Abstract
To explore environmental differentiation in the stoichiometric characteristics of carbon (C), nitrogen (N), phosphorus (P), potassium (K), and their ratios among tree species organs in different lithological karst regions of Guizhou, six common species were selected from the limestone karst area of Liping and the dolomite karst area of Shibing. The analysis focused on the differences in the contents of C, N, P, and K, as well as the ratios of C/N, C/P, C/K, and N/P, and the correlations among these elements in branches, leaves, and roots, with the aim of investigating the allocation distribution patterns of nutrient elements among organs in tree species under different lithological karst conditions. The results showed the following: In both dolomite and limestone karst regions, the tree species exhibited a distinct characteristic: their leaves exhibited high C levels, but relatively low N and P levels. Tree species in both karst regions showed high C/N, C/P, and C/K ratios, indicative of rapid C assimilation rates and efficient utilization of N, P, and K. Plant growth in the dolomite region was N-limited, while plant growth in the limestone region was P-limited. However, plant growth in neither region was K-limited. The nutrient characteristics and adaptation strategies of plants in dolomite and limestone karst regions were the result of multiple factors, such as lithology, plant organs, and interactions among elements. In conclusion, considering the differences in lithology during vegetation restoration and management in karst regions, targeted application of nitrogen or phosphorus fertilizers is expected to promote plant growth and further enhance the carbon sequestration capacity of both karst plants and soils.
1. Introduction
Ecological stoichiometry is a scientific discipline that integrates multiple disciplines, such as botany, chemistry, and ecology, to study the energy balance of ecosystems and the balance of multiple chemical elements within biological organisms [1,2,3]. This research field organically unifies the theoretical research at different hierarchical levels of biology, including at the molecular, cellular, individual, population, ecosystem, and global scales [4]. It is widely applied in research areas such as individual plant growth, population dynamics, community succession, identification of limiting elements, and ecosystem stability [5,6,7], providing new insights into exploring nutrient recycling mechanisms in different ecosystems [8].
Leaves, branches, and roots are essential components of embryophyta, performing distinct functions during plant growth. The root system functions as the principal organ for nutrient absorption in plants [9,10], while leaves accumulate photosynthetic products through photosynthesis [11]. Branches provide storage space for nutrients and connect leaves to roots [12]. Carbon, nitrogen, phosphorus, and potassium are the fundamental nutritive components of plants [11], and their nutrient contents and ecological stoichiometric properties can determine the stability of plant organs and the allocation ratios and inter-relationships among these elements in different organs [13]. This understanding aids in elucidating plant growth and maturation sequences, community composition and organization, diversity, and functionality, and reflects plants’ adaptability to environmental changes [14,15]. Differences in organ functions, as well as variations in the physical and biological environments of plants, directly lead to disparities in nutrient allocation among their organs [16]. Leaves require more nitrogen and phosphorus for rapid growth and high photosynthetic rates, while root respiration and nutrient uptake demand more energy. Pooter’s research [17] showed that, under low-nitrogen conditions, plants may allocate more nitrogen to their roots to enhance nutrient absorption capacity. Plants adjust their resource allocation strategies based on environmental conditions such as light and nutrient supply. Schindler [18] pointed out that under nitrogen-limited conditions, plants may exhibit higher C/N. Drought stress may lead to a shift in carbon allocation toward roots to enhance water absorption capacity, thereby altering the root C/N ratio [19,20]. Reich and Oleksyn’s research [21] demonstrated that there are significant differences in C/N and C/P between herbaceous and woody plants. Therefore, conducting in-depth research on the distribution of nutrients in leaves, branches, and roots is essential for understanding plants’ environmental adaptation strategies and their mechanisms of nutrient allocation [16].
The region of southwestern China is a mountainous plateau area, with the largest distribution and most intense development of tropical and subtropical karst landforms in China and the world. It is also the most completely developed continuous karst zone globally [22]. Formed by the dissolution and transformation of two types of carbonate rocks, dolomite and limestone, this region has a unique landform, with special structures and functions characterized by high ecological sensitivity, low environmental carrying capacity, and poor stability [23]. The variability in karst lithologies results in heterogeneous habitats for vegetation, leading to differences in the composition [24] and internal environments [25] of forest ecosystems, which, in turn, affect nutrient distribution patterns. Plants exhibit variations in stoichiometric characteristics among their organs across regions, due to the diverse demands of physiological processes, nutrient storage capacities, and functional specializations of each organ [7,26]. Therefore, exploring the ecological stoichiometric traits of C, N, P, and K in different organs of common plants across distinct karst lithological areas can enhance our understanding of plant growth regulation mechanisms and adaptive strategies, which is crucial for reasonably evaluating plant growth potential. It is also crucial for gaining insights into nutrient cycling, plant growth succession, and vegetation recovery and re-establishment, as well as the management of rocky desertification ecosystems [23,27]. This study focused on six tree species common to the limestone karst area of Liping and the dolomite karst area of Shibing. It analyzed the differences in C, N, P, and K contents and ecological stoichiometric characteristics among organs of these tree species in distinct karst lithological regions, and explored the differences in nutrient limitation and resource competitiveness, as well as the differentiated responses and adaptation strategies of these tree species to the environment.
