111. [Google Scholar] [CrossRef]
- Miller, D.R.; Allison, J.D.; Crowe, C.M.; Dickinson, D.M.; Eglitis, A.; Hofstetter, R.W.; Munson, A.S.; Poland, T.M.; Reid, L.S.; Steed, B.E.; et al. Pine sawyers (Coleoptera: Cerambycidae) attracted to α-pinene, monochamol, and ipsenol in North America. J. Econ. Entomol. 2016, 109, 1205–1214. [Google Scholar] [CrossRef]
- Allison, J.D.; McKenney, J.L.; Millar, J.G.; McElfresh, J.S.; Mitchell, R.F.; Hanks, L.M. Response of the woodborers Monochamus carolinensis and Monochamus titillator to known cerambycid pheromones in the presence and absence of the host plant volatile alpha-pinene. Environ. Entomol. 2012, 41, 1587–1596. [Google Scholar] [CrossRef]
- Allison, J.D.; Guignard, Q.; Ochoa, I.; Sousa, E.; Bonifácio, L. Asymmetric semiochemical-mediated interactions of Monochamus spp. (Coleoptera: Cerambycidae) and associated bark beetles in Portugal and Canada. Environ. Entomol. 2024, 54, 46–53. [Google Scholar] [CrossRef]
- Allison, J.D.; Borden, J.H.; McIntosh, R.L.; de Groot, P.; Gries, R. Kairomonal response by four Monochamus species (Coleoptera: Cerambycidae) to bark beetle pheromones. J. Chem. Ecol. 2001, 27, 633–646. [Google Scholar] [CrossRef]
- Allison, J.D.; Morewood, W.D.; Borden, J.H.; Hein, K.E.; Wilson, I.M. Differential bio-activity of Ips and Dendroctonus (Coleoptera: Scolytidae) pheromone components for Monochamus clamator and M. scutellatus (Coleoptera: Cerambycidae). Environ. Entomol. 2003, 32, 23–30. [Google Scholar] [CrossRef]
- De Groot, P.; Nott, W. Response of the whitespotted sawyer beetle, Monochamus scutellatus, and associated woodborers to pheromones of some Ips and Dendroctonus bark beetles. J. Appl. Entomol. 2004, 128, 483–487. [Google Scholar] [CrossRef]
- Pajares, J.A.; Ibeas, F.; Diez, J.J.; Gallego, D. Attractive responses by Monochamus galloprovincialis (Col., Cerambycidae) to host and bark beetle semiochemicals. J. Appl. Entomol. 2004, 128, 633–638. [Google Scholar]
- Pajares, J.A.; Alvarez, G.; Hall, D.R.; Douglas, P.; Centeno, F.; Ibarra, N.; Schroeder, M.; Teale, S.A.; Wang, Z.; Yan, S.; et al. 2-(Undecyloxy)-ethanol is a major component of the male-produced aggregation pheromone of Monochamus sutor. Entomol. Exp. Appl. 2013, 149, 118–127. [Google Scholar] [CrossRef]
- Miller, D.R.; Asaro, C. Ipsenol and Ipsdienol attract Monochamus titillator (Coleoptera: Cerambycidae) and associated large pine woodborers in Southeastern United States. J. Econ. Entomol. 2005, 98, 2033–2040. [Google Scholar] [CrossRef]
- Miller, D.R.; Asaro, C.; Crowe, C.; Duerr, D. Bark beetle pheromones and pine volatiles: Attractant kairomone lure blend for longhorn beetles (Cerambycidae) in pine stands of the Southeastern United States. J. Econ. Entomol. 2011, 104, 1245–1257. [Google Scholar] [CrossRef]
- Miller, D.R.; Crowe, C.M.; Barnes, B.F.; Gandhi, K.J.K.; Duerr, D.A. Attaching lures to multiple-funnel traps targeting saproxylic beetles (Coleoptera) in pine stands: Inside or outside funnels? J. Econ. Entomol. 2013, 106, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Dodds, K.J.; Graber, C.; Stephen, F.M. Facultative intraguild predation by larval Cerambycidae (Coleoptera) on bark beetle larvae (Coleoptera: Scolytidae). Environ. Entomol. 2001, 30, 17–22. [Google Scholar] [CrossRef]
