
Soil Carbohydrates and Glomalin-Related Soil Proteins Affect Aggregate Characteristics in Chinese Fir Plantations with Different Stand Types



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Litter and Soil Sampling
2.4. Indicator Measurements
2.5. Data Processing
3. Results
3.1. Soil Aggregate Characteristics
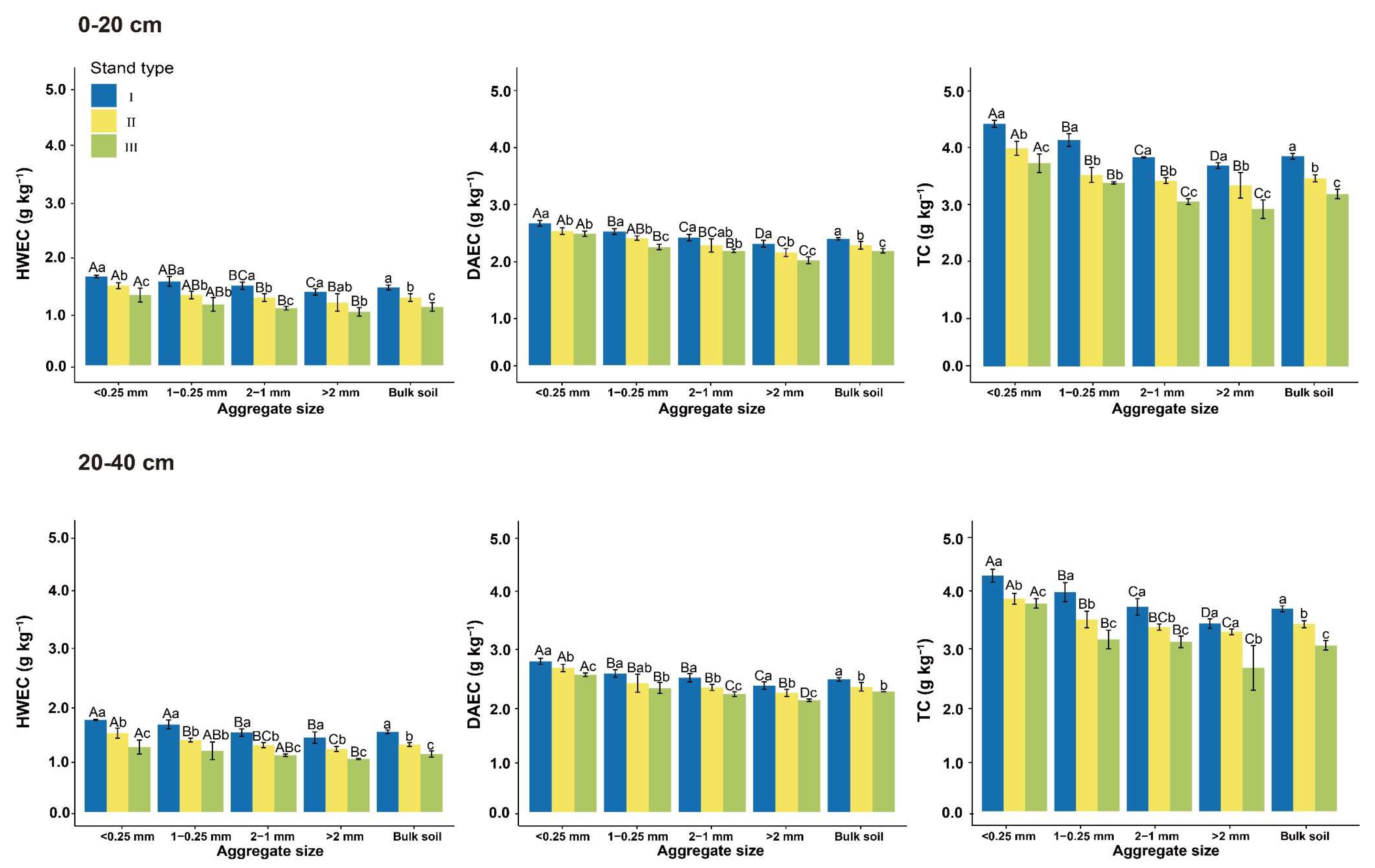
3.2. Soil Aggregate-Associated Carbohydrates
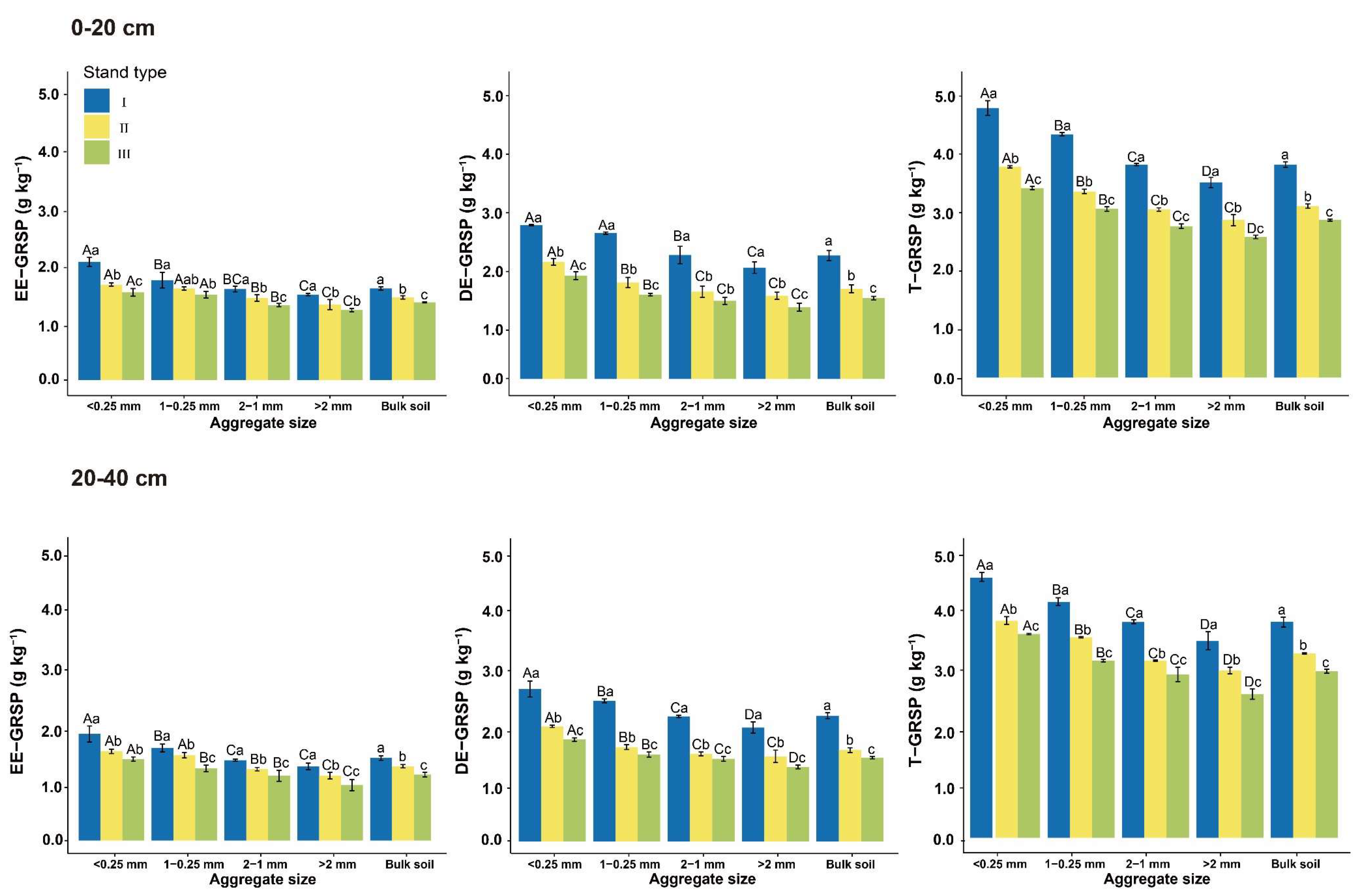
3.3. Soil Aggregate-Associated GRSP
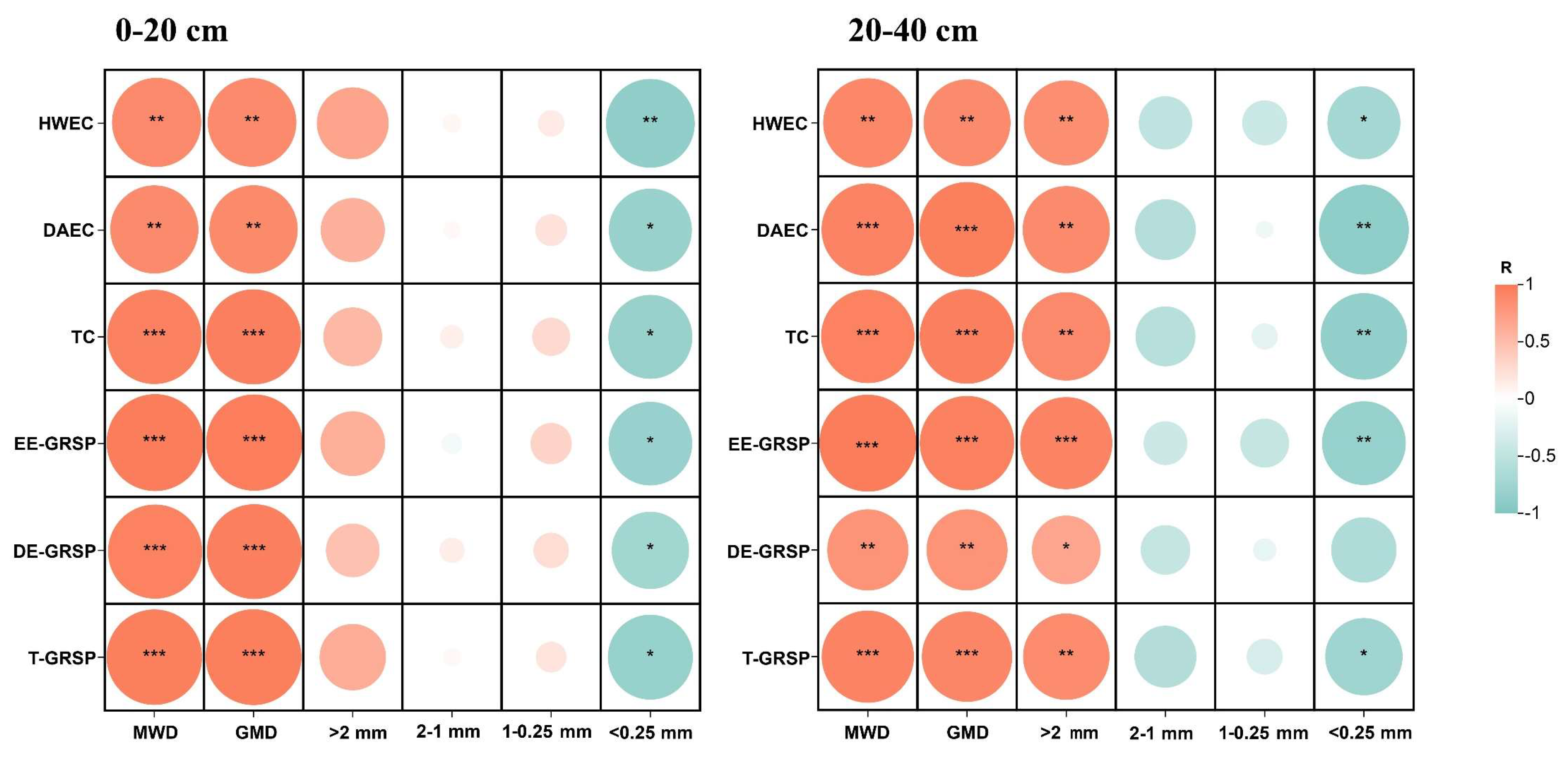
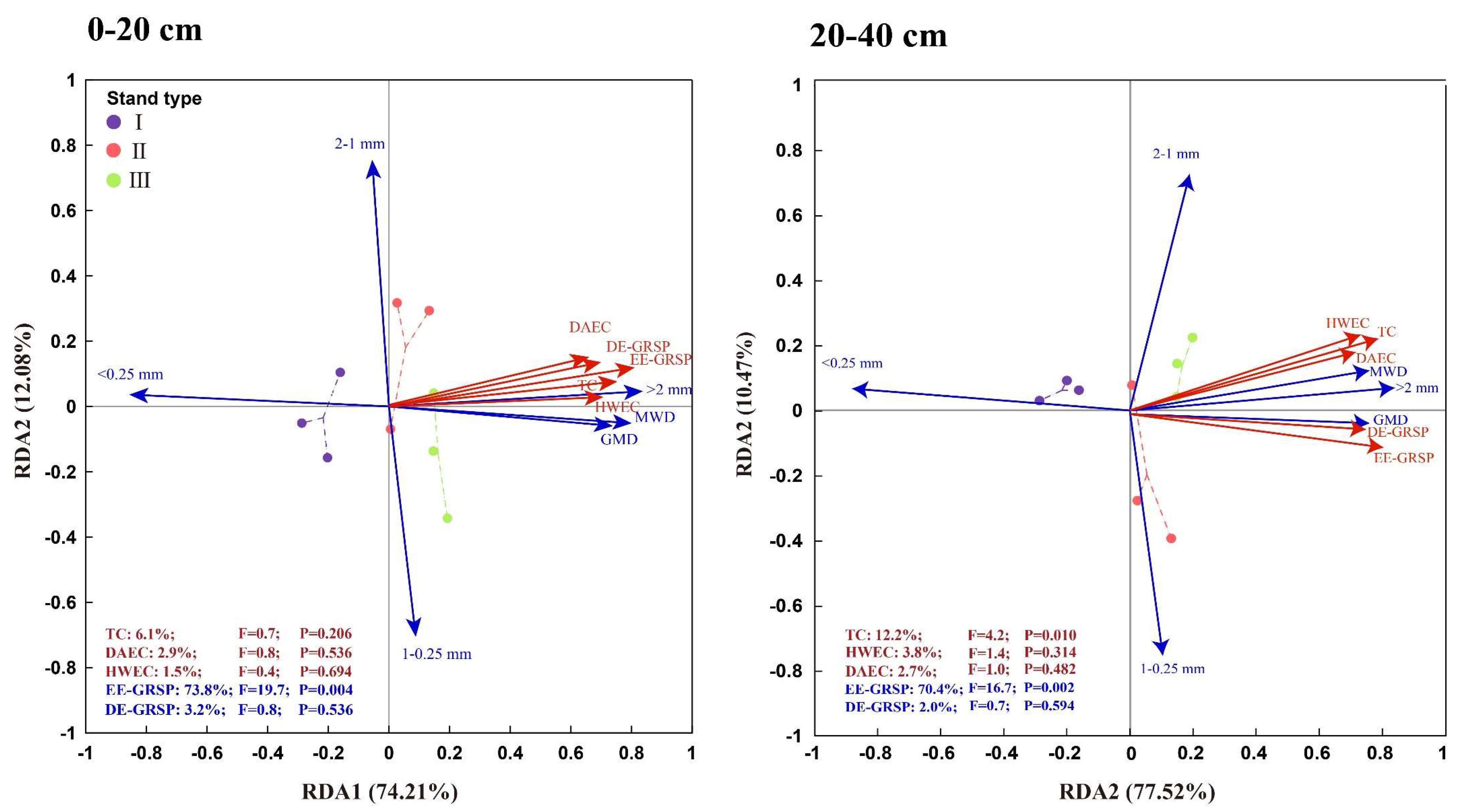
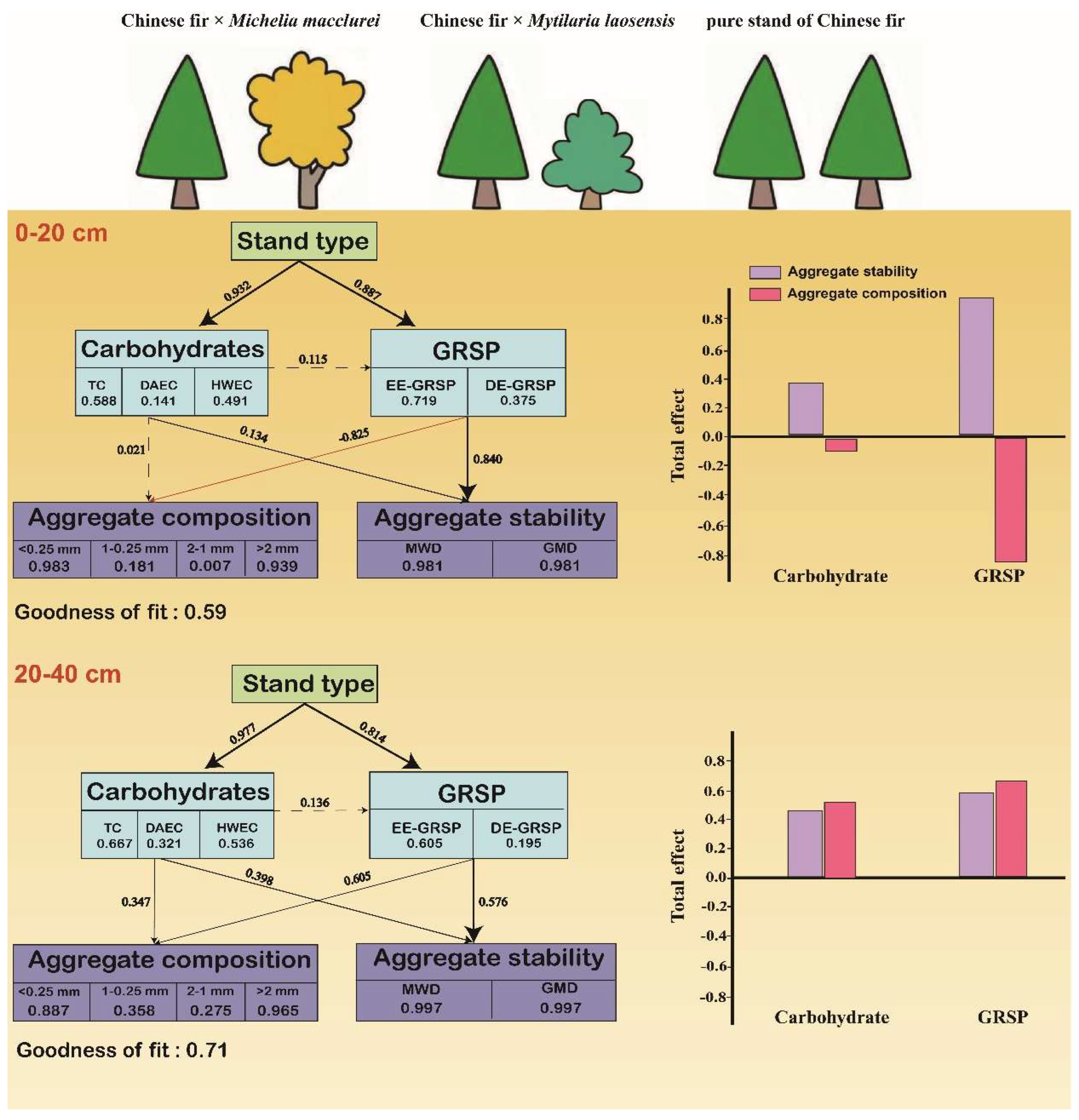
3.4. Regulation of Carbohydrates and GRSP on Soil Aggregates
4. Discussion
4.1. Soil Aggregate Characteristics
4.2. Soil Aggregate-Associated Carbohydrates and GRSP
4.3. Regulation of Carbohydrates and GRSP on Soil Aggregates
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Yang, H.; Viña, A.; Winkler, J.A.; Chung, M.G.; Huang, Q.; Dou, Y.; McShea, W.J.; Songer, M.; Zhang, J.; Liu, J. A global assessment of the impact of individual protected areas on preventing forest loss. Sci. Total Environ. 2021, 777, 145995. [Google Scholar] [CrossRef]
- Bai, Y.; Zhou, Y.; Du, J.; Zhang, X. Tree species identity affects nutrient accumulation and stoichiometric in soil aggregates in mixed plantations of subtropical China. Catena 2024, 236, 107752. [Google Scholar] [CrossRef]
- Sun, C. Exploration on the development status of forest resources in China. For. Investig. Des. 2020, 49, 22–24. [Google Scholar]
- Guo, J.; Sun, J.; Feng, H.; Cao, P.; Yu, Y. Research progress on evolution trends and maintenance measures of soil fertility quality in Chinese fir plantations. J. Zhejiang A F Univ. 2020, 37, 801–809. [Google Scholar]
- Gao, G.; Huang, X.; Xu, H.; Wang, Y.; Shen, W.; Zhang, W.; Yan, J.; Su, X.; Liao, S.; You, Y. Conversion of pure Chinese fir plantation to multi-layered mixed plantation enhances the soil aggregate stability by regulating microbial communities in subtropical China. For. Ecosyst. 2022, 9, 100078. [Google Scholar] [CrossRef]