2. Materials and Methods
2.1. Research Sites
The study site is situated in two county-level regions, Shibing and Liping, which are 400 km apart. Their geographic coordinates are 26°57′42.4558″ N, 108°14′45.63919″ E and 109°7′18.15″ N, 25°59′15.08″ E, respectively (Figure 1). The site experiences a mid-subtropical humid monsoon climate. The typical annual temperature is 16.3 °C, and precipitation amounts to approximately 1182.4 mm per year, primarily concentrated from April to October. The annual average relative humidity is 80.0%. The period without frost lasts 294 days, while the total annual sunshine time amounts to 1195 h. The average elevation of the study area ranges from 500 to 850 m. Based on lithological characteristics, Shibing and Liping are classified under karst landscapes predominantly characterized by dolomite and limestone, respectively, which have undergone weathering to form limestone soils. There is an absence of continuous sedimentary layers from deposits of the Quaternary age or other ages over carbonate rocks. The soil layer is thin (approximately 20 cm) and discontinuous, predominantly found within rock fissures or gullies formed by exposed rocks. It is rich in calcium and basic cations, as well as having a high organic matter content [28]. The vegetation types in the two study areas are subtropical evergreen deciduous broad-leaved forests and limestone evergreen broad-leaved forests. The dominant tree species in the canopy layer include Platycarya strobilacea Sieb. et Zucc., Quercus fabri Hance, Quercus acutissima Carr., Cyclobalanopsis phanera (Chun) Y. C. Hsu et H. W. Jen, etc. The shrub layer is primarily composed of Ligustrum quihoui Carr., Loropetalum chinense (R. Br.) Oliver, Lindera glauca (Sieb. et Zucc.) Bl., and Manglietia fordiana (Hemsl.) OliV [29].
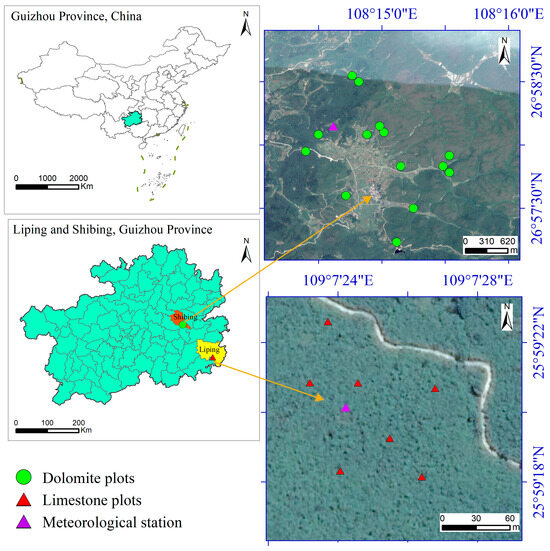
Figure 1.
The locations of the study sites in Liping and Shibing.
2.2. Research Approach
2.2.1. Field Sampling Survey
From July to August during 2017 to 2019, we established 14 and 7 sample plots with dimensions of 20 m × 20 m in Shibing and Liping, respectively, for field investigations. The slope, aspect, longitude, latitude, and altitude of individual fields were measured by means of a hand-held GPS and a compass, and canopy density, shrub herb coverage, rock exposure, and slope position were estimated using a visual method.
2.2.2. Sample Collection, Processing, and Measurement
The species commonly found in both dolomite and limestone karst areas were selected for this study. In the dolomite stand, the average diameter at breast height (DBH) of trees was 25.5 cm, 9.9 cm, 7.3 cm, 16.0 cm, 7.5cm and 5.9 cm for Pinus massoniana, Quercus acutissima, Carpinus turczaninowii, Liquidambar formosana, Cyclobalanopsis glauca and Lindera glauca, respectively, while the average height of trees was 21 m, 13 m, 11 m, 16 m, 12 m and 11 m for them, respectively. The average DBH of trees in the limestone stand was 15.7 cm, 9.5 cm, 7.7 cm, 8.3 cm, 8.7 cm and 7.2 cm for Pinus massoniana, Quercus acutissima, Carpinus turczaninowii, Liquidambar formosana, Cyclobalanopsis glauca and Lindera glauca, respectively, while the average height of trees was 11 m, 13 m, 12 m, 12 m, 12 m and 11 m for them, respectively. Within typical sample plots, three healthy trees of each species, close to the average condition of that species, were chosen for sample collection. One sample was taken from each organ of each tree. Leaf and branch samples were collected using the standard branch pruning method. Standard branches were selected and cut off from four different directions of the tree canopy at the base using a pole pruner. Intact, fully expanded, and disease-free mature leaves without petioles were collected and thoroughly mixed, and a leaf sample was composed using the quartering method (with a sample size of ≥30 leaves). Tree branch disk samples were collected, taking into full consideration the distance along the base of the branch and the weight of different branch orders. A mixed branch sample was then retained using the quartering method (one-quarter of the disk). Fine roots (≤2 mm in diameter) from the soil surface, at a depth of approximately 20 cm, around the base of the sample trees were excavated using a small shovel, and any soil and impurities adhering to the surface of the fine roots were removed. The gathered branches, leaves, and roots were sealed and kept in a portable refrigerator for future analysis.
The gathered plant samples, including leaves, branches, and roots, were rinsed with ultrapure water, blotted with filter paper to absorb excess moisture, and heated at 105 °C in an oven for 15 min, followed by drying at a constant temperature of 65 °C for 48 h until they achieved a stable weight. After being crushed and passed through a 0.25 mm sieve, the samples were used to quantify the total carbon (C), total nitrogen (N), total phosphorus (P), and total potassium (K) contents of the leaves, branches, and roots. The potassium dichromate-external heating method, using a heating digestion device (SPH120; Alva Instrument Co., LTD, Jinan, China) and a burette (Brand titrette 50 mL, BRAND GMBH+CO KG, Wertheim, Germany), was applied to quantify the C content, with an uncertainty of 1%–3%. With the aid of a Kjeldahl apparatus (KN520; Jinan Alva Instrument Co., LTD, Jinan, China), the semi-micro Kjeldahl method was utilized to determine the nitrogen (N) content, achieving an accuracy within 1%–2%. The P content was quantified via the molybdenum–antimony-anti-colorimetry method [30], utilizing a spectrophotometer (TU-1810; Beijing Puyang General Instrument Co., LTD, Beijing, China) to determine the absorbance, with an uncertainty of 2%–5%. The K content was assayed using flame photometry (FP6450; Shanghai Xin Yi Precision Instrument Co., LTD, Shanghai, China), quantified with a flame photometer, with an uncertainty of 1%–3%.