- Jaworski, T.; Plewa, R.; Dziuk, A.; Sukovata, L. Different responses of Monochamus galloprovincialis and three non-target species to trap type, colour, and lubricant treatment. Ann. For. Res. 2022, 65, 31–46. [Google Scholar] [CrossRef]
- Mellin, M.; Vihervuori, L.; Koivula, M.; Velmala, S. Pheromone-based monitoring of invasive alien insects along the border of Finland and Russia—Methods and unintentionally caught species. Balt. For. 2023, 28, 639. [Google Scholar] [CrossRef]
- Tamutis, V.; Zalubas, P. Non-target beetles trapped in Ips typographus L. pheromone traps. Balt. J. Coleopterol. 2001, 1, 65–70. [Google Scholar]
- Galko, J.; Nikolov, C.; Kunca, A.; Vakula, J.; Gubka, A.; Zubrik, M.; Konopka, B. Effectiveness of pheromone traps for the European spruce bark beetle: A comparative study of four commercial products and two new models. Lesn. Cas. For. J. 2016, 62, 207–215. [Google Scholar] [CrossRef]
- Bracalini, M.; Croci, F.; Ciardi, E.; Mannucci, G.; Papucci, E.; Gestri, G.; Tiberi, R.; Panzavolta, T. Ips sexdentatus Mass-Trapping: Mitigation of Its Negative Effects on Saproxylic Beetles Larger Than the Target. Forests 2021, 12, 175. [Google Scholar] [CrossRef]
- Chiverton, P.A.; Sotherton, N.W. The effects on beneficial arthropods of the exclusion of herbicides from cereal crop edges. J. Appl. Ecol. 1991, 28, 1027–1039. [Google Scholar] [CrossRef]
- Aukema, B.H.; Dahlsten, D.L.; Raffa, K.F. Exploiting behavioral disparities among predators and prey to selectively remove pests: Maximizing the ratio of bark beetles to predators removed during semiochemically based trap-out. Environ. Entomol. 2000, 29, 651–660. [Google Scholar] [CrossRef]
- Paradise, C.J.; Penev, Y.; Yu, P.; Grant, D.; Stanback, M. Accumulation of non-target arthropods on sticky tree bands. J. N. C. Acad. Sci. 2016, 132, 1–5. [Google Scholar] [CrossRef]
- Fite, T.; Damte, T.; Tefera, T.; Negeri, M. Evaluation of commercial trap types and lures on the population dynamics of Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) and its effects on non-targets insects. Cogent Food Agric. 2020, 6, 1771116. [Google Scholar] [CrossRef]
- Martín, A.; Etxebeste, I.; Pérez, G.; Álvarez, G.; Sánchez, E.; Pajares, J. Modified pheromone traps help reduce bycatch of bark-beetle natural enemies. Agric. For. Entomol. 2013, 15, 86–97. [Google Scholar] [CrossRef]
- Chinery, M. Guide to the Insects of Britain and Western Europe; Collins Education: Glasgow, UK, 1986; 320p. [Google Scholar]
- Ferreira, M.C.; Cabral, M.T. Pragas do Pinhal; Estação Florestal Nacional: Lisbon, Portugal, 1999; 159p. (In Portuguese) [Google Scholar]
- Ferreira, M.C.; Ferreira, G.W.S. Pragas das Resinosas. Guia de Campo, 3rd ed.; D.G.P.A./Ministério Agricultura, Pescas e Alimentação: Lisbon, Portugal, 1990; 108p. (In Portuguese) [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 2nd ed.; Prantice-Hall International Inc.: New Jersey, NJ, USA, 1984; 718p. [Google Scholar]
- Stat Soft Inc. Statistica 64, version 12; Stat Soft Inc.: Tulsa, OK, USA, 2014.
