
Holocene Forest Dynamics in Western Mediterranean Islands: Rates, Periodicity, and Trends

 , , , , , ,
, , , , , ,  , , , , , ,
, , , , , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
- Records for which detailed pollen counts are not available;
- Records with less than four chronological constraints (radiocarbon dates or dated tephras);
- Records spanning less than 3000 years.
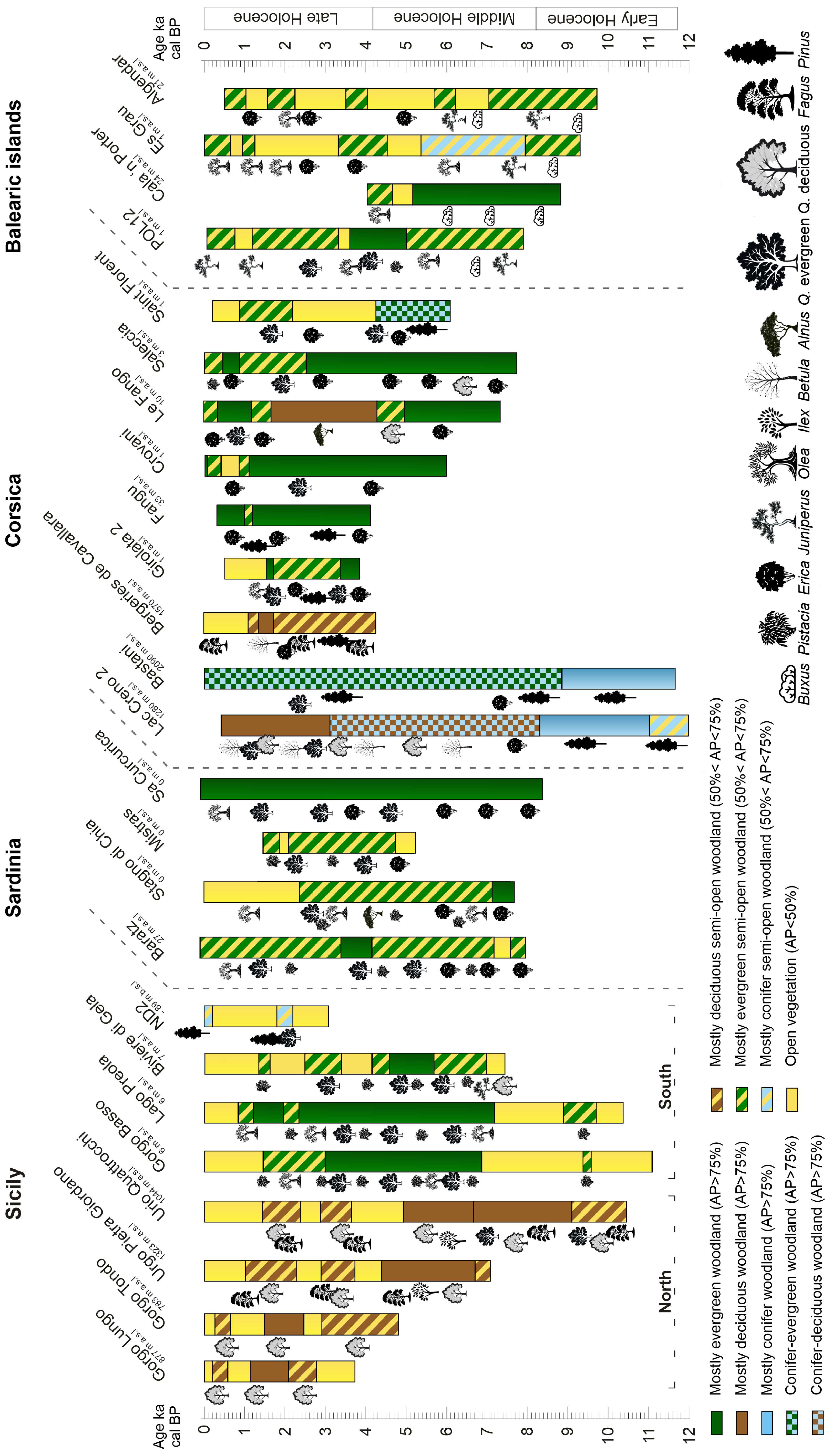
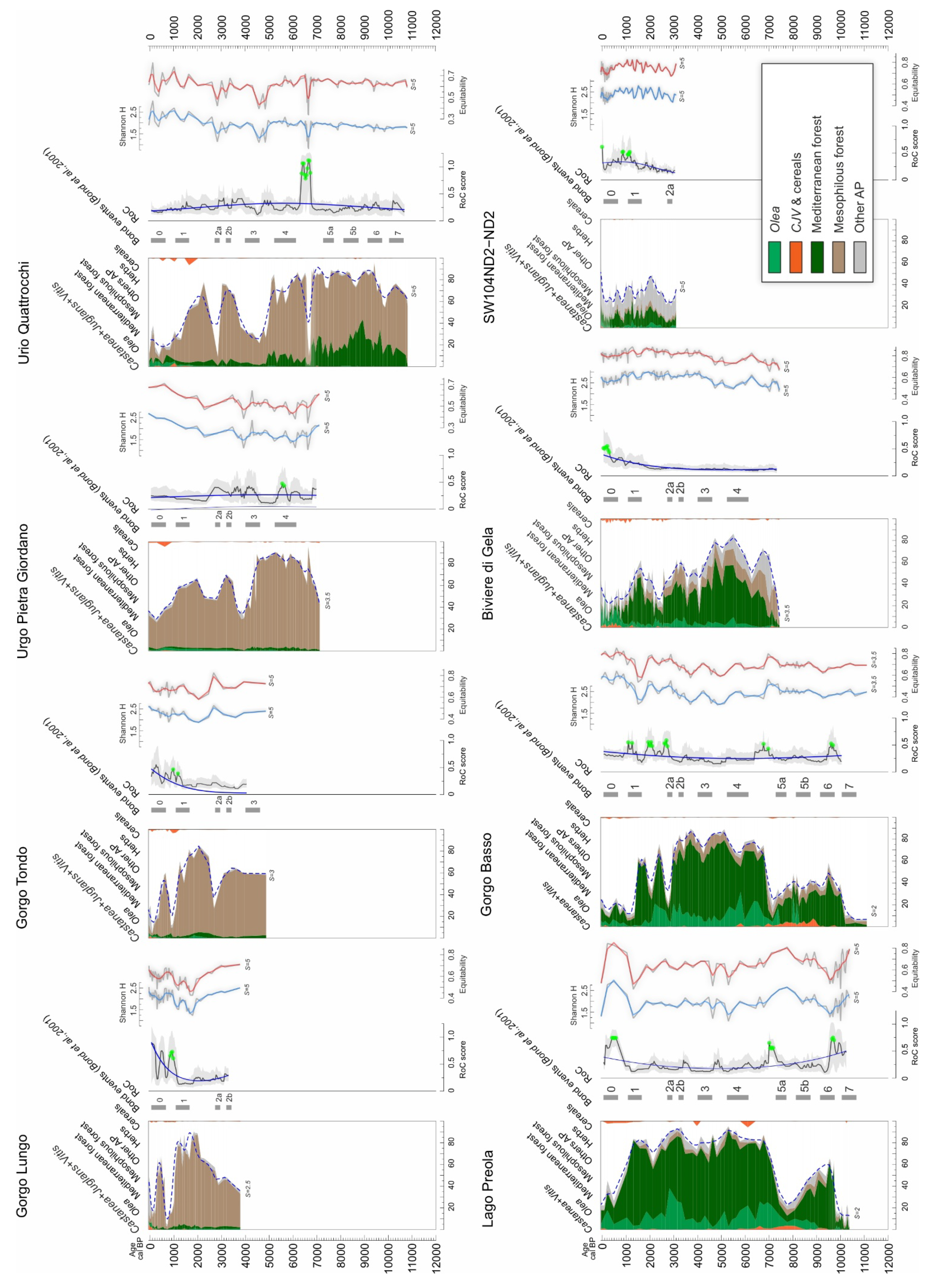
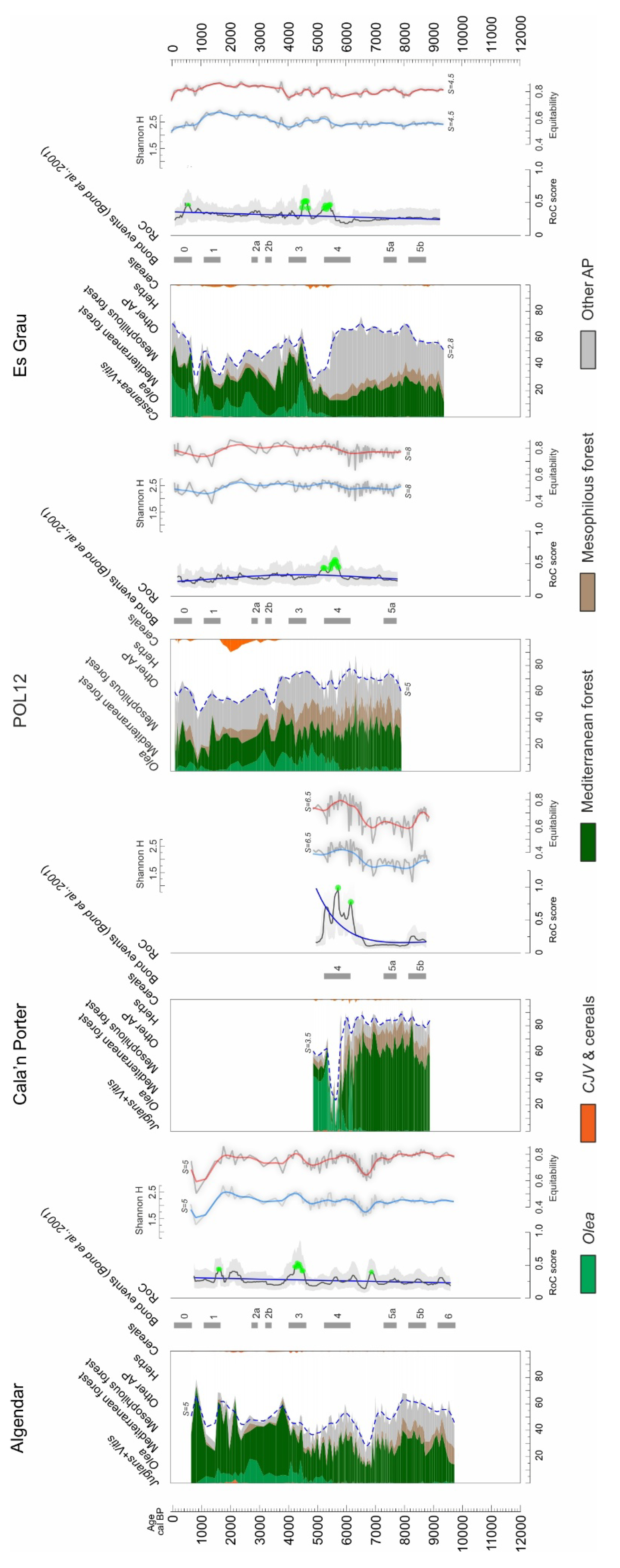
3. Results
3.1. Sicily
3.2. Sardinia
3.3. Corsica
3.4. Balearic Islands
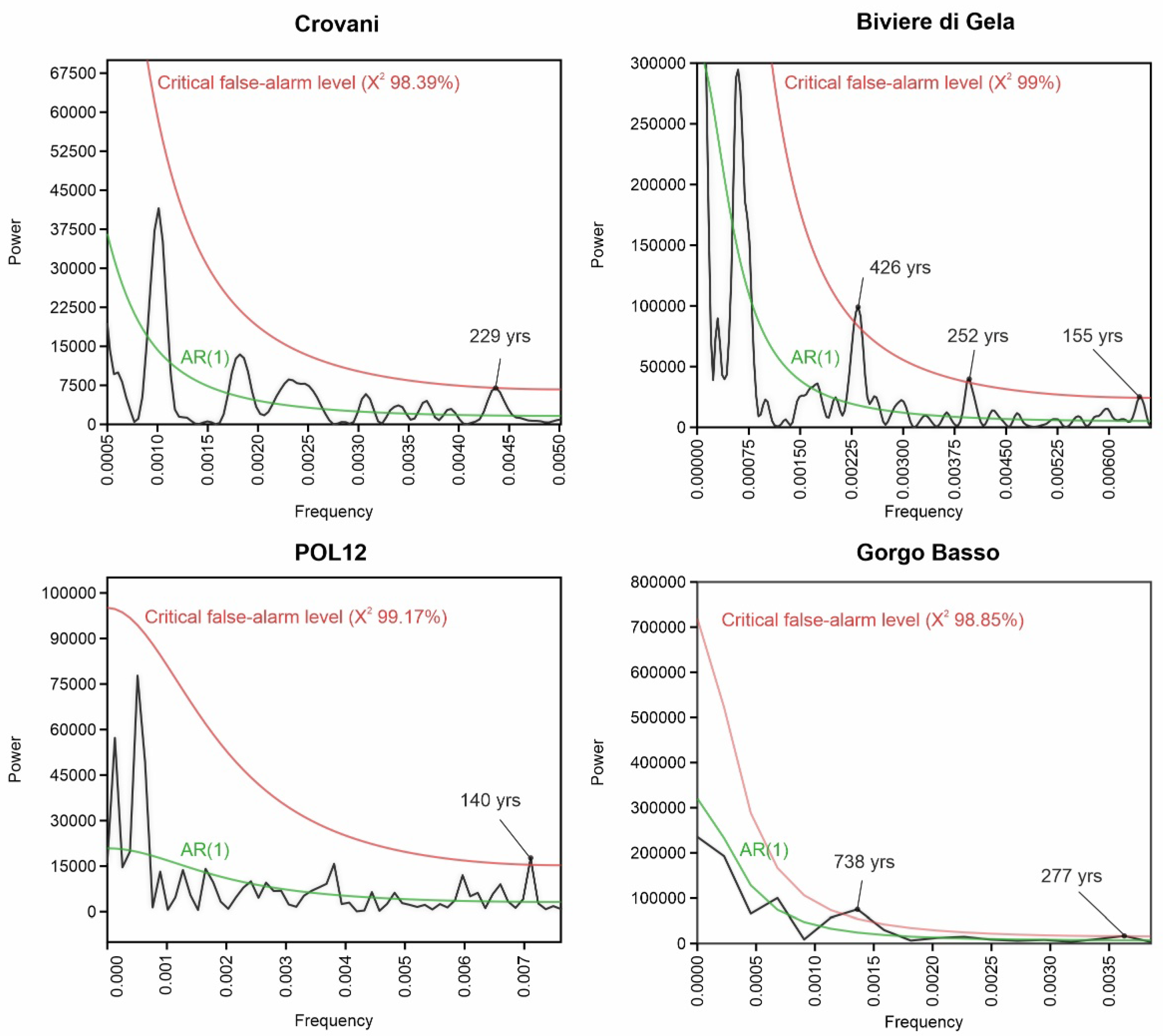
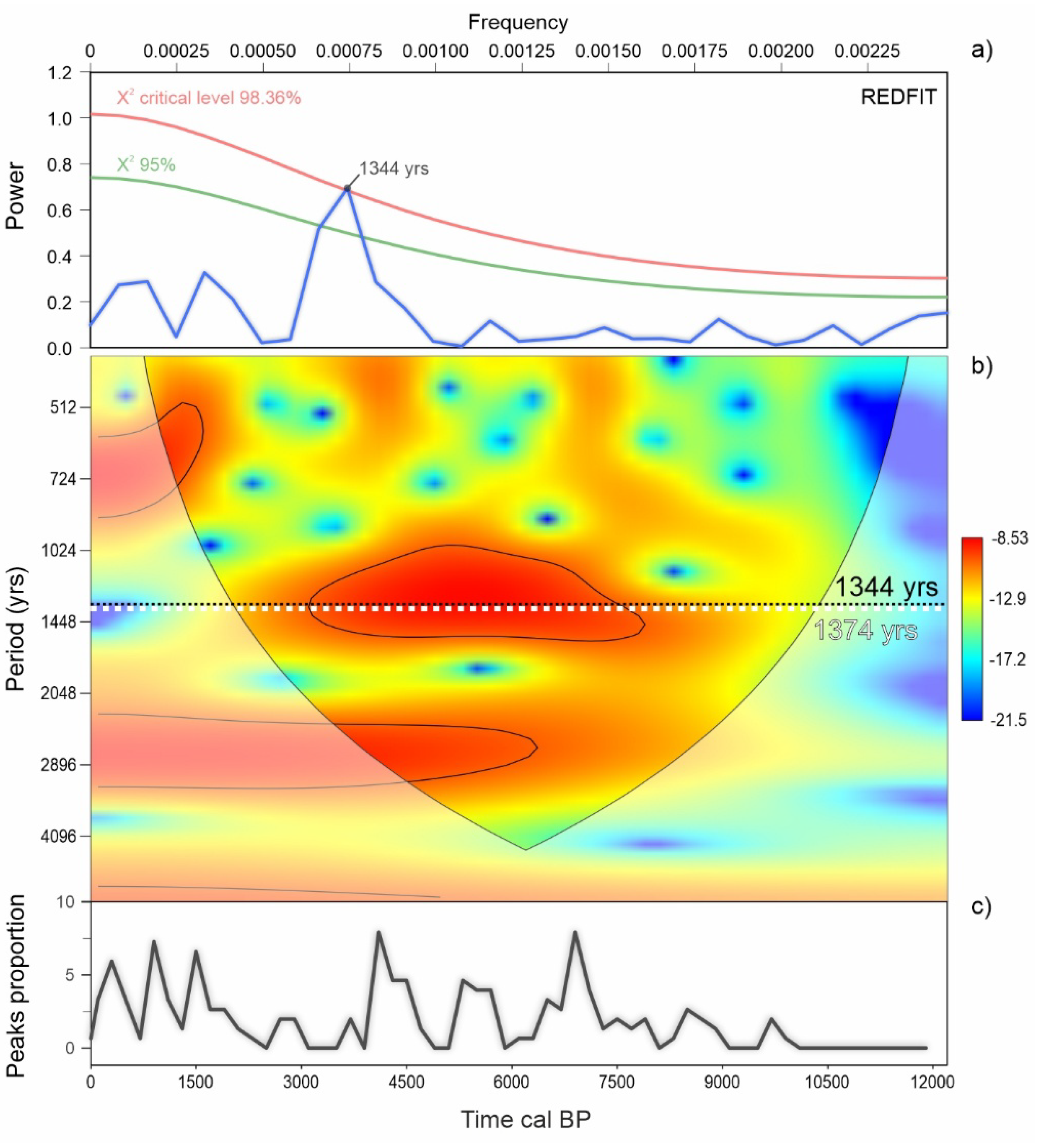
3.5. REDFIT Analysis of the AP Time Series
4. Discussion
4.1. Rate of Change Analysis and Rapid Climate Changes (RCCs)
4.2. Fundamental Tempo of Forest Cover Variability
5. Conclusions
- The RoC analysis proved to be a powerful tool for detecting rapid changes in vegetation composition that may not be apparent in changes in tree cover. This approach is effective in identifying past vegetation responses to RCCs in large Mediterranean islands, especially where compensatory vegetation dynamics between oak woodlands and shrublands/heathlands may result in near-stable AP percentages over time.
- The timing of rapid changes in forest composition detected across large Mediterranean islands is consistent with centennial-scale hydroclimatic oscillations linked to Bond events. This correspondence highlights a teleconnection between the North Atlantic climate system and western Mediterranean forest ecosystems, suggesting that shifts in ocean–atmosphere mechanisms may have influenced regional hydroclimatic conditions.
- The response of insular Mediterranean ecosystems to these climate events could have been more complex, with varying degrees of resilience and adaptation depending on local factors and ecosystem characteristics.
- The spectral analysis carried out on RoC and AP time series revealed periodicities that have already been identified in paleoclimate and palaeoenvironmental records worldwide. Most of them (140–150, 230–250, and 420 years) are related to the influence of solar activity, possibly in relation to its effect on latitudinal shifts of ITCZ and the consequent reorganization of climate patterns. Other periodicities (270, 740, and 1350 years) may derive from fundamental modes of solar activity through a rectification operated by complex oceanic–atmospheric circulation mechanisms possibly involving AMOC, ENSO, and NAO, whose influence in climate variability of the Mediterranean regions is well known.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harter, D.E.V.; Irl, S.D.H.; Seo, B.; Steinbauer, M.J.; Gillespie, R.; Triantis, K.A.; Fernández-Palacios, J.-M.; Beierkuhnlein, C. Impacts of Global Climate Change on the Floras of Oceanic Islands—Projections, Implications and Current Knowledge. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 160–183. [Google Scholar] [CrossRef]
- Taylor, S.; Kumar, L. Global Climate Change Impacts on Pacific Islands Terrestrial Biodiversity: A Review. Trop. Conserv. Sci. 2016, 9, 203–223. [Google Scholar] [CrossRef]
- Médail, F. The Specific Vulnerability of Plant Biodiversity and Vegetation on Mediterranean Islands in the Face of Global Change. Reg. Environ. Chang. 2017, 17, 1775–1790. [Google Scholar] [CrossRef]
- Mcleod, E.; Bruton-Adams, M.; Förster, J.; Franco, C.; Gaines, G.; Gorong, B.; James, R.; Posing-Kulwaum, G.; Tara, M.; Terk, E. Lessons from the Pacific Islands—Adapting to Climate Change by Supporting Social and Ecological Resilience. Front. Mar. Sci. 2019, 6, 289. [Google Scholar] [CrossRef]
- Fernández-Palacios, J.M.; Kreft, H.; Irl, S.D.H.; Norder, S.; Ah-Peng, C.; Borges, P.A.V.; Burns, K.C.; De Nascimento, L.; Meyer, J.-Y.; Montes, E.; et al. Scientists’ Warning—The Outstanding Biodiversity of Islands Is in Peril. Glob. Ecol. Conserv. 2021, 31, e01847. [Google Scholar] [CrossRef]
- Heywood, V.; Culham, A. The Impacts of Climate Change on Plant Species in Europe. Report T-PVS/Inf9E. In Convention on the Conservation of European Wildlife and Natural Habitats, Proceedings of the 29th Meeting of the Standing Committee, Bern, Switzerland, 23–26 November 2009; University of Reading: Reading, UK, 2009. [Google Scholar]