Soil sampling: Each plot was evenly divided into four subplots, and one surface soil sample (0–20 cm) was collected from each subplot; these samples were then mixed to form a composite sample. The composite samples were purified, dried, ground, sieved, and processed further in preparation for element determination. For information on the measurement methods of soil samples, please refer to reference [29]. The soil nutrient content in the two regions is shown in Table 1.

Table 1.
The nutrient content (mean ± standard deviation) of topsoil in karst areas with different lithologies.
2.3. Data Analysis and Statistics
Data processing and statistical analysis were performed using Excel 2016 and SPSS 21.0, while plotting was conducted with Origin 2021. The effects of the lithologic karst areas and organs on the nutrient content and stoichiometric ratio were analyzed using two-way ANOVA. Using one-way ANOVA and LSD, the variations in nutrient content and stoichiometric ratios were analyzed across different lithologic karst areas and plant organs, respectively; the significance level was 0.05. The correlation of each index was analyzed using Pearson correlation analysis.
3. Results
3.1. C, N, P, and K Contents in Various Plant Organs in Karst Regions with Different Lithologies
Across different organs, the C content in both dolomite and limestone regions showed a trend of leaves > branches > roots, with coefficient of variation (CV) ranges of 10%–16% and 8%–13%, respectively. In the dolomite region, the leaves had a markedly higher C concentration (499.62 g/kg), while the roots had a correspondingly lower one (p < 0.05), whereas in the limestone region, the C content in leaves (499.68 g/kg) was notably elevated compared with both branches and roots (Figure 2a). Regarding the N content, the dolomite region showed a trend of branches (9.07 g/kg) > leaves (6.78 g/kg) > roots (5.94 g/kg), with CV ranges of 49%–75%. A notable variation was found in the N content between branches and roots (p < 0.05) in this region. In the limestone region, the trend was leaves (15.84 g/kg) > branches (7.62 g/kg) > roots (5.94 g/kg), with the N concentration in leaves being distinctly different from that in branches and roots (p < 0.05), and with CV ranges of 32%–65% (Figure 2c). For the P content, the dolomite region demonstrated a trend of leaves (1.16 g/kg) > branches (1.15 g/kg) > roots (0.86 g/kg), with CV ranges of 20%–57%. In contrast, the limestone region exhibited a trend of leaves (0.50 g/kg) > branches (0.27 g/kg) > roots (0.20 g/kg), with the P content in leaves being significantly higher compared to that in other organs, and with CV ranges of 32%–48% (Figure 2b). For the K content, the dolomite region showed a trend of leaves (5.08 g/kg) > roots (3.96 g/kg) > branches (2.97 g/kg), with marked variation in the K content between leaves and branches (p < 0.05). However, in the limestone region, the trend was branches (8.28 g/kg) > leaves (6.13 g/kg) > roots (6.08 g/kg), with CV ranges of 50%–97% and 30%–73%, respectively. (Figure 2d). Variance analysis indicated that the dolomite regions had significantly higher C in branches and roots, as well as a significantly higher P content in all organs, compared with the limestone regions (p < 0.05). Conversely, limestone regions had significantly greater N in leaves and K in branches (p < 0.05) (Figure 2).
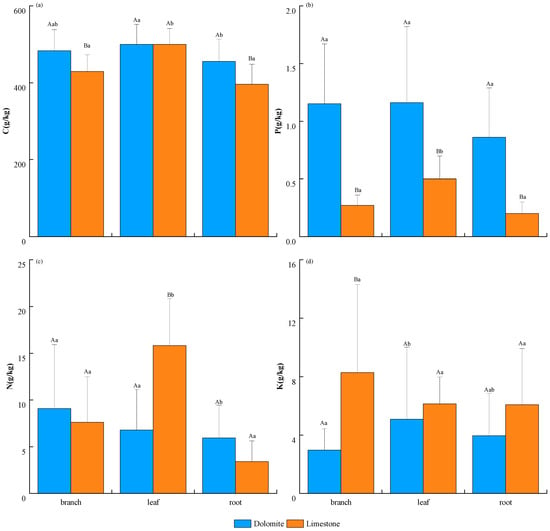
Figure 2.
Variations in nutrient elements across plant organs in karst areas with distinct lithologic compositions. (a) the content of C, (b) the content of P, (c) the content of N, and (d) the content of K. Uppercase letters indicate significant disparities in the same organ across regions (p < 0.05), while lowercase letters signify significant contrasts among organs within the same region (p < 0.05).
