
Comparison of the Microbiome and Metabolome in the Phloem of Straight and Twisted Trunks of Pinus yunnanensis Franch

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Processing
2.2. DNA Extraction and HIGH-Throughput Sequencing
2.3. Statistical Analysis of Microbiota
2.4. Extraction of Phloem Metabolites
2.5. Metabolites Detection
2.6. Statistical Analysis of Metabolomic Data
2.7. Correlation Between Microbiota and Metabolome of P. yunnanensis
3. Results
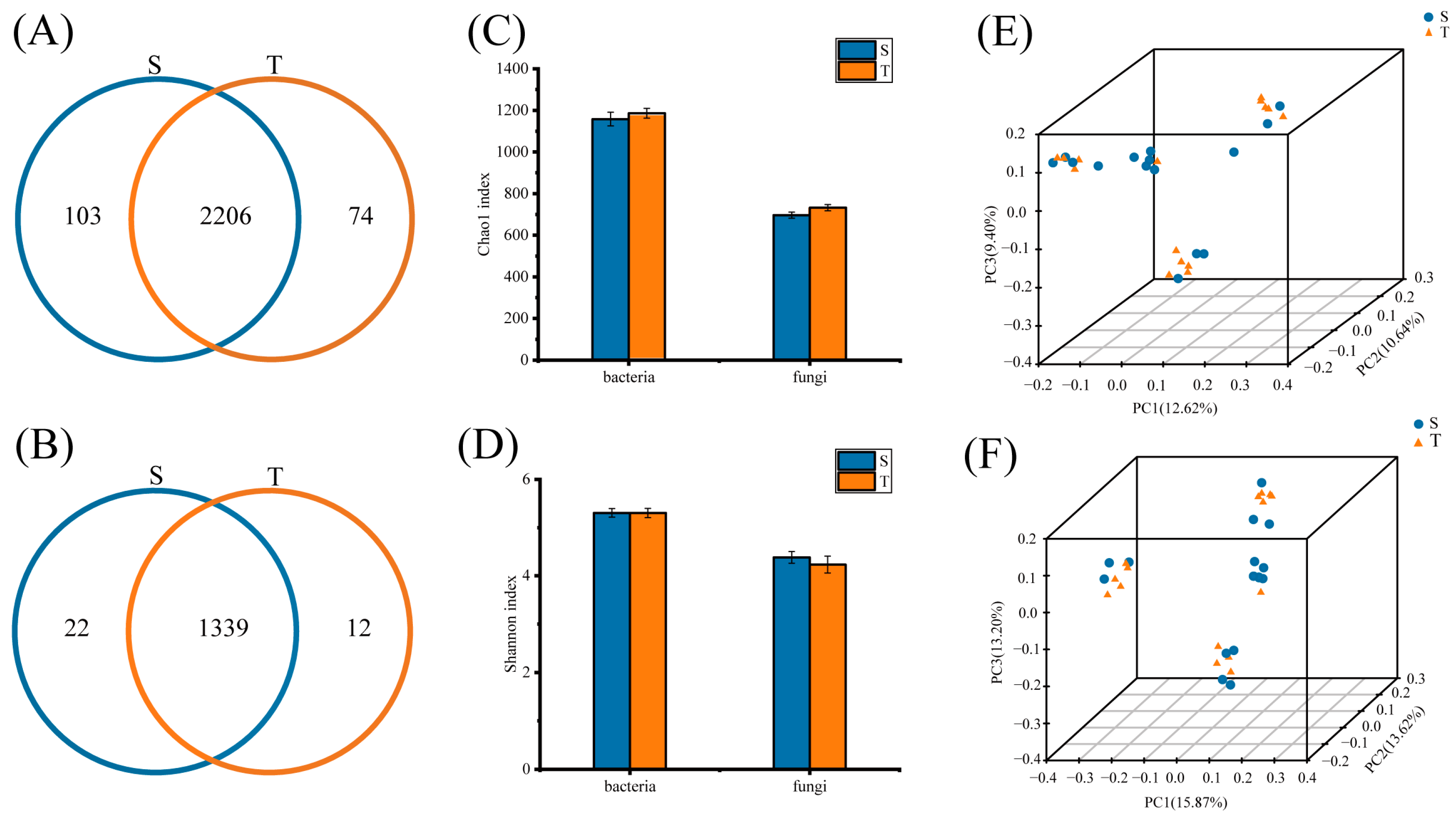
3.1. Sequencing and Phloem Microbial Diversity Analysis
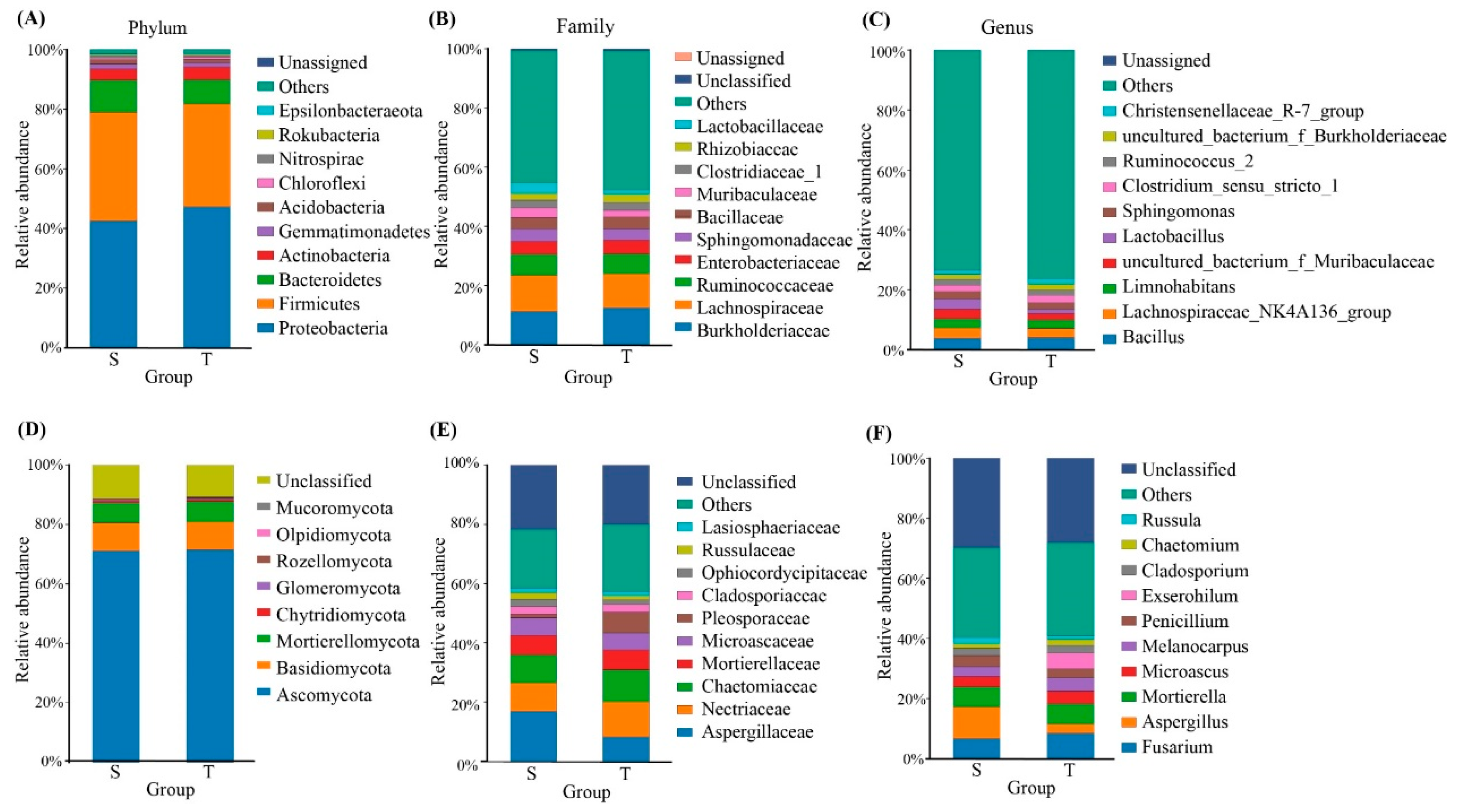
3.2. Community Composition of Phloem Bacterial and Fungal Communities
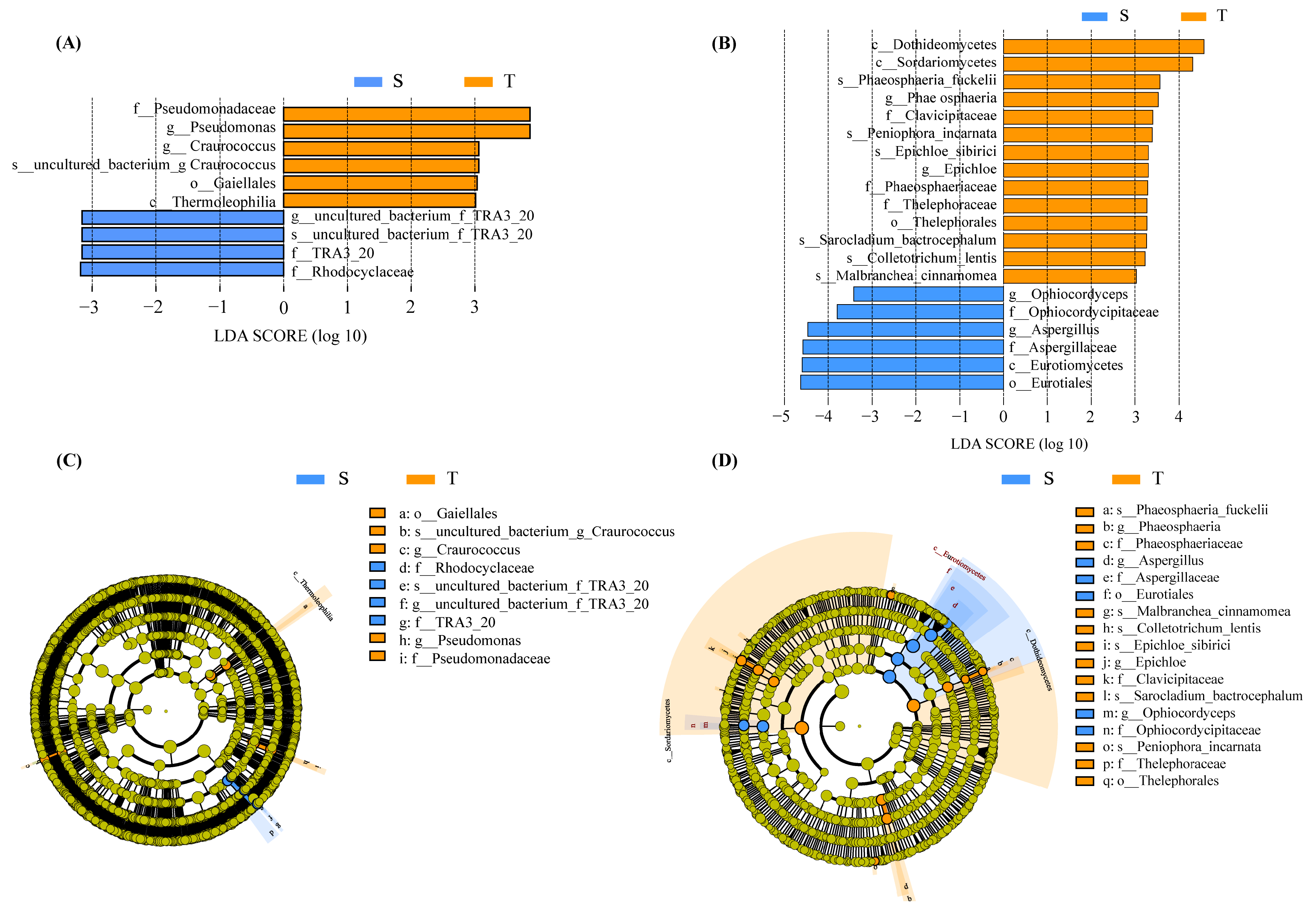
3.3. Microbial Functional Group Analysis
3.4. Metabolomics Analyses Between Two Different Trunk Types of P. yunnanensis
3.5. Co-Occurrence Network and Metabolites Correlated with Keystone Genus

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Liu, D.; Du, K.C.; Wang, A.D.; Meng, D.L.; Li, J.L. Secondary Metabolites from the Fresh Leaves of Pinus yunnanensis Franch. Chem. Biodivers. 2022, 19, e202100707. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zheng, W.; Han, M.; Su, J. Progress in Genetic Improvement of Pinus yunnanensis and Analysis of Its Breeding Strategies. West. J. For. Sci. 2010, 39, 104–110. [Google Scholar] [CrossRef]
- He, C.; Wu, Z.; Shen, D.; Gan, P.; Zhou, A.; Zong, D. Study on the Genetic Basis of Different Stem Forms of Pinus yunnanensis Based on SRAP Markers. J. Southwest For. Univ. Nat. Sci. 2021, 41, 1–10+197. Available online: https://kns.cnki.net/kcms2/article/abstract?v=Zw74qSZOFgjJZlaQOtHk1mM5ynHUuRjSlZLhnAoaU0A14-75uh1ETfH3Y1GjlesUOykrJ4B9snzbdAz9q5jShOuPTHmteQ1qLmCu81AUVQF7RpxKMhNRCpsUimI4XoflgDdfOC62DUkTlsiN0IefKyFS-V2uMSZ4BPhqYRuXtWjCYRzjghGL6ztGFOKqfEIm_g4FqZcvZ48=&uniplatform=NZKPT&language=CHS (accessed on 20 March 2025).
- Hong, Y.; Song, G.; Huang, R.; Jiang, H.-Q. A Preliminary Study on Genetic Variation and Relationships of Pinus yunnanensis and Its Closely Related Species. J. Integr. Plant Biol. 2000, 42, 107–110. [Google Scholar]
- Luo, L.; Xu, L. A study on the effect of oblique grain on the physical and mechanical properties of Pinus yunnanensis wood. For. Sci. 1983, 382–389+448. Available online: https://kns.cnki.net/kcms2/article/abstract?v=Zw74qSZOFgi0459KVPkuXKUSJ-57m0XU86g8C_uy6bzf-jfAxFvhWkq7RkSSqNLU-38DImPTiOZlQHAzfX9MLLY1kZsUDPmcD4F124Tr6K3y-nrZX6URGlMyvTfJE_fxgArPvpy28ZnbaBH6G7jksqTm7eKrVjPZzsufAGHvG3pW_Q8Uokve9mQMRLU_8ZYy&uniplatform=NZKPT&language=CHS (accessed on 20 March 2025).
- Luo, L.; Han, M.; Zheng, W.; Su, J.; Li, W.; Zheng, S.; Gong, J. The Causes and Classification of Low-Quality and Low-Efficiency Forests of Pinus yunnanensis. West. For. Sci. 2009, 38, 94–99. [Google Scholar] [CrossRef]
- Zhou, A.; Zong, D.; Luo, J.; Shen, D.; He, R.; Tian, B.; Xu, Y.; He, C. AFLP Analysis on Genetic Variation of Stem Forms in Pinus yunnanensis. Mol. Plant Breed. 2016, 14, 186–194. [Google Scholar] [CrossRef]
- Wang, C.; Wang, J.; Jiang, H.-Q. Morphological Characteristics of Stem of Pinus yunnanensis and Its Related Species in Different Habitats. J. West China For. Sci. 2009, 38, 23–27+125. [Google Scholar] [CrossRef]