- Medail, F.; Quezel, P. Hot-Spots Analysis for Conservation of Plant Biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Médail, F. Plant Biogeography and Vegetation Patterns of the Mediterranean Islands. Bot. Rev. 2022, 88, 63–129. [Google Scholar] [CrossRef]
- Quézel, P. The Study of Groupings in the Countries Surrounding the Mediterranean: Some Methodological Aspects; Mediterranean-type shrublands; Elsevier: Amsterdam, The Netherlands, 1981; pp. 87–93. [Google Scholar]
- Vogiatzakis, I.N.; Mannion, A.M.; Sarris, D. Mediterranean Island Biodiversity and Climate Change: The Last 10,000 Years and the Future. Biol. Conserv. 2016, 25, 2597–2627. [Google Scholar] [CrossRef]
- Lionello, P.; Malanotte-Rizzoli, P.; Boscolo, R.; Alpert, P.; Artale, V.; Li, L.; Luterbacher, J.; May, W.; Trigo, R.; Tsimplis, M.; et al. The Mediterranean Climate: An Overview of the Main Characteristics and Issues. In Developments in Earth and Environmental Sciences; Lionello, P., Malanotte-Rizzoli, P., Boscolo, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 4, pp. 1–26. ISBN 1571-9197. [Google Scholar]
- Lionello, P. The Climate of the Mediterranean Region: From the Past to the Future; Elsevier: London, UK, 2012; ISBN 978-0-12-416042-2. [Google Scholar]
- Zielhofer, C.; Köhler, A.; Mischke, S.; Benkaddour, A.; Mikdad, A.; Fletcher, W.J. Western Mediterranean Hydro-Climatic Consequences of Holocene Ice-Rafted Debris (Bond) Events. Clim. Past 2019, 15, 463–475. [Google Scholar] [CrossRef]
- Rousseau, D.-D.; Bagniewski, W.; Ghil, M. Abrupt Climate Changes and the Astronomical Theory: Are They Related? Clim. Past 2022, 18, 249–271. [Google Scholar] [CrossRef]
- Bond, G.; Kromer, B.; Beer, J.; Muscheler, R.; Evans, M.N.; Showers, W.; Hoffmann, S.; Lotti-Bond, R.; Hajdas, I.; Bonani, G. Persistent Solar Influence on North Atlantic Climate During the Holocene. Science 2001, 294, 2130–2136. [Google Scholar] [CrossRef] [PubMed]
- Pèlachs, A.; Julià, R.; Pérez-Obiol, R.; Soriano, J.M.; Bal, M.-C.; Cunill, R.; Catalan, J. Potential Influence of Bond Events on Mid-Holocene Climate and Vegetation in Southern Pyrenees as Assessed from Burg Lake LOI and Pollen Records. Holocene 2011, 21, 95–104. [Google Scholar] [CrossRef]
- Di Rita, F.; Fletcher, W.J.; Aranbarri, J.; Margaritelli, G.; Lirer, F.; Magri, D. Holocene Forest Dynamics in Central and Western Mediterranean: Periodicity, Spatio-Temporal Patterns and Climate Influence. Sci. Rep. 2018, 8, 8929. [Google Scholar] [CrossRef] [PubMed]
- Wanner, H.; Buetikofer, J. Holocene Bond Cycles: Real or Imaginary. Geografie 2008, 113, 338–349. [Google Scholar] [CrossRef]
- Bradley, R.S.; Bakke, J. Is There Evidence for a 4.2 Ka BP Event in the Northern North Atlantic Region? Clim. Past 2019, 15, 1665–1676. [Google Scholar] [CrossRef]
- Reille, M. New Pollen-Analytical Researches in Corsica: The Problem of Quercus ilex L. and Erica arborea L., the Origin of Pinus halepensis Miller Forests. New Phytol. 1992, 122, 359–378. [Google Scholar] [CrossRef]
- Burjachs, F.; Pérez-Obiol, R.; Roure, J.M.; Julià, R. Dinámica de La Vegetación Durante El Holoceno En La Isla de Mallorca. In Trabajos de Palinología Básica y Aplicada; Universitat de València: València, Spain, 1994; pp. 199–210. [Google Scholar]
- Yll, E.-I.; Perez-Obiol, R.; Pantaleon-Cano, J.; Roure, J.M. Palynological Evidence for Climatic Change and Human Activity during the Holocene on Minorca (Balearic Islands). Quat. Res. 1997, 48, 339–347. [Google Scholar] [CrossRef]
- Tinner, W.; Vescovi, E.; van Leeuwen, J.F.N.; Colombaroli, D.; Henne, P.D.; Kaltenrieder, P.; Morales-Molino, C.; Beffa, G.; Gnaegi, B.; van der Knaap, W.O.; et al. Holocene Vegetation and Fire History of the Mountains of Northern Sicily (Italy). Veg. Hist. Archaeobot. 2016, 25, 499–519. [Google Scholar] [CrossRef]
- Burjachs, F.; Pérez-Obiol, R.; Picornell-Gelabert, L.; Revelles, J.; Servera-Vives, G.; Expósito, I.; Yll, E.-I. Overview of Environmental Changes and Human Colonization in the Balearic Islands (Western Mediterranean) and Their Impacts on Vegetation Composition during the Holocene. J. Archaeol. Sci. Rep. 2017, 12, 845–859. [Google Scholar] [CrossRef]
- Revelles, J.; Ghilardi, M.; Rossi, V.; Currás, A.; López-Bultó, O.; Brkojewitsch, G.; Vacchi, M. Coastal Landscape Evolution of Corsica Island (W. Mediterranean): Palaeoenvironments, Vegetation History and Human Impacts since the Early Neolithic Period. Quat. Sci. Rev. 2019, 225, 105993. [Google Scholar] [CrossRef]