- Mao, L.; Tang, L.; Ye, S.; Wang, S. Soil organic C and total N as well as microbial biomass C and N affect aggregate stability in a chronosequence of Chinese fir plantations. Eur. J. Soil Biol. 2021, 106, 103347. [Google Scholar] [CrossRef]
- Tang, L.; Wang, S. Dynamics of soil aggregate-related C-N-P stoichiometric characteristics with stand age and soil depth in Chinese fir plantations. Land Degrad. Dev. 2022, 33, 1290–1306. [Google Scholar] [CrossRef]
- Tang, L.; Mao, L.; Wang, Z.; Ye, S.; Wang, S. Mixed with broadleaf tree species improved soil aggregate stability in Chinese fir plantations: Based on the Le Bissonnais method. J. Soil Sci. Plant Nutr. 2023, 23, 2110–2121. [Google Scholar] [CrossRef]
- Zhang, H.; Li, X.; Wang, S.; Jiang, C.; Cui, Y.; Fan, R.; Lan, Y.; Zhang, Q.; Ye, S. Tree–litter–soil system C:N:P stoichiometry and tree organ homeostasis in mixed and pure Chinese fir stands in south subtropical China. Front. For. Glob. Change 2024, 7, 1293439. [Google Scholar] [CrossRef]
- Tisdall, J.; Oades, J.M. Organic matter and water-stable aggregates in soils. Eur. J. Soil Sci. 1982, 33, 141–163. [Google Scholar] [CrossRef]
- Six, J.; Paustian, K. Aggregate-associated soil organic matter as an ecosystem property and a measurement tool. Soil Biol. Biochem. 2014, 68, 4–9. [Google Scholar] [CrossRef]
- Xu, L.; Zhou, Y.; Miao, C.; Chen, H.; Zhang, J.; Qian, H.; Hou, P.; Ding, Y.; Liu, Z.; Li, W.; et al. Long-term straw return increases fungal residual contribution to soil microaggregate nitrogen pool: An eco-enzymatic stoichiometric study. Soil Till. Res. 2024, 244, 106278. [Google Scholar] [CrossRef]
- He, Y.; Zhang, Q.; Wang, S.; Jiang, C.; Lan, Y.; Zhang, H.; Ye, S. Mixed plantations induce more soil macroaggregate formation and facilitate soil nitrogen accumulation. Forests 2023, 14, 735. [Google Scholar] [CrossRef]
- Xie, H.; Li, J.; Zhu, P.; Peng, C.; Wang, J.; He, H.; Zhang, X. Long-term manure amendments enhance neutral sugar accumulation in bulk soil and particulate organic matter in a Mollisol. Soil Biol. Biochem. 2014, 78, 45–53. [Google Scholar] [CrossRef]