3.2. Ecological Stoichiometric Ratios of Plant Organs Across Karst Regions with Varying Lithologies
In various plant organs, the C/N ratio demonstrated a trend of leaves (115.15) > branches (107.53) > roots (107.39) in the dolomite region. Conversely, in the limestone region, the roots (187.70) had a notably higher C/N ratio than the branches (87.52) and leaves (35.76) (Figure 3a). Regarding the C/P ratio, the trend was roots > leaves > branches, with no significant difference in the dolomite region (ratios of 619.47, 532.72, and 501.3, respectively). However, in the limestone region, the C/P ratio was found to be significantly elevated in roots (2373.93) compared to that in leaves (1164.68) (Figure 3b). The C/K ratio showed a trend of branches > roots > leaves in the dolomite region, whereas in the limestone region, the trend was leaves > roots > branches, with ranges of 196.71–207.8 and 69.02–87.56, respectively (Figure 3c). For the N/P ratio, the trend in the dolomite region was roots > leaves > branches, while in the limestone region, it was leaves > branches > roots, with ranges of 6.09–7.39 and 16.93–37.52, respectively (Figure 3d). The N/K exhibited a trend of branches > roots > leaves in the dolomite area and leaves > branches > roots in the limestone area, with variation ranges of 1.78–3.88 and 0.58–2.64, respectively (Figure 3e). Finally, the K/P showed a trend of roots > leaves > branches in the dolomite area and roots > branches > leaves in the limestone area, with ranges of 3.04–5.51 and 14.23–34.58, respectively (Figure 3f). Variance analysis revealed that the dolomite regions had significantly higher C/N in leaves and C/K in all organs compared with the limestone regions. Conversely, the limestone regions had significantly higher C/P in branches and roots, along with significantly higher N/P and K/P in all organs (p < 0.05) (Figure 3).
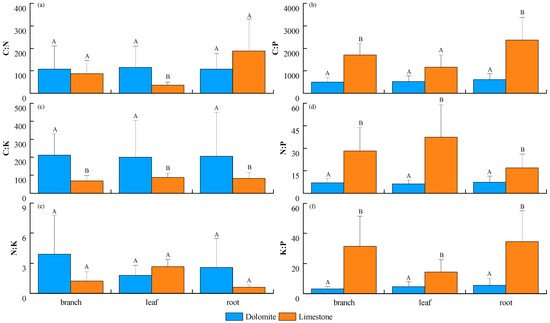
Figure 3.
Differences in the stoichiometric ratios of nutrient elements in various organs across regions. (a) the ratio of C:N, (b) the ratio of C:P, (c) the ratio of C:K, (d) the ratio of N:P, (e) the ratio of N:K, and (f) the ratio of K:P. Uppercase letters indicate significant disparities in the same organ across regions (p < 0.05).
3.3. Correlations Among Nutrient Elements and Stoichiometric Ratios in Various Plant Organs
The analysis of correlations among the nutrient contents of various organs is shown in Figure 4a. Among the 66 pairwise comparisons of elements conducted both within individual organs and across different organs among various tree species, ten pairs in the dolomite region and nine in the limestone region exhibited significant correlations. Specifically, within the organs of tree species in the dolomite region, there were extremely strong positive links between branch N and branch P, as well as between leaf N and leaf K. Furthermore, leaf P correlated significantly positively with leaf K. Conversely, in the limestone region, within the organs of tree species, the only significant positive correlation observed was between leaf C and leaf K. Across organs, significant negative correlations existed between branch N and both leaf P and leaf K; leaf N and branch P; leaf P and root K; and leaf K and root C; and an extremely notable negative correlation existed between branch P and leaf K. Additionally, branch N showed a significant positive correlation with root P. In contrast, in the limestone region, there were highly significant positive correlations between leaf C and both branch C and root C, as well as between branch C and root C. Significant positive correlations were also noted between leaf P and both branch P and root N, between branch K and both leaf K and root K, and between root P and branch P. Evidently, the degree of nutrient element synergism among organs was markedly higher compared to that within organs. Figure 4b exhibits the results of the analysis exploring the correlations between nutrient contents and stoichiometric characteristics across lithological karst regions. In both karst regions, there was a highly significant positive correlation between N and P, along with one between N/P and N/K. Furthermore, strong and statistically significant negative correlations were observed not only between N/K and K/P, but also between C/K and K/P. C/N showed notable correlations with other stoichiometric ratios, except for K/P (and C/K). In the dolomite region, highly significant positive correlations were observed between C/P and C/K, in addition to those between N/K and C/K. Additionally, both N and P showed significant positive correlations with K. In limestone areas, both N and P showed strong positive correlations with C. It was evident that, in karst regions, plants often exhibited synergistic absorption characteristics for elements including N, P, and K.
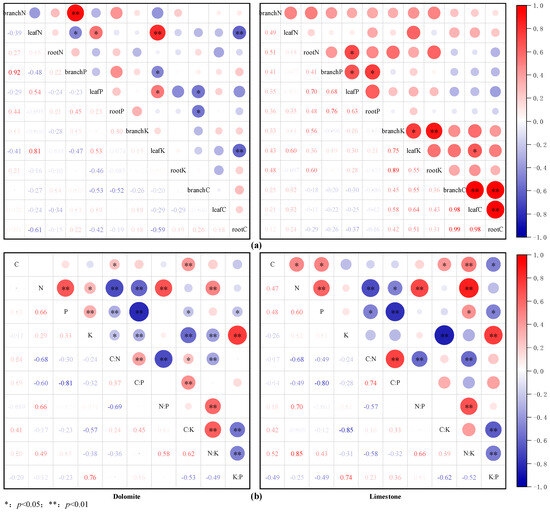
Figure 4.
Analysis of nutrient content correlations among various organs (a); analysis of correlations between nutrient levels and stoichiometric characteristics across lithological karst regions (b).
3.4. The Influence of Different Lithological Karst Regions and Plant Organs on the Nutrient Content and Stoichiometric Ratios of Karst Plants
Both plant organs and lithological karst regions significantly affected the C content, with plant organs having a more pronounced effect. Plant organs also significantly affected the N content. Different lithological karst regions significantly impacted the P and K contents and C/K. Plant organs, lithological karst regions, and their synergistic effects had extremely significant effects on C/P, N/P, and K/P. Additionally, the interaction between plant organs and lithological karst regions had a marked influence on the N content and the C/N. The characteristics of variation in element content in karst plants were the comprehensive results of their responses to changes in organ and lithological regional environments (Table 2).