- Linsley, E.G. Ecology of cerambycidae. Ann. Rev. Entomol. 1959, 4, 99–138. [Google Scholar] [CrossRef]
- Hanks, L.M. Influence of the larval host plant on reproductive strategies of cerambycid beetles. Annu. Rev. Entomol. 1999, 44, 483–505. [Google Scholar] [CrossRef]
- Allison, J.D.; Borden, J.H.; Seybold, S.J. A review of the chemical ecology of the Cerambycidae (Coleoptera). Chemoecology 2004, 14, 123–150. [Google Scholar] [CrossRef]
- De Groot, P.; Nott, R. Response of Monochamus (Col., Cerambycidae) and some Buprestidae to flight intercept traps. J. Appl. Entomol. 2003, 127, 548–552. [Google Scholar] [CrossRef]
- Morewood, W.D.; Heln, K.E.; Katanic, P.J.; Borden, J.H. An improved trap for large wood-boring insects with special reference to Monochamus scutellatus (Coleoptera: Cerambycidae). Can. J. For. Res. 2002, 32, 519–525. [Google Scholar] [CrossRef]
- Ibeas, F.; Gallego, D.; Diez, J.J.; Pajares, J.A. An operative kairomonal lure for managing pine sawyer beetle Monochamus galloprovincialis (Coleoptera: Cerymbycidae). J. Appl. Entomol. 2007, 131, 13–20. [Google Scholar] [CrossRef]
- Billings, R.F. Southern pine bark beetles and associated insects. Effects of rapidly-released host volatiles on response to aggregation pheromones. J. Appl. Ent. 1985, 99, 483–491. [Google Scholar]
- Billings, R.F.; Cameron, R.S. Kairomonal responses of coleoptera, Monochamus titillator (Cerambycidae), Thanasimus dubius (Cleridae), and Temnochila virescens (Trogossitidae), to behavioral chemicals of southern pine bark beetles (Coleoptera: Scolytidae). Environ. Entomol. 1984, 13, 1542–1548. [Google Scholar] [CrossRef]
- Jurc, M.; Hauptman, T.; Pavlin, R.; Borkovic, D. Target and non-target beetles in semiochemical-baited cross vane funnel traps used in monitoring Bursaphelenchus xylophilus (PWN) vectors in pine stands. Phytoparasitica 2016, 44, 151–164. [Google Scholar] [CrossRef]
- Etxebeste, I.; Álvarez, G.; Pérez, G.; Pajares, J.A. Field response of the six-toothed pine bark beetle, Ips sexdentatus (Col.: Curculionidae, Scolytinae), to pheromonal blend candidates. J. Appl. Entomol. 2012, 136, 431–444. [Google Scholar] [CrossRef]
- Panzavolta, T.; Bracalini, M.; Bonuomo, L.; Croci, F.; Tiberi, R. Field response of non-target beetles to Ips sexdentatus aggregation pheromone and pine volatiles. J. Appl. Entomol. 2014, 138, 586–599. [Google Scholar] [CrossRef]
- Seybold, S.J.; Huber, D.P.W.; Lee, J.C.; Graves, A.D.; Bohlmann, J. Pine monoterpenes and pine bark beetles: A marriage of convenience for defense and chemical communication. Phytochem. Rev. 2006, 5, 143–178. [Google Scholar] [CrossRef]
- Serez, M. Verwendung des Aggregationspheromon-Präparats “Ipslure” gegen den mediterranen Kiefernborkenkäfer, Ips (Orthotomicus) erosus (Woll.) (Col., Scolytidae). Schadlingskd. Pfl. 1987, 60, 94–95. [Google Scholar] [CrossRef]
- Raffa, R.F.; Aukema, B.H.; Bentz, B.J.; Carroll, A.L.; Hicke, J.A.; Kolb, T.E. Responses of tree-killing bark beetles to a changing climate. In Climate Change and Insect Pests; Björkman, C., Niemelä, P., Eds.; CABI: Wallingford, UK, 2015; pp. 173–201. [Google Scholar]