- Low, K.E.; Tingley, J.P.; Klassen, L.; King, M.; Xing, X.; Watt, C.; Hoover, S.E.R.; Gorzelak, M.; Abbott, D.W. Carbohydrate flow through agricultural ecosystems: Implications for synthesis and microbial conversion of carbohydrates. Biotechnol. Adv. 2023, 69, 108245. [Google Scholar] [CrossRef]
- Yousefi, M.; Hajabbasi, M.; Shariatmadari, H. Cropping system effects on carbohydrate content and water-stable aggregates in a calcareous soil of Central Iran. Soil Till. Res. 2008, 101, 57–61. [Google Scholar] [CrossRef]
- Rakhsh, F.; Golchin, A. Carbohydrate concentrations and enzyme activities as influenced by exchangeable cations, mineralogy and clay content. Appl. Clay Sci. 2018, 163, 214–226. [Google Scholar] [CrossRef]
- Carrizo, M.E.; Alesso, C.A.; Cosentino, D.; Imhoff, S. Aggregation agents and structural stability in soils with different texture and organic carbon contents. Sci. Agric. 2015, 72, 75–82. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Allen, M.F.; Field, C.B. Rise in carbon dioxide changes soil structure. Nature 1999, 400, 628. [Google Scholar] [CrossRef]
- Wright, S.F.; Franke-Snyder, M.; Morton, J.B.; Upadhyaya, A. Time-course study and partial characterization of a protein on hyphae of arbuscular mycorrhizal fungi during active colonization of roots. Plant Soil 1996, 181, 193–203. [Google Scholar] [CrossRef]
- Wu, Q.; Cao, M.; Zou, Y.; He, X. Direct and indirect effects of glomalin, mycorrhizal hyphae and roots on aggregate stability in rhizosphere of trifoliate orange. Sci. Rep. 2014, 4, 5823. [Google Scholar] [CrossRef]
- Huang, B.; Yan, G.; Liu, G.; Sun, X.; Wang, X.; Xing, Y.; Wang, Q. Effects of long-term nitrogen addition and precipitation reduction on glomalin-related soil protein and soil aggregate stability in a temperate forest. Catena 2022, 214, 106284. [Google Scholar] [CrossRef]
- Cai, C.; Huang, F.; Yang, Y.; Yu, S.; Wang, S.; Fan, Y.; Wang, Q.; Liu, W. Effects of glomalin-related soil protein driven by root on forest soil aggregate stability and carbon sequestration during urbanization in Nanchang, China. Plants 2023, 12, 1847. [Google Scholar] [CrossRef]