- Kumar, A.; Verma, J.P. Does plant—Microbe interaction confer stress tolerance in plants: A review? Microbiol. Res. 2018, 207, 41–52. [Google Scholar] [CrossRef]
- Sharma, M.; Sudheer, S.; Usmani, Z.; Rani, R.; Gupta, P. Deciphering the Omics of Plant-Microbe Interaction: Perspectives and New Insights. Curr. Genom. 2020, 21, 343–362. [Google Scholar] [CrossRef]
- Rospond, B.; Chłopicka, J. Funkcje biologiczne L-karnityny i jej zawartość w wybranych produktach spozywczych [The biological function of L-carnitine and its content in the particular food examples]. Przegl. Lek. 2013, 70, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Zhu, J.; Zhao, X.; Shi, J.; Jiang, C.; Shao, D. Beneficial effects of endophytic fungi colonization on plants. Appl. Microbiol. Biotechnol. 2019, 103, 3327–3340. [Google Scholar] [CrossRef]
- De-La-Pena, C.; Loyola-Vargas, V.M. Biotic interactions in the rhizosphere: A diverse cooperative enterprise for plant productivity. Plant Physiol. 2014, 166, 701–719. [Google Scholar] [CrossRef]
- Rosier, A.; Bishnoi, B.; Lakshmanan, V.; Sherrier, D.J.; Bais, H.P. A perspective on inter-kingdom signaling in plant-beneficial microbe interactions. Plant Mol. Biol. 2016, 90, 537–548. [Google Scholar] [CrossRef]
- Le Cocq, K.; Gurr, S.J.; Hirsch, P.R.; Mauchline, T.H. Exploitation of endophytes for sustainable agricultural intensification. Mol. Plant Pathol. 2017, 18, 469–473. [Google Scholar] [CrossRef]
- Nataraja, K.N.; Suryanarayanan, T.S.; Shaanker, R.U.; Senthil-Kumar, M.; Oelmüller, R. Plant–microbe interaction: Prospects for crop improvement and management. Plant Physiol. Rep. 2019, 24, 461–462. [Google Scholar] [CrossRef]
- Feussner, I.; Polle, A. What the transcriptome does not tell—Proteomics and metabolomics are closer to the plants’ patho-phenotype. Curr. Opin. Plant Biol. 2015, 26, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Sun, D.; Chen, L.; An, Y. Integrative analysis of the microbiome and metabolome in understanding the causes of sugarcane bitterness. Sci. Rep. 2021, 11, 6024. [Google Scholar] [CrossRef]
- Li, Y.-H.; Yang, Y.-Y.; Wang, Z.-G.; Chen, Z. Emerging Function of Ecotype-Specific Splicing in the Recruitment of Commensal Microbiome. Int. J. Mol. Sci. 2022, 23, 4860. [Google Scholar] [CrossRef]
- Kang, S.H.; Cho, H.S.; Cheong, H.; Choongmin, R.; Kim, J.H.; Park, S.H. Two bacterial entophytes eliciting both plant growth promotion and plant defense on pepper (Capsicum annuum L.). J. Microbiol. Biotechnol. 2007, 17, 96. [Google Scholar] [CrossRef]
- Scherling, C.; Kristina, U.; Ewald, D.; Wolfram, W. A metabolic signature of the beneficial interaction of the endophyte Paenibacillus sp. isolate and in vitro-grown poplar plants revealed by metabolomics. Mol. Plant-Microbe Interact. 2009, 22, 1032–1037. [Google Scholar] [CrossRef] [PubMed]
- Balmer, D.; De Papajewski, D.V.; Planchamp, C.; Glauser, G.; Mauch-Mani, B. Induced resistance in maize is based on organ-specific defence responses. Plant J. Cell Mol. Biol. 2013, 74, 213–225. [Google Scholar] [CrossRef]
- Ważny, R.; Rozpądek, P.; Domka, A.; Jędrzejczyk, R.J.; Nosek, M.; Hubalewska-Mazgaj, M.; Lichtscheidl, I.; Kidd, P.; Turnau, K. The effect of endophytic fungi on growth and nickel accumulation in Noccaea hyperaccumulators. Sci. Total Environ. 2021, 768, 144666. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics—The link between genotypes and phenotypes. Plant Mol. Biol. 2002, 48, 155–171. [Google Scholar]
