


Soil Moisture Loss in Planted Forests and Its Driving Factors: A Case Study of the Nanpan River Basin
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Research Data and Methodology
2.2.1. Soil Texture Data
2.2.2. Soil Moisture and Meteorological Data
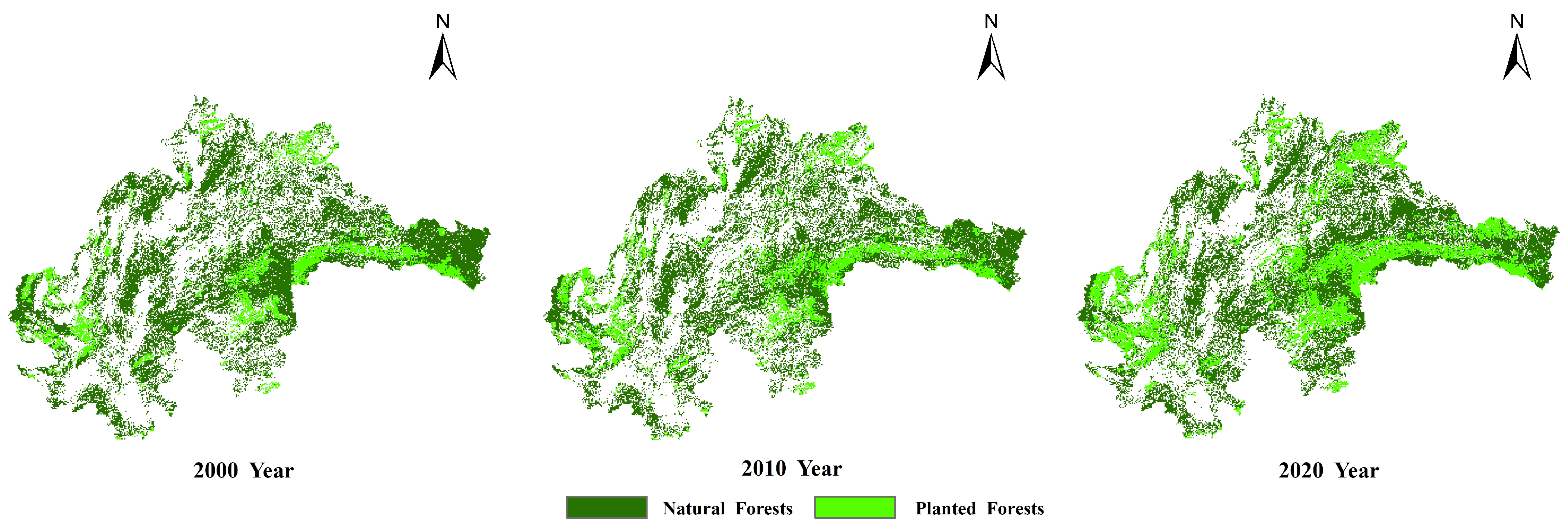
2.2.3. Data on Planted and Natural Forests
2.2.4. Linear Trend Analysis
- A.
- Calculation of Trend Slopes
- B.
- Trend Significance Testing
2.2.5. Geodetectors
3. Results
3.1. Spatiotemporal Evolution Characteristics of Soil Moisture
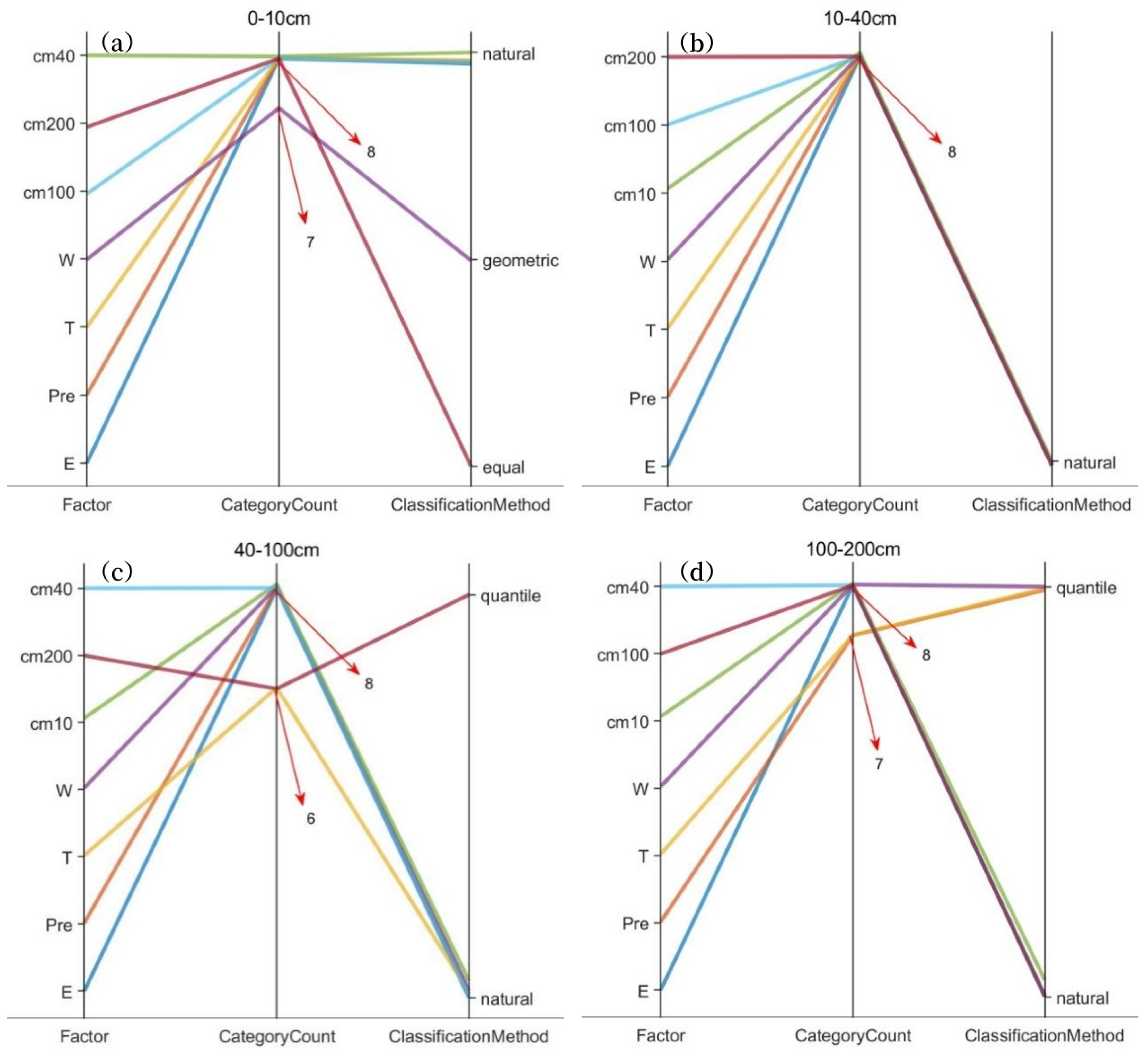
3.2. Soil Moisture Responses to Composite Factors by Forest Attributes
3.2.1. Soil Moisture Response in Horizontal Soil Layers by Forest Attributes
3.2.2. Soil Moisture Response in Vertical Soil Layers by Forest Attributes
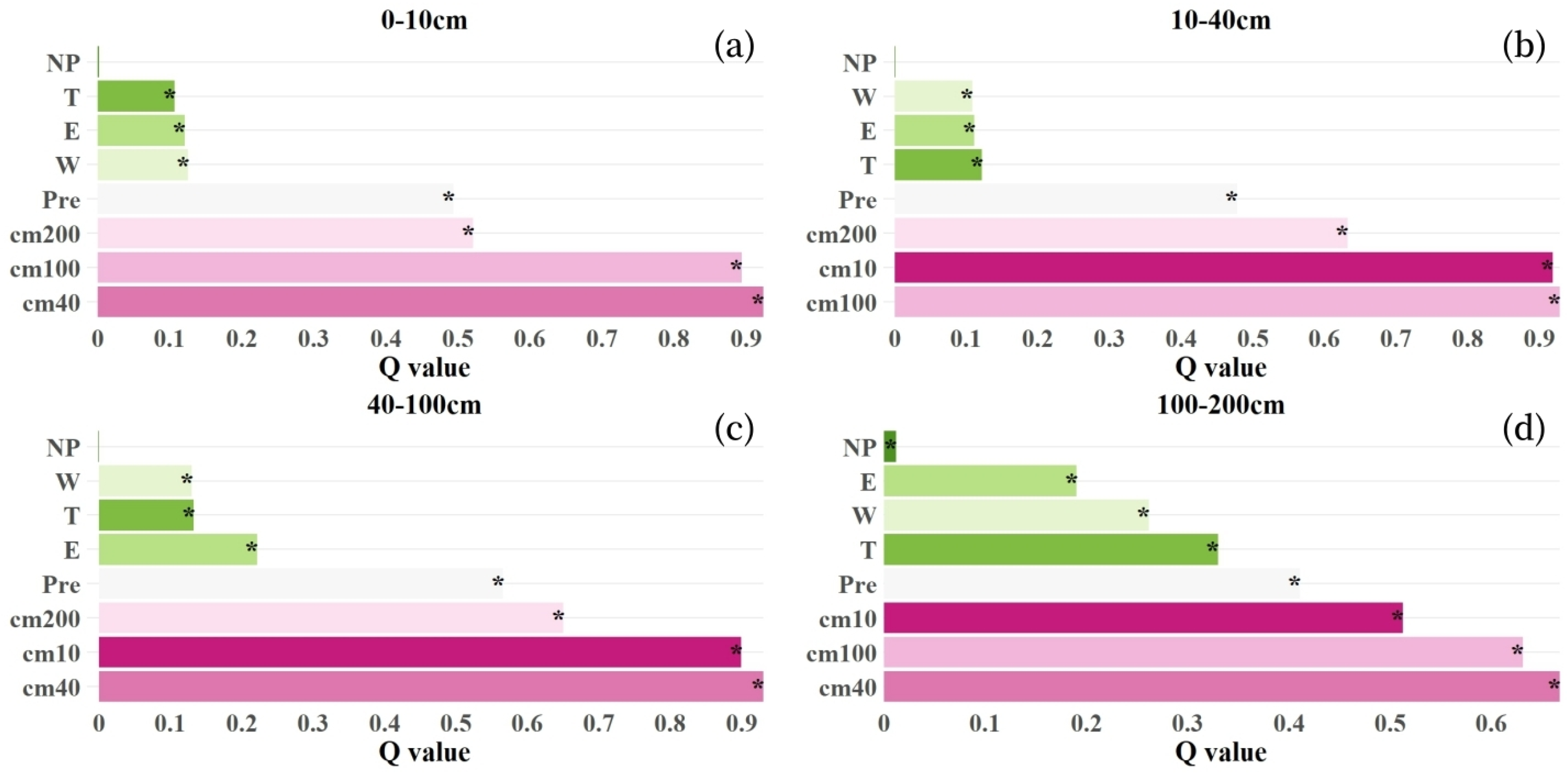
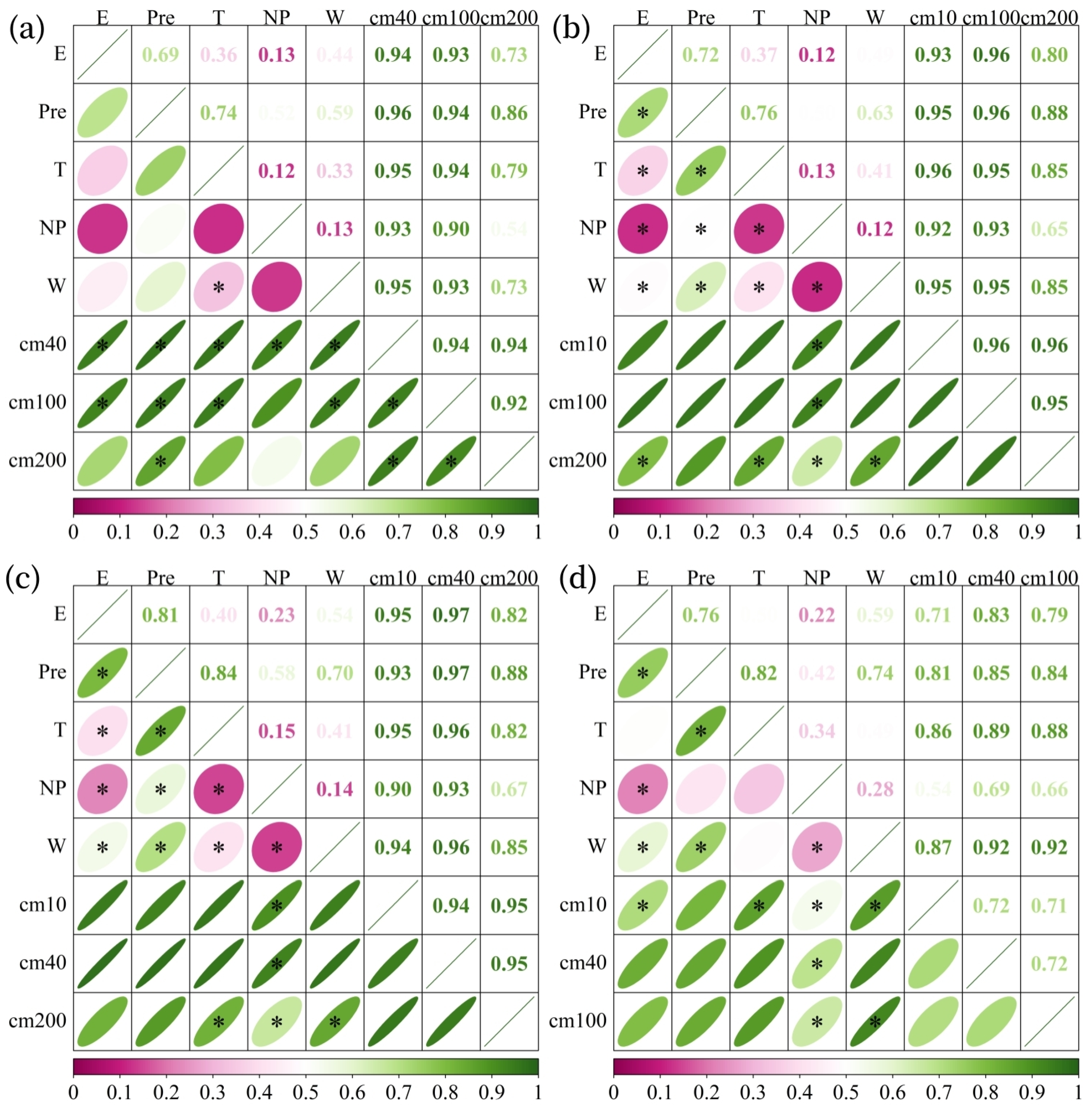
3.3. Driving Factors of Soil Moisture Changes
3.3.1. Spatial Pattern Differentiation and Factor Detection
3.3.2. Interaction Effects on the Spatial Patterns
4. Discussion
4.1. Analysis of Data and Model Applicability
4.2. Response of Soil Moisture to Soil Texture
4.3. Driving Factors of Soil Moisture Content Variation
4.4. Limitations and Future Work
5. Conclusions
- (1)
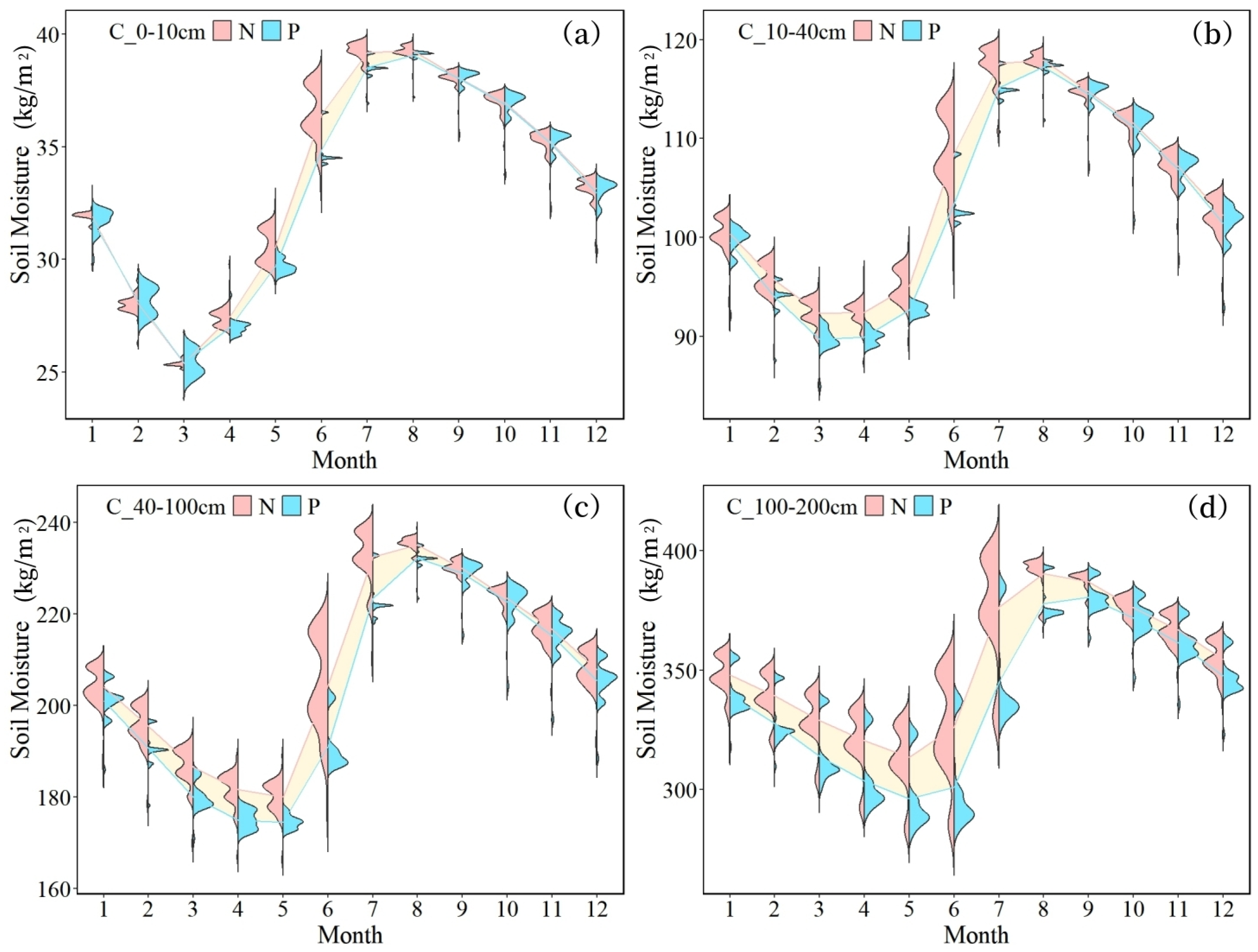
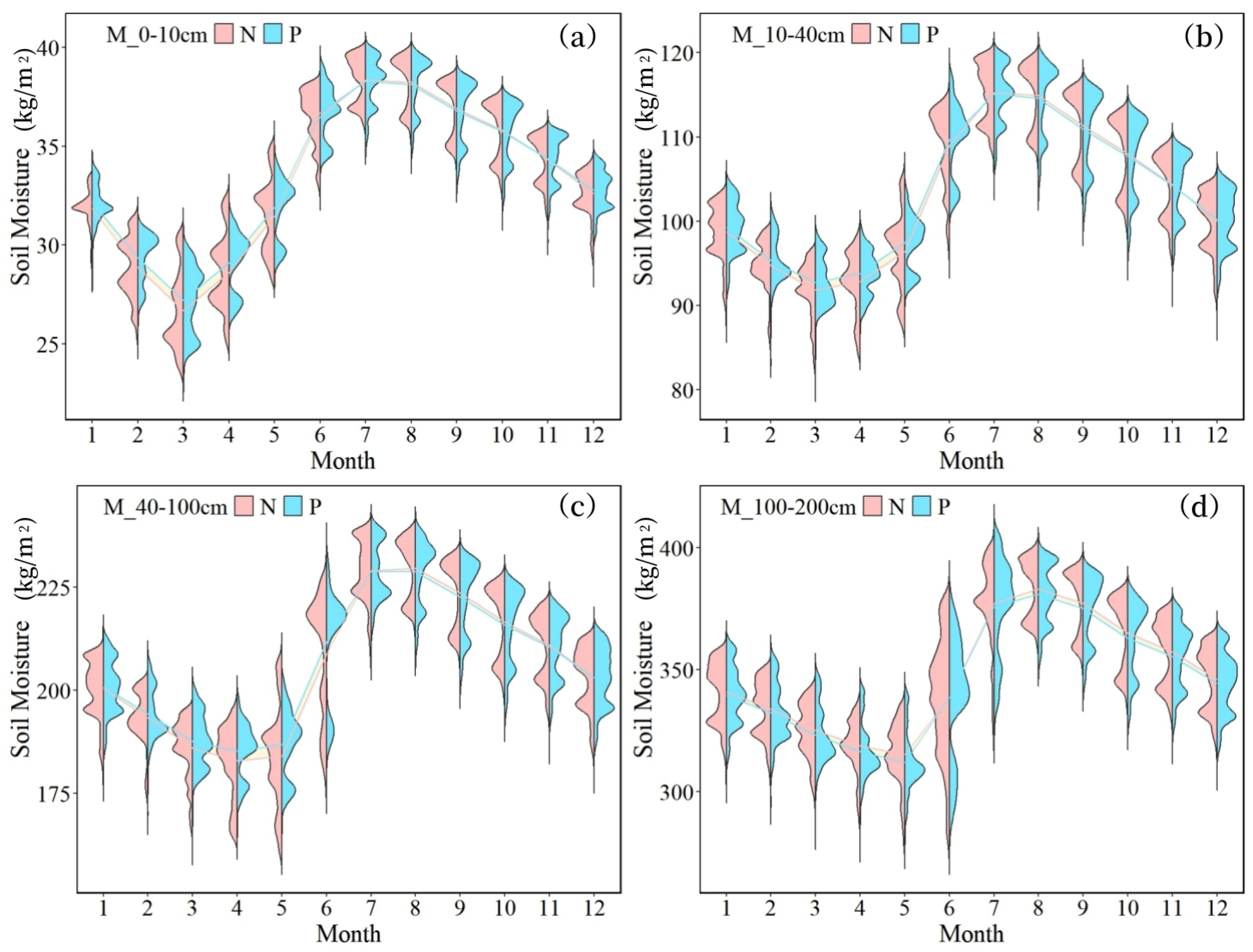
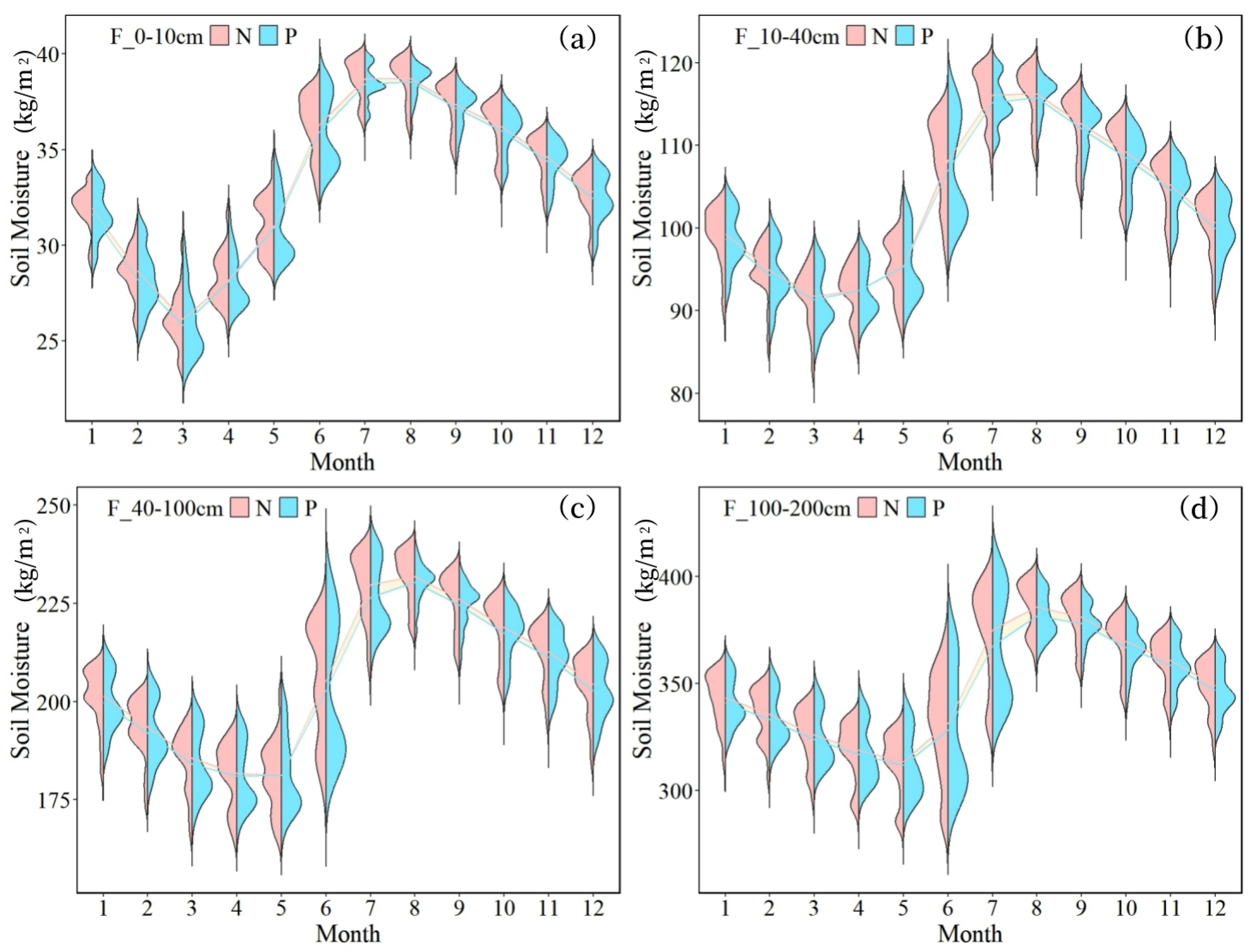