- Cissé, G.; Essi, M.; Kedi, B.; Mollier, A.; Staunton, S. Contrasting effects of long term phosphorus fertilization on glomalin-related soil protein (GRSP). Eur. J. Soil Biol. 2021, 107, 103363. [Google Scholar] [CrossRef]
- Han, S.; Lucas-Borja, M.E.; Chen, W.; Huang, Q. Soil glomalin-related protein affects aggregate N2O fluxes by modulating denitrifier communities in a fertilized soil. Sci. Total Environ. 2023, 880, 163147. [Google Scholar] [CrossRef]
- Wu, Q.; He, X.; Zou, Y.; He, K.; Sun, Y.; Cao, M. Spatial distribution of glomalin-related soil protein and its relationships with root mycorrhization, soil aggregates, carbohydrates, activity of protease and β-glucosidase in the rhizosphere of Citrus unshiu. Soil Biol. Biochem. 2012, 45, 181–183. [Google Scholar] [CrossRef]
- Lu, R. High-Temperature External Heating Potassium Dichromate Oxidation-Titration Method. Methods for Agrochemical Analysis of Soils; China Agricultural Science and Technology Press: Beijing, China, 2000. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis: Part 3 C; Sparks, D.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis: Part 3; Sparks, D.L., Ed.; American Society of Agronomy: Madison, WI, USA, 1996; pp. 1085–1121. [Google Scholar]
- Brink, R.H.; Dubach, P.; Lynch, D.L. Measurement of carbohydrates in soil hydrolyzates with anthrone. Soil Sci. 1960, 89, 157–166. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Till. Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Cotching, W.E. Organic matter in the agricultural soils of Tasmania, Australia—A review. Geoderma 2018, 312, 170–182. [Google Scholar] [CrossRef]
- Fang, X.; Zhong, X.; Cui, Z.; Zhang, Y.; Du, L.; Yang, Y.; Lv, J. Distribution and remediation techniques of heavy metals in soil aggregates perspective: A review. Water Air Soil Pollut. 2023, 234, 631. [Google Scholar] [CrossRef]
- Larson, S.L.; Ballard, J.H.; Runge, K.A.; Zhang, H.; Breland, B.R.; Nick, Z.H.; Weiss, C.A.; Han, F. Adsorption and characterization of exopolysaccharides from Rhizobium tropici on clay minerals. Carbohydr. Polym. Technol. Appl. 2023, 5, 100314. [Google Scholar] [CrossRef]