- Chong, J.; Xia, J. Computational Approaches for Integrative Analysis of the Metabolome and Microbiome. Metabolites 2017, 7, 62. [Google Scholar] [CrossRef]
- Mishra, S.; Priyanka; Sharma, S. Metabolomic Insights Into Endophyte-Derived Bioactive Compounds. Front. Microbiol. 2022, 13, 835931. [Google Scholar] [CrossRef]
- Heuberger, A.L.; Robison, F.M.; Lyons, S.M.A.; Broeckling, C.D.; Prenni, J.E. Evaluating plant immunity using mass spectrometry-based metabolomics workflows. Front. Plant Sci. 2014, 5, 291. [Google Scholar] [CrossRef]
- Wolf, R.T.S. Phloem Transport: Cellular Pathways and Molecular Trafficking. Annu. Rev. Plant Biol. 2009, 60, 207–221. [Google Scholar] [CrossRef]
- Lucas, W.J.; Groover, A.; Lichtenberger, R.; Furuta, K.; Yadav, S.-R.; Helariutta, Y.; He, X.-Q.; Fukuda, H.; Kang, J.; Brady, M.S.; et al. The plant vascular system: Evolution, development and functions. J. Integr. Plant Biol. 2013, 55, 294–388. [Google Scholar] [CrossRef]
- Yoo, S.C.; Chen, C.; Rojas, M.; Daimon, Y.; Ham, B.K.; Araki, T.; Lucas, W. Phloem long-distance delivery of FLOWERING LOCUS T (FT) to the apex. Plant J. 2013, 75, 456–468. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, C.X.; Chen, R.; He, S.Y. Challenging battles of plants with phloem-feeding insects and prokaryotic pathogens. Proc. Natl. Acad. Sci. USA 2019, 116, 23390–23397. [Google Scholar] [CrossRef] [PubMed]
- Miyashima, S.; Roszak, P.; Sevilem, I.; Toyokura, K.; Helariutta, Y. Mobile PEAR transcription factors integrate positional cues to prime cambial growth. Nature 2019, 565, 490–494. [Google Scholar] [CrossRef] [PubMed]
- Smetana, O.; Makila, R.; Lyu, M.; Amiryousefi, A.; Rodriguez, F.S.; Wu, M.F.; Sole-Gil, A.; Gavarron, M.L.; Siligato, R.; Miyashima, S. High levels of auxin signalling define the stem-cell organizer of the vascular cambium. Nature 2019, 565, 485–489. [Google Scholar] [CrossRef]
- Mago, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Khaleel, S.S.; Huang, H.; Wu, C.H. Software for pre-processing Illumina next-generation sequencing short read sequences. Source Code Biol. Med. 2014, 9, 8. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Elmar, P.; Christian, Q.; Katrin, K.; Fuchs, B.M.; Wolfgang, L.; Jrg, P.; Oliver, G.F. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Kõljalg, U.; Larsson, K.H.; Abarenkovenrik, K.; Nilsson, R.H.; Alexander, I.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; et al. UNITE: A database providing web-based methods for the molecular identification of ectomycorrhizal fungi. New Nhytologist 2005, 166, 1063–4068. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2: An improved and customizable approach for metagenome inference. bioRxiv 2020. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.R.; Schilling, J.; Kennedy, P. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Ulvcrona, A.U.; Ulvcrona, T.; Nilsson, U.; Lundmark, T. Stand density and fertilization effects on aboveground allocation patterns and stem form of Pinus yunnanensis in young stands. Scand. J. For. Res. 2014, 29, 197–209. [Google Scholar] [CrossRef]