- Carrión, J.S.; Munuera, M.; Ochando Tomás, J.; López-Sáez, J.A.; Casas-Gallego, M.; González-Sampériz, P.; Badal, E.; Pérez-Díaz, S.; Carrión-Marco, Y.; Jiménez-Moreno, G.; et al. Paleoflora and Paleovegetation of Iberia III: Holocene; Universidad Politécnica de Cartagena, Ministerio de Ciencia e Innovación y Fundación Séneca: Cartagena, Spain, 2022; ISBN 978-84-17853-89-1. [Google Scholar]
- Ghilardi, M.; Revelles, J.; Mary, J.-B.; Rita, F.D.; Delhon, C.; Delanghe, D.; Robresco, S. Mid- to Late-Holocene Coastal Morphological Evolution, Vegetation History and Land-Use Changes of the Porto Gulf UNESCO World Heritage Site and Its Surroundings (NW Corsica Island, Western Mediterranean). Holocene 2023, 33, 1023–1044. [Google Scholar] [CrossRef]
- Ghilardi, M.; Revelles, J.; Fagel, N.; Schmidt, S.; Delanghe, D. Reconstructing the Mid- to Late Holocene Human-Environments Interactions in Cape Corsica (Corsica Island, Western Mediterranean) Based on Sedimentology, Pollen Analyses and Geochemistry. J. Archaeol. Sci. Rep. 2025, 62, 104985. [Google Scholar] [CrossRef]
- Servera-Vives, G.; Riera, S.; Picornell-Gelabert, L.; Moffa-Sánchez, P.; Llergo, Y.; Garcia, A.; Mus-Amezquita, M.; García Álvarez, S.; Calvo Trías, M. The Onset of Islandscapes in the Balearic Islands: A Study-Case of Addaia (Northern Minorca, Spain). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2018, 498, 9–23. [Google Scholar] [CrossRef]
- Williams, J.W.; Grimm, E.C.; Blois, J.L.; Charles, D.F.; Davis, E.B.; Goring, S.J.; Graham, R.W.; Smith, A.J.; Anderson, M.; Arroyo-Cabrales, J.; et al. The Neotoma Paleoecology Database, a Multiproxy, International, Community-Curated Data Resource. Quat. Res. 2018, 89, 156–177. [Google Scholar] [CrossRef]
- Fyfe, R.M.; de Beaulieu, J.-L.; Binney, H.; Bradshaw, R.H.W.; Brewer, S.; Le Flao, A.; Finsinger, W.; Gaillard, M.-J.; Giesecke, T.; Gil-Romera, G.; et al. The European Pollen Database: Past Efforts and Current Activities. Veg. Hist. Archaeobot. 2009, 18, 417–424. [Google Scholar] [CrossRef]
- Blaauw, M.; Christen, J.A. Flexible Paleoclimate Age-Depth Models Using an Autoregressive Gamma Process. Bayesian Anal. 2011, 6, 457–474. [Google Scholar] [CrossRef]
- Blaauw, M.; Christen, J.A.; Bennett, K.D.; Reimer, P.J. Double the Dates and Go for Bayes—Impacts of Model Choice, Dating Density and Quality on Chronologies. Quat. Sci. Rev. 2018, 188, 58–66. [Google Scholar] [CrossRef]
- Li, C.; Postl, A.K.; Böhmer, T.; Cao, X.; Dolman, A.M.; Herzschuh, U. Harmonized Chronologies of a Global Late Quaternary Pollen Dataset (LegacyAge 1.0). Earth Syst. Sci. Data 2022, 14, 1331–1343. [Google Scholar] [CrossRef]
- Bennett, K. Psimpoll’ and ‘Pscomb’: C Programs for Analysing Pollen Data and Plotting Pollen Diagrams (Version 4.27); The 14Chrono Centre, Archaeology and Palaeoecology, Queen’s University of Belfast: Belfast, UK, 2009. [Google Scholar]
- Mottl, O.; Grytnes, J.-A.; Seddon, A.W.R.; Steinbauer, M.J.; Bhatta, K.P.; Felde, V.A.; Flantua, S.G.A.; Birks, H.J.B. Rate-of-Change Analysis in Paleoecology Revisited: A New Approach. Rev. Palaeobot. Palynol. 2021, 293, 104483. [Google Scholar] [CrossRef]
- Mottl, O.; Flantua, S.G.A.; Bhatta, K.P.; Felde, V.A.; Giesecke, T.; Goring, S.; Grimm, E.C.; Haberle, S.; Hooghiemstra, H.; Ivory, S.; et al. Global Acceleration in Rates of Vegetation Change over the Past 18,000 Years. Science 2021, 372, 860–864. [Google Scholar] [CrossRef]
- Birks, H.J.B.; Line, J.M. The Use of Rarefaction Analysis for Estimating Palynological Richness from Quaternary Pollen-Analytical Data. Holocene 1992, 2, 1–10. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A. Past: Paleontological Statistics Software Package for Educaton and Data Anlysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and Extrapolation with Hill Numbers: A Framework for Sampling and Estimation in Species Diversity Studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef]
- Xie, S.; Li, K.; Li, F.; Yang, X.; Reitalu, T. Modern Pollen–Plant Diversity Relationships for Reliable Pollen-Based Reconstruction of Past Plant Taxonomic and Functional Diversity: A Case Study in Southwest Shandong, China. Ecol. Indic. 2024, 167, 112739. [Google Scholar] [CrossRef]
- Schulz, M.; Mudelsee, M. REDFIT: Estimating red-noise spectra directly from unevenly spaced paleoclimatic time series. Comput. Geosci. 2002, 28, 421–426. [Google Scholar] [CrossRef]
- Vachula, R.S.; Cheung, A.H. A Meta-Analysis of Studies Attributing Significance to Solar Irradiance. Earth Space Sci. 2023, 10, e2022EA002466. [Google Scholar] [CrossRef]