Table 2.
Two-factor analysis of element concentration and stoichiometric characteristics.
4. Discussion
Plant physiological traits and growth conditions often influence nutrient distribution, causing different changes [31]. As an organ with a high nutrient content in plants, leaves can better reflect plants’ ability to utilize the nutrients in their habitats [32]. The leaf C content of tree species in the dolomite and limestone karst areas of this study was higher than that of leaves in the Puding karst area (431.35 g/kg) [23], of leaves in the peak cluster depression region of the Guangxi karst area (427.5 g/kg) [33], and of leaves in central Guizhou karst area (438.93 g/kg) [27], and similar to that of leaves in the peak cluster depression region of the Qiannan karst area (496.15 g/kg) [34]. It exceeded the global terrestrial leaf C value (464.00 g/kg) [35] and that in other parts of the country (between 438.3 and 476.9 g/kg) [36,37,38]. A high C content in the leaves indicates that a tree species has a strong defense ability in a harsh environment [31]. The karst area is high in temperature and precipitation, which is conducive to improving the carbon storage capacity of plants. This indicates that the tree species in karst areas can produce and store a large amount of organic matter, reflecting the plants’ strategy for optimizing utilization of nutrients in a resource-limited environment, that is, increasing the leaf C content to enhance resistance to environmental stress. The plants’ C assimilation capacity is influenced by the C/N, C/P, and C/K values in their leaves, reflecting the nutrient use efficiency of the plants to some extent [37]. This analysis revealed values that were substantially above the global average for leaves (22.50, 232.00, and 30.73) [1], which was consistent with the results of the karst area of the Guizhou Plateau [23], indicating that the tree species in the karst area had a higher C assimilation rate, and demonstrating their resilience to nutrient limitation, which reflects the high adaptability of karst plants to low-resource environments.
As N and P are essential nutrient elements that can be easily limited in plants, they are important factors limiting the productivity of terrestrial ecosystems [39]. The amounts of leaf N and P in this study were beneath those of plants in the Guangxi karst area, which were 21.2 g/kg and 1.2 g/kg [33], as well as lower than those of karst plants in central Guizhou, which were 16.26 g/kg and 1.73 g/kg [27]. They were also lower than the global leaf levels of 20.60 g/kg and 1.99 g/kg [35] and the domestic terrestrial levels of 20.2 g/kg and 1.46 g/kg [40]. This may be due to the extensive rock exposure, shallow soil layer, inherently minimal amount of nitrogen and phosphorus in the soil, and high precipitation rates in the karst area, where there is a serious loss of N due to leaching [41], reducing the available N content of trees, which leads to a low N content in the leaves of trees. The P content of leaves in China is significantly lower than the average on a global scale [42], and the results of this study verify this conclusion. Organic matter decomposes and mineralizes rapidly in karst areas, which can easily lead to ineffective consumption of soil nutrients and a relative lack of soil P [22]. An imbalance in the soil nitrogen-to-phosphorus ratio can restrict plants’ uptake of P to some degree [31]. The leaf N and P contents varied significantly between the two karst areas, which may be due to the varying requirements for N and P for plant growth in highly heterogeneous karst environments, and the leaves were more sensitive to the highly heterogeneous environment in the karst area. The N/P threshold hypothesis [43] proposes that the N/P content in mature foliage can indicate the nutrients that limit plant growth activities. Generally, an N/P ratio of lower than 14 means that plants tend to be restricted by N, while a ratio greater than 16 indicates P limitation, and within the range of 14 to 16, both N and P constrain plant growth. In the dolomite area, mature leaves exhibited an N/P ratio below 14, indicating that plant growth in these areas was primarily N-limited. In limestone areas, mature leaves exhibited an N/P ratio exceeding 16, suggesting that plant growth in this area was primarily P-limited, and this result was similar to the average N/P ratio of three constructive species in the peak cluster depression karst area of southern Guizhou (22.02 ± 3.79) [34]. In limestone areas, the elevated N/P ratio of mature leaves was primarily attributed to soil phosphorus deficiency. In addition, due to the relatively strong acidity of the soil in limestone areas (pH = 5.7), elements such as iron and aluminum form insoluble compounds with phosphate radicals, decreasing the effectiveness of phosphorus. In contrast, the soil in dolomite areas is usually neutral-to-slightly acidic, which limits the bioavailability of N, making plants more susceptible to N limitation [44].
The average K level in the leaves of this study was under the global average leaf K content of 15.1 g/kg [45] and that of karst plants in central Guizhou, which equaled 12.25 g/kg [46]. This may be related to the soil properties. The K content of limestone soil itself is low, and it is prone to K and Ca ion exchange. Consequently, this hinders the plants’ capacity to absorb and make use of the potassium present in the soil [41]. The higher K concentration in the limestone areas compared with dolomite areas may be due to the fact that soil particles in the limestone area are gathered in the rock fractures and underground void system [44]. A higher amount of roots can facilitate the absorption of potassium from rock crevices at different depths. Venterink et al. [43] have shown that when N/K is greater than 2.1 and K/P is lower than 3.4, plant growth is limited by potassium. Combined with the results of the N/K average of 2.74 and 1.48 and the K/P average of 4.37 and 26.77 in the leaves of this study area, the plant growth in the two karst areas was less affected by potassium. This aligns with the findings of Liu Qingyang et al. [47]. It is related to the efficient utilization of K by karst plants. K has strong variability and is an easy-to-move element in plants. During plant growth, K will be transferred, in accordance with the shifting growth center, to meet the plant’s nutritional needs. In addition, the leaves of karst plants are rich in Ca and Mg, and the enrichment of these elements helps plants to maintain their physiological functions in a low-K environment [48].