- Shavnin, S.A.; Golikov, D.Y.; Montile, A.A. Rotational Activity of Trunk Parts in Scots Pine Trees: Seasonal Dynamics and Patterns. Dokl. Biol. Sci. 2022, 503, 63–67. [Google Scholar] [CrossRef]
- Gapare, W.; Hathorn, A.; Kain, D.; Matheson, C.; Wu, H. Inheritance of spiral grain in the juvenile core of Pinus yunnanensis. Can. J. For. Res. 2006, 37, 116–127. [Google Scholar] [CrossRef]
- Hannrup, B.; Säll, H.; Jansson, G. Genetic parameters for spiral grain in Scots pine and Norway spruce. Silvae Genet. 2003, 52, 215–220. [Google Scholar] [CrossRef]
- Pang, Z.; Xu, J.; Yi, C. Linking Plant Secondary Metabolites and Plant Microbiomes: A Review. Front. Plant Sci. 2021, 11, 621276. [Google Scholar] [CrossRef]
- Hiruma, K.; Gerlach, N.; Sacristán, S.; Nakano, R.T.; Hacquard, S.; Kracher, B.; Neumann, U.; Ramírez, D.; Bucher, M.; O’Connell, R.J.; et al. Root Endophyte Colletotrichum tofieldiae Confers Plant Fitness Benefits that Are Phosphate Status Dependent. Cell 2016, 165, 464–474. [Google Scholar] [CrossRef]
- Cregger, M.A.; Veach, A.M.; Yang, Z.K.; Crouch, M.J.; Vilgalys, R.; Tuskan, G.A.; Schadt, C.W. The Populus holobiont: Dissecting the effects of plant niches and genotype on the microbiome. Microbiome 2018, 6, 31. [Google Scholar] [CrossRef]
- Li, P.; Zong, D.; Gan, P.; Li, H.; Wu, Z.; Li, F.; Zhao, C.; Li, L.; He, C. Comparison of the diversity and structure of the rhizosphere microbial community between the straight and twisted-trunk types of Pinus yunnanensis. Front. Microbiol. 2023, 14, 1066805. [Google Scholar] [CrossRef]
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Raaijmakers, J.M.; Irene, D.B.; Ole, N.; Marc, O. Natural functions of lipopeptides from Bacillus and Pseudomonas: More than surfactants and antibiotics. Fems Microbiol. Rev. 2010, 34, 1037–1062. [Google Scholar] [CrossRef] [PubMed]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: Looking back and future perspectives. Front. Microbiol. 2014, 5, 148. [Google Scholar] [CrossRef]
- Nadana, R.V.G. Isolation of Pseudomonas Species from Rhizospheric Soil and its Antagonistic Effect on Plant Pathogen. Int. J. Res. Appl. Sci. Eng. Technol. 2021, 9, 2405–2408. [Google Scholar] [CrossRef]
- Munakata, Y.; Heuson, E.; Daboudet, T.; Deracinois, B.; Duban, M.; Hehn, A.; Coutte, F.; Slezack-Deschaumes, S. Screening of Antimicrobial Activities and Lipopeptide Production of Endophytic Bacteria Isolated from Vetiver Roots. Microorganisms 2022, 10, 209. [Google Scholar] [CrossRef]
- Bao, C.; Sha, L.; Yingjie, W.; Jiayuan, Y.; Qiong, W.; Xiaomeng, X.; Fengshan, P.; Khan, K.Y.; Ying, F.; Xiaoe, Y. The Effects of the Endophytic Bacterium Pseudomonas fluorescens Sasm05 and IAA on the Plant Growth and Cadmium Uptake of Sedum alfredii Hance. Front. Microbiol. 2017, 8, 2538. [Google Scholar] [CrossRef]
- Ghaderi, F.; Habibi, A.; Sharifnabi, B. Phylogenetic Analysis of Phaeosphaeria Species Using Mating Type Genes and Distribution of Mating Types in Iran. Plant Pathol. J. 2022, 38, 78–79. [Google Scholar] [CrossRef]
- Lazarevi, J.; Menkis, A. Fungal Diversity in the Phyllosphere of Pinus heldreichii H. Christ—An Endemic and High-Altitude Pine of the Mediterranean Region. Diversity 2020, 12, 172. [Google Scholar] [CrossRef]
- Laihonen, M.; Saikkonen, K.; Helander, M.; de Aldana, B.R.V.; Zabalgogeazcoa, I.; Fuchs, B. Epichloë Endophyte-Promoted Seed Pathogen Increases Host Grass Resistance Against Insect Herbivory. Front. Microbiol. 2021, 12, 786619. [Google Scholar] [CrossRef]