- The overall soil moisture consumption in planted forests is greater than that in natural forests, with variations in moisture consumption differing across soil textures. The difference is most pronounced in coarse-textured soils. In June, the soil moisture content in plantations was consistently lower than in natural forests across all soil layers, with measured differences of 1.60 kg/m2 (0–10 cm), 5.07 kg/m2 (10–40 cm), 13.35 kg/m2 (40–100 cm), and 31.31 kg/m2 (100–200 cm).
- (2)
- The spatial differentiation of mean soil moisture content across soil layers is primarily determined by the moisture content in the subsoil layer (10–40 cm), which exhibits Q-values of 92.36% (0–10 cm), 93.00% (40–100 cm), and 66.72% (100–200 cm). Precipitation constitutes the secondary influencing factor, while the surface 0–10 cm soil layer shows greater susceptibility to wind speed effects.
- (3)
- Notably, only the soil moisture in the 100–200 cm layer is affected by forest attributes. This is because the dominant planted species in the basin, such as Eucalyptus and Cunninghamia, have main root systems concentrated at depths of 100–200 cm. In this layer, the substantial water consumption by planted forests leads to spatial differentiation in soil moisture.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Pre | precipitation |
| T | temperature |
| W | wind speed |
| NP | forest attributes |
| E | evapotranspiration |
References
- Xue, S.; Wu, G. Causal inference of root zone soil moisture performance in drought. Agric. Water Manag. 2024, 305, 109123. [Google Scholar] [CrossRef]
- Luo, M.; Meng, F.; Sa, C.; Duan, Y.; Bao, Y.; Liu, T.; De Maeyer, P. Response of vegetation phenology to soil moisture dynamics in the Mongolian Plateau. CATENA 2021, 206, 105505. [Google Scholar] [CrossRef]