Leaves, branches, and roots, as distinct plant organs, exhibited variations in nutrient allocation. In different organs, the order of C content was consistent with the study of Pi Fajian [27]. The ranking of N and P contents consistently showed leaves having the highest content, followed by branches, and then roots, with the exception of N content in the dolomite area. Since roots serve as channels for nutrient absorption, and they transport and store relatively few nutrients, this results in them having lowest nutrient content among the organs. In contrast, the nutrient content in leaves was relatively high. As assimilatory organs, leaves require substantial quantities of N for the synthesis of chlorophyll and other compounds during photosynthesis. Meanwhile, the assimilatory process and photophosphorylation in photosynthesis also demand substantial amounts of P. Therefore, to further enhance the photosynthetic rate of leaves, the amount of N and P allocated to leaves is notably greater than that allocated to roots [49]. Studies have shown that nutrient allocation strategies within plant organs correlate with their functional characteristics, regardless of the scale of the region, ecosystem, or individual plant [34].
Within the same organs of trees, there were three pairs of significant correlations across different elements in dolomite areas, whereas limestone areas showed only one pair. This reflects the covariability of nutrient element supply to these organs by the environment [27]; it is a comprehensive reflection of the utilization of different nutrient elements by plants [34]. Between different organs of trees, seven element combinations in dolomite areas and eight in limestone areas demonstrated significant correlations. Notably, the contents of the same elements (C, P, and K) in the leaf and branch tissues in the limestone area showed a significant correlation. As a transport organ between leaves and stems, branches play an important intermediary role, Therefore, a close link was found between the nutrient constituents and their stoichiometric ratios in leaves as opposed to branches. This indicated that there was a synergistic proportional relationship in the uptake and utilization of nutrients among different organs of trees. The dolomite region displayed no correlation between the same elements across different organs. This suggested that plants exhibited a certain level of homeostasis, ensuring that the deficiency of elements in one organ did not compromise another. It was evident that the correlation among various elements showed notable differences across lithological areas. This phenomenon could be connected to the distinctive environment of the karst area, where various microhabitats, such as stone surfaces, stone crevices, and stone ditches, are staggered in space, with high heterogeneity, and there is the stress of alternating dry and wet conditions [50]. The karstification of limestone is stronger than that of dolomite, and the formed microhabitats and soil properties are different. The thin soil layer and the high rock exposure rate in the karst area provide plants and rocks with a more direct element–nutrition relationship [24]. These niches and soil properties will inevitably affect the physiological activities of some tree species [51]. To survive in this extreme environment, plants exhibit a unique form of nutrient distribution. They can adapt to the barren environment by adjusting the distribution of nutrients among various organs and employing distinct growth strategies.
5. Conclusions
Tree species in dolomite and limestone karst regions adapted to N and P limitations through various physiological mechanisms, including enhanced carbon assimilation, efficient utilization of N, P, and K, synergistic absorption of elements, and coordination among plant organs. The nutrient characteristics and adaptation strategies of plants in these two karst regions were the result of multiple factors acting together, including lithology, plant organs, and interactions among elements. Based on differences in lithology, targeted application of nitrogen or phosphorus fertilizers during vegetation restoration and management in karst areas can stimulate plant growth and further strengthen the carbon sequestration capacity of karst plants and soils.
Author Contributions
Conceptualization, W.Z.; methodology, P.W.; validation, P.W. and H.Z.; investigation, Y.Y., X.J. and Y.H.; resources, H.Z.; data curation, H.Z. and X.J.; writing—original draft preparation, W.Z.; funding acquisition, H.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (No. 32460275), the Guizhou Provincial Key Technology R&D Program (No. [2021]457), and the Science and Technology Planning Project of Guizhou Province (QKHFQ[2023]009).
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
We are deeply appreciative of Liu Yanhui and Cui Yingchun for their invaluable assistance in both the fieldwork and laboratory tasks.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Elser, J.J.; Sterner, R.W.; Gorokhova, E.; Fagan, W.F.; Markow, T.A.; Cotner, J.B.; Harrison, J.F.; Hobbie, S.E.; Odell, G.M.; Weider, L.W. Biological stoichiometry from genes to ecosystems. Ecol. Lett. 2000, 3, 540–550. [Google Scholar] [CrossRef]
