
Brenneria goodwinii and Gibbsiella quercinecans as a Threat to Quercus coccifera L.
 ,
,  , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling and Symptom Assessment
2.2. Plant Diagnosis
2.3. Bacterial Isolation and 16S Barcoding
2.4. Pathogenicity Tests
3. Results
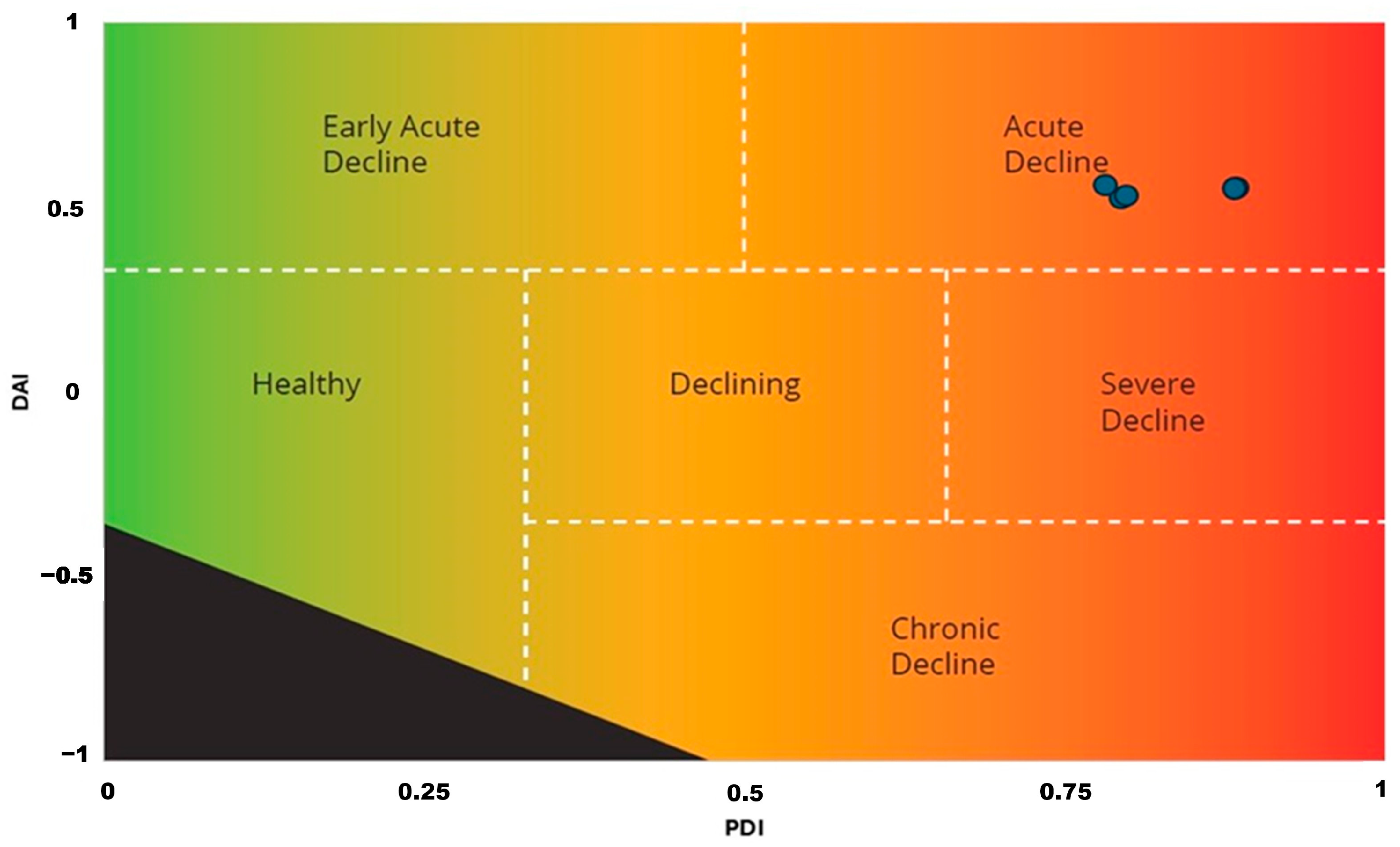
3.1. Symptomatology and Diagnosis
3.2. Isolation, Sequencing, and Phylogenetic Analysis
3.3. Pathogenicity Assessments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Martínez-Ferri, E.; Balaguer, L.; Valladares, F.; Chico, J.M.; Manrique, E. Energy Dissipation in Drought-Avoiding and Drought-Tolerant Tree Species at Midday during the Mediterranean Summer. Tree Physiol. 2000, 20, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Cañellas, I.; San Miguel, A. Litter Fall and Nutrient Turnover in Kermes Oak (Quercus coccifera L.) Shrublands in Valencia (Eastern Spain). Ann. Sci. For. 1998, 55, 589–597. [Google Scholar] [CrossRef]
- Jasprica, N.; Škvorc, Ž.; Dolina, K.; Ruščić, M.; Kovačić, S.; Franjić, J. Composition and Ecology of the Quercus coccifera L. Communities along the Eastern Adriatic Coast (NE Mediterranean). Plant Biosyst. 2016, 150, 1140–1155. [Google Scholar] [CrossRef]
- Rodà, F.; Retana, J.; Gracia, C.A.; Bellot, J. Ecology of Mediterranean Evergreen Oak Forests; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- Tsiouvaras, C.N. Ecology and Management of Kermes Oak (Quercus coccifera L.) Shrublands in Greece: A Review. J. Range Manag. 1987, 40, 542. [Google Scholar] [CrossRef]
- FO.RE.S.T.A.S. Sardegna Foreste–Quercia Spinosa. 2024. Available online: https://www.sardegnaforeste.it/flora/quercia-spinosa (accessed on 15 November 2024).