- Zhang, D.-H.; Li, X.-R.; Zhang, F.; Zhang, Z.-S.; Chen, Y.-L. Effects of rainfall intensity and intermittency on woody vegetation cover and deep soil moisture in dryland ecosystems. J. Hydrol. 2016, 543, 270–282. [Google Scholar] [CrossRef]
- Demir, G.; Guswa, A.J.; Filipzik, J.; Metzger, J.C.; Römermann, C.; Hildebrandt, A. Root water uptake patterns are controlled by tree species interactions and soil water variability. Hydrol. Earth Syst. Sci. 2024, 28, 1441–1461. [Google Scholar] [CrossRef]
- Mi, J.; Xiao, X.; Guan, Q.; Wang, Q.; Zhang, J.; Zhang, Z.; Yang, E. Exploring the spatiotemporal distribution characteristics and driving factors of water erosion in mountain area based on RUSLE-SDR. J. Hydrol. 2025, 649, 132451. [Google Scholar] [CrossRef]
- Mei, X.-m.; Ma, L.; Zhu, Q.-k.; Wang, S.; Zhang, D.; Wang, Y. Responses of soil moisture to vegetation restoration type and slope length on the loess hillslope. J. Mt. Sci. 2018, 15, 548–562. [Google Scholar] [CrossRef]
- Meng, T.; Sun, P. Variations of deep soil moisture under different vegetation restoration types in a watershed of the Loess Plateau, China. Sci. Rep. 2023, 13, 4957. [Google Scholar] [CrossRef]
- Tian, R.; Li, J.; Zheng, J.; Liu, L.; Han, W.; Liu, Y. Changes in vegetation phenology and its response to different layers of soil moisture in the dry zone of Central Asia, 1982–2022. J. Hydrol. 2025, 646, 132314. [Google Scholar] [CrossRef]