- Zhong, R.; Bastías, D.A.; Zhang, X.; Li, C.; Nan, Z. Vertically Transmitted Epichloë Systemic Endophyte Enhances Drought Tolerance of Achnatherum inebrians Host Plants through Promoting Photosynthesis and Biomass Accumulation. Fungi 2022, 8, 512. [Google Scholar] [CrossRef]
- Newman, J.A.; Gillis, S.; Hager, H.A. Costs, Benefits, Parasites and Mutualists: The Use and Abuse of the Mutualism-Parasitism Continuum Concept for Epichloë Fungi. bioRxiv. 2021. [Google Scholar] [CrossRef]
- Wang, Z.; Li, M.; Ju, W.; Ye, W.; Xue, L.; Boufford, D.E.; Gao, X.; Yue, B.; Liu, Y.; Pierce, N.E. The entomophagous caterpillar fungus Ophiocordyceps sinensis is consumed by its lepidopteran host as a plant endophyte. Fungal Ecol. 2020, 47, 100989. [Google Scholar] [CrossRef]
- de Menezes, T.A.; Aburjaile, F.F.; Quintanilha-Peixoto, G.; Tomé, L.M.R.; Fonseca, P.L.C.; Mendes-Pereira, T.; Araújo, D.S.; Melo, T.S.; Kato, R.B.; Delabie, J.H.C.; et al. Unraveling the Secrets of a Double-Life Fungus by Genomics: Ophiocordyceps australis CCMB661 Displays Molecular Machinery for Both Parasitic and Endophytic Lifestyles. Fungi 2023, 9, 110. [Google Scholar] [CrossRef] [PubMed]
- Pyszynski, W. Mechanism of formation of spiral grain in Aesculus stems: Dissymmetry of deformation of stems caused by cyclic torsion. Acta Soc. Bot. Pol. 2015, 46, 501–522. [Google Scholar] [CrossRef]
- Hagag, A.; Abdelwahab, M.F.; El-Kader, A.M.A.; Fouad, M.A. The endophytic Aspergillus strains: A bountiful source of natural products. J. Appl. Microbiol. 2022, 132, 4150–4169. [Google Scholar] [CrossRef] [PubMed]
- Toghueo, R.M.K.; Boyom, F.F. Endophytic Penicillium species and their agricultural, biotechnological, and pharmaceutical applications. 3 Biotech 2020, 10, 107. [Google Scholar] [CrossRef]
- Zhao, W.; Bi, X.; Peng, Y.; Bai, M. Research advances of the phosphorus-accumulating organisms of Candidatus Accumulibacter, Dechloromonas and Tetrasphaera: Metabolic mechanisms, applications and influencing factors. Chemosphere 2022, 307, 135675. [Google Scholar] [CrossRef]
- Hedden, P.; Sponsel, V. A Century of Gibberellin Research. J. Plant Growth Regul. 2015, 34, 740–760. [Google Scholar] [CrossRef]
- Hartmann, M.; Zeier, T.; Bernsdorff, F.; Reichel-Deland, V.; Kim, D.; Hohmann, M.; Scholten, N.; Schuck, S.; Bräutigam, A.; Hölzel, T.; et al. Flavin Monooxygenase-Generated N-Hydroxypipecolic Acid Is a Critical Element of Plant Systemic Immunity. Cell 2018, 173, 456. [Google Scholar] [CrossRef]
- López-Sámano, M.; Beltrán, L.F.L.-A.; Sánchez-Thomas, R.; Dávalos, A.; Villaseñor, T.; García-García, J.D.; García-de los Santos, A. A novel way to synthesize pantothenate in bacteria involves β-lanine synthase present in uracil degradation pathway. MicrobiologyOpen 2020, 9, e1006. [Google Scholar] [CrossRef]
- Wright, L.D.; Miller, C.S. Uracil in growth and pyrimidine nucleotide synthesis of Lactobacillus bulgaricus 09. Proc. Soc. Exp. Biol. Med. 1952, 81, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Wang, J.; E, J.; Cheng, Z.; Zhang, J. The effect of uracil on the freeze-dried survival rate of Lactobacillus plantarum LIP-1 and its mechanism of action. Trans. Chin. Soc. Agric. Eng. 2022, 38, 308–316. [Google Scholar]