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological Maps for the Main European Woody Species. Data Br. 2017, 12, 662–666. [Google Scholar] [CrossRef]
- EFI. EUFORGEN- Quercus Coccifera. 2020. Available online: https://www.euforgen.org/species/quercus-coccifera (accessed on 26 November 2024).
- Ruffner, B.; Schneider, S.; Meyer, J.B.; Queloz, V.; Rigling, D. First Report of Acute Oak Decline Disease of Native and Non-native Oaks in Switzerland. New Dis. Rep. 2020, 41, 18. [Google Scholar] [CrossRef]
- Tkaczyk, M.; Celma, L.; Ruņģis, D.E.; Bokuma, G. First Report of Brenneria Goodwinii and Gibbsiella Quercinecans Bacteria, Detected on Weaken Oak Trees in Poland. Balt. For. 2021, 27, 166–169. [Google Scholar] [CrossRef]
- Tkaczyk, M.; Sikora, K.; Plewa, R. Dieback of Small-Leaved Lime Trees (Tilia Cordata Mill.) Caused by Gibsiella quercinecans in Urban Areas in Poland. For. Pathol. 2024, 54, e12861. [Google Scholar] [CrossRef]
- Fernandes, C.; Duarte, L.; Naves, P.; Sousa, E.; Cruz, L. First Report of Brenneria goodwinii Causing Acute Oak Decline on Quercus suber in Portugal. J. Plant Pathol. 2022, 104, 837–838. [Google Scholar] [CrossRef]
- Carluccio, G.; Sabella, E.; Greco, D.; Vergine, M.; Delle Donne, A.G.; Nutricati, E.; Aprile, A.; De Bellis, L.; Luvisi, A. Acute and Chronic Oak Decline in Urban and Forest Ecosystems in Southern Italy. For. Int. J. For. Res. 2024, 97, 739–749. [Google Scholar] [CrossRef]
- Brady, C.; Denman, S.; Kirk, S.; Venter, S.; Rodríguez-Palenzuela, P.; Coutinho, T. Description of Gibbsiella quercinecans Gen. Nov., Sp. Nov., Associated with Acute Oak Decline. Syst. Appl. Microbiol. 2010, 33, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Denman, S.; Brady, C.; Kirk, S.; Cleenwerck, I.; Venter, S.; Coutinho, T.; De Vos, P. Brenneria goodwinii Sp. Nov., Associated with Acute Oak Decline in the UK. Int. J. Syst. Evol. Microbiol. 2012, 62, 2451–2456. [Google Scholar] [CrossRef] [PubMed]
- Brady, C.; Arnold, D.; McDonald, J.; Denman, S. Taxonomy and Identification of Bacteria Associated with Acute Oak Decline. World J. Microbiol. Biotechnol. 2017, 33, 143. [Google Scholar] [CrossRef]
- Denman, S.; Kirk, S.; Webber, J. Managing Acute Oak Decline; FC Practice Note 15; FC: Edinburgh, Scotland, 2010; p. 6. [Google Scholar]
- Denman, S.; Brown, N.; Kirk, S.; Jeger, M.; Webber, J. A Description of the Symptoms of Acute Oak Decline in Britain and a Comparative Review on Causes of Similar Disorders on Oak in Europe. Forestry 2014, 87, 535–551. [Google Scholar] [CrossRef]
- Broberg, M.; Doonan, J.; Mundt, F.; Denman, S.; McDonald, J.E. Integrated Multi-Omic Analysis of Host-Microbiota Interactions in Acute Oak Decline. Microbiome 2018, 6, 21. [Google Scholar] [CrossRef]
- González, A.J.; Ciordia, M. Brenneria Goodwinii and Gibbsiella Quercinecans Isolated from Weeping Cankers on Quercus robur L. in Spain. Eur. J. Plant Pathol. 2020, 156, 965–969. [Google Scholar] [CrossRef]
- Zalkalns, O.; Celma, L. The Distribution of Bacteria Gibbsiella quercinecans and Brenneria goodwinii in Oak (Quercus robur L.) Stands in Latvia. IOP Conf. Ser. Earth Environ. Sci. 2021, 875, 012033. [Google Scholar] [CrossRef]