- Liu, G.; Duan, X.; Yan, G.; Sun, X.; Jiang, S.; Xing, Y.; Wang, Q. Changes in soil aggregates and glomalin-related soil protein stability during the successional process of boreal forests. J. Soil Sci. Plant Nutr. 2024, 24, 1335–1348. [Google Scholar] [CrossRef]
- Udom, B.; Ogunwole, J.; Wokocha, C. Aggregate characteristics and aggregate-associated soil organic carbon and carbohydrates of soils under contrasting tree land use. Sains Tanah 2021, 18, 126–135. [Google Scholar] [CrossRef]
- Cotrufo, M.; Wallenstein, M.; Boot, C.; Denef, K.; Paul, E. The microbial efficiency-matrix stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Glob. Change Biol. 2013, 19, 508–512. [Google Scholar] [CrossRef]
- Tang, H.; Liu, L.; Wang, L.; Ba, C. Effect of land use type on profile distribution of glomalin. Chin. J. Eco-Agric. 2009, 17, 1137–1142. [Google Scholar] [CrossRef]
- Li, F.; Chen, L.; Li, Y.; Li, P.; Wang, Y.; Han, Y. Research progress in the effect of fungi on soil aggregate formation. J. Henan Agric. Univ. 2021, 55, 800–806. [Google Scholar]
- Wang, Q.; Lu, H.; Chen, J.; Hong, H.; Liu, J.; Li, J.; Yan, C. Spatial distribution of glomalin-related soil protein and its relationship with sediment carbon sequestration across a mangrove forest. Sci. Total Environ. 2018, 45, 181–183. [Google Scholar] [CrossRef]
- Wu, B.; Umer, M.; Guo, Y.; He, M.; Han, X.; Shen, K.; Xia, T.; He, Y.; He, X. Positive responses of soil nutrients and enzyme activities to AM fungus under interspecific and intraspecific competitions when associated with litter addition. Rhizosphere. 2023, 27, 100728. [Google Scholar] [CrossRef]
- Guo, Z.; Liu, J.; Wu, J.; Yang, D.; Mei, K.; Li, H.; Lu, H.; Yan, C. Spatial heterogeneity in chemical composition and stability of glomalin-related soil protein in the coastal wetlands. Sci. Total Environ. 2022, 835, 155351. [Google Scholar] [CrossRef]
- Qi, D.; Wu, Z.; Yang, C.; Xie, G.; Li, Z.; Yang, X.; Li, D. Can intercropping with native trees enhance structural stability in young rubber (Hevea brasiliensis) agroforestry system? Eur. J. Agron. 2021, 130, 126353. [Google Scholar] [CrossRef]