- Lobley, C.M.C.; Schmitzberger, F.; Kilkenny, M.L.; Whitney, H.; Ottenhof, H.H.; Chakauya, E.; Webb, M.E.; Birch, L.M.; Tuck, K.L.; Abell, C. Structural insights into the evolution of the pantothenate-biosynthesis pathway. Biochem Soc Trans 2003, 31, 563–571. [Google Scholar] [CrossRef]
- Leonardi, R.; Jackowski, S. Biosynthesis of Pantothenic Acid and Coenzyme A. Ecosal Plus 2007, 2. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Lou, K.; Li, C. Promotion of plant growth by phytohormone-producing endophytic microbes of sugar beet. Biol. Fertil. Soils 2009, 45, 645–653. [Google Scholar] [CrossRef]
- Bian, X.; Qu, C.; Zhang, M.; Li, D.; Jiang, J.; Liu, G. Transcriptome analysis provides new insights into leaf shape variation in birch. Trees: Struct. Funct. 2019, 33, 1265–1281. [Google Scholar] [CrossRef]
- Eklund, L.; Säll, H.; Linder, S. Enhanced growth and ethylene increases spiral grain formation in Picea abies and Abies balsamea trees. Trees 2003, 17, 81–86. [Google Scholar] [CrossRef]
- Suharti, W.S.; Nose, A.; Zheng, S.H. Metabolomic study of two rice lines infected by Rhizoctonia solani in negative ion mode by CE/TOF-MS. J. Plant Physiol. 2016, 1, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Hinsch, J.; Galuszka, P.; Tudzynski, P. Functional characterization of the first filamentous fungal tRNA-isopentenyltransferase and its role in the virulence of Claviceps purpurea. New Phytol. 2016, 211, 980–992. [Google Scholar] [CrossRef]
- Mishra, D.C.; Arora, D.; Budhlakoti, N.; Solanke, A.U.; Mithra, S.V.A.C.; Kumar, A.; Pandey, P.S.; Srivastava, S.; Kumar, S.; Farooqi, M.S. Identification of Potential Cytokinin Responsive Key Genes in Rice Treated With trans-Zeatin Through Systems Biology Approach. Front. Genet. 2022, 12, 780599. [Google Scholar] [CrossRef]
- Gajdosová, S.; Spíchal, L.; Kamínek, M.; Hoyerová, K.; Novák, O.; Dobrev, P.I.; Galuszka, P.; Klíma, P.; Gaudinová, A.; Žižková, E.; et al. Distribution, biological activities, metabolism, and the conceivable function of cis-Zeatin-type cytokinins in plants. J. Exp. Bot. 2011, 62, 2827–2840. [Google Scholar] [CrossRef] [PubMed]
- Perostová, S.; Jaroová, J.; Dobrev, P.; Hlusková, L.; Motyka, V.; Filepová, R.; Knirsch, V.; Gaudinova, A.; Kieber, J.; Vaňková, R. Heat Stress Targeting Individual Organs Reveals the Central Role of Roots and Crowns in Rice Stress Responses. Front. Plant Sci. 2022, 12, 799249. [Google Scholar] [CrossRef]
- Mengiste, T. Plant immunity to necrotrophs. Annu. Rev. Phytopathol. 2012, 50, 267–294. [Google Scholar] [CrossRef]
- Yuan, H.M.; Liu, W.C.; Lu, Y.T. CATALASE2 Coordinates SA-Mediated Repression of Both Auxin Accumulation and JA Biosynthesis in Plant Defenses. Cell Host Microbe 2017, 21, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, R.F.; Yuan, H.M.; Li, T.T.; Wang, L.F.; Lu, K.K.; Guo, J.X.; Liu, W.C. Overexpressing the N-terminus of CATALASE2 enhances plant jasmonic acid biosynthesis and resistance to necrotrophic pathogen Botrytis cinerea B05.10. Mol Plant Pathol. 2021, 22, 1226–1238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yang, C.; Liu, J. Phytopathogenic bacterial effectors. Science Bulletin. 2023, 68, 4895–4917. Available online: https://kns.cnki.net/kcms2/article/abstract?v=jNHD1hIvxn0DsP0kHfLF5LuMZb05LXk0DkkbkQpXL2kkW4cxyYdBISTX-e7db4264z35RJa7UJilRHmIwHjS3hJzOsTB0DuPTRcbR3ge-m6-lyFCojsBB5IOTemcGQGLU1atdskxezuY0HHYNWrQGPWjuJtLqrGBOV1kBaX5_yDoDBr-C4NlGngmEbXjaKXpqMYS79gfnog=&uniplatform=NZKPT&language=CHS (accessed on 20 March 2025).