- Molenaar, R.E.; Kleidorfer, M.; Kohl, B.; Teuling, A.J.; Achleitner, S. Does afforestation increase soil water buffering? A demonstrator study on soil moisture variability in the Alpine Geroldsbach catchment, Austria. J. Hydrol. 2024, 643, 131984. [Google Scholar] [CrossRef]
- Fang, X.; Zhao, W.; Wang, L.; Feng, Q.; Ding, J.; Liu, Y.; Zhang, X. Variations of deep soil moisture under different vegetation types and influencing factors in a watershed of the Loess Plateau, China. Hydrol. Earth Syst. Sci. 2016, 20, 3309–3323. [Google Scholar] [CrossRef]
- Geng, J.; Li, H.; Shi, Y.; Pang, J.; Shao, Y. Comparison and Optimization of Water Conservation Function in Natural Forests and Tea Plantations in a Headwater Catchment, Taihu Lake Basin. Forests 2024, 15, 189. [Google Scholar] [CrossRef]
- Hou, L.; Zhang, Y.; Li, Z.; Shao, G.; Song, L.; Sun, Q. Comparison of Soil Properties, Understory Vegetation Species Diversities and Soil Microbial Diversities Between Chinese Fir Plantation and Close-to-Natural Forest. Forests 2021, 12, 632. [Google Scholar] [CrossRef]
- Liu, X.; Feng, T.; Zhang, Y.; Liu, Y.; Wang, P. Vegetation restoration affects soil hydrological processes in typical natural and planted forests on the Loess Plateau. J. Hydrol. 2025, 650, 132465. [Google Scholar] [CrossRef]
- Oggioni, S.D.; Rossi, L.M.W.; Avanzi, C.; Marchetti, M.; Piotti, A.; Vacchiano, G. Drought responses of Italian silver fir provenances in a climate change perspective. Dendrochronologia 2024, 85, 126184. [Google Scholar] [CrossRef]