- He, P.; Deng, X.; Liu, J.; Li, M.; Cheng, F. Non-additive responses of litter decomposition, litter chemical traits, and soil C:N:P stoichiometry to mixing with Eucalyptus in plantation environments. Plant Soil. 2024, 499, 457–472. [Google Scholar] [CrossRef]
- Zhou, L.; Sun, Y.; Saeed, S.; Zhang, B.; Luo, M. The difference of soil properties between pure and mixed Chinese fir plantations depends on tree species. Glob. Ecol. Conserv. 2020, 22, e01009. [Google Scholar] [CrossRef]
- Ma, X.; Ni, X.; Guo, Z.; Zou, X.; Chen, J.; Shen, W.; Kuzyakov, Y. Nitrogen addition influences fine root growth and mycorrhizal symbiosis formation in trees with contrasting root morphology. Appl. Soil Ecol. 2023, 189, 104987. [Google Scholar] [CrossRef]
- Li, L.; McCormack, M.L.; Chen, F.; Wang, H.; Ma, Z.; Guo, D. Different responses of absorptive roots and arbuscular mycorrhizal fungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir. For. Ecol. Manag. 2019, 433, 64–72. [Google Scholar] [CrossRef]
- Yang, L.; Wang, W.; Zhang, F. Effects of reclamationon content and microorganism community metabolic diversity of soil aggregates. Agric. Res. Arid Areas 2018, 36, 215–222. [Google Scholar]
- Yu, G.; Xie, X. Effects of afforestation on soil aggregate stability and microbial carbon metabolism activity in karst area. Res. Soil Water Conserv. 2020, 27, 21–27. [Google Scholar]
- Sun, Z.; Duan, S.; Jiang, Y.; Wang, Q.; Zhang, G. Dominant aggregate binding agent dynamics of quaternary ancient red soils under different land use patterns. Agronomy 2023, 13, 1572. [Google Scholar] [CrossRef]
- Ortner, M.; Seidel, M.; Semella, S.; Udelhoven, T.; Vohland, M.; Thiele-Bruhn, S. Content of soil organic carbon and labile fractions depend on local combinations of mineral-phase characteristics. Soil 2022, 8, 113–131. [Google Scholar] [CrossRef]
- Carrizo, M.E.; Alesso, C.A.; Soares Franco, H.H.; Bernabé Ferreira, C.J.; Imhoff, S. Tensile strength of mollisols of contrasting texture under influence of plant growth and crop residues addition. Geoderma 2018, 329, 1–10. [Google Scholar] [CrossRef]