- Darino, M.; Chia, K.S.; Marques, J.; Aleksza, D.; Soto-Jiménez, L.M.; Saado, I.; Uhse, S.; Borg, M.; Betz, R.; Bindics, J.; et al. Ustilago maydis effector Jsi1 interacts with Topless corepressor, hijacking plant jasmonate/ethylene signaling. New Phytol 2021, 229, 3393–3407. [Google Scholar] [CrossRef]
- Nilsson, A.K.; Fahlberg, P.; Johansson, O.N.; Hamberg, M.; Andersson, M.X.; Ellerström, M. The activity of HYDROPEROXIDE LYASE 1 regulates accumultation of galactolipids containing 12-oxo-phytodienoic acid in Arabidopsis. J. Exp. Bot. 2016, 64, 5133–5144. [Google Scholar] [CrossRef]
- Kumar, A.; Yogendra, K.N.; Karre, S.; Kushalappa, A.C.; Dion, Y.; Choo, T.M. WAX INDUCER1 (HvWIN1) transcription factor regulates free fatty acid biosynthetic genes to reinforce cuticle to resist Fusarium head blight in barley spikelets. J. Exp. Bot. 2016, 67, 4127–4139. [Google Scholar] [CrossRef]
- Zhu, K.; Gu, S.; Liu, J.; Luo, T.; Khan, Z.; Zhang, K.; Hu, L. Wood Vinegar as a Complex Growth Regulator Promotes the Growth, Yield, and Quality of Rapeseed. Agronomy 2021, 11, 510. [Google Scholar] [CrossRef]
- Yu, H.; Lin, Z.; Li, Y.; Yuan, L.; Zhao, B. Effects of spraying low molecular organic compounds on growth and nutrients uptake of rape (Brassica Chinensis L.). J. Plant Nutr. Fertil. 2014, 20, 1560–1568. [Google Scholar] [CrossRef]
- Gourkhede, P.H.; Patil, V.D.; Narale, S. Effect of Foliar Feeding of Gluconate and EDTA Chelated Plant Nutrients on Yield, Quality and Nutrient Concentration of Bt-Cotton. Int. J. Trop. Agric. 2015, 33, 1875–1879. [Google Scholar]
- Marra, R.; Coppola, M.; Pironti, A.; Grasso, F.; Lombardi, N.; D’Errico, G.; Sicari, A.; Censi, S.B.; Woo, S.L.; Rao, R. The Application of Trichoderma Strains or Metabolites Alters the Olive Leaf Metabolome and the Expression of Defense-Related Genes. J. Fungi 2020, 6, 369. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Wang, X.; Yan, X.; Ke, J.; Yang, Y.; Li, Y.; Xu, D.; Zhuo, Z.; Yan, X. Research progress on the application of transcriptomics and metabolomics in tree fungal disease defense responses. World For. Res. 2022, 35, 27–32. [Google Scholar] [CrossRef]
- Mosquera, S.; Leveau, J.H.J.; Stergiopoulos, I. Repeated exposure of Aspergillus niger spores to the antifungal bacterium Collimonas fungivorans Ter331 selects for delayed spore germination. Appl. Environ. Microbiol. 2021, 87, e00233-21. [Google Scholar] [CrossRef]
- Li, J.; Pan, M.; Zhang, X.; Zhou, Y.; Feng, G.-D.; Zhu, H. Collimonas silvisoli sp. nov. and Collimonas humicola sp. nov., two novel species isolated from forest soil. Microbiol. Soc. 2021, 71, 005061. [Google Scholar] [CrossRef]
- Uroz, S.; Geisler, O.; Fauchery, L.; Lami, R.; Rodrigues, A.M.S.; Morin, E.; Leveau, J.H.J.; Oger, P. Genomic and transcriptomic characterization of the Collimonas quorum sensing genes and regulon. FEMS Microbiol. Ecol. 2022, 98, 1–22. [Google Scholar] [CrossRef]