- Bazan, G.; Marino, P.; Guarino, R.; Domina, G.; Schicchi, R. Bioclimatology and Vegetation Series in Sicily: A Geostatistical Approach. Ann. Bot. Fenn. 2015, 52, 1–18. [Google Scholar] [CrossRef]
- Bisculm, M.; Colombaroli, D.; Vescovi, E.; van Leeuwen, J.F.N.; Henne, P.D.; Rothen, J.; Procacci, G.; Pasta, S.; La Mantia, T.; Tinner, W. Holocene Vegetation and Fire Dynamics in the Supra-Mediterranean Belt of the Nebrodi Mountains (Sicily, Italy). J. Quat. Sci. 2012, 27, 687–698. [Google Scholar] [CrossRef]
- Tinner, W.; van Leeuwen, J.F.N.; Colombaroli, D.; Vescovi, E.; van der Knaap, W.O.; Henne, P.D.; Pasta, S.; D’Angelo, S.; La Mantia, T. Holocene Environmental and Climatic Changes at Gorgo Basso, a Coastal Lake in Southern Sicily, Italy. Quat. Sci. Rev. 2009, 28, 1498–1510. [Google Scholar] [CrossRef]
- Calò, C.; Henne, P.D.; Curry, B.; Magny, M.; Vescovi, E.; La Mantia, T.; Pasta, S.; Vannière, B.; Tinner, W. Spatio-Temporal Patterns of Holocene Environmental Change in Southern Sicily. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 323–325, 110–122. [Google Scholar] [CrossRef]
- Grassa, F.; Favara, R.; Valenza, M. Moisture Source in the Hyblean Mountains Region (South-Eastern Sicily, Italy): Evidence from Stable Isotopes Signature. Appl. Geochem. 2006, 21, 2082–2095. [Google Scholar] [CrossRef]
- Noti, R.; van Leeuwen, J.F.N.; Colombaroli, D.; Vescovi, E.; Pasta, S.; La Mantia, T.; Tinner, W. Mid- and Late-Holocene Vegetation and Fire History at Biviere Di Gela, a Coastal Lake in Southern Sicily, Italy. Veg. Hist. Archaeobot. 2009, 18, 371–387. [Google Scholar] [CrossRef]
- Michelangeli, F.; Di Rita, F.; Lirer, F.; Lubritto, C.; Bellucci, L.G.; Cascella, A.; Bonomo, S.; Margaritelli, G.; Magri, D. Vegetation History of SE Sicily from Feudal Land Management to Post-War Agricultural Industrialization. Rev. Palaeobot. Palynol. 2022, 296, 104547. [Google Scholar] [CrossRef]
- Michelangeli, F.; Di Rita, F.; Celant, A.; Tisnérat-Laborde, N.; Lirer, F.; Magri, D. Three Millennia of Vegetation, Land-Use, and Climate Change in SE Sicily. Forests 2022, 13, 102. [Google Scholar] [CrossRef] [PubMed]
- Schwörer, C.; Morales-Molino, C.; Gobet, E.; Henne, P.D.; Pasta, S.; Pedrotta, T.; van Leeuwen, J.F.N.; Vannière, B.; Tinner, W. Simulating Past and Future Fire Impacts on Mediterranean Ecosystems. J. Ecol. 2024, 112, 954–970. [Google Scholar] [CrossRef]
- Di Rita, F.; Melis, R.T. The Cultural Landscape near the Ancient City of Tharros (Central West Sardinia): Vegetation Changes and Human Impact. J. Archaeol. Sci. 2013, 40, 4271–4282. [Google Scholar] [CrossRef]
- Beffa, G.; Pedrotta, T.; Colombaroli, D.; Henne, P.D.; van Leeuwen, J.F.N.; Süsstrunk, P.; Kaltenrieder, P.; Adolf, C.; Vogel, H.; Pasta, S.; et al. Vegetation and Fire History of Coastal North-Eastern Sardinia (Italy) under Changing Holocene Climates and Land Use. Veg. Hist. Archaeobot. 2016, 25, 271–289. [Google Scholar] [CrossRef]
- Melis, R.T.; Di Rita, F.; French, C.; Marriner, N.; Montis, F.; Serreli, G.; Sulas, F.; Vacchi, M. 8000 years of Coastal Changes on a Western Mediterranean Island: A Multiproxy Approach from the Posada Plain of Sardinia. Mar. Geol. 2018, 403, 93–108. [Google Scholar] [CrossRef]
- Pedrotta, T.; Gobet, E.; Schwörer, C.; Beffa, G.; Butz, C.; Henne, P.D.; Morales-Molino, C.; Pasta, S.; van Leeuwen, J.F.N.; Vogel, H.; et al. 8000 Years of Climate, Vegetation, Fire and Land-Use Dynamics in the Thermo-Mediterranean Vegetation Belt of Northern Sardinia (Italy). Veg. Hist. Archaeobot. 2021, 30, 789–813. [Google Scholar] [CrossRef]
- Reille, M. Contribution Pollen Analytique a l’histoire Tardiglaciaire et Holocene de la Vegetation de la Montagne Corse. Ph.D. Thesis, Université d’Aix-Marseille, Marseille, France, 1975. [Google Scholar]
- Di Rita, F.; Ghilardi, M.; Fagel, N.; Vacchi, M.; Warichet, F.; Delanghe, D.; Sicurani, J.; Martinet, L.; Robresco, S. Natural and Anthropogenic Dynamics of the Coastal Environment in Northwestern Corsica (Western Mediterranean) over the Past Six Millennia. Quat. Sci. Rev. 2022, 278, 107372. [Google Scholar] [CrossRef]
- Lestienne, M.; Jouffroy-Bapicot, I.; Leyssenne, D.; Sabatier, P.; Debret, M.; Albertini, P.-J.; Colombaroli, D.; Didier, J.; Hély, C.; Vannière, B. Fires and Human Activities as Key Factors in the High Diversity of Corsican Vegetation. Holocene 2020, 30, 244–257. [Google Scholar] [CrossRef]