- Liu, R.; Gao, W.; Srivastava, A.K.; Zou, Y.; Kuča, K.; Hashem, A.; Abd Allah, E.F.; Wu, Q. Differential effects of exogenous glomalin-related soil proteins on plant growth of trifoliate orange through regulating auxin changes. Front. Plant Sci. 2021, 12, 745402. [Google Scholar] [CrossRef]
- Sun, X.; Xing, Y.; Yan, G.; Liu, G.; Wang, X.; Wang, Q. Dynamics of glomalin-related soil protein and soil aggregates during secondary succession in the temperate forest. Catena 2024, 234, 107602. [Google Scholar] [CrossRef]
- Agnihotri, R.; Sharma, M.; Prakash, A.; Ramesh, A.; Bhattacharjya, S.; Patra, A.; Manna, M.; Kurganova, I.; Kuzyakov, Y. Glycoproteins of arbuscular mycorrhiza for soil carbon sequestration: Review of mechanisms and controls. Sci. Total Environ. 2022, 806, 150571. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, J.; Ma, L.; He, X.; Liu, Z.; Wang, F.; Chu, G.; Tang, X. Accumulation of glomalin-related soil protein benefits soil carbon sequestration: Tropical coastal forest restoration experiences. Land Degrad. Dev. 2022, 33, 1541–1551. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Stand Type | ||
|---|---|---|---|
| I | II | III | |
| Altitude (m) | 730 | 723 | 726 |
| Slop (°) | 28 | 24 | 29 |
| Aspect | South | South | South |
| Stand age (year) | 30 | 30 | 30 |
| Crown density | 0.9 | 0.9 | 0.9 |
| Litter quantity (g m−2) | 324 ± 46 b | 455 ± 24 a | 504 ± 22 a |
| Litter C/N ratio | 35 ± 4 a | 33 ± 1 a | 32 ± 2 a |
| Soil Depth | Item | Stand Type | ||
|---|---|---|---|---|
| I | II | III | ||
| 0–20 cm | Sand (%) | 26 ± 1 a | 20 ± 2 b | 15 ± 1 c |
| Silt (%) | 60 ± 1 b | 62 ± 2 b | 66 ± 2 a | |
| Clay (%) | 14 ± 2 c | 18 ± 3 b | 19 ± 3 a | |
| Bulk density (g m−3) | 1.17 ± 0.03 b | 1.26 ± 0.01 a | 1.28 ± 0.02 a | |
| pH | 4.31 ± 0.02 a | 4.28 ± 0.04 ab | 4.22 ± 0.04 b | |
| Organic carbon (g kg−1) | 23.0 ± 1.1 a | 19.7 ± 1.1 b | 17.3 ± 0.60 c | |
| Total nitrogen (g kg−1) | 1.10 ± 0.04 a | 0.98 ± 0.05 b | 0.84 ± 0.07 c | |
| 20–40 cm | Sand (%) | 21 ± 3 a | 15 ± 1 b | 19 ± 3 ab |
| Silt (%) | 62 ± 2 ab | 64 ± 2 a | 58 ± 2 b | |
| Clay (%) | 17 ± 1 c | 21 ± 1 b | 23 ± 1 a | |
| Bulk density (g m−3) | 1.19 ± 0.03 b | 1.27 ± 0.03 a | 1.30 ± 0.02 a | |