- Bonell, M.; Purandara, B.K.; Venkatesh, B.; Krishnaswamy, J.; Acharya, H.A.K.; Singh, U.V.; Jayakumar, R.; Chappell, N. The impact of forest use and reforestation on soil hydraulic conductivity in the Western Ghats of India: Implications for surface and sub-surface hydrology. J. Hydrol. 2010, 391, 47–62. [Google Scholar] [CrossRef]
- Jaroszewicz, B.; Cholewińska, O.; Chećko, E.; Wrzosek, M. Predictors of diversity of deadwood-dwelling macrofungi in a European natural forest. For. Ecol. Manag. 2021, 490, 119123. [Google Scholar] [CrossRef]
- Qiu, D.; Zhu, G.; Lin, X.; Jiao, Y.; Lu, S.; Liu, J.; Liu, J.; Zhang, W.; Ye, L.; Li, R.; et al. Dissipation and movement of soil water in artificial forest in arid oasis areas: Cognition based on stable isotopes. CATENA 2023, 228, 107178. [Google Scholar] [CrossRef]
- Zheng, J.; Jin, X.; Li, Q.; Lang, J.; Yin, X. Soil moisture variation and affecting factors analysis in the Zhangjiakou–Chengde district based on modified temperature vegetation dryness index. Ecol. Indic. 2024, 168, 112775. [Google Scholar] [CrossRef]
- Liang, X.; Yan, J.; Liang, W.; Li, B.; Liu, X.; Feng, F.; Wei, J. Ecosystem water limitation shifts driven by soil moisture in the Loess Plateau, China. Glob. Planet. Change 2024, 243, 104625. [Google Scholar] [CrossRef]
- Gu, X.; Jamshidi, S.; Sun, H.; Niyogi, D. Identifying multivariate controls of soil moisture variations using multiple wavelet coherence in the U.S. Midwest. J. Hydrol. 2021, 602, 126755. [Google Scholar] [CrossRef]
- Wen, Y.; Li, M.; Xu, R.; Qiu, D.; Gao, P.; Mu, X. Spatial distribution characteristics and influencing factors of shallow and deep soil moisture under ecological restoration in the loess plateau, China. Hydrol. Process. 2024, 38, e15109. [Google Scholar] [CrossRef]