- Poher, Y.; Ponel, P.; Médail, F.; Andrieu-Ponel, V.; Guiter, F. Holocene Environmental History of a Small Mediterranean Island in Response to Sea-Level Changes, Climate and Human Impact. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2017, 465, 247–263. [Google Scholar] [CrossRef]
- Picornell-Gelabert, L.; Servera-Vives, G. Landscape Practices and Everyday Life in Domestic Spaces in Bronze Age Mallorca (Balearic Islands): Perspectives for and Archaeology of Fuel and Firewood. Quat. Int. 2017, 431, 73–89. [Google Scholar] [CrossRef]
- Di Domenico, F.; Lucchese, F.; Magri, D. Buxus in Europe: Late Quaternary Dynamics and Modern Vulnerability. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 354–362. [Google Scholar] [CrossRef]
- Ninyerola, M.; Sáez, L.; Pérez-Obiol, R. Relating Postglacial Relict Plants and Holocene Vegetation Dynamics in the Balearic Islands Through Field Surveys, Pollen Analysis and GIS Modeling. Plant Biosyst. 2007, 141, 292–304. [Google Scholar] [CrossRef]
- Kaniewski, D.; Marriner, N.; Cheddadi, R.; Morhange, C.; Ontiveros, M.Á.C.; Fornós, J.J.; Giaime, M.; Trichon, V.; Otto, T.; Luce, F.; et al. Recent Anthropogenic Climate Change Exceeds the Rate and Magnitude of Natural Holocene Variability on the Balearic Islands. Anthropocene 2020, 32, 100268. [Google Scholar] [CrossRef]
- Onac, B.P.; Polyak, V.J.; Mitrovica, J.X.; Ginés, J.; Gràcia, F.; Fornós, J.J.; Ginés, A.; Asmerom, Y. Submerged Bridge Constructed at Least 5600 Years Ago Indicates Early Human Arrival in Mallorca Spain. Commun. Earth Environ. 2024, 5, 457. [Google Scholar] [CrossRef]
- Carrión, J.S.; Fernández, S.; González-Sampériz, P.; Gil-Romera, G.; Badal, E.; Carrión-Marco, Y.; López-Merino, L.; López-Sáez, J.A.; Fierro, E.; Burjachs, F. Expected Trends and Surprises in the Lateglacial and Holocene Vegetation History of the Iberian Peninsula and Balearic Islands. Rev. Palaeobot. Palynol. 2010, 162, 458–475. [Google Scholar] [CrossRef]
- Di Rita, F.; Magri, D. An Overview of the Holocene Vegetation History from the Central Mediterranean Coasts. J. Mech. Eng. Sci. 2012, 4, 35–52. [Google Scholar] [CrossRef]
- Buosi, C.; Del Rio, M.; Orrù, P.; Pittau, P.; Scanu, G.G.; Solinas, E. Sea Level Changes and Past Vegetation in the Punic Period (5th–4th Century BC): Archaeological, Geomorphological and Palaeobotanical Indicators (South Sardinia—West Mediterranean Sea). Quat. Int. 2017, 439, 141–157. [Google Scholar] [CrossRef]
- Vella, M.-A.; Andrieu-Ponel, V.; Cesari, J.; Leandri, F.; Pêche-Quilichini, K.; Reille, M.; Poher, Y.; Demory, F.; Delanghe, D.; Ghilardi, M.; et al. Early Impact of Agropastoral Activities and Climate on the Littoral Landscape of Corsica since Mid-Holocene. PLoS ONE 2019, 14, e0226358. [Google Scholar] [CrossRef] [PubMed]
- Di Rita, F.; Lirer, F.; Bonomo, S.; Cascella, A.; Ferraro, L.; Florindo, F.; Insinga, D.D.; Lurcock, P.C.; Margaritelli, G.; Petrosino, P.; et al. Late Holocene Forest Dynamics in the Gulf of Gaeta (Central Mediterranean) in Relation to NAO Variability and Human Impact. Quat. Sci. Rev. 2018, 179, 137–152. [Google Scholar] [CrossRef]
- Jalut, G.; Dedoubat, J.J.; Fontugne, M.; Otto, T. Holocene Circum-Mediterranean Vegetation Changes: Climate Forcing and Human Impact. Quat. Int. 2009, 200, 4–18. [Google Scholar] [CrossRef]
- Pérez-Obiol, R.; Jalut, G.; Julià, R.; Pèlachs, A.; Iriarte, M.J.; Otto, T.; Hernández-Beloqui, B. Mid-Holocene Vegetation and Climatic History of the Iberian Peninsula. Holocene 2011, 21, 75–93. [Google Scholar] [CrossRef]
- Combourieu-Nebout, N.; Peyron, O.; Bout-Roumazeilles, V.; Goring, S.; Dormoy, I.; Joannin, S.; Sadori, L.; Siani, G.; Magny, M. Holocene Vegetation and Climate Changes in the Central Mediterranean Inferred from a High-Resolution Marine Pollen Record (Adriatic Sea). Clim. Past 2013, 9, 2023–2042. [Google Scholar] [CrossRef]
- Cipolla, S.S.; Montaldo, N. On the Impacts of Historical and Future Climate Changes to the Sustainability of the Main Sardinian Forests. Remote Sens. 2022, 14, 4893. [Google Scholar] [CrossRef]
- Magri, D.; Parra, I. Late Quaternary Western Mediterranean Pollen Records and African Winds. Earth Planet. Sci. Lett. 2002, 200, 401–408. [Google Scholar] [CrossRef]
- Di Rita, F.; Magri, D. Holocene Drought, Deforestation and Evergreen Vegetation Development in the Central Mediterranean: A 5500 Year Record from Lago Alimini Piccolo, Apulia, Southeast Italy. Holocene 2009, 19, 295–306. [Google Scholar] [CrossRef]