- Finch, J.P.; Brown, N.; Beckmann, M.; Denman, S.; Draper, J. Index Measures for Oak Decline Severity Using Phenotypic Descriptors. For. Ecol. Manag. 2021, 485, 118948. [Google Scholar] [CrossRef]
- Crampton, B.G.; Plummer, S.J.; Kaczmarek, M.; McDonald, J.E.; Denman, S. A Multiplex Real-time PCR Assay Enables Simultaneous Rapid Detection and Quantification of Bacteria Associated with Acute Oak Decline. Plant Pathol. 2020, 69, 1301–1310. [Google Scholar] [CrossRef]
- Moradi-Amirabad, Y.; Rahimian, H.; Babaeizad, V.; Denman, S. Brenneria Spp. and Rahnella victoriana Associated with Acute Oak Decline Symptoms on Oak and Hornbeam in Iran. For. Pathol. 2019, 49, e12535. [Google Scholar] [CrossRef]
- Pettifor, B. Survival of Brenneria Goodwinii and Gibbsiella quercinecans, Associated with Lesion Formation in Acute Oak Decline, in Rainwater and Forest Soil. Master’s Thesis, Bangor University, Gwynedd, UK, 2019; pp. 1–90. [Google Scholar]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Wiley: Chichester, England, 1991; pp. 115–175. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J Mol Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Eichenlaub, L.; Denman, S.; Brady, C.; Maddock, D.; Robledo-Garcia, F.; Aubert, A.; Husson, C.; Robin, C. First Report of Brenneria goodwinii, Gibbsiella quercinecans and Rahnella victoriana in Declining Oaks in France. New Dis. Rep. 2024, 49, 2023–2024. [Google Scholar] [CrossRef]
- Carluccio, G.; Vergine, M.; Vita, F.; Sabella, E.; Delle Donne, A.; De Bellis, L.; Luvisi, A. Long-Distance Finding of AOD-Related Bacteria in the Natural Environment: Risks to Quercus ilex (L.) in Italy. Forests 2024, 15, 2055. [Google Scholar] [CrossRef]
- Denman, S.; Doonan, J.; Ransom-Jones, E.; Broberg, M.; Plummer, S.; Kirk, S.; Scarlett, K.; Griffiths, A.R.; Kaczmarek, M.; Forster, J.; et al. Microbiome and Infectivity Studies Reveal Complex Polyspecies Tree Disease in Acute Oak Decline. ISME J. 2018, 12, 386–399. [Google Scholar] [CrossRef]
- Maddock, D.; Brady, C.; Denman, S.; Arnold, D. Bacteria Associated with Acute Oak Decline: Where Did They Come From? We Know Where They Go. Microorganisms 2023, 11, 2789. [Google Scholar] [CrossRef]
- Wang, L.; Li, C.; Wang, X.; Wang, G.; Shang, S.; Dou, Z.; Luo, Y. Gut Lignocellulose Activity and Microbiota in Asian Longhorned Beetle and Their Predicted Contribution to Larval Nutrition. Front. Microbiol. 2022, 13, 899865. [Google Scholar] [CrossRef]
- Wang, X.; Wang, H.; Zeng, J.; Cui, Z.; Geng, S.; Song, X.; Zhang, F.; Su, X.; Li, H. Distinct Gut Bacterial Composition in Anoplophora Glabripennis Reared on Two Host Plants. Front. Microbiol. 2023, 14, 1199994. [Google Scholar] [CrossRef]
- Zhang, H. Abiotic Stress Responses in Plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Le Floch, A.; Jourdes, M.; Teissedre, P.L. Polysaccharides and Lignin from Oak Wood Used in Cooperage: Composition, Interest, Assays: A Review. Carbohydr. Res. 2015, 417, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Daniel, G. Variations in Cell Wall Ultrastructure and Chemistry in Cell Types of Earlywood and Latewood in English Oak (Quercus robur). IAWA J. 2016, 37, 383–401. [Google Scholar] [CrossRef]
- Martínez, M.T.; Cuenca, B.; Mosteiro, F.; Piñeiro, P.; Pérez, F.; Solla, A.; Corredoira, E. Screening of Cork Oak for Resistance to Phytophthora cinnamomi and Micropropagation of Tolerant Seedlings. Horticulturae 2023, 9, 692. [Google Scholar] [CrossRef]