- Wang, J.B.; Peng, J.; Zhang, R.X.; Xu, Z.H.; Liu, Y.X. Research progress and prospect of karst ecosystem services. J. Guizhou Norm. Univ. (Nat. Sci.) 2024, 42, 77–84. [Google Scholar] [CrossRef]
- De Lannoy, G.J.M.; Koster, R.D.; Reichle, R.H.; Mahanama, S.P.P.; Liu, Q. An updated treatment of soil texture and associated hydraulic properties in a global land modeling system. J. Adv. Model. Earth Syst. 2014, 6, 957–979. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; Wang, W. Inter-comparison of satellite-retrieved and Global Land Data Assimilation System-simulated soil moisture datasets for global drought analysis. Remote Sens. Environ. 2019, 220, 1–18. [Google Scholar] [CrossRef]
- Zhang, Z.; Song, Y.; Wu, P. Robust geographical detector. Int. J. Appl. Earth Obs. Geoinf. 2022, 109, 102782. [Google Scholar] [CrossRef]
- Wang, J.-F.; Zhang, T.-L.; Fu, B.-J. A measure of spatial stratified heterogeneity. Ecol. Indic. 2016, 67, 250–256. [Google Scholar] [CrossRef]
- Sun, X.; Lai, P.; Wang, S.; Song, L.; Ma, M.; Han, X.J. Monitoring of Extreme Agricultural Drought of the Past 20 Years in Southwest China Using GLDAS Soil Moisture. Remote Sens. 2022, 14, 1323. [Google Scholar] [CrossRef]
- Guan, Y.; Gu, X.; Slater, L.J.; Li, J.; Kong, D.; Zhang, X. Spatio-temporal variations in global surface soil moisture based on multiple datasets: Intercomparison and climate drivers. J. Hydrol. 2023, 625, 130095. [Google Scholar] [CrossRef]