- Bakke, A.; Kvamme, T. Kairomone response in Thanasimus predators to pheromone components of Ips typographus. J. Chem. Ecol. 1981, 7, 305–312. [Google Scholar] [CrossRef]
- Mendel, Z.; Podoler, H.; Livne, H. Interactions between Aulonium ruficorne (Coleoptera: Colydiidae) and other natural enemies of bark beetles (Coleoptera: Scolytidae). Entomophaga 1990, 35, 99–105. [Google Scholar] [CrossRef]
- Larrañaga, I.E.; Hernández, A.B.M.; Escolar, G.P.; Fernández, M.F.; Casero, J.J.D.; Alonso, J.A.P. Evaluación de compuestos semioquímicos para su empleo en estrategias de aumento de enemigos naturales de Ips sexdentatus (Col.: Scolytidae). Cuad. Soc. Esp. Cienc. Forestales. 2008, 32, 27–32. [Google Scholar]
- Etxebeste, I.; Lencina, J.L.; Pajares, J. Saproxylic community, guild and species responses to varying pheromone components of a pine bark beetle. Bull. Entomol. Res. 2013, 103, 497–510. [Google Scholar] [CrossRef]
- Schroeder, L.M. Prolonged development time of the bark beetle predator Thanasimus formicarius (Col.; Cleridae) in relation to its prey species Tomicus piniperda (L.) and Ips typographus (L.) (Col.; Scolytidae). Agr. For. Entomol. 1999, 1, 127–135. [Google Scholar] [CrossRef]
- Warzée, N.; Grégoire, J.C. Biodiversité forestiére et annemies naturels des scolytes: Le cas examplaire de Thanasimus formicarius. Forêt Wallonne. 2003, 66, 2–6. [Google Scholar]
- Piper, R. UK Saproxylic Beetles. Available online: https://www.rosspiper.net/2020/01/10/saproxylic-beetles/ (accessed on 18 October 2022).
- Krista, L.; Ryall, L.F. Habitat loss decreases predator-prey ratios in a pine-bark beetle system. Oikos 2005, 110, 265–270. [Google Scholar]
- Bonifácio, L. Evolution and Impact of the Pine Wilt Disease in the Affected Zone South of Tagus River. Ph.D. Thesis, University of Lisbon, Lisbon, Portugal, 2009; 208p. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonifácio, L.; Sousa, E. Optimization of Traps Used in the Management of Monochamus galloprovincialis (Coleoptera: Cerambycidae), the Insect-Vector of Pinewood Nematode, to Reduce By-Catches of Non-Target Insects. Forests 2025, 16, 1017. https://doi.org/10.3390/f16061017
Bonifácio L, Sousa E. Optimization of Traps Used in the Management of Monochamus galloprovincialis (Coleoptera: Cerambycidae), the Insect-Vector of Pinewood Nematode, to Reduce By-Catches of Non-Target Insects. Forests. 2025; 16(6):1017. https://doi.org/10.3390/f16061017
Chicago/Turabian StyleBonifácio, Luís, and Edmundo Sousa. 2025. "Optimization of Traps Used in the Management of Monochamus galloprovincialis (Coleoptera: Cerambycidae), the Insect-Vector of Pinewood Nematode, to Reduce By-Catches of Non-Target Insects" Forests 16, no. 6: 1017. https://doi.org/10.3390/f16061017
APA StyleBonifácio, L., & Sousa, E. (2025). Optimization of Traps Used in the Management of Monochamus galloprovincialis (Coleoptera: Cerambycidae), the Insect-Vector of Pinewood Nematode, to Reduce By-Catches of Non-Target Insects. Forests, 16(6), 1017. https://doi.org/10.3390/f16061017