- Brown, N.; Jeger, M.; Kirk, S.; Xu, X.; Denman, S. Spatial and Temporal Patterns in Symptom Expression within Eight Woodlands Affected by Acute Oak Decline. For. Ecol. Manag. 2016, 360, 97–109. [Google Scholar] [CrossRef]
- Ducousso, A.; Bordacs, S. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Pedunculate and Sessile Oaks (Quercus robur and Q. petraea); The International Plant Genetic Resources Institute: Maccarese, Italy, 2004; pp. 1–6. [Google Scholar]
- Basavand, E.; Khodaygan, P.; Doonan, J.M.; Rahimian, H. Gibbsiella quercinecans as New Pathogen Involved in Bacterial Canker of Russian Olive. 3 Biotech 2021, 11, 286. [Google Scholar] [CrossRef]
- Allahverdipour, T.; Shahryari, F.; FalahiCharkhabi, N. Gibbsiella quercinecans and Brenneria roseae subsp. roseae Associated to the Canker Disease of Walnut Trees in Northwestern Iran. Eur. J. Plant Pathol. 2021, 161, 783–797. [Google Scholar] [CrossRef]
- Brown, N.; Inward, D.J.G.; Jeger, M.; Denman, S. A Review of A Review of Agrilus biguttatus in UK Forests and Its Relationship with Acute Oak Decline. Forestry 2015, 88, 53–63. [Google Scholar] [CrossRef]
- Moraal, L.G.; Hilszczanski, J. The Oak Buprestid Beetle, Agrilus biguttatus (F.) (Col., Buprestidae), a Recent Factor in Oak Decline in Europe. Anz. Schädlingskunde 2000, 73, 134–138. [Google Scholar] [CrossRef]
- Sallé, A. Native buprestid and longhorn beetles in the Mediterranean Basin. In Insects and Diseases of Mediterranean Forest Systems; Springer International Publishing: Cham, Germany, 2016; pp. 199–210. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | Primers and Probes | Sequence (5′–3′) |
|---|---|---|
| Brenneria goodwinii | Bg99F Bg179R Bg179P | CTGGCCGAGCCTGGAAAC AGTTCAGGAAGGAGAGTTCGC CCAGAATCTCATATTCGAACTCCACCATGTT |
| Gibsiella quercinecans | Gq284F Gq418R Gq342P | GGCTTTGATAGTGGTGGCC CGTTCCGTTATCACCGTGG AACAGTTCCAGCGCCATTTTCTTCG |
| Rahnella victoriana | Rv15F Rv134R Rv57P | CACCCAGACTTACGTGCAT TCAGTGTGATTGGTGAAGGT AGTGATTGGCGATACTGACGTGACC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carluccio, G.; Vergine, M.; De Pascali, M.; Bene, A.; Portaccio, L.; Delle Donne, A.; De Bellis, L.; Luvisi, A. Brenneria goodwinii and Gibbsiella quercinecans as a Threat to Quercus coccifera L. Forests 2025, 16, 789. https://doi.org/10.3390/f16050789
Carluccio G, Vergine M, De Pascali M, Bene A, Portaccio L, Delle Donne A, De Bellis L, Luvisi A. Brenneria goodwinii and Gibbsiella quercinecans as a Threat to Quercus coccifera L. Forests. 2025; 16(5):789. https://doi.org/10.3390/f16050789
Chicago/Turabian StyleCarluccio, Giambattista, Marzia Vergine, Mariarosaria De Pascali, Alessandro Bene, Letizia Portaccio, Angelo Delle Donne, Luigi De Bellis, and Andrea Luvisi. 2025. "Brenneria goodwinii and Gibbsiella quercinecans as a Threat to Quercus coccifera L." Forests 16, no. 5: 789. https://doi.org/10.3390/f16050789
APA StyleCarluccio, G., Vergine, M., De Pascali, M., Bene, A., Portaccio, L., Delle Donne, A., De Bellis, L., & Luvisi, A. (2025). Brenneria goodwinii and Gibbsiella quercinecans as a Threat to Quercus coccifera L. Forests, 16(5), 789. https://doi.org/10.3390/f16050789