- Elser, J.J.; Dobberfuhl, D.R.; MacKay, N.A.; Schampe, J.H. Organism size, life history, and N:P stoichiometry. Bioscience 1996, 48, 674. [Google Scholar] [CrossRef]
- Feng, W.; Yang, J.; Xu, L.; Zhang, G. The spatial variations and driving factors of C, N, P stoichiometric characteristics of plant and soil in the terrestrial ecosystem. Sci. Total Environ. 2024, 951, 175543. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Bai, Y.; Han, X. Application of N:P Stoichiometry to Ecology Studies. Acta Bot. Sin. 2003, 45, 1009–1018. [Google Scholar]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Matzek, V.; Vitousek, P.M. N:P stoichiometry and protein: RNA ratios in vascular plants: An evaluation of the growth-rate hypothesis. Ecol. Lett. 2009, 12, 765–771. [Google Scholar] [CrossRef]
- Gong, Z.; Sheng, M. Ecological stoichiometry of C, N, P and Si of Karst Masson pine forests: Insights for the forest management in southern China. Sci. Total Environ. 2023, 912, 169490. [Google Scholar] [CrossRef]
- Wu, P.; Cui, Y.; Zhao, W.; Shu, D.; Hou, Y.; Ding, F.; Yang, W. Characteristics of soil stoichiometric in natural restoration process of Maolan karst forest vegetation, southwestern China. J. Beijing For. Univ. 2019, 41, 80–92. [Google Scholar]
- Fu, X.; Meng, S.; Kou, L.; Dai, X.; Wang, H. Multiple responses of fine root resource uptake strategies to gravel content in a subtropical plantation. Can. J. For. Res. 2022, 52, 293–300. [Google Scholar] [CrossRef]
- Su, L.; Du, H.; Zeng, F.; Peng, W.; Rizwan, M.; Núñez-Delgado, A.; Zhou, Y.; Song, T.; Wang, H. Soil and fine roots ecological stoichiometry in different vegetation restoration stages in a karst area, southwest China. J. Environ. Manag. 2019, 252, 109694. [Google Scholar] [CrossRef]
- Wang, L.; Arif, M.; Zheng, J.; Li, C. Patterns and drivers of plant carbon, nitrogen, and phosphorus stoichiometry in a novel riparian ecosystem. Front. Plant Sci. 2024, 15, 1354222. [Google Scholar] [CrossRef] [PubMed]
- Zhao, N.; Yu, G.; He, N.; Xia, F.; Wang, Q.; Wang, R.; Xu, Z.; Jia, Y. Invariant allometric scaling of nitrogen and phosphorus in leaves, stems, and fine roots of woody plants along an altitudinal gradient. J. Plant Res. 2016, 29, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Zhou, H.; Cui, Y.; Zhao, W.; Hou, Y.; Tan, C.; Yang, G.; Ding, F. Stoichiometric Characteristics of Leaf, Litter and Soil during Vegetation Succession in Maolan National Nature Reserve, Guizhou, China. Sustainability 2022, 14, 16517. [Google Scholar] [CrossRef]
- Niklas, K.J.; Owens, T.; Reich, P.B.; Cobb, E.D.; Chave, J. Nitrogen/phosphorus leaf stoichiometry and the scaling of plant growth. Ecol. Lett. 2005, 8, 636–642. [Google Scholar] [CrossRef]
- Liao, J.; Dou, Y.; An, S. Plant community productivity is associated with multiple ecological stoichiometry in restoration grasslands. Ecol. Eng. 2023, 187, 106845. [Google Scholar] [CrossRef]
- Shen, Y.; Tian, D.; Jiang, L.; Wang, J.; Chen, X.; Li, Y.; Wang, B.; Li, Z.; Fu, Y.; Hou, J.; et al. Different responses and links of N:P ratio among ecosystem components under nutrient addition in a temperate forest. J. Geophys. Res.-Biogeosci. Sect. 2019, 124, 3158–3167. [Google Scholar] [CrossRef]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Sterner, R.W.; Elser, J.J. Ecological Stoichiometry: The Biology of Elements from Molecules to the Biosphere; Princeton University Press: Princeton, NJ, USA, 2002; pp. 167–196. [Google Scholar]
- Liu, Y.; Li, P.; Wang, T.; Liu, Q.; Wang, W. Root respiration and belowground carbon allocation respond to drought stress in a perennial grass (Bothriochloa ischaemum). Catena 2020, 188, 104449. [Google Scholar] [CrossRef]
- Xiong, J.; Dong, L.; Lu, J. Variation in plant carbon, nitrogen and phosphorus contents across the drylands of China. Funct. Ecol. 2022, 36, 174–186. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Global patterns of plant leaf N and P in relation to temperature and latitude. Proc. Natl. Acad. Sci. USA 2004, 101, 11001–11006. [Google Scholar] [CrossRef]
- Xiao, S.Z.; He, J.H.; Ceng, C.; Xiao, H.; Shi, S.N. Soil nutrient and ecological stoichiometry characteristic in dolomite karst area. J. Henan Univ. Sci. Technol. 2020, 41, 76–81. [Google Scholar]
- Liu, L.; Zhong, Q.; Ni, J. Ecosystem C:N:P stoichiometry and storages of a secondary plateau-surface karst forest in Guizhou Province, southwestern China. Acta Ecol. Sin. 2019, 39, 8606–8614. [Google Scholar]
- Hou, M.F.; Jiang, Z.C. Species diversity of karst original forest in different geochemical environments in Maolan. Ecol. Environ. 2006, 15, 572–576. [Google Scholar]
- Jia, S.; Yu, L. Soil properties and correlation analysis on karst rocky desertification areas of limestone and dolomite: A case study in Xingyi City of Guizhou. Guizhou Sci. 2010, 28, 29–33. [Google Scholar]
- Minden, V.; Kleyer, M. Internal and external regulation of plant organ stoichiometry. Plant Biol. 2014, 18, 897–907. [Google Scholar] [CrossRef]
- Pi, F.; Shu, L.; Yu, L.; Yan, L.; Zhou, C.; Wu, Z.; Yuan, C. Study on ecological stoichiometry characteristics and correlation of plants within different organs of 10 dominant tree species in karst region of central Guizhou. Ecol. Environ. Sci. 2017, 26, 628–634. [Google Scholar]