- Di Rita, F.; Magri, D. The 4.2 Ka Event in the Vegetation Record of the Central Mediterranean. Clim. Past 2019, 15, 237–251. [Google Scholar] [CrossRef]
- Di Rita, F.; Michelangeli, F.; Celant, A.; Magri, D. Sign-Switching Ecological Changes in the Mediterranean Basin at 4.2 Ka BP. Glob. Planet. Chang. 2022, 208, 103713. [Google Scholar] [CrossRef]
- Bond, G.; Showers, W.; Cheseby, M.; Lotti, R.; Almasi, P.; deMenocal, P.; Priore, P.; Cullen, H.; Hajdas, I.; Bonani, G. A Pervasive Millennial-Scale Cycle in North Atlantic Holocene and Glacial Climates. Science 1997, 278, 1257–1266. [Google Scholar] [CrossRef]
- Wirth, S.B.; Glur, L.; Gilli, A.; Anselmetti, F.S. Holocene Flood Frequency across the Central Alps—Solar Forcing and Evidence for Variations in North Atlantic Atmospheric Circulation. Quat. Sci. Rev. 2013, 80, 112–128. [Google Scholar] [CrossRef]
- Le Houedec, S.; Liebrand, D.; Hennekam, R.; Mojtahid, M. Assessing Atmospheric and Oceanic Teleconnections between the Eastern and Western Mediterranean over the Past 8000 Years. Holocene 2024, 34, 162–174. [Google Scholar] [CrossRef]
- Incarbona, A.; Sprovieri, M. The Postglacial Isotopic Record of Intermediate Water Connects Mediterranean Sapropels and Organic-Rich Layers. Paleoceanogr. Paleoclimatol. 2020, 35, e2020PA004009. [Google Scholar] [CrossRef]
- Chiessi, C.M.; Mulitza, S.; Groeneveld, J.; Silva, J.B.; Campos, M.C.; Gurgel, M.H.C. Variability of the Brazil Current during the Late Holocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 415, 28–36. [Google Scholar] [CrossRef]
- Dima, M.; Lohmann, G. Conceptual Model for Millennial Climate Variability: A Possible Combined Solar-Thermohaline Circulation Origin for the ~1,500-Year Cycle. Clim. Dyn. 2009, 32, 301–311. [Google Scholar] [CrossRef]
- Berner, K.S.; Koç, N.; Divine, D.; Godtliebsen, F.; Moros, M. A Decadal-scale Holocene Sea Surface Temperature Record from the Subpolar North Atlantic Constructed Using Diatoms and Statistics and Its Relation to Other Climate Parameters. Paleoceanography 2008, 23, 2006PA001339. [Google Scholar] [CrossRef]
- Russell, J.M.; Johnson, T.C.; Talbot, M.R. A 725 Yr Cycle in the Climate of Central Africa during the Late Holocene. Geology 2003, 31, 677–680. [Google Scholar] [CrossRef]
- Fletcher, W.J.; Debret, M.; Goñi, M.F.S. Mid-Holocene Emergence of a Low-Frequency Millennial Oscillation in Western Mediterranean Climate: Implications for Past Dynamics of the North Atlantic Atmospheric Westerlies. Holocene 2013, 23, 153–166. [Google Scholar] [CrossRef]
- Volkov, D.L.; Baringer, M.; Smeed, D.; Johns, W.; Landerer, F.W. Teleconnection between the Atlantic Meridional Overturning Circulation and Sea Level in the Mediterranean Sea. J. Clim. 2019, 32, 935–955. [Google Scholar] [CrossRef]
- Petridi, N.; Polychroni, I.; Hatzaki, M. The Impact of the Atlantic Meridional Overturning Circulation (AMOC) Variability on the Mediterranean Climate. Environ. Sci. Proc. 2023, 26, 160. [Google Scholar] [CrossRef]
- Stuiver, M.; Braziunas, T.F. Atmospheric 14C and Century-Scale Solar Oscillations. Nature 1989, 338, 405–408. [Google Scholar] [CrossRef]
- Damon, P.E.; Jirikowic, J.L. The Sun as a Low-Frequency Harmonic Oscillator. Radiocarbon 1992, 34, 199–205. [Google Scholar] [CrossRef]
- Yu, Z.; Ito, E. The 400-Year Wet-Dry Climate Cycle in Interior North America and Its Solar Connection. In Proceedings of the Proceedings of the Nineteenth Annual Pacific Climate Workshop, Pacific Grove, CA, USA, 3–6 March 2002. [Google Scholar]
- Sabatier, P.; Nicolle, M.; Piot, C.; Colin, C.; Debret, M.; Swingedouw, D.; Perrette, Y.; Bellingery, M.-C.; Chazeau, B.; Develle, A.-L.; et al. Past African Dust Inputs in the Western Mediterranean Area Controlled by the Complex Interaction between the Intertropical Convergence Zone, the North Atlantic Oscillation, and Total Solar Irradiance. Clim. Past 2020, 16, 283–298. [Google Scholar] [CrossRef]
- Martín-Chivelet, J.; Muñoz-García, M.B.; Edwards, R.L.; Turrero, M.J.; Ortega, A.I. Land Surface Temperature Changes in Northern Iberia since 4000 Yr BP, Based on δ13C of Speleothems. Glob. Planet. Chang. 2011, 77, 1–12. [Google Scholar] [CrossRef]
- Degeai, J.-P.; Devillers, B.; Dezileau, L.; Oueslati, H.; Bony, G. Major Storm Periods and Climate Forcing in the Western Mediterranean during the Late Holocene. Quat. Sci. Rev. 2015, 129, 37–56. [Google Scholar] [CrossRef]