- Cheng, K.; Su, Y.; Guan, H.; Tao, S.; Ren, Y.; Hu, T.; Ma, K.; Tang, Y.; Guo, Q. Mapping China’s planted forests using high resolution imagery and massive amounts of crowdsourced samples. ISPRS J. Photogramm. Remote Sens. 2023, 196, 356–371. [Google Scholar] [CrossRef]
- Cheng, K.; Yang, H.; Tao, S.; Su, Y.; Guan, H.; Ren, Y.; Hu, T.; Li, W.; Xu, G.; Chen, M.; et al. Carbon storage through China’s planted forest expansion. Nat. Commun. 2024, 15, 4106. [Google Scholar] [CrossRef]
- Hillel, D. Fundamentals of Soil Physics; Academic Press: Cambridge, MA, USA, 1980. [Google Scholar] [CrossRef]
- Graetz, D.A. Biogeochemistry: An Analysis of Global Change. J. Environ. Qual. 1992, 21, 153–154. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Jactel, H.; Parrotta, J.A.; Quine, C.P.; Sayer, J. Plantation forests and biodiversity: Oxymoron or opportunity? Biodivers. Conserv. 2008, 17, 925–951. [Google Scholar] [CrossRef]
- Farley, K.A.; Jobbágy, E.G.; Jackson, R.B. Effects of afforestation on water yield: A global synthesis with implications for policy. Glob. Change Biol. 2005, 11, 1565–1576. [Google Scholar] [CrossRef]
- Li, S.L.Y.; Zhou, M. Relationship between Vertical Root Distribution of Planted Eucalyptus and Soil Bulk Density and Moisture. Soil Water Conserv. China 2022, 10, 67–70. [Google Scholar] [CrossRef]
- Daniels, W.L. The Nature and Properties of Soils, 15th Edition Ray R. Weil and Nyle C. Brady. Pearson Press, Upper Saddle River NJ, 2017. 1086 p. $164.80. ISBN-10: 0-13-325448-8; ISBN-13: 978-0-13-325448-8. Also available as eText for $67.99. Soil Sci. Soc. Am. J. 2016, 80, 1428. [Google Scholar] [CrossRef]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils. J. Range Manag. 2002, 5, 333. [Google Scholar] [CrossRef]