- Akinsanya, M.A.; Goh, J.K.; Lim, S.P.; Adeline Su Yien Ting, A.S.Y. Diversity, antimicrobial and antioxidant activities of culturable bacterial endophyte communities in Aloe vera. FEMS Microbiol. Lett. 2015, 362, fnv184. [Google Scholar] [CrossRef]
- Jeong, J.J.; Sang, M.K.; Wan, L.D.; Choi, I.G.; Kim, K.D. Chryseobacterium phosphatilyticum sp. nov., a phosphate-solubilizing endophyte isolated from cucumber (Cucumis sativus L.) root. Int. J. Syst. Evol. Microbiol. 2019, 69, 610–615. [Google Scholar] [CrossRef]
- Singburaudom, N. The alkaloid Berberine isolated from Coscinium fenestratum is an inhibitor of phytopathogenic fungi. J. Biopestic. 2015, 8, 28–36. [Google Scholar] [CrossRef]
- Glenz, R.; Kaiping, A.; Göpfert, D.; Weber, H.; Lambour, B.; Sylvester, M.; Fröschel, C.; Mueller, M.J.; Osman, M.; Waller, F. The major plant sphingolipid long chain base phytosphingosine inhibits growth of bacterial and fungal plant pathogens. Sci. Rep. 2022, 12, 1081. [Google Scholar] [CrossRef] [PubMed]
- Kleber, H.P. Bacterial carnitine metabolism. FEMS Microbiol. Lett. 1997, 147, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Millican, M. The Role of Plant-Derived Quaternary Ammonium Compounds, Including Carnitine and Choline-O-sulfate, on the Biology of the Plant Pathogen Pseudomonas syringae and Its Interactions with the Host Species Phaseolus vulgaris. Doctoral Thesis, Iowa State University, Ames, IA, USA, 2015. [Google Scholar] [CrossRef]
- Rosenfeld, H.; Feigelson, P. Product induction in Pseudomonas acidovorans of a permease system which transports L-tryptophan. J. Bacteriol. 1969, 97, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Kelly, Y.L.; Yinghao, W.; Tszking, L.; Chuenfai, K.; Wingping, Y.; Zhongzhen, Z. Correlation between quality and geographical origins of Leonuri Herba revealed by the qualitative fngerprint profling and quantitative determination of chemical components. Chin. Med. 2022, 17, 2–14. [Google Scholar] [CrossRef]
- Jung, H.; Lee, D.; Lee, S.; Kong, H.J.; Park, J.; Seo, Y.S. Comparative genomic analysis of Chryseobacterium species: Deep insights into plant-growth-promoting and halotolerant capacities. Microb. Genom. 2023, 9, 001108. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, J.; Zhang, X.; Li, H.; Yang, W.; Wang, C.; Qiu, Y.; Luo, Y.; Gan, P.; He, C.; Li, P.; et al. Comparison of the Microbiome and Metabolome in the Phloem of Straight and Twisted Trunks of Pinus yunnanensis Franch. Forests 2025, 16, 574. https://doi.org/10.3390/f16040574
Feng J, Zhang X, Li H, Yang W, Wang C, Qiu Y, Luo Y, Gan P, He C, Li P, et al. Comparison of the Microbiome and Metabolome in the Phloem of Straight and Twisted Trunks of Pinus yunnanensis Franch. Forests. 2025; 16(4):574. https://doi.org/10.3390/f16040574
Chicago/Turabian StyleFeng, Jiayu, Xiaolin Zhang, Hailin Li, Wenwu Yang, Cai Wang, Yingqing Qiu, Yiran Luo, Peihua Gan, Chengzhong He, Peiling Li, and et al. 2025. "Comparison of the Microbiome and Metabolome in the Phloem of Straight and Twisted Trunks of Pinus yunnanensis Franch" Forests 16, no. 4: 574. https://doi.org/10.3390/f16040574
APA StyleFeng, J., Zhang, X., Li, H., Yang, W., Wang, C., Qiu, Y., Luo, Y., Gan, P., He, C., Li, P., & Zong, D. (2025). Comparison of the Microbiome and Metabolome in the Phloem of Straight and Twisted Trunks of Pinus yunnanensis Franch. Forests, 16(4), 574. https://doi.org/10.3390/f16040574