| pH | 4.29 ± 0.03 a | 4.25 ± 0.04 ab | 4.21 ± 0.02 b | |
| Organic carbon (g kg−1) | 17.5 ± 1.5 a | 14.4 ± 1.8 a | 10.9 ± 1.7 b | |
| Total nitrogen (g kg−1) | 0.91 ± 0.13 a | 0.87 ± 0.08 ab | 0.72 ± 0.06 b | |
| Soil Depth | Stand Type | Soil Aggregate Composition | Soil Aggregate Stability | ||||
|---|---|---|---|---|---|---|---|
| >2 mm (%) | 2–1 mm (%) | 1–0.25 mm (%) | <0.25 mm (%) | MWD (mm) | GMD (mm) | ||
| 0–20 cm | I | 37 ± 3 Aa | 23 ± 2 Ba | 25 ± 1 Bab | 15 ± 2 Cb | 1.80 ± 0.09 a | 1.12 ± 0.07 a |
| II | 28 ± 1 Ab | 26 ± 2 ABa | 22 ± 3 ABb | 24 ± 3 Ba | 1.54 ± 0.04 b | 0.86 ± 0.05 b | |
| III | 25 ± 1 ABb | 22 ± 2 Ba | 26 ± 0 Aa | 27 ± 2 Aa | 1.39 ± 0.02 c | 0.74 ± 0.03 c | |
| 20–40 cm | I | 32 ± 1 Aa | 20 ± 1 Ba | 29 ± 3 Aab | 19 ± 1 Bb | 1.60 ± 0.04 a | 0.93 ± 0.01 a |
| II | 23 ± 2 Bb | 24 ± 2 Ba | 26 ± 1 ABb | 27 ± 3 Aa | 1.35 ± 0.07 b | 0.73 ± 0.05 b | |
| III | 18 ± 2 Cc | 22 ± 2 Ba | 30 ± 1 Aa | 30 ± 3 Aa | 1.17 ± 0.08 c | 0.63 ± 0.06 c | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Du, L.; Yao, X.; Guo, Y.; Ye, S.; Wang, S. Soil Carbohydrates and Glomalin-Related Soil Proteins Affect Aggregate Characteristics in Chinese Fir Plantations with Different Stand Types. Forests 2025, 16, 444. https://doi.org/10.3390/f16030444
Wang Z, Du L, Yao X, Guo Y, Ye S, Wang S. Soil Carbohydrates and Glomalin-Related Soil Proteins Affect Aggregate Characteristics in Chinese Fir Plantations with Different Stand Types. Forests. 2025; 16(3):444. https://doi.org/10.3390/f16030444
Chicago/Turabian StyleWang, Zhiyao, Lei Du, Xianyu Yao, Yili Guo, Shaoming Ye, and Shengqiang Wang. 2025. "Soil Carbohydrates and Glomalin-Related Soil Proteins Affect Aggregate Characteristics in Chinese Fir Plantations with Different Stand Types" Forests 16, no. 3: 444. https://doi.org/10.3390/f16030444
APA StyleWang, Z., Du, L., Yao, X., Guo, Y., Ye, S., & Wang, S. (2025). Soil Carbohydrates and Glomalin-Related Soil Proteins Affect Aggregate Characteristics in Chinese Fir Plantations with Different Stand Types. Forests, 16(3), 444. https://doi.org/10.3390/f16030444