- Lin, H. Earth’s Critical Zone and hydropedology: Concepts, characteristics, and advances. Hydrol. Earth Syst. Sci. 2010, 6, 3417–3481. [Google Scholar] [CrossRef]
- Monteith, J.L. Principles of environmental physics. Phys. Today 1974, 27, 51. [Google Scholar] [CrossRef]
- Yeh, J.F.; Eltahir, E.A.B. Stochastic analysis of the relationship between topography and the spatial distribution of soil moisture. Water Resour. Res. 1998, 34, 2075. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, H.; Cui, W.; He, Z.; Yang, M.; Tan, H.; Yang, Q. Soil Moisture Loss in Planted Forests and Its Driving Factors: A Case Study of the Nanpan River Basin. Forests 2025, 16, 665. https://doi.org/10.3390/f16040665
Yu H, Cui W, He Z, Yang M, Tan H, Yang Q. Soil Moisture Loss in Planted Forests and Its Driving Factors: A Case Study of the Nanpan River Basin. Forests. 2025; 16(4):665. https://doi.org/10.3390/f16040665
Chicago/Turabian StyleYu, Huan, Wengang Cui, Zhonghua He, Mei Yang, Hongmei Tan, and Qiuyun Yang. 2025. "Soil Moisture Loss in Planted Forests and Its Driving Factors: A Case Study of the Nanpan River Basin" Forests 16, no. 4: 665. https://doi.org/10.3390/f16040665
APA StyleYu, H., Cui, W., He, Z., Yang, M., Tan, H., & Yang, Q. (2025). Soil Moisture Loss in Planted Forests and Its Driving Factors: A Case Study of the Nanpan River Basin. Forests, 16(4), 665. https://doi.org/10.3390/f16040665