- Zhao, W.; Cui, Y.; Zhou, T.; Hou, Y.; Zhou, H.; Wu, P. Specific leaf area for 68 species of plant functional groups in Maolan National Nature Reserve. Guizhou For. Sci. Technol. 2022, 50, 1–7. [Google Scholar]
- Zhou, H.; Xu, X.; Jiang, X.; Ding, B.; Wu, P.; Ding, F. Plant functional trait responses to dolomite and limestone karst forests in Southwest China. Forests 2022, 13, 2187. [Google Scholar] [CrossRef]
- Wang, M.; Chen, H.; Zhang, W.; Wang, K. Soil nutrients and stoichiometric ratios as affected by land use and lithology at county scale in a karst area, southwest China. Sci. Total Environ. 2018, 619–620, 1299–1307. [Google Scholar] [CrossRef]
- Zhang, T.; Qiu, Z.; Wu, Z.; Chen, Z.; Hu, H.; Zhou, G.; Zhao, H.; Li, Z.; Cai, Z. Stoichiometric characteristics of Carbon, Nitrogen, Phosphorus and Potassium in organs of coniferous-broadleaved mixed forest in Northern Guangdong. For. Res. 2021, 34, 149–157. [Google Scholar]
- Li, W.; Bai, X.; He, B.; Zou, S.; Huang, J. Responses of soil properties and foliar traits of a dominant tree species Pseudotsuga sinensis to rocky desertification in a karst ecosystem. Glob. Ecol. Conserv. 2024, 55, e3260. [Google Scholar] [CrossRef]
- Zeng, Z.; Wang, K.; Liu, X.; Ceng, F.; Song, T.; Peng, W.; Zhang, H.; Du, H. Stoichiometric characteristics of plants, litter and soils in karst plant communities of North-west Guangxi. Chin. J. Plant Ecol. 2015, 39, 682–693. [Google Scholar]
- Cai, G.; Suo, P.; Zhang, L.; Fu, Y.; Li, A. C, N, P stoichiometric characteristics in different organs of three constructive plants in Karst peak-cluster depressions in southern Guizhou, Southwest China. J. Guizhou Norm. Univ. 2021, 39, 36–44. [Google Scholar]
- Elser, J.J.; Fagan, W.F.; Denno, R.F.; Dobberfuhl, D.R. Nutritional constraints in terrestrial and freshwater food webs. Nature 2000, 408, 578. [Google Scholar] [CrossRef]
- Zheng, S.; Shangguan, Z. Spatial patterns of leaf nutrient traits of the plants in the Loess Plateau of China. Trees 2008, 21, 357–370. [Google Scholar] [CrossRef]
- Huang, J.; Wang, X. Leaf nutrient and structural characteristics of 32 evergreen broad-leaved species. J. Eastchina Norm. Univ. 2003, 1, 92–97. [Google Scholar]
- Liu, W.; Su, J.; Li, S.; Zhang, Z.; Li, Z. Stoichiometry study of C, N and P in plant and soil at different successional stages of monsoon evergreen broad-leaved forest in Pu’er, Yunnan Province. Acta Ecol. Sin. 2010, 30, 6581–6590. [Google Scholar]
- Zheng, C.; Wan, L.; Wang, R.; Wang, G.; Dong, L.; Yang, T.; Yang, Q.; Zhou, J. Effects of rock lithology and soil nutrients on nitrogen and phosphorus mobility in trees in non-karst and karst forests of southwest China. Forest Ecol. Manag. 2023, 548, 121392. [Google Scholar] [CrossRef]
- Han, W.; Fang, J.; Guo, D.; Zhang, Y. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol. 2005, 168, 377–385. [Google Scholar] [CrossRef]
- Luo, T.; Huang, F.; Li, J.; Lu, F.; Wen, S.; Ruan, P.; Li, X. Ecological stoichiometry characteristics of plant dominant species leaf and soil at different restoration stages of vegetation in the karst area of the Lijiang River Basin in Guangxi. J. Plant Resour. Environ. 2024, 33, 80–90. [Google Scholar]
- Ren, S.; Yu, G.; Tao, B.; Wang, S. Leaf Nitrogen and Phosphorus stoichiometry across 654 terrestrial plant species in NSTEC. Environ. Sci. 2007, 28, 2665–2673. [Google Scholar]
- Venterink, H.O.; Wassen, M.J.; Verkroost, A.W.M.; de Ruiter, P.C. Species richness-productivity patterns differ between N-, P-, and K-limited wetlands. Ecology 2003, 84, 2191–2199. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, K.; Qi, X.; Yue, Y.; Tong, X. Spatial and temporal vegetation changes under geological settings of dolomite and limestone based on TM images. Acta Ecol. Sin. 2016, 36, 180–189. [Google Scholar]
- Qin, H.; Li, J.; Gao, S.; Li, C.; Li, R.; Shen, X. Characteristics of leaf element contents for eight nutrients across 660 terrestrial plant species in China. Acta Ecol. Sin. 2010, 30, 1247–1257. [Google Scholar]
- Pi, F.; Yuan, C.; Yu, L.; Yan, L.; Wu, L.; Yang, R. Ecological stoichiometry characteristics of plant leaves from the main dominant species of natural secondary forest in the central of Guizhou. Ecol. Environ. Sci. 2016, 25, 801–807. [Google Scholar]
- Liu, Q.; Wang, Z. Nutrient characteristics of typical plant leaves in karst and non-karst regions of southwest China. J. Hunan Ecol. Sci. 2024, 11, 10–17. [Google Scholar]
- Cui, P.; Shen, Z.; Fu, P.; Bai, K.; Jiang, Y.; Cao, K. Comparison of foliar element contents of plants from natural forests with different substrates in southern China. Acta Ecol. Sin. 2020, 40, 9148–9163. [Google Scholar]
- Luo, J.; Yang, S.; Luo, H.; Li, X.; Li, B.; Guo, W.; Wang, L. Stoichiometric characteristics of C, N and P of five Fagaceae species in subtropical evergreen broad-leaved forests. Acta Ecol. Sin. 2024, 44, 5318–5327. [Google Scholar]
- Guo, K.; Liu, Z.; Dong, M. Ecological adaptation of plants and control of rocky-desertification on karst region of southwest China. Chin. J. Plant Ecol. 2011, 35, 991–999. [Google Scholar] [CrossRef]
- Zou, J.; Yu, L.; Li, Y.; Huang, Z. Changes in δ13C value of plant-litter-soil system in soils developed from different lithologies in Karst area. J. Soil Water Conserv. 2020, 34, 186–194. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).