- Azuara, J.; Sabatier, P.; Lebreton, V.; Jalali, B.; Sicre, M.-A.; Dezileau, L.; Bassetti, M.-A.; Frigola, J.; Combourieu-Nebout, N. Mid- to Late-Holocene Mediterranean Climate Variability: Contribution of Multi-Proxy and Multi-Sequence Comparison Using Wavelet Spectral Analysis in the Northwestern Mediterranean Basin. Earth-Sci. Rev. 2020, 208, 103232. [Google Scholar] [CrossRef]
- Gao, G.; Niu, H.; Song, L.; Wang, J.; Meng, M.; Yang, Z.; Liu, L.; Wei, Y.; Chen, N.; Yu, J.; et al. Solar Forcing and ENSO Regulated Rates of Chang. of Ecosystems in Northeast China since the Last Deglaciation. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2023, 627, 111728. [Google Scholar] [CrossRef]
- Piervitali, E.; Colacino, M. Evidence of Drought in Western Sicily during the Period 1565–1915 from Liturgical Offices. Clim. Chang. 2001, 49, 225–238. [Google Scholar] [CrossRef]
- Vaquero, J.M.; Gallego, M.C.; García, J.A. A 250-Year Cycle in Naked-Eye Observations of Sunspots. Geophys. Res. Lett. 2002, 29, 58-1–58-4. [Google Scholar] [CrossRef]
- Ogurtsov, M.G.; Nagovitsyn, Y.A.; Kocharov, G.E.; Jungner, H. Long-Period Cycles of the Sun’s Activity Recorded in Direct Solar Data and Proxies. Sol. Phys. 2002, 211, 371–394. [Google Scholar] [CrossRef]
- Matthews, J.A.; Olaf Dahl, S.; Nesje, A.; Berrisford, M.S.; Andersson, C. Holocene Glacier Variations in Central Jotunheimen, Southern Norway Based on Distal Glaciolacustrine Sediment Cores. Quat. Sci. Rev. 2000, 19, 1625–1647. [Google Scholar] [CrossRef]
- Borgmark, A. Holocene Climate Variability and Periodicities in South-Central Sweden, as Interpreted from Peat Humification Analysis. Holocene 2005, 15, 387–395. [Google Scholar] [CrossRef]
- Dykoski, C.A.; Edwards, R.L.; Cheng, H.; Yuan, D.; Cai, Y.; Zhang, M.; Lin, Y.; Qing, J.; An, Z.; Revenaugh, J. A High-Resolution, Absolute-Dated Holocene and Deglacial Asian Monsoon Record from Dongge Cave, China. Earth Planet. Sci. Lett. 2005, 233, 71–86. [Google Scholar] [CrossRef]
- Swindles, G.T.; Plunkett, G.; Roe, H.M. A Delayed Climatic Response to Solar Forcing at 2800 Cal. BP: Multiproxy Evidence from Three Irish Peatlands. Holocene 2007, 17, 177–182. [Google Scholar] [CrossRef]
- Swindles, G.T.; Patterson, R.T.; Roe, H.M.; Galloway, J.M. Evaluating Periodicities in Peat-Based Climate Proxy Records. Quat. Sci. Rev. 2012, 41, 94–103. [Google Scholar] [CrossRef]
- Di Rita, F. A possible solar pacemaker for Holocene fluctuations of a salt-marsh in southern Italy. Quat. Int. 2013, 288, 239–248. [Google Scholar] [CrossRef]
- Castagnoli, G.C.; Bonino, G.; Della Monica, P.; Taricco, C.; Bernasconi, S.M. Solar Activity in the Last Millennium Recorded in the δ18O Profile of Planktonic Foraminifera of a Shallow Water Ionian Sea Core. Sol. Phys. 1999, 188, 191–202. [Google Scholar] [CrossRef]
- Ma, L.; Vaquero, J.M. New Evidence of the Suess/de Vries Cycle Existing in Historical Naked-Eye Observations of Sunspots. Open Astron. 2020, 29, 28–31. [Google Scholar] [CrossRef]
- Jiang, D.; Lang, X.; Tian, Z.; Ju, L. Mid-Holocene East Asian Summer Monsoon Strengthening: Insights from Paleoclimate Modeling Intercomparison Project (PMIP) Simulations. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 369, 422–429. [Google Scholar] [CrossRef]
- Turney, C.S.M.; Palmer, J.; Hogg, A.; Fogwill, C.J.; Jones, R.T.; Bronk Ramsey, C.; Fenwick, P.; Grierson, P.; Wilmshurst, J.; O’Donnell, A.; et al. Multidecadal Variations in Southern Hemisphere Atmospheric 14C: Evidence against a Southern Ocean Sink at the End of the Little Ice Age CO2 Anomaly. Glob. Biogeochem. Cycles 2016, 30, 211–218. [Google Scholar] [CrossRef]
- Spiridonov, A.; Balakauskas, L.; Stankevič, R.; Kluczynska, G.; Gedminienė, L.; Stančikaitė, M. Holocene Vegetation Patterns in Southern Lithuania Indicate Astronomical Forcing on the Millennial and Centennial Time Scales. Sci. Rep. 2019, 9, 14711. [Google Scholar] [CrossRef] [PubMed]
- Ron, C.; Chapanov, Y.; Vondrak, J. Solar Excitation of Bicentennial Earth Rotation Oscillations. Acta Geodyn. Geomater. 2012, 9, 167. [Google Scholar]
- Incarbona, A.; Bonomo, S.; Cacho, I.; Lirer, F.; Margaritelli, G.; Pecoraro, D.; Ziveri, P. Solar Forcing for Nutricline Depth Variability Inferred by Coccoliths in the Pre-Industrial Northwestern Mediterranean. Glob. Planet. Chang. 2023, 224, 104102. [Google Scholar] [CrossRef]
- Pinardi, N.; Zavatarelli, M.; Adani, M.; Coppini, G.; Fratianni, C.; Oddo, P.; Simoncelli, S.; Tonani, M.; Lyubartsev, V.; Dobricic, S.; et al. Mediterranean Sea Large-Scale Low-Frequency Ocean Variability and Water Mass Formation Rates from 1987 to 2007: A Retrospective Analysis. Prog. Oceanogr. 2015, 132, 318–332. [Google Scholar] [CrossRef]
- Somot, S.; Houpert, L.; Sevault, F.; Testor, P.; Bosse, A.; Taupier-Letage, I.; Bouin, M.-N.; Waldman, R.; Cassou, C.; Sanchez-Gomez, E.; et al. Characterizing, Modelling and Understanding the Climate Variability of the Deep Water Formation in the North-Western Mediterranean Sea. Clim. Dyn. 2018, 51, 1179–1210. [Google Scholar] [CrossRef]
- Seyfried, L.; Estournel, C.; Marsaleix, P.; Richard, E. Dynamics of the North Balearic Front during an Autumn Tramontane and Mistral Storm: Air–Sea Coupling Processes and Stratification Budget Diagnostic. Ocean Sci. 2019, 15, 179–198. [Google Scholar] [CrossRef]
- Briffa, K.R.; Jones, P.D.; Bartholin, T.S.; Eckstein, D.; Schweingruber, F.H.; Karlén, W.; Zetterberg, P.; Eronen, M. Fennoscandian Summers from Ad 500: Temperature Changes on Short and Long Timescales. Clim. Dyn. 1992, 7, 111–119. [Google Scholar] [CrossRef]
- Loope, W.L.; Arbogast, A.F. Dominance of an ∼150-Year Cycle of Sand-Supply Change in Late Holocene Dune-Building along the Eastern Shore of Lake Michigan. Quat. Res. 2000, 54, 414–422. [Google Scholar] [CrossRef]
- Raspopov, O.M.; Dergachev, V.A.; Esper, J.; Kozyreva, O.V.; Frank, D.; Ogurtsov, M.; Kolström, T.; Shao, X. The Influence of the de Vries (∼200-Year) Solar Cycle on Climate Variations: Results from the Central Asian Mountains and Their Global Link. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 259, 6–16. [Google Scholar] [CrossRef]
- Hou, G.; E, C.; Liu, X.; Zeng, F. Reconstruction of Integrated Temperature Series of the Past 2000 Years on the Tibetan Plateau with 10-Year Intervals. Theor. Appl. Climatol. 2013, 113, 259–269. [Google Scholar] [CrossRef]
- Huang, C.; Yang, Y.; Chen, F.; Kong, D.; Xu, J.; Chen, C.; Zhu, Q.; Huang, X. External Forcing Mechanisms Controlling the Qiongdong Upwelling in the Northern South China Sea during the Holocene. Glob. Planet. Chang. 2023, 220, 104021. [Google Scholar] [CrossRef]
- Ptitsyna, N.G.; Demina, I.M. Frequency Modulation as a Cause of Additional Branches of the Gleissberg Cycle in Solar Activity. Geomagn. Aeron. 2021, 61, S48–S61. [Google Scholar] [CrossRef]
- Azharuddin, S.; Govil, P.; Singh, A.D.; Mishra, R.; Shekhar, M. Solar Insolation Driven Periodicities in Southwest Monsoon and Its Impact on NE Arabian Sea Paleoceanography. Geosci. Front. 2019, 10, 2251–2263. [Google Scholar] [CrossRef]
- Poore, R.Z.; Dowsett, H.J.; Verardo, S.; Quinn, T.M. Millennial- to Century-Scale Variability in Gulf of Mexico Holocene Climate Records. Paleoceanography 2003, 18, 1048. [Google Scholar] [CrossRef]
- Nogué, S.; de Nascimento, L.; Froyd, C.A.; Wilmshurst, J.M.; de Boer, E.J.; Coffey, E.E.D.; Whittaker, R.J.; Fernández-Palacios, J.M.; Willis, K.J. Island Biodiversity Conservation Needs Palaeoecology. Nat. Ecol. Evol. 2017, 1, 0181. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michelangeli, F.; De Luca, E.; Magri, D.; De Santis, S.; Celant, A.; Ghilardi, M.; Vacchi, M.; Revelles, J.; Melis, R.T.; Ochando, J.; et al. Holocene Forest Dynamics in Western Mediterranean Islands: Rates, Periodicity, and Trends. Forests 2025, 16, 808. https://doi.org/10.3390/f16050808
Michelangeli F, De Luca E, Magri D, De Santis S, Celant A, Ghilardi M, Vacchi M, Revelles J, Melis RT, Ochando J, et al. Holocene Forest Dynamics in Western Mediterranean Islands: Rates, Periodicity, and Trends. Forests. 2025; 16(5):808. https://doi.org/10.3390/f16050808
Chicago/Turabian StyleMichelangeli, Fabrizio, Elisa De Luca, Donatella Magri, Simone De Santis, Alessandra Celant, Matthieu Ghilardi, Matteo Vacchi, Jordi Revelles, Rita Teresa Melis, Juan Ochando, and et al. 2025. "Holocene Forest Dynamics in Western Mediterranean Islands: Rates, Periodicity, and Trends" Forests 16, no. 5: 808. https://doi.org/10.3390/f16050808
APA StyleMichelangeli, F., De Luca, E., Magri, D., De Santis, S., Celant, A., Ghilardi, M., Vacchi, M., Revelles, J., Melis, R. T., Ochando, J., Carrión, J., Pini, R., Servera-Vives, G., & Di Rita, F. (2025). Holocene Forest Dynamics in Western Mediterranean Islands: Rates, Periodicity, and Trends. Forests, 16(5), 808. https://doi.org/10.3